

Abstract
Background
Hemangioma is a common benign tumor in the childhood; however our knowledge about the molecular mechanisms of hemangioma development and progression are still limited. Currently, microRNAs (miRNAs) have been shown as gene expression regulators with an important role in disease pathogenesis. Our goals were to identify miRNA-mRNA expression networks associated with infantile hemangioma.
Methods
We performed a meta-analysis of previously published gene expression datasets including 98 hemangioma samples. Deregulated genes were further used to identify microRNAs as potential regulators of gene expression in infantile hemangioma. Data were integrated using bioinformatics methods, and genes were mapped in proteins, which were then used to construct protein-protein interaction networks.
Results
Deregulated genes play roles in cell growth and differentiation, cell signaling, angiogenesis and vasculogenesis. Regulatory networks identified included microRNAs miR-9, miR-939 and let-7 family; these microRNAs showed the most number of interactions with deregulated genes in infantile hemangioma, suggesting that they may have an important role in the molecular mechanisms of disease. Additionally, results were used to identify drug-gene interactions and druggable gene categories using Drug-Gene Interaction Database. We show that microRNAs and microRNA-target genes may be useful biomarkers for the development of novel therapeutic strategies for patients with infantile hemangioma.
Conclusions
microRNA-regulated pathways may play a role in infantile hemangioma development and progression and may be potentially useful for future development of novel therapeutic strategies for patients with infantile hemangioma.
Keywords: Infantile hemangioma, MicroRNAs, Gene expression, Protein-protein interaction networks, Molecular pathogenesis, Treatment
Background
Infantile Hemangioma, a common benign tumor in childhood, occurs in 10 % of children, more frequently in prematures and females [1]. It shows a cycle with three phases: an initial proliferative phase (rapid growth during the first year), a plateau and an involution phase (spontaneous regression over 1–8 years) [2, 3]. In a study by Chang et al. [4], growth characteristics were examined in a large number (n = 526) of infantile hemangiomas and the results showed that infantile hemangioma growth occurred mainly in infancy, at a mean age of 3 months. Infantile hemangiomas may be deep or superficial, classified based on the depth of lesions. Deep infantile hemangiomas usually appear later in life and may be associated with a longer growth phase compared to the superficial form. Superficial infantile hemangiomas may be focal or segmental [4]. Segmental lesions are associated with a longer proliferative phase and could require a longer period of treatment [5].
The body area more frequently affected by hemangioma is the head and neck, mainly the face, with an association with the embryological development of the face [6]. Available treatment for patients with infantile hemangioma includes the use of corticosteroids and/or surgical resection of the tumor. The standard of care treatment strategy is the use of propranolol hydrochloride, a β-blocker entered as a safer form of treatment for proliferating infantile hemangioma [7, 8]. Although the advances in therapeutic strategies for infantile hemangioma, the main clinical problems are still the lack of reliable parameters able to distinguish proliferative from involuting IH lesions and the diverse response rates of patients to treatment.
Therefore, the identification of genetic and epigenetic alterations in proliferating and involuting infantile hemangioma lesions will likely contribute to better understand the underlying molecular mechanisms of development and progression of this disease, which is a leading cause of morbidity in affected children. Indeed, differences in the expression of genomic biomarkers have been reported in infantile hemangioma; e.g., insulin-like growth factor 2 (IGF-2) was found as highly expressed in proliferative lesions compared to involuting lesions [9].
In infantile hemangioma, neural crest markers (NG2 and nestin), pericytes markers (δ-like kinase, smooth muscle actin, calponin and CD90) and stem cell markers (OCT4, NANOG and SOX2) are frequently over-expressed both at mRNA and protein levels. In addition, pericytes (perivascular cells surrounding microvessels and that are related to the development and regulation of angiogenesis) and the derm of the face are derived from neural crest, suggesting that the neural crest may be involved in disease pathogenesis [10]. Importantly, expression of lymphatic endothelial hyaluronan receptor-1 (LYVE-1) has been reported in kaposiform hemangioendothelioma and tufted angioma [11]. LYVE-1 was detected as strongly expressed in proliferative infantile hemangiomas but not in pyogenic granulomas or intramuscular hemangioma lesions, suggesting an important role of these markers in the biology of infantile hemangioma [12]. microRNAs (miRNAs) play an important role in gene expression regulation and have been demonstrated to play a role in the pathogenesis of several human diseases [13]. miRNAs are small, non-coding RNAs containing ~18–24 nucleotides. They can bind to the 3’ and 5’ends of the mRNA, leading, in most cases, to translation inhibition or mRNA degradation [14, 15]. Furthermore, miRNAs are related to important biological processes, such as embryonic development, differentiation, apoptosis, cell proliferation [16–18] and oncogenesis [19–21]. To date, 2588 miRNAs were identified and characterized as to their sequence and function in the human genome (http://www.mirbase.org/cgi-bin/browse.pl?org=hsa) [22–26].
Different mechanisms can lead to deregulated miRNA expression, including genomic alterations, such as DNA gains or amplifications and mutations, epigenomic changes including DNA methylation and defects in miRNA biogenesis, including transcription and processing of miRNAs [27, 28]. miRNAs that are altered by these mechanisms may lead to deregulated gene expression.
The understanding of genetic and epigenetic mechanisms, such as deregulated miRNA and target gene expression, as well as molecular pathways regulated by miRNAs, may contribute for the development of new strategies for diagnosis and treatment of complex human diseases [13, 14, 16]. Currently, it is known that miRNAs and miRNA-target genes may represent useful biomarkers to help improve diagnosis, prognosis and treatment of human diseases, such as cancer [29, 30]. Although some gene expression studies have been previously published [31–37], there are no current data on miRNAs or deregulated protein-protein interaction networks in hemangioma. Such data integration strategy is important to understand the functional significance of deregulated genes, miRNAs and molecular pathways involved in hemangioma development and progression. In addition, miRNAs and their target genes may be clinically applicable as therapeutic targets. Indeed, a systematic integration of data derived from multiple sources may achieve the appropriate statistical power and lead to robust, reproducible and accurate predictions [38].
To the best of our knowledge, there are no studies on global miRNA expression in infantile hemangioma. A recent PubMed search (August 19, 2015) showed only one published study on the involvement of miRNAs in senile hemangioma [39], which reported decreased miR-424 expression and increased levels of CCNE1 and MEK1 proteins, which are targeted by miR-424, in patient samples. This study suggested that abnormal proliferation in senile hemangioma may be regulated, at least in part, by miR-424 [39].
Herein, we performed a comprehensive meta-analysis of gene expression data in infantile hemangioma and identified miRNAs as potential regulators of target genes in these tumors. Gene expression datasets were integrated with miRNAs for the identification of molecular pathways potentially involved in infantile hemangioma development and progression. These data may be clinically valuable to predict which infantile hemangioma lesions may respond and which lesions will be resistant to currently available treatment modalities. Furthermore, these data are useful for the identification of robust biomarkers applicable in the development of novel and better molecularly-targeted treatment strategies in infantile hemangioma.
Methods
Meta-analysis of gene expression data in infantile hemangioma
Meta-analysis study design followed the stages of the PRISMA Statement [40] (Fig. 1). Herein, we performed a meta-analysis of previously published gene expression data in infantile hemangioma, by searching PubMed (http://www.ncbi.nlm.nih.gov/pubmed). Key words used were: “infantile hemangioma AND global gene expression”, “infantile hemangioma AND gene signature”, “infantile hemangioma AND microRNAs”, “microRNA in infantile hemangioma”, “infantile hemangioma AND microarray”, infantile hemangioma AND mRNA expression”. Meta-analysis searches comprised studies published between the years of 2000–2015. Considering that our searches did not retrieve any records on miRNA studies in infantile hemangioma, we included only gene expression studies in this meta-analysis. Deregulated genes reported in selected studies were further used for bioinformatics prediction of miRNAs as potential regulators of gene expression, as described below.
