

Abstract
The mammalian cochlea is a remarkable sensory organ, capable of perceiving sound over a range of 1012 in pressure, and discriminating both infrasonic and ultrasonic frequencies in different species. The sensory hair cells of the mammalian cochlea are exquisitely sensitive, responding to atomic‐level deflections at speeds on the order of tens of microseconds. The number and placement of hair cells are precisely determined during inner ear development, and a large number of developmental processes sculpt the shape, size and morphology of these cells along the length of the cochlear duct to make them optimally responsive to different sound frequencies. In this review, we briefly discuss the evolutionary origins of the mammalian cochlea, and then describe the successive developmental processes that lead to its induction, cell cycle exit, cellular patterning and the establishment of topologically distinct frequency responses along its length.
Keywords: Cochlea, Organ of Corti, Hair cells, Sensory, tonotopy, Notch, BMP, FGF, Shh
The evolutionary origins of the mammalian cochlea
Although the term ‘cochlea’ derives from the Latin description of the coiled snail‐like auditory structure in the mammalian inner ear, the term is habitually also applied to the homologous shorter, uncoiled structures in birds, crocodiles and alligators (archosaurs), snakes and lizards (lepidosaurs), and turtles. Regardless of their length and curvature, these outgrowths from the rest of the inner ear contain a patch of sensory epithelium – the basilar papilla – that responds to sound using mechanosensitive hair cells. In mammals, the basilar papilla is more commonly known as the organ of Corti.
All major vertebrate groups, even those lacking a cochlea, show some form of sensitivity to sound (the exception being lampreys and hagfish where very little information about auditory responses is currently available). In teleost fish, sound perception is carried out by an otolithic organ, the saccular macula, housed in the saccule, which also plays an important role in balance (Popper & Fay, 1999). In some species of fish, sound detection is also performed by a second sensory macula housed in an evagination of the saccule wall termed the lagena. Amphibians also possess saccular and lagenar maculae, but in addition have two extra outgrowths of the saccular wall, housing a very short basilar papilla and another hearing organ, the amphibian papilla, that appears to be a unique amphibian derivation (Smotherman & Narins, 2004). The basilar papilla and lagenar macula are often found in close proximity in amphibians, with the basilar papilla frequently housed in the lagenar recess. Interestingly, such an arrangement of sensory organs is also seen in the closest living relative of tetrapods, the coelacanth Laterimeria (Fritzsch, 1987, 2003), which has led to the idea that the basilar papilla may have arisen in ancestral lobe‐finned fish (Sarcopterygii) and was retained in their tetrapod relatives (Fritzsch, 1992; Fritzsch et al. 2011). In such a scheme, summarized in Fig. 1, the basilar papilla of the amniote cochlea had its origins as a small sensory papilla close to the lagenar macula in lobe‐finned fishes. As the basilar papilla enlarged in the course of evolution, the lagenar macular was displaced to the distal portion of the growing lagenar recess as it transformed into the cochlear duct (Smotherman & Narins, 2004; Fritzsch et al. 2011, 2013; Fritzsch & Straka, 2014). Such an arrangement is seen in modern birds, crocodiles and alligators, which have a banana‐shaped cochlear duct with a basilar papilla running the length of the duct and a small lagenar macula at its apex. Supporting this model, egg‐laying monotreme mammals also have a small lagena at the apex of their cochlear duct (Ladhams & Pickles, 1996), although the lagena has been lost in therian (marsupial and placental) mammals and independently in other groups such as lungfish and caecilians (Fritzsch, 1992). Although modern therian mammals have a characteristically long, coiled cochlear duct, the cochlea of egg‐laying mammals is quite short, and fossil evidence suggests that the modern therian cochlea arose as recently as 100 million years ago, with elongation and coiling occurring to some degree independently of one another. These evolutionary changes are reviewed in detail by Manley (2012).
Figure 1.