Fig. 1.
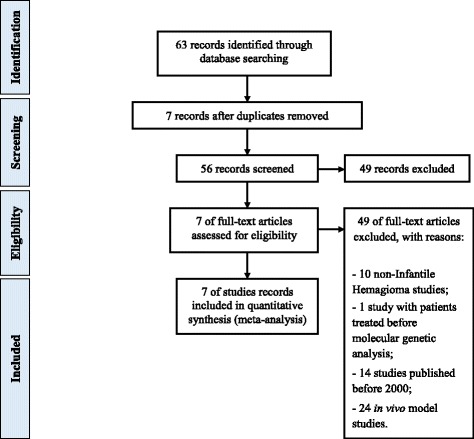
Flowchart of meta-analysis process
Inclusion criteria were: gene expression data in primary patient samples of infantile hemangioma or pure cell populations of infantile hemangioma, any subtype of disease, inclusion of normal tissues for comparison, data subjected to independent validation. Exclusion criteria were: non- infantile hemangioma, patients treated before molecular genetic analysis and in vivo model studies.
Identification of miRNAs as potential modulators of deregulated genes in infantile hemangioma
Deregulated genes identified in the meta-analysis were used for bioinformatics prediction of miRNAs as regulators of gene expression. We used microRNA Data Integration Portal, mirDIP [41], a computational tool that integrates several predicted and validated miRNA databases. mirDIP allows searching for genes that are targeted by miRNAs as well as for miRNAs predicted to regulate genes. Additionally, relevant biological pathways for differentially expressed genes were identified using Biological Networks Gene Ontology (BiNGO) tool, application available in Cytoscape v3.1.1 [42]. BiNGO allows recognizing which of Gene Ontology (GO) categories are statistically more represented in a specific set of genes. Protein-protein interaction (PPI) networks were then generated using Metasearch STRING v9.1 [43, 44] and visualization and annotation data of PPI and miRNA-gene interaction networks were generated using Cytoscape v3.1.1 [45, 46]. Furthermore, we identified drug-gene interactions using Drug-Gene Interaction Database (DGIdb), a database and web-interface for identifying known and potential drug-gene relationships. Genes were defined by Entrez Gene and Ensembl and matched with genes from drug-gene interactions and druggable gene categories. Drugs were defined by searching PubChem and then matched with drugs from drug-gene interaction data. Drug-gene interactions were obtained from multiple sources, including DrugBank, Therapeutic Target Database (TTD) and Pharmacogenomics Knowledge Base (PharmGKB) [47].
Results
Protein-protein interaction networks Identified in infantile hemangioma
According to the meta-analysis study design and the inclusion and exclusion criteria (Fig. 1), we selected 7 studies reporting gene expression data in infantile hemangioma [31–37] (Table 1). Altogether, these studies reported a total of 54 differentially expressed genes (36 over- and 18 under-expressed) in 98 patient samples (Table 2).
Table 1.
Description of publicly available studies used in the meta-analysis
| Reference ID | Sample size | Gene expression analysis and validation analysis platforms |
|---|---|---|
| [31] | 6 hemangiomas and 7 normal term placental tissues | U95Av2 GeneChip oligonucleotide microarrays (Affymetrix) |
| [32] | 7 hemangiomas (3 proliferating, 4 involuting) and 3 normal term placental tissues | Human Genome U133 Plus 2.0 (Affymetrix) |
| [33] | 4 pairs of early proliferative stage and spontaneously early involution stage of the same hemangiomas, 11 hemangiomas (6 proliferative and 5 involuting), 5 controls (normal skin) Serum from 69 patients with hemangioma (46 proliferative and 23 involuting), 20 patients with venous malformations and 31 negative controls (children with cheilopalatognathus) | Illumina Human-6 bead chip, QRT-PCR |
| [34] GSE43742 | HEMECs, HDMVECs, 16 infantile hemangioma, 4 normal controls (neonatal foreskin) | Illumina HumanHT-12 V4.0 expression beadchip Immunohistochemistry |
| [35] | hemSCs, bm-MPCs, HDMECs, cbEPCs and abEPCs | RQ-PCR, Functional assays, Immunofluorescence |
| [36] | HemSCs, HemECs, HDMECs and MSCs | RQ-PCR, Immunofluorescence |
| [37] | 48 hemangiomas, 9 vascular malformations and vascular tumor specimens, 11 neonatal foreskin controls and HemECs from proliferating hemangioma | GeneFilter GF211 (Invitrogen) |
QRT-PCR quantitative reverse-transcription polymerase chain reaction, HEMECs infantile hemangioma endothelial cells, HDMVECs dermal microvascular endothelial cells, HemECs hemangioma-derived endothelial cells, hemSCs proliferating hemangioma-derived CD133+ cells, HDMECs human dermal microvascular endothelial cells, cbEPCs cord blood endothelial progenitor cells, abEPCs adult blood endothelial progenitor cells, bm-MPCs bone marrow-mesenchymal progenitor cells, HemSCs hemangioma-derived stem cells, MSCs mesenchymal stem cells
Table 2.
List of 54 deregulated genes identified in infantile hemangioma, as reported by the seven studies included in the meta-analysis
| Gene symbol | Gene name | Gene function | Gene ID |
|---|---|---|---|
| Over-expressed | |||
| SMARCE1 | SWI/SNF related, matrix associated, actin dependent regulator of chromatin, subfamily e, member 1 | chromatin remodelling | 6605 |
| RGS5 | Regulator of G-protein signaling 5 | cell signaling | 8490 |
| CTAG2 | Cancer/testis antigen 2 | autoimmunogenic tumor antigen | 30848 |
| LTBP2 | Latent transforming growth factor beta binding protein 2 | cell growth and differentiation | 4053 |
| ANG | Angiogenin, ribonuclease, RNase A family, 5 | cell growth and differentiation | 283 |
| IGF2 | Insulin-like growth factor 2 | cell growth and differentiation | 3481 |
| TBX2 | T-box 2 | transcription factor | 6909 |
| NOTCH3 | Notch 3 | cell fate and signalling | 4854 |
| HSD17B2 | Hydroxysteroid (17-beta) dehydrogenase 2 | uncharacterized | 3294 |
| TFPI2 | Tissue factor pathway inhibitor 2 | tumor supressor | 7980 |
| GNG11 | Guanine nucleotide binding protein (G protein), gamma 11 | cell signaling | 2791 |
| NID1 | Nidogen 1 | cell interactions | 4811 |
| COL4A1 | Collagen, type IV, alpha 1 | basement membrane/metabolism | 1282 |
| KDR | Kinase insert domain receptor (a type III receptor tyrosine kinase) | cell growth and differentiation | 3791 |
| FCGR2B | Fc fragment of IgG, low affinity IIb, receptor (CD32) | immunocomplex phagocytosis/antibody production regulation | 2213 |
| PLAGL1 | Pleiomorphic adenoma gene-like 1 | tumor supressor | 5325 |
| DLK1 | Delta-like 1 homolog (Drosophila) | cell growth and differentiation | 8788 |
| JAM3 | Junctional adhesion molecule 3 | cell adhesion | 83700 |
| NID2 | Nidogen 2 (osteonidogen) | cell adhesion | 22795 |
| MEOX2 | Mesenchyme homeobox 2 | myogenesis regulation | 4223 |
| GABRE | Gamma-aminobutyric acid (GABA) A receptor, epsilon | synaptic transmission | 2564 |
| CEACAM1 | Carcinoembryonic antigen-related cell adhesion molecule 1 (biliary glycoprotein) | cell adhesion | 634 |
| BET1 | Bet1 golgi vesicular membrane trafficking protein | vesicular transport | 10282 |
| MXRA5 | Matrix-remodelling associated 5 | matrix remodelling | 25878 |
| IGFBP7 | Insulin-like growth factor binding protein 7 | cell growth and differentiation | 3490 |
| NETO2 | Neuropilin (NRP) and tolloid (TLL)-like 2 | cell signaling | 81831 |
| BAI3 | Brain-specific angiogenesis inhibitor 3 | angiogenesis | 577 |
| PLXDC1 | Plexin domain containing 1 | uncharacterized | 57125 |
| JAG1 | Jagged 1 | hematopoiesis | 182 |
| EDNRA | Endothelin receptor type A | cell signaling | 1909 |
| ICAM2 | Intercellular adhesion molecule 2 | cell adhesion | 3384 |
| NOTCH4 | Notch 4 | cell fate | 4855 |
| STAB1 | Stabilin 1 | cell growth and differentiation | 23166 |
| EPHB3 | EPH receptor B3 | cell signaling | 2049 |
| LPHN1 | Latrophilin 1 | cell adhesion/signal transduction | 22859 |
| NPR1 | Natriuretic peptide receptor 1 | cell signaling | 4881 |
| Under-expressed | |||
| GPR37 | G protein-coupled receptor 37 (endothelin receptor type B-like) | cell signaling | 2861 |
| IGFBP3 | Insulin-like growth factor binding protein 3 | cell growth and differentiation | 3486 |
| FLT1 | Fms-related tyrosine kinase 1 | cell growth and differentiation | 2321 |
| PDGFRA | Platelet-derived growth factor receptor, alpha polypeptide | cell growth and differentiation | 5156 |
| TGFBR3 | Transforming growth factor, beta receptor III | cell growth and differentiation | 7049 |
| LPAR1 | Lysophosphatidic acid receptor 1 | cell growth and differentiation | 1902 |
| IGFBP5 | Insulin-like growth factor binding protein 5 | cell growth and differentiation | 3488 |
| EDNRB | Endothelin receptor type B | cell signaling | 1910 |
| PDGFC | Platelet derived growth factor C | cell growth and differentiation | 56034 |
| BMP4 | Bone morphogenetic protein 4 | cell growth and differentiation | 652 |
| ANGPTL1 | Angiopoietin-like 1 | cell growth and differentiation | 9068 |
| VCAM1 | Vascular cell adhesion molecule 1 | cell adhesion | 7412 |
| BMP5 | bone morphogenetic protein 5 | cell growth and differentiation | 653 |
| IGF1R | Insulin-like growth factor 1 receptor | cell growth and differentiation | 3480 |
| ANGPT2 | Angiopoietin 2 | vascular remodeling | 285 |
| ANTXR1 | Anthrax toxin receptor 1 | cell signaling | 84168 |
| CLDN11 | Claudin 11 | cell adhesion | 5010 |
| KISS1 | KiSS-1 metastasis-suppressor | cell adhesion | 3814 |
Enrichment pathways analysis showed information on the biological role of differentially expressed genes in infantile hemangioma. Gene Ontology (GO) categories were divided into 3 hierarchically structured groups, in order to identify proteins encoded by deregulated genes in infantile hemangioma, and associated with biological processes, molecular functions and cellular components. The top 10 statistically significant enriched GO terms are shown in Fig. 2. Integrated, complex interactome analysis for deregulated genes in infantile hemangioma and functional annotations are shown in Fig. 3. A higher number of interactions was identified between genes with roles in vascular and matrix remodeling, hematopoiesis, cell growth and differentiation and transcriptional control. An interaction network between genes and miRNAs predicted to regulate the expression of these specific genes is shown in Fig. 4. The red and green triangles represent up-regulated and down-regulated genes, respectively. Notably, DGIdb data showed that 5 genes were predicted to interact with drugs that have been demonstrated as clinically useful in other tumor types (Table 3).
Fig. 2.
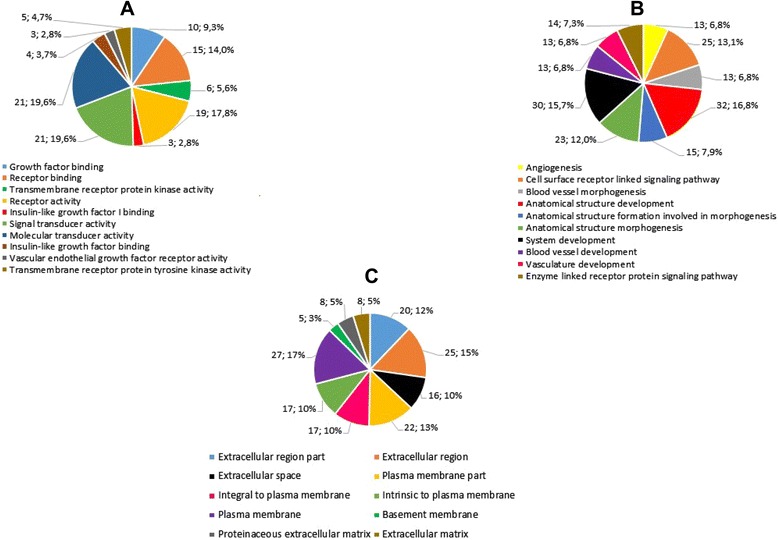
The top 10 enriched Gene Ontology terms of differentially expressed genes. a. molecular function (P value ≤ 1,87E-02); b. biological process (P value ≤ 3,98E-08); c. cellular component (P value ≤ 7,98E-02)
Fig. 3.
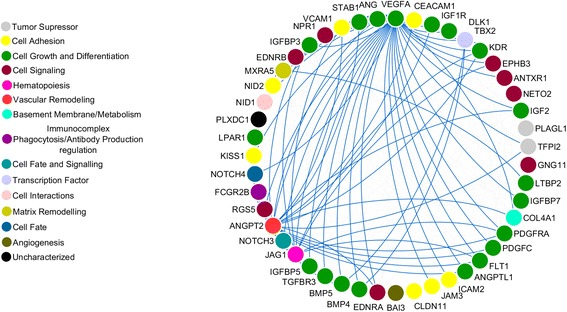
Protein-protein interaction network. The blue lines highlight the interactions of VEGFA, TBX2, JAG1, ANGPT2 and MXRA5. STRING v9.1 was used to generate protein interactions and the result of the network was visualized using Cytoscape v3.1.1
Fig. 4.
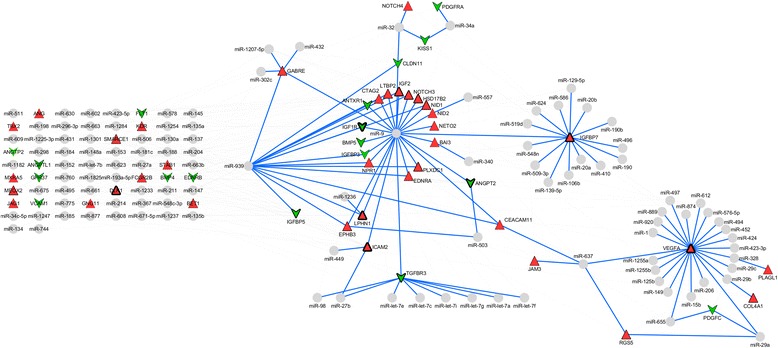
miRNA-gene interaction network. The larger the triangle edge, the higher is the number of interactions identified. The gray circles are miRNAs and blue lines highlights the interactions. The grid only groups miRNAs and genes with few interactions. The miRNA-gene interactions were visualized using Cytoscape v3.1.1
Table 3.