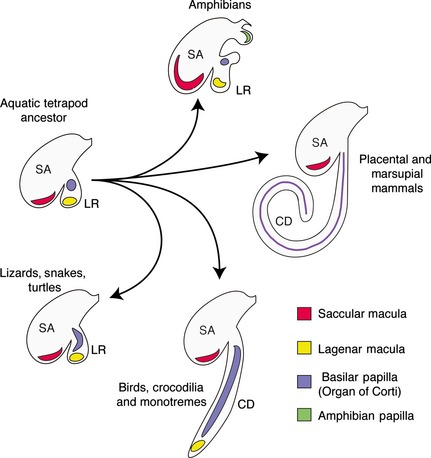
Evolutionary divergence of the inner ear showing the emergence of the cochlea. The aquatic ancestor of modern tetrapods likely had an evagination of the saccule (SA), termed the lagenar recess (LR) that contained the macula lagena (yellow) and a small basilar papilla (purple). This arrangement is seen today in the coelacanth Latimeria (Fritzsch, 1987, 2003) and persists to varying degrees in modern lizards, snakes and turtles, and in many modern amphibians that also have a second unique auditory organ, the amphibian papilla (green). In birds, crocodilians and monotremes, the basilar papilla has elongated to different extents, with the lagenar macula being displaced to the distal tip of the cochlear duct (CD). In therian mammals, the lagena has been lost and the elongated basilar papilla (purple) running the length of the cochlear duct is termed the organ of Corti. In each case, only the pars inferior of the inner ear (saccule, lagenar recess and cochlea duct) are shown in the diagram. This diagram is intended to show the basic trends occurring during the evolution of the cochlea, although in reality considerable variation occurs in the shape and size of the sensory organs in each of the main groups shown in the diagram (Gleich et al. 2004; Manley, 2004, 2012; Smotherman & Narins, 2004; Vater et al. 2004; Fritzsch et al. 2013).
Later in the review, we discuss some of the signals that lead to the differentiation of auditory and vestibular sensory patches in the mammalian inner ear. We currently have very little idea of the molecular and genetic signals that allowed new sensory patches of the ear such as the basilar papilla to arise during evolution. However, loss‐of‐function studies in mice have revealed a number of genes and signals that regulate the outgrowth of the cochlear duct (reviewed in Fritzsch et al. 2011), and it is possible that some of these genes were upregulated or redeployed as the cochlear duct enlarged in amniotes. Moreover, the coordinated elongation of the duct and differentiation of the sensory epithelium into the organ of Corti are tightly coupled, as mutations that affect the length of the mouse cochlear duct typically cause abnormal arrangements of the sensory hair cells of the organ of Corti (Ma et al. 2000; Pauley et al. 2006; Chen et al. 2008).
The embryonic origins of the mammalian cochlea: patterning the early inner ear
The inner ear begins its development as a thickening of ectoderm on either side of the developing hindbrain called the otic placode (Groves, 2005). The otic placode, along with all other craniofacial sensory placodes, derives from a region bordering the anterior neural plate called the preplacodal region (Streit, 2007). The preplacodal region is induced by a combination of signals from both the neural plate and the cranial mesoderm beneath it, requiring Fibroblast Growth Factors (FGFs) FGFs and the suppression of Wnt and bone morphogenetic protein (BMP) signals (Litsiou et al. 2005; Streit, 2007; Grocott et al. 2012; Groves & LaBonne, 2014). After formation of the preplacodal region, FGF signaling at the level of the posterior hindbrain then acts on preplacodal tissue to induce the otic placode (Ohyama et al. 2007). The location and specific identity of the FGFs responsible for otic placode induction vary from species to species, but it has been established that FGFs are both necessary and sufficient to induce some of the earliest markers of the otic placode (Ohyama et al. 2007; Groves & Fekete, 2012). Some of these early otic placode markers include Pax8 (in anamniotes) and Pax2 (in amniotes), which are known to be responsive to FGFs, as well as Foxi1/3 and Sox9 (Pfeffer et al. 1998; Heller & Brandli, 1999; Groves & Bronner‐Fraser, 2000; Nissen et al. 2003; Solomon et al. 2003; Wright & Mansour, 2003; Khatri & Groves, 2013; Khatri et al. 2014).