Potential target agents identified based on protein-protein interaction networks of deregulated genes in infantile hemangioma
| Gene symbol | Gene name | Selected target agent | Clinical relevance |
|---|---|---|---|
| EDNRA | endothelin receptor type A | Zibotentan, Atrasentan | Colorectal [62], prostate [63] and renal cell [64] carcinomas |
| IGF1R | insulin-like growth factor 1 receptor | Linsitinib, Ganitumab, Figitumumab, Dalotuzumab, Cixutumumab, Robatumumab | Adrenocortical [65], ovarian [66], non-small cell lung [67], colorectal [68, 69] carcinomas and soft tissue sarcoma [70] |
| PDGFC | platelet derived growth factor C | Sunitinib | Renal cell carcinoma [71] and breast carcinomas [72] |
| PDGFRA | platelet-derived growth factor receptor, alpha polypeptide | Motesanib, Ramucirumab, Midostaurin, Amuvatinib, Nintedanib, Pazopanib, Tandutinib, Crenolanib, Nilotinib, Masitinib, Sorafenib, Sunitinib, Regorafenib, Dovitinib, Telatinib, Vatalanib, Axitinib, Lenvatinib, Imatinib | Colorectal [73, 74], hepatocellular [75–78], kidney [79], non-small [80] and small cell lung [81], pancreatic [82–84], colon [85], gastrointestinal [86], renal cell [71, 87], breast [72, 88], melanoma [89], thyroid [90] carcinomas and soft tissue sarcoma [91] |
| VEGFA | vascular endothelial growth factor A | Ziv-aflibercept, Bevacizumab, Sorafenib tosylate, Lenalidomide, Thalidomide, Aflibercept | Colorectal [92, 93] ovarian [94], non-small cell lung [95], hepatocellular [96] carcinomas and multiple myeloma [97] |
Discussion
Molecular pathways deregulated in hemangioma
Molecular pathogenesis of infantile hemangioma is not well understood. Advances in methods of global genetic and epigenetic analyses represent an extremely valuable approach for the identification of disease development mechanisms and have the potential to identify biomarkers and/or pathways that may be useful for the development of better treatment approaches, including molecularly-targeted therapies.
Our meta-analysis approach allowed us to integrate mRNA expression data in infantile hemangioma and to predict which miRNAs are potential regulators of gene expression. Among the different mechanisms that can lead to gene expression alterations; miRNA alteration is an important mechanism of over- or under-expression of target genes [48]. Herein, we aimed to utilize data on deregulated genes in infantile hemangioma, in order to predict which miRNAs could potentially regulate these genes, and to construct interaction networks between genes and miRNAs.
Gene enrichment analysis showed that deregulated genes previously reported in infantile hemangioma [31–37] are mainly involved in cell signaling and angiogenesis, functioning in vascular and matrix remodeling, hematopoiesis, cell growth and differentiation and transcriptional control.
It is known that the formation of vascular tumors including infantile hemangioma is partly related to increased expression of angiogenic growth factors, such as basic fibroblast growth factor (bFGF) and vascular endothelial growth factor (VEGF), which lead to the development of a disorganized blood vessel mass [49]. Indeed, angiogenesis is mainly regulated by the vascular endothelium [50].
miRNAs control and modulate cell response of vascular endothelium to angiogenic stimuli; for example, miR-126 is a positive regulator of angiogenic signaling and vascular endothelial integrity. Vascular development defects were demonstrated in an in vitro model of miR-126-depleted cells, which did not respond to bFGF and VEGF angiogenic factors [51]. Angiogenic response is also controlled by miRNAs, such as miR-221 and miR-222, which play a role as inhibitors of stem cell factors. Other miRNAs, such as miR-27b and miR-let-7f, play a pro-angiogenic role, since their expression promotes angiogenesis [51]. Notably, miRNA expression in vascular endothelial cells can be modified in response to cellular stimuli or to the microenvironment. For example, a hypoxic environment promotes the production of miR-210, which has pro-angiogenic activity. Therefore, increase in pro-angiogenic miRNA expression in endothelial cells may stimulate the production of angiogenic factors, contributing to the process of tumorigenesis [51].
VEGFA plays an important role in vascular development and in pathological angiogenesis and its protein is highly expressed in vessels of proliferating infantile hemangioma [52]. Interestingly, angiogenin protein (ANG), which is required for cell proliferation and is an important mediator of blood vessel formation, regulates VEGFA expression [53]. Although VEGFA was not identified among the deregulated genes reported in the studies used for meta-analysis, VEGFA is shown in the PPI and miRNA-gene interaction networks, likely due to its important role in angiogenesis and to its indirect interaction with other proteins in the network.
To our knowledge, the only available previously published study on miRNAs in hemangioma identified miR-424 under-expression in senile hemangioma [39]. miR-424 is shown interacting with VEGFA in our miRNA-gene network analysis. miR-424 over-expression has been associated with greater cell motility, decreased cell adhesion and other alterations associated with epithelial-to-mesenchymal transition (EMT) [54]. Notably, this study showed that miR-424 expression levels are increased in primary tumors and decreased in metastasis compared to primary breast tumors and additional functional data suggested that miR-424 may play different roles in the different stages of tumor development and progression [54].
Several genes with roles in vascular and matrix remodeling, hematopoiesis, cell growth and differentiation and transcriptional regulation were shown in the PPI and miRNA-gene interation network. Among these, ANGPT2, MXRA5, JAG1, VEGFA and TBX2 showed a large number of interactions. Interestingly, some of these genes also play roles in cell signaling pathways that have been linked to the pathogenesis of infantile hemangioma [55]; namely, VEGFA in the VEGF/VEGFR pathway, ANGPT2 and ANGPTL1 in the Tie2/Angiopoietin signaling pathway and NOTCH3, NOTCH4 and JAG1 in the Notch pathway. Growth factors and angiopoietins have roles in embryonic development and angiogenesis-dependent diseases and Notch components are involved in modulation of cell fate and differentiation [55].
Herein, miRNA-gene interaction networks generated by integrative meta-analysis showed miRNAs with a large number of interactions (miR-9, miR-939, and let-7 family of miRNAs); these miRNAs are likely acting as main regulators in the network. Notably, miR-9 has been demonstrated as pro-metastatic and suppressor of E-cadherin in breast cancer cells, promoting cell motility and increasing invasive potential of carcinoma cells, besides activating β-catenin signaling, which in turn contributes to high VEGFA expression and consequently to induction of angiogenesis [56]. Increased miR-9 expression levels were also associated with EMT in breast cancer cells and with poor prognosis of patients with breast cancer [57]. In ovarian cancer, miR-939 plays an important role in the progression and regulation of cell growth and cell cycle; it has been demonstrated that ES-2 cells transfected with miR-939 mimic show APC2 decreased expression, suggesting that APC2 may be a target of miR-939 [58].
A recent meta-analysis suggested that the let-7 family of miRNAs are potential biomarkers for tumor grade prediction in breast cancer [59] as well as in other cancers, since let-7 family is highly conserved across species [60].
Among other miRNAs with a significant number of interactions, miR-637 links to main networks through direct interactions with VEGFA and CEACAM; the latter interacting with miR-9 and miR-939. Functional data has shown that miR-637 is one of the effective regulators of HER2 signaling; in HER2-positive trastuzumab non-responsive cell lines, miR-637 was efficient to inhibit breast cancer cell growth [61].
Conclusion
Herein, we identified several interconnected genes and miRNAs as potential regulators of gene expression. Such miRNAs and genes may play important roles in the development and progression of infantile hemangioma. Additionally, these molecules show potential to be targets for drugs that may be clinically useful in the development of new therapies for infants and children affected by this tumor. Data generated herein may be used for validation of expression of miRNAs and genes regulated by miRNAs in infantile hemangioma. Validation analysis in a large representative cohort of primary untreated patient samples is necessary in order to establish robust biomarkers for prediction of treatment response and for the development of better treatment modalities.
Abbreviations
- miRNA or miR
microRNA
- miRDIP
microRNA data integration portal
- BiNGO
biological networks gene ontology
- GO
gene ontology
- PPI
protein-protein interaction
- DGIdb
drug-gene interaction database
- TTD
therapeutic target database
- PharmaGKB
pharmacogenomics knowledge base
- bFGF
basic fibroblast growth factor
- VEGF
vascular endothelial growth factor
- ANG
angiogenin protein
- EMT
epithelial-to-mesenchymal transition
- QRT-PCR
quantitative reverse-transcription polymerase chain reaction
- HEMECs
Infantile hemangioma endothelial cells
- HDMVECs
dermal microvascular endothelial cells
- HemECs
hemangioma-derived endothelial cells
- hemSCs
proliferating hemangioma-derived CD133+ cells
- HDMECs
human dermal microvascular endothelial cells
- cbEPCs
cord blood endothelial progenitor cells
- abEPCs
adult blood endothelial progenitor cells
- bm-MPCs
bone marrow-mesenchymal progenitor cells
- HemSCs
hemangioma-derived stem cells
- MSCs
mesenchymal stem cells
Footnotes
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
PPR designed the study, helped analyze the data and wrote the manuscript. NB helped with study design, performed the meta-analysis and wrote the manuscript. FES performed bioinformatics data analyses and generated figures. LMSP, RM and WBY helped with study design and with writing of the manuscript. All authors read and approved the final version of the manuscript and agreed to be accountable for all aspects of the work.
Authors’ information
Since 2002, WBY and RM coordinate the Vascular Malformations laboratory at a university-based, teaching hospital at the Faculty of Medicine, São Paulo State University, Botucatu, SP, Brazil. The oncopediatric service of our institution (LMSP) work closely with the vascular malformations laboratory. The clinical group (WBY, RM, LMSP) develops multiple quality of life projects for patients with vascular malformations and tumors. Our research group (NB, FES and PPR) is investigating the molecular basis of infantile hemangioma with the main goal of identifying and validating genetic and epigenetic pathways involved in disease development and progression. Such pathways may represent clinically useful biomarkers for prediction of treatment response, or as a basis for the development of novel therapeutic targets for treatment of children affected by hemangioma.
Contributor Information
Natália Bertoni, Email: bertoni.na@gmail.com.
Lied M. S. Pereira, Email: lied.pereira@gmail.com
Fábio E. Severino, Email: fabio.bjj@gmail.com
Regina Moura, Email: rmoura@fmb.unesp.br.
Winston B. Yoshida, Phone: +55 (14) 3880-1443, Email: winston@fmb.unesp.br
Patricia P. Reis, Phone: +55 (14) 3880-1451, Email: preis@fmb.unesp.br
References
- 1.Itinteang T, Tan ST, Brasch HD, Steel R, Best HA, Vishvanath A, et al. Infantile haemangioma expresses embryonic stem cell markers. J Clin Pathol. 2012;65(5):394–398. doi: 10.1136/jclinpath-2011-200462. [DOI] [PubMed] [Google Scholar]
- 2.Holland KE, Drolet BA. Infantile hemangioma. Pediatr Clin North Am. 2010;57(5):1069–1083. doi: 10.1016/j.pcl.2010.07.008. [DOI] [PubMed] [Google Scholar]
- 3.Vikkula M, Boon LM, Mulliken JB. Molecular genetics of vascular malformations. Matrix Biol. 2001;20(5–6):327–335. doi: 10.1016/S0945-053X(01)00150-0. [DOI] [PubMed] [Google Scholar]
- 4.Chang LC, Haggstrom AN, Drolet BA, Baselga E, Chamlin SL, Garzon MC, et al. Growth characteristics of infantile hemangiomas: implications for management. Pediatrics. 2008;122(2):360–367. doi: 10.1542/peds.2007-2767. [DOI] [PubMed] [Google Scholar]
- 5.Drolet BA, Pope E, Juern AM, Sato T, Howell B, Puttgen KB, et al. Gastrointestinal bleeding in infantile hemangioma: a complication of segmental, rather than multifocal, infantile hemangiomas. J Pediatr. 2012;160(6):1021–1026. doi: 10.1016/j.jpeds.2011.12.026. [DOI] [PubMed] [Google Scholar]
- 6.Waner M, North PE, Scherer KA, Frieden IJ, Waner A, Mihm MC. The nonrandom distribution of facial hemangiomas. Arch Dermatol. 2003;139(7):869–875. doi: 10.1001/archderm.139.7.869. [DOI] [PubMed] [Google Scholar]
- 7.Hogeling M, Adams S, Wargon O. A randomized controlled trial of propranolol for infantile hemangiomas. Pediatrics. 2011;128(2):e259–e266. doi: 10.1542/peds.2010-0029. [DOI] [PubMed] [Google Scholar]
- 8.Léauté-Labrèze C, de la Roque DE, Hubiche T, Boralevi F, Thambo JB, Taïeb A. Propranolol for severe hemangiomas of infancy. N Engl J Med. 2008;358(24):2649–2651. doi: 10.1056/NEJMc0708819. [DOI] [PubMed] [Google Scholar]
- 9.Ritter MR, Dorrell MI, Edmonds J, Friedlander SF, Friedlander M. Insulin-like growth factor 2 and potential regulators of hemangioma growth and involution identified by large-scale expression analysis. Proc Natl Acad Sci U S A. 2002;99(11):7455–7460. doi: 10.1073/pnas.102185799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Spock CL, Tom LK, Canadas K, Sue GR, Sawh-Martinez R, Maier CL, et al. Infantile hemangiomas exhibit neural crest and pericyte markers. Ann Plast Surg. 2014. [DOI] [PubMed]
- 11.Le Huu AR, Jokinen CH, Rubin BP, Ruben BP, Mihm MC, Weiss SW, et al. Expression of prox1, lymphatic endothelial nuclear transcription factor, in Kaposiform hemangioendothelioma and tufted angioma. Am J Surg Pathol. 2010;34(11):1563–1573. doi: 10.1097/PAS.0b013e3181f6076f. [DOI] [PubMed] [Google Scholar]
- 12.Dadras SS, North PE, Bertoncini J, Mihm MC, Detmar M. Infantile hemangiomas are arrested in an early developmental vascular differentiation state. Mod Pathol. 2004;17(9):1068–1079. doi: 10.1038/modpathol.3800153. [DOI] [PubMed] [Google Scholar]
- 13.Boyd SD. Everything you wanted to know about small RNA but were afraid to ask. Lab Invest. 2008;88(6):569–578. doi: 10.1038/labinvest.2008.32. [DOI] [PubMed] [Google Scholar]
- 14.He L, Hannon GJ. MicroRNAs: small RNAs with a big role in gene regulation. Nat Rev Genet. 2004;5(7):522–531. doi: 10.1038/nrg1379. [DOI] [PubMed] [Google Scholar]
- 15.Di Leva G, Calin GA, Croce CM. MicroRNAs: fundamental facts and involvement in human diseases. Birth Defects Res C Embryo Today. 2006;78(2):180–189. doi: 10.1002/bdrc.20073. [DOI] [PubMed] [Google Scholar]
- 16.Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116(2):281–297. doi: 10.1016/S0092-8674(04)00045-5. [DOI] [PubMed] [Google Scholar]
- 17.Harfe BD. MicroRNAs in vertebrate development. Curr Opin Genet Dev. 2005;15(4):410–415. doi: 10.1016/j.gde.2005.06.012. [DOI] [PubMed] [Google Scholar]
- 18.Cummins JM, Velculescu VE. Implications of micro-RNA profiling for cancer diagnosis. Oncogene. 2006;25(46):6220–6227. doi: 10.1038/sj.onc.1209914. [DOI] [PubMed] [Google Scholar]
- 19.Rinaldi A, Poretti G, Kwee I, Zucca E, Catapano CV, Tibiletti MG, et al. Concomitant MYC and microRNA cluster miR-17-92 (C13orf25) amplification in human mantle cell lymphoma. Leuk Lymphoma. 2007;48(2):410–412. doi: 10.1080/10428190601059738. [DOI] [PubMed] [Google Scholar]