Once the otic placode has been induced, it undergoes a process not dissimilar to neurulation in which the otic placode begins to close, forming the otic cup. Once this invagination and closure are completed, the resulting structure is known as the otocyst. In teleosts like the zebrafish, the otocyst and neural tube are formed in a slightly different manner, as the structures are instead hollowed out in a process of cavitation (Haddon & Lewis, 1996; Whitfield et al. 2002). It is during the process of otocyst formation that much of the axial patterning of the otocyst occurs, which is vital for the correct formation of many of the sensory structures of the inner ear, including the cochlea.
Anterior–posterior (A–P) axis formation in the mammalian otocyst
The three cardinal axes of the otocyst are established at different times and by different sets of signals (Fig. 2). In chickens, axial patterning occurs after otic placode induction, with the A–P axis becoming fixed before the dorsal–ventral (D–V) axis (Wu et al. 1998; Bok et al. 2005). As early as the otic cup stage, there are already discernable indications of a clear A–P polarity, revealed by the anterior expression of genes defining the early neurosensory region of the otocyst, such as Fgf10, Lunatic Fringe (Lfng), Delta‐like 1, Neurogenin1 (Neurog1) and NeuroD (Morsli et al. 1998, 1999; Cole et al. 2000; Pauley et al. 2003; Alsina et al. 2004). However, despite the asymmetrical expression of these early markers, there is evidence from a series of otic transplantation studies that the A–P axis is not completely fixed until around the time of otic cup closure (Wu, 1998).
Figure 2.
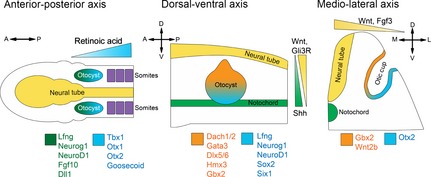
Cardinal axis determination of the amniote inner ear. As the otic placode invaginates to form the otic cup and eventually closes to form the otocyst, the earliest axes established are the M–L axis and the A–P axis, with the D–V axis determined shortly afterwards. A posterior source of RA provides a gradient that allows for the expression of posterior and anterior otic genes. The neural tube provides a source of Wnt to create a dorsalizing gradient, and these dorsalizing signals are augmented by a gradient of inhibitory Gli3R, while the notochord sets up a gradient of Shh, establishing a ventral character. During the otic placode and otic cup stages, Wnt and Fgf3 gradients from the neural tube help establish a medial and a lateral identity.
Efforts to find a source for the establishment of the A–P axis initially hypothesized that the proximity of the otocyst to the rhombomeres of the hindbrain, specifically rhombomeres 5 (r5) and 6 (r6), exposed it to a source of polarizing signals, but it was found that when r5 and r6 were surgically manipulated there was no effect on otocyst A–P patterning in the chick (Bok et al. 2005), although rotation of these rhombomeres did affect the shape and pattern of the cochlear duct (Liang et al. 2010). It was proposed that ectoderm surrounding the otocyst may be playing a role in A–P patterning when it was found that if ectoderm surrounding the otocyst was included during otocyst transplantations to reverse A–P polarity, there was a greater incidence of axis reversal than in cases where ectoderm was absent (Bok et al. 2011). Furthermore, it was shown that retinoic acid (RA) produced by the embryonic ectoderm was a determining factor in establishment of the A–P axis (Bok et al. 2011). Treatment with RA resulted in an expansion of posterior character in the chick and mouse, including an expansion of Tbx1 (Bok et al. 2011), while introducing an anterior source of RA resulted in a mirroring of the posterior otocyst with either the absence of the cochlea or formation of a rudimentary cochlear duct lacking sensory epithelium (Bok et al. 2011). This work supported previous evidence that Tbx1 may be a downstream target of extrinsic A–P patterning signals (Vitelli et al. 2002; Raft et al. 2004). Tbx1 is expressed posteriorly in the mouse otocyst, and the loss of Tbx1 in mice exhibited a posterior expansion of anteriorly expressed genes like Lfng and Neurog1, as well as the loss of posteriorly expressed genes like Otx1, Otx2 and Goosecoid (Vitelli et al. 2002; Raft et al. 2004). In addition, expression of multiple copies of the human TBX1 in mice resulted in a reduction of the anteriorly expressed genes Neurog1 and NeuroD1 (Raft et al. 2004). Together, these data present a model in which a RA signaling gradient used for general determination of A–P polarity in embryonic axis development is co‐opted in the A–P axis patterning of the developing mammalian otocyst (Fig. 2). In other species, notably zebrafish, other signaling pathways such as FGF and Shh have been shown to play a role in A–P patterning of the otocyst in addition to RA signaling (Hammond et al. 2003, 2010; Waldman et al. 2007; Hammond & Whitfield, 2011; Radosevic et al. 2011; Groves & Fekete, 2012; Maier & Whitfield, 2014), but their role in A–P patterning of the mammalian otocyst is less clear.