- 20.Frankel LB, Christoffersen NR, Jacobsen A, Lindow M, Krogh A, Lund AH. Programmed cell death 4 (PDCD4) is an important functional target of the microRNA miR-21 in breast cancer cells. J Biol Chem. 2008;283(2):1026–1033. doi: 10.1074/jbc.M707224200. [DOI] [PubMed] [Google Scholar]
- 21.Reis PP, Tomenson M, Cervigne NK, Machado J, Jurisica I, Pintilie M, et al. Programmed cell death 4 loss increases tumor cell invasion and is regulated by miR-21 in oral squamous cell carcinoma. Mol Cancer. 2010;9:238. doi: 10.1186/1476-4598-9-238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kozomara A, Griffiths-Jones S. miRBase: annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res. 2014;42(Database issue):D68–D73. doi: 10.1093/nar/gkt1181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kozomara A, Griffiths-Jones S. miRBase: integrating microRNA annotation and deep-sequencing data. Nucleic Acids Res. 2011;39(Database issue):D152–D157. doi: 10.1093/nar/gkq1027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Griffiths-Jones S, Saini HK, van Dongen S, Enright AJ. miRBase: tools for microRNA genomics. Nucleic Acids Res. 2008;36(Database issue):D154–D158. doi: 10.1093/nar/gkm952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Griffiths-Jones S, Grocock RJ, van Dongen S, Bateman A, Enright AJ. miRBase: microRNA sequences, targets and gene nomenclature. Nucleic Acids Res. 2006;34(Database issue):D140–D144. doi: 10.1093/nar/gkj112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Griffiths-Jones S. The microRNA Registry. Nucleic Acids Res. 2004;32(Database issue):D109–D111. doi: 10.1093/nar/gkh023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kasinski AL, Slack FJ. Epigenetics and genetics. MicroRNAs en route to the clinic: progress in validating and targeting microRNAs for cancer therapy. Nat Rev Cancer. 2011;11(12):849–864. doi: 10.1038/nrc3166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ha M, Kim VN. Regulation of microRNA biogenesis. Nat Rev Mol Cell Biol. 2014;15(8):509–524. doi: 10.1038/nrm3838. [DOI] [PubMed] [Google Scholar]
- 29.Chen X, Ba Y, Ma L, Cai X, Yin Y, Wang K, et al. Characterization of microRNAs in serum: a novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res. 2008;18(10):997–1006. doi: 10.1038/cr.2008.282. [DOI] [PubMed] [Google Scholar]
- 30.Mitchell PS, Parkin RK, Kroh EM, Fritz BR, Wyman SK, Pogosova-Agadjanyan EL, et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc Natl Acad Sci U S A. 2008;105(30):10513–10518. doi: 10.1073/pnas.0804549105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Barnés CM, Huang S, Kaipainen A, Sanoudou D, Chen EJ, Eichler GS, et al. Evidence by molecular profiling for a placental origin of infantile hemangioma. Proc Natl Acad Sci U S A. 2005;102(52):19097–19102. doi: 10.1073/pnas.0509579102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Calicchio ML, Collins T, Kozakewich HP. Identification of signaling systems in proliferating and involuting phase infantile hemangiomas by genome-wide transcriptional profiling. Am J Pathol. 2009;174(5):1638–1649. doi: 10.2353/ajpath.2009.080517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Jiang C, Lin X, Hu X, Chen H, Jin Y, Ma G, et al. Angiogenin: a potential serum marker of infantile hemangioma revealed by cDNA microarray analysis. Plast Reconstr Surg. 2014;134(2):231e–239e. doi: 10.1097/PRS.0000000000000367. [DOI] [PubMed] [Google Scholar]
- 34.Stiles JM, Rowntree RK, Amaya C, Diaz D, Kokta V, Mitchell DC, et al. Gene expression analysis reveals marked differences in the transcriptome of infantile hemangioma endothelial cells compared to normal dermal microvascular endothelial cells. Vasc Cell. 2013;5(1):6. doi: 10.1186/2045-824X-5-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Todorovich SM, Khan ZA. Elevated T-box 2 in infantile hemangioma stem cells maintains an adipogenic differentiation-competent state. Dermatoendocrinol. 2013;5(3):352–357. doi: 10.4161/derm.26739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Wu JK, Adepoju O, De Silva D, Baribault K, Boscolo E, Bischoff J, et al. A switch in Notch gene expression parallels stem cell to endothelial transition in infantile hemangioma. Angiogenesis. 2010;13(1):15–23. doi: 10.1007/s10456-009-9161-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Yu Y, Wylie-Sears J, Boscolo E, Mulliken JB, Bischoff J. Genomic imprinting of IGF2 is maintained in infantile hemangioma despite its high level of expression. Mol Med. 2004;10(7–12):117–123. doi: 10.2119/2004-00045.Bischoff. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Ramasamy A, Mondry A, Holmes CC, Altman DG. Key issues in conducting a meta-analysis of gene expression microarray datasets. PLoS Med. 2008;5(9) doi: 10.1371/journal.pmed.0050184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Nakashima T, Jinnin M, Etoh T, Fukushima S, Masuguchi S, Maruo K, et al. Down-regulation of mir-424 contributes to the abnormal angiogenesis via MEK1 and cyclin E1 in senile hemangioma: its implications to therapy. PLoS One. 2010;5(12) doi: 10.1371/journal.pone.0014334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Moher D, Liberati A, Tetzlaff J, Altman DG, Group P. Preferred reporting items for systematic reviews and meta-analyses: the PRISMA Statement. Open Med. 2009;3(3):e123–e130. [PMC free article] [PubMed] [Google Scholar]
- 41.Shirdel EA, Xie W, Mak TW, Jurisica I. NAViGaTing the micronome--using multiple microRNA prediction databases to identify signalling pathway-associated microRNAs. PLoS One. 2011;6(2) doi: 10.1371/journal.pone.0017429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Maere S, Heymans K, Kuiper M. BiNGO: a Cytoscape plugin to assess overrepresentation of gene ontology categories in biological networks. Bioinformatics. 2005;21(16):3448–3449. doi: 10.1093/bioinformatics/bti551. [DOI] [PubMed] [Google Scholar]
- 43.Snel B, Lehmann G, Bork P, Huynen MA. STRING: a web-server to retrieve and display the repeatedly occurring neighbourhood of a gene. Nucleic Acids Res. 2000;28(18):3442–3444. doi: 10.1093/nar/28.18.3442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Franceschini A, Szklarczyk D, Frankild S, Kuhn M, Simonovic M, Roth A, et al. STRING v9.1: protein-protein interaction networks, with increased coverage and integration. Nucleic Acids Res. 2013;41(Database issue):D808–D815. doi: 10.1093/nar/gks1094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, Ramage D, et al. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 2003;13(11):2498–2504. doi: 10.1101/gr.1239303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Saito R, Smoot ME, Ono K, Ruscheinski J, Wang PL, Lotia S, et al. A travel guide to Cytoscape plugins. Nat Methods. 2012;9(11):1069–1076. doi: 10.1038/nmeth.2212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Griffith M, Griffith OL, Coffman AC, Weible JV, McMichael JF, Spies NC, et al. DGIdb: mining the druggable genome. Nat Methods. 2013;10(12):1209–1210. doi: 10.1038/nmeth.2689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Iorio MV, Croce CM. MicroRNA dysregulation in cancer: diagnostics, monitoring and therapeutics. A comprehensive review. EMBO Mol Med. 2012;4(3):143–159. doi: 10.1002/emmm.201100209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Boscolo E, Bischoff J. Vasculogenesis in infantile hemangioma. Angiogenesis. 2009;12(2):197–207. doi: 10.1007/s10456-009-9148-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Tille JC, Pepper MS. Hereditary vascular anomalies: new insights into their pathogenesis. Arterioscler Thromb Vasc Biol. 2004;24(9):1578–1590. doi: 10.1161/01.ATV.0000137390.56554.df. [DOI] [PubMed] [Google Scholar]