D–V axis formation in the mammalian otocyst
The inner ear is commonly defined as being divided into two separate functional units: the dorsal pars superior encompassing the dorsal vestibular system (the semicircular canals, their associated sensory cristae and the utricular macula); and the ventral pars inferior, which contains the cochlear duct and saccular macula. Thus, the establishment of the D–V axis of the inner ear is one of the most important first steps in cochlear development. As early as the otic cup stage, the dorsal expression of genes like Dlx5, Dlx6, Hmx2, Hmx3 and Gbx2 (used as markers of dorsal character), and the ventral expression of genes like Lfng, Neurog1, NeuroD1, Sox2 and Six1 (used as markers of ventral character) suggest that D–V differentiation is already apparent in both the chick and mouse (Morsli et al. 1998; Brigande et al. 2000b; Cantos et al. 2000; Fekete & Wu, 2002; Groves & Fekete, 2012; Wu & Kelley, 2012). Despite the fact that markers identifying D–V and A–P polarity appear at approximately the same time, transplantation experiments in the chick manipulating otocyst orientation indicate that the D–V axis is not committed until after the A–P axis (Wu et al. 1998).
The hindbrain and notochord are thought to provide signals that specify D–V polarity in the otocyst (Giraldez, 1998; Bok et al. 2005). Rotation of the hindbrain about its D–V axis resulted in the otocyst expressing ventral genes such as Lfng, NeuroD1 and Six1 in a more dorsal pattern, while expression of dorsal genes like Gbx2 was abolished (Bok et al. 2005). Some of the signals produced by the hindbrain that are thought to be responsible for setting up the D–V axis are the Wnt family of signaling molecules and the morphogen Sonic Hedgehog (Shh; Riccomagno et al. 2002, 2005; Bok et al. 2005, 2007a ,b; Brown & Epstein, 2011; Wu & Kelley, 2012). In this regard, the D–V patterning of the otocyst shares much in common with the patterning of the neural tube and hindbrain (Fig. 2). In this model, the expression of Wnts dorsally by the hindbrain promotes a dorsal character in the otocyst, and the expression of Shh ventrally by the notochord and floorplate of the hindbrain promotes a ventral character. This model is supported by the fact that in the case of Wnt1/Wnt3a double knockout mice there is an absence of Dlx5, Dlx6 and Gbx2, as well as the loss of development of vestibular structures (Riccomagno et al. 2005).
Ventral patterning of the otocyst has been shown to require Shh in both mouse and chick (Riccomagno et al. 2002, 2005; Bok et al. 2005, 2007a,b; Brown & Epstein, 2011; Wu & Kelley, 2012). Shh null mice show a decrease in the expression of ventral marker genes Otx1, Otx2, Lfng, Fgf3, Neurog1 and NeuroD1, as well as the loss of ventral structures such as the cochlea and saccule (Riccomagno et al. 2002; Bok et al. 2005). In addition, when hybridoma cells expressing antibodies that block SHH activity were implanted in the ventral midlines of chicks during the otic cup stage, it resulted in the elimination of ventral structures (Bok et al. 2005). However, in each of these cases, SHH signaling has been eliminated in a manner that caused effects on other tissues in the embryo. To address whether Shh signaling acts directly on the otocyst, the Shh receptor smoothened (Smo) was deleted specifically in the inner ear (Brown & Epstein, 2011). Smo conditional mutant mice show a loss of ventral structures like the cochlea, but developed normal dorsal structures (Brown & Epstein, 2011). A simple gradient model for Shh patterning the ventral otocyst is further questioned by consideration of Shh‐responsive transcription factors, the Gli proteins. Different combinations of Gli mutant alleles suggest the presence of the Gli3 repressor must balance the activity of Gli2/3 activators (Bok et al. 2007a,b). Gli2/Gli3 null mice fail to form the most ventral structures like the cochlear duct, similar to Shh nulls (Bok et al. 2007a,b). Structures farther from the most ventral region of the otocyst are lost in Shh nulls, but restored when combined with the loss of Gli3 repressor activity (Bok et al. 2007a,b). Furthermore, the loss of the Gli3 repressor results in malformed dorsal structures, reinforcing the case for a precise balance between Gli activators and repressors (Bok et al. 2007a,b).
Further evidence arguing against a simple two‐gradient model of Wnt and Shh signaling comes from a comparison of loss of function of the two pathways in the otocyst. Shh null mice show a ventral expansion of the dorsal markers Dlx5 and Gbx2, but the loss of Wnt1/3 signaling does not expand ventral otocyst markers like Gli1, Neurog1, Otx2 or Pax2 (Riccomagno et al. 2002, 2005). In addition, the loss of Wnt signaling results in the loss of only some dorsal genes, while others like Hmx3 and Wnt2b appear unaffected (Riccomagno et al. 2005). This suggests that there are likely to be other pathways or signals participating in D–V patterning. One candidate pathway is the BMP pathway, as it is expressed in the right time and place in the dorsal neural tube (Liem et al. 1995, 2000). A problem in discerning the role of BMPs on D–V axis formation is the fact that the otocyst itself expresses BMPs at various times and places during its development (Morsli et al. 1998). Another gene that seems to play an important role in D–V polarity is the transcription factor Six1. It is expressed ventrally in the otocyst and its loss seems to result in a phenocopy of the Shh null, albeit with the additional expansion of dorsal genes Hmx3, Gata3, Dach1 and Dach2 (Zheng et al. 2003; Ozaki et al. 2004). However, it also appears to be independent of Shh regulation, as Six1 is itself not altered in Shh null mice (Ozaki et al. 2004).
Medio–lateral (M–L) axis formation in the mammalian otocyst
The M–L axis is important for establishing the identity of several structures in the developing inner ear, particularly the endolymphatic duct (a dorsal and medial structure), and the lateral semicircular canal and ampulla. However, little is known about which tissues are providing M–L identity to the developing inner ear. In addition, fate‐mapping in the chick suggests that some structures such as the A–P semicircular canals and their ampullae, as well as the cochlear duct may form from a combination of M–L domains (Brigande et al. 2000a; Fekete & Wu, 2002). Some genes appear to be expressed in a M–L gradient as early as the otic placode stage, including the expression of Wnt reporter genes (Ohyama et al. 2006; Jayasena et al. 2008). However, in surgical manipulations altering the M–L orientation of the otocyst in chicks, the otocysts underwent malformations during development without the respecification of medial or lateral genes (Wu et al. 1998). These experiments also showed that M–L specification may occur approximately at the same time as A–P axis formation (Wu et al. 1998).
The hindbrain has been implicated as a source of M–L patterning signals (Fig. 2). Certain mouse mutants that cause hindbrain defects like Hoxa1, Fgf3 and Kreisler/MafB mutants exhibit r5/r6 defects and defects in FGF signaling, and also lack the presence of an endolymphatic duct in addition to other inner ear defects (Deol, 1964; Mark et al. 1993; McKay et al. 1996; Choo et al. 2006). Together, these data suggest that hindbrain‐derived Fgf3 may play a role in M–L axis patterning (Lin et al. 2005). Further investigation of the role of the hindbrain and its signaling centers in Kreisler mutants showed they lack the endolymphatic duct, but still possess the lateral canal and its ampulla (Choo et al. 2006). Genes expressed dorsal–medially, Gbx2 and Wnt2b, were downregulated in the ears of these mice as well, while Otx2, a gene expressed in the lateral otocyst, expanded medially (Choo et al. 2006). Wnt signaling has also been proposed to play a role in M–L patterning, as fate mapping of Wnt‐responsive cells in the otic cup indicated that these cells made broad contributions to the inner ear, especially the entire medial wall (Riccomagno et al. 2005; Brown et al. 2015). DiI fate‐mapping of the chick otic cup also revealed that the bulk of the lateral wall of the otocyst originated from the ventral–posterior rim of the otic cup (Brigande et al. 2000a).