- 51.Fish JE, Srivastava D. MicroRNAs: opening a new vein in angiogenesis research. Sci Signal. 2009;2(52) doi: 10.1126/scisignal.252pe1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Takahashi K, Mulliken JB, Kozakewich HP, Rogers RA, Folkman J, Ezekowitz RA. Cellular markers that distinguish the phases of hemangioma during infancy and childhood. J Clin Invest. 1994;93(6):2357–2364. doi: 10.1172/JCI117241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Kishimoto K, Liu S, Tsuji T, Olson KA, Hu GF. Endogenous angiogenin in endothelial cells is a general requirement for cell proliferation and angiogenesis. Oncogene. 2005;24(3):445–456. doi: 10.1038/sj.onc.1208223. [DOI] [PubMed] [Google Scholar]
- 54.Drasin DJ, Guarnieri AL, Neelakantan D, Kim J, Cabrera JH, Wang CA, et al. TWIST1-Induced miR-424 Reversibly Drives Mesenchymal Programming while Inhibiting Tumor Initiation. Cancer Res. 2015;75(9):1908–1921. doi: 10.1158/0008-5472.CAN-14-2394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Ji Y, Chen S, Li K, Li L, Xu C, Xiang B. Signaling pathways in the development of infantile hemangioma. J Hematol Oncol. 2014;7(1):13. doi: 10.1186/1756-8722-7-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Ma L, Young J, Prabhala H, Pan E, Mestdagh P, Muth D, et al. miR-9, a MYC/MYCN-activated microRNA, regulates E-cadherin and cancer metastasis. Nat Cell Biol. 2010;12(3):247–256. doi: 10.1038/ncb2024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Gwak JM, Kim HJ, Kim EJ, Chung YR, Yun S, Seo AN, et al. MicroRNA-9 is associated with epithelial-mesenchymal transition, breast cancer stem cell phenotype, and tumor progression in breast cancer. Breast Cancer Res Treat. 2014;147(1):39–49. doi: 10.1007/s10549-014-3069-5. [DOI] [PubMed] [Google Scholar]
- 58.Ying X, Li-Ya Q, Feng Z, Yin W, Ji-Hong L. MiR-939 promotes the proliferation of human ovarian cancer cells by repressing APC2 expression. Biomed Pharmacother. 2015;71:64–69. doi: 10.1016/j.biopha.2015.02.020. [DOI] [PubMed] [Google Scholar]
- 59.Oztemur Y, Bekmez T, Aydos A, Yulug IG, Bozkurt B, Dedeoglu BG. A ranking-based meta-analysis reveals let-7 family as a meta-signature for grade classification in breast cancer. PLoS One. 2015;10(5) doi: 10.1371/journal.pone.0126837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Dai Y, Wang D, Tian X, Zhang L, Sui C, Meng F, et al. Insights into the application of let-7 family as promising biomarker in cancer screening. Tumour Biol. 2015. [DOI] [PubMed]
- 61.Leivonen SK, Sahlberg KK, Mäkelä R, Due EU, Kallioniemi O, Børresen-Dale AL, et al. High-throughput screens identify microRNAs essential for HER2 positive breast cancer cell growth. Mol Oncol. 2014;8(1):93–104. doi: 10.1016/j.molonc.2013.10.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Haque SU, Dashwood MR, Heetun M, Shiwen X, Farooqui N, Ramesh B, et al. Efficacy of the specific endothelin a receptor antagonist zibotentan (ZD4054) in colorectal cancer: a preclinical study. Mol Cancer Ther. 2013;12(8):1556–1567. doi: 10.1158/1535-7163.MCT-12-0975. [DOI] [PubMed] [Google Scholar]
- 63.Younis IR, George DJ, McManus TJ, Hurwitz H, Creel P, Armstrong AJ, et al. Clinical pharmacology of an atrasentan and docetaxel regimen in men with hormone-refractory prostate cancer. Cancer Chemother Pharmacol. 2014;73(5):991–997. doi: 10.1007/s00280-014-2432-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Groenewegen G, Walraven M, Vermaat J, de Gast B, Witteveen E, Giles R, et al. Targeting the endothelin axis with atrasentan, in combination with IFN-alpha, in metastatic renal cell carcinoma. Br J Cancer. 2012;106(2):284–289. doi: 10.1038/bjc.2011.515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Fassnacht M, Berruti A, Baudin E, Demeure MJ, Gilbert J, Haak H, et al. Linsitinib (OSI-906) versus placebo for patients with locally advanced or metastatic adrenocortical carcinoma: a double-blind, randomised, phase 3 study. Lancet Oncol. 2015;16(4):426–435. doi: 10.1016/S1470-2045(15)70081-1. [DOI] [PubMed] [Google Scholar]
- 66.Beltran PJ, Calzone FJ, Mitchell P, Chung YA, Cajulis E, Moody G, et al. Ganitumab (AMG 479) inhibits IGF-II-dependent ovarian cancer growth and potentiates platinum-based chemotherapy. Clin Cancer Res. 2014;20(11):2947–2958. doi: 10.1158/1078-0432.CCR-13-3448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Scagliotti GV, Bondarenko I, Blackhall F, Barlesi F, Hsia TC, Jassem J, et al. Randomized, phase III trial of figitumumab in combination with erlotinib versus erlotinib alone in patients with nonadenocarcinoma nonsmall-cell lung cancer. Ann Oncol. 2015;26(3):497–504. doi: 10.1093/annonc/mdu517. [DOI] [PubMed] [Google Scholar]
- 68.Doi T, Muro K, Yoshino T, Fuse N, Ura T, Takahari D, et al. Phase 1 pharmacokinetic study of MK-0646 (dalotuzumab), an anti-insulin-like growth factor-1 receptor monoclonal antibody, in combination with cetuximab and irinotecan in Japanese patients with advanced colorectal cancer. Cancer Chemother Pharmacol. 2013;72(3):643–652. doi: 10.1007/s00280-013-2240-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Lin EH, Lenz HJ, Saleh MN, Mackenzie MJ, Knost JA, Pathiraja K, et al. A randomized, phase II study of the anti-insulin-like growth factor receptor type 1 (IGF-1R) monoclonal antibody robatumumab (SCH 717454) in patients with advanced colorectal cancer. Cancer Med. 2014;3(4):988–997. doi: 10.1002/cam4.263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Chugh R, Griffith KA, Davis EJ, Thomas DG, Zavala JD, Metko G, et al. Doxorubicin plus the IGF-1R antibody cixutumumab in soft tissue sarcoma: a phase I study using the TITE-CRM model. Ann Oncol. 2015. [DOI] [PMC free article] [PubMed]
- 71.Takayoshi K, Sagara K, Uchino K, Kusaba H, Sakamoto N, Iguchi A, et al. A case of metastatic renal cell carcinoma and bile duct carcinoma treated with a combination of sunitinib and gemcitabine. BMC Cancer. 2015;15(1):426. doi: 10.1186/s12885-015-1443-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Spanheimer PM, Lorenzen AW, De Andrade JP, Kulak MV, Carr JC, Woodfield GW, et al. Receptor Tyrosine Kinase Expression Predicts Response to Sunitinib in Breast Cancer. Ann Surg Oncol. 2015;22(13):4287–4294. doi: 10.1245/s10434-015-4597-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Tebbutt N, Kotasek D, Burris HA, Schwartzberg LS, Hurwitz H, Stephenson J, et al. Motesanib with or without panitumumab plus FOLFIRI or FOLFOX for the treatment of metastatic colorectal cancer. Cancer Chemother Pharmacol. 2015;75(5):993–1004. doi: 10.1007/s00280-015-2694-y. [DOI] [PubMed] [Google Scholar]