In summary, for the inner ear and cochlea to develop with the correct patterning and structures, the establishment of the cardinal axes is vital. To establish these axes and subsequently pattern the otocyst, many of the same developmental paradigms used to pattern the embryo proper are redeployed: the same signaling process used to pattern the D–V axis of the neural tube is utilized in the otocyst, and the process used to confer A–P identity to the embryo is used for the otocyst A–P axis formation.
Mapping the origins of the cochlea and the organ of Corti
The signals described in the preceding sections help partition the otocyst into its three cardinal axes. It is well‐established that the cochlear duct grows out from the ventral otocyst (Li et al. 1978), with a portion of cells located posterior–ventrally beginning to form a protrusion around embryonic day 11 in the mouse that marks the beginning of the cochlear duct (Riccomagno et al. 2005; Wu & Kelley, 2012; Brown et al. 2015). Over the next 5 days, the cochlear duct continues to elongate and coil in an anterior–medial direction until it reaches its full one and three‐quarters turns (Fig. 3). In contrast to the vestibular sensory organs (Wu & Oh, 1996; Morsli et al. 1998; Wu & Kelley, 2012; Raft & Groves, 2015), there is currently only a small amount of fate‐mapping or gene expression data that describes the early location of cochlear progenitors in general and progenitors of the organ of Corti in particular (Wu & Kelley, 2012). Fate‐mapping studies using a Neurog1‐CreER transgenic mouse to track the fate of the Neurog1‐expressing progenitors in the anterior and ventral regions of the otocyst between embryonic days 8 and 13 show few descendants of this region in the cochlear duct and almost no cells in the organ of Corti itself (Koundakjian et al. 2007; Raft et al. 2007). Fate‐mapping of Wnt‐responsive progenitors using a CreER mouse line driven by Lef/TCF‐binding sites (TOP‐CreER) shows that some Wnt‐responsive progenitors in the medial wall of the otocyst between E9.5 and 11.5 will contribute to sensory epithelium of the cochlear duct (Riccomagno et al. 2005). Regardless of the precise origin of cochlear duct progenitors, by embryonic day 11, the ventral cochlear outgrowth expresses a broad domain of Sox2 and Jag1 that will eventually resolve into a central Sox2‐expressing prosensory domain that will give rise of the organ of Corti, a stripe of Jag1 running along the neural side of the cochlear duct (closest to the spiral ganglion) and a stripe of Bmp4 expression running along the abneural side of the cochlear duct (Ohyama et al. 2010; Wu & Kelley, 2012; Fig. 3). In the following sections, we describe how the prosensory domain exits the cell cycle, receives patterning information and differentiates into the organ of Corti.
Figure 3.