- 74.Mross K, Frost A, Scheulen ME, Krauss J, Strumberg D, Schultheiss B, et al. Phase I study of telatinib (BAY 57–9352): analysis of safety, pharmacokinetics, tumor efficacy, and biomarkers in patients with colorectal cancer. Vasc Cell. 2011;3:16. doi: 10.1186/2045-824X-3-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Chuma M, Terashita K, Sakamoto N. New molecularly targeted therapies against advanced hepatocellular carcinoma: From molecular pathogenesis to clinical trials and future directions. Hepatol Res. 2014. [DOI] [PubMed]
- 76.Qi XS, Guo XZ, Han GH, Li HY, Chen J. MET inhibitors for treatment of advanced hepatocellular carcinoma: A review. World J Gastroenterol. 2015;21(18):5445–5453. doi: 10.3748/wjg.v21.i18.5445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.D’Alessandro R, Refolo MG, Lippolis C, Carella N, Messa C, Cavallini A, et al. Modulation of Regorafenib effects on HCC cell lines by epidermal growth factor. Cancer Chemother Pharmacol. 2015;75(6):1237–1245. doi: 10.1007/s00280-015-2751-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Stiuso P, Potenza N, Lombardi A, Ferrandino I, Monaco A, Zappavigna S, et al. MicroRNA-423-5p Promotes Autophagy in Cancer Cells and Is Increased in Serum From Hepatocarcinoma Patients Treated With Sorafenib. Mol Ther Nucleic Acids. 2015;4 doi: 10.1038/mtna.2015.8. [DOI] [PubMed] [Google Scholar]
- 79.Mirza Z, Schulten HJ, Farsi HM, Al-Maghrabi JA, Gari MA, Chaudhary AG, et al. Molecular interaction of a kinase inhibitor midostaurin with anticancer drug targets, S100A8 and EGFR: transcriptional profiling and molecular docking study for kidney cancer therapeutics. PLoS One. 2015;10(3) doi: 10.1371/journal.pone.0119765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Wang P, Song L, Ge H, Jin P, Jiang Y, Hu W, et al. Crenolanib, a PDGFR inhibitor, suppresses lung cancer cell proliferation and inhibits tumor growth in vivo. Onco Targets Ther. 2014;7:1761–1768. doi: 10.2147/OTT.S68773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Mita M, Gordon M, Rosen L, Kapoor N, Choy G, Redkar S, et al. Phase 1B study of amuvatinib in combination with five standard cancer therapies in adults with advanced solid tumors. Cancer Chemother Pharmacol. 2014;74(1):195–204. doi: 10.1007/s00280-014-2481-1. [DOI] [PubMed] [Google Scholar]
- 82.Awasthi N, Hinz S, Brekken RA, Schwarz MA, Schwarz RE. Nintedanib, a triple angiokinase inhibitor, enhances cytotoxic therapy response in pancreatic cancer. Cancer Lett. 2015;358(1):59–66. doi: 10.1016/j.canlet.2014.12.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Deplanque G, Demarchi M, Hebbar M, Flynn P, Melichar B, Atkins J, et al. A randomized, placebo-controlled phase III trial of masitinib plus gemcitabine in the treatment of advanced pancreatic cancer. Ann Oncol. 2015;26(6):1194–1200. doi: 10.1093/annonc/mdv133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Dragovich T, Laheru D, Dayyani F, Bolejack V, Smith L, Seng J, et al. Phase II trial of vatalanib in patients with advanced or metastatic pancreatic adenocarcinoma after first-line gemcitabine therapy (PCRT O4-001) Cancer Chemother Pharmacol. 2014;74(2):379–387. doi: 10.1007/s00280-014-2499-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Ponnurangam S, Standing D, Rangarajan P, Subramaniam D. Tandutinib inhibits the Akt/mTOR signaling pathway to inhibit colon cancer growth. Mol Cancer Ther. 2013;12(5):598–609. doi: 10.1158/1535-7163.MCT-12-0907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Blay JY, Shen L, Kang YK, Rutkowski P, Qin S, Nosov D, et al. Nilotinib versus imatinib as first-line therapy for patients with unresectable or metastatic gastrointestinal stromal tumours (ENESTg1): a randomised phase 3 trial. Lancet Oncol. 2015;16(5):550–560. doi: 10.1016/S1470-2045(15)70105-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Minguet J, Smith KH, Bramlage CP, Bramlage P. Targeted therapies for treatment of renal cell carcinoma: recent advances and future perspectives. Cancer Chemother Pharmacol. 2015. [DOI] [PubMed]
- 88.Grassadonia A, Caporale M, Tinari N, Zilli M, DeTursi M, Gamucci T, et al. Effect of targeted agents on the endocrine response of breast cancer in the neoadjuvant setting: a systematic review. J Cancer. 2015;6(6):575–582. doi: 10.7150/jca.11566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Langdon CG, Held MA, Platt JT, Meeth K, Iyidogan P, Mamillapalli R, et al. The broad-spectrum receptor tyrosine kinase inhibitor dovitinib suppresses growth of BRAF-mutant melanoma cells in combination with other signaling pathway inhibitors. Pigment Cell Melanoma Res. 2015. [DOI] [PMC free article] [PubMed]
- 90.Cabanillas ME, Schlumberger M, Jarzab B, Martins RG, Pacini F, Robinson B, et al. A phase 2 trial of lenvatinib (E7080) in advanced, progressive, radioiodine-refractory, differentiated thyroid cancer: A clinical outcomes and biomarker assessment. Cancer. 2015. [DOI] [PMC free article] [PubMed]
- 91.Haas RL, Gelderblom H, Sleijfer S, van Boven HH, Scholten A, Dewit L, et al. A phase I study on the combination of neoadjuvant radiotherapy plus pazopanib in patients with locally advanced soft tissue sarcoma of the extremities. Acta Oncol. 2015;1–7. [DOI] [PubMed]
- 92.De Souza A, Daly KP, Yoo J, Saif MW. Safety and efficacy of combined yttrium 90 resin radioembolization with aflibercept and folfiri in a patient with metastatic colorectal cancer. Case Rep Oncol Med. 2015;2015:461823. doi: 10.1155/2015/461823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Pietrantonio F, Mazzaferro V, Miceli R, Cotsoglou C, Melotti F, Fanetti G, et al. Pathological response after neoadjuvant bevacizumab- or cetuximab-based chemotherapy in resected colorectal cancer liver metastases. Med Oncol. 2015;32(7):638. doi: 10.1007/s12032-015-0638-3. [DOI] [PubMed] [Google Scholar]
- 94.Selle F, Sevin E, Ray-Coquard I, Mari V, Berton-Rigaud D, Favier L, et al. A phase II study of lenalidomide in platinum-sensitive recurrent ovarian carcinoma. Ann Oncol. 2014;25(11):2191–2196. doi: 10.1093/annonc/mdu392. [DOI] [PubMed] [Google Scholar]
- 95.Villaruz LC, Socinski MA. The role of anti-angiogenesis in non-small-cell lung cancer: an update. Curr Oncol Rep. 2015;17(6):448. doi: 10.1007/s11912-015-0448-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Llovet JM. Focal gains of VEGFA: candidate predictors of sorafenib response in hepatocellular carcinoma. Cancer Cell. 2014;25(5):560–562. doi: 10.1016/j.ccr.2014.04.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Patel UH, Mir MA, Sivik JK, Raheja D, Pandey MK, Talamo G. Central neurotoxicity of immunomodulatory drugs in multiple myeloma. Hematol Rep. 2015;7(1):5704. doi: 10.4081/hr.2015.5704. [DOI] [PMC free article] [PubMed] [Google Scholar]