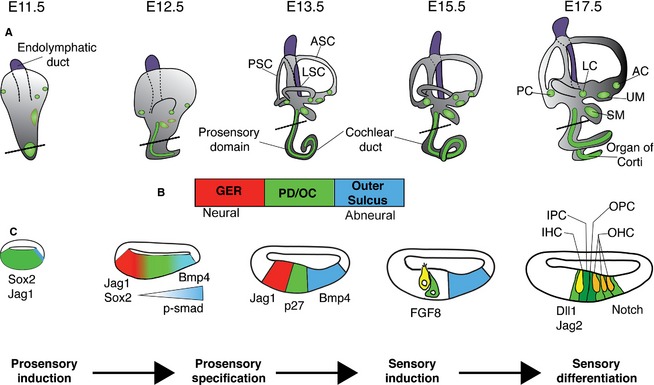
Development and specification of the prosensory domain (PD) and the organ of Corti (OC). (A) Time course of inner ear development, viewed from the lateral side of the embryo. Between E11.5 and E17.5, the cochlear duct elongates from the ventral wall of the otocyst. Between E12.5 and E13.5, the plates in the vestibular portion undergo extensive rearrangement to form the semicircular canals with cristae (green circles) developing at their base. By E15.5, the vestibular labyrinth and its associated sensory organs have developed (cristae and maculae are shown in green, the approximate position of maculae at E11.5 and E12.5 are shown with red outlines), while the OC continues to differentiate. (B) The bar represents the orientation of the cochlear sections shown in (C). The PD, which gives rise to the OC, is flanked by non‐sensory tissue. On the neural side the PD is flanked by the greater epithelial ridge (GER; red) and on the abneural side by the cells that will form the outer sulcus (blue). (C) Schematic diagrams of cochlear sections at the level of the dotted lines shown in (A). Initially the sensory competent region of the cochlea expresses Sox2 and Jag1. A gradient of BMP4 (visualized by the readout of phosphorylated SMAD1/5/8) from the abneural side of the cochlea refines the prosensory region as development proceeds. By E13.5, cells in the PD exit the cell cycle and contain all the progenitors for the OC. At E14.5, at the border between the GER and the PD, inner hair cells (IHCs) in the base of the cochlea begin to differentiate in response to an as‐yet unidentified signal. As IHCs differentiate, they become a source of FGF8, which together with Notch signaling will help establish the patterning of the OC. By E17.5, the base of the cochlea exhibits its final pattern of one row of IHCs and three rows of outer hair cells (OHCs). Over the next 2 days, this patterning will extend throughout the length of the cochlea. The endolymphatic duct, which grows out from the medial side of the otocyst, is colored purple to distinguish it from the rest of the inner ear. AC, anterior crista ampullaris; ASC, anterior semicircular canal; IPC, inner pillar cell; LC, lateral crista ampullaris; LSC, lateral semicircular canal; OPC, outer pillar cell; PC, posterior crista ampullaris; PSC, posterior semicircular canal; SM, saccular macula; UM, utricular macula.
Coordination of cell cycle exit and differentiation in the cochlea
The mammalian organ of Corti is one of the most precisely patterned structures in vertebrates, showing a level of cellular organization comparable to the almost crystalline arrangement of ommatadia in the insect compound eye. Three rows of outer hair cells and one row of inner hair cells run along the length of the cochlear duct, with each cell surrounded by dedicated types of supporting cells: inner phalangeal and border cells surround the inner hair cells; and Deiters’ cells surround the outer hair cells. In addition, an inner and outer pillar cell separate these two domains and form the tunnel of Corti (Fig. 4). This precise arrangement of cells is repeated hundreds of times along the length of the cochlea in an invariant fashion. To form such a sophisticated pattern, strict control of proliferation and differentiation of hair cell and supporting cells progenitors is necessary. As described above, the organ of Corti is derived from a prosensory domain of Sox2 + progenitors located in the ventral region of the otocyst. After an initial period of proliferation and extension (between E10 and E12 in the mouse), the progenitor cells of the prosensory domain undergo cell cycle exit at E12.5, starting in the apex (tip) of the cochlear duct (Ruben, 1967; Matei et al. 2005; Doetzlhofer et al. 2006). A wave of cell cycle exit then proceeds along the prosensory domain from apex to base over the next 2–2.5 days, with a few cells in the most basal region completing their final division between E14.5 and E15.0 (Fig. 3).
Figure 4.
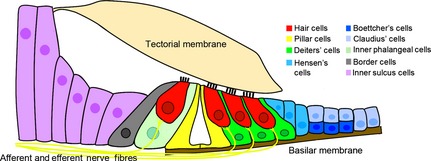
The structure of the organ of Corti. The organ of Corti rests on the basilar membrane of the cochlear duct. It consists of epithelial cells that are varied in both structure and function. Cochlear hair cells (red) can be anatomically and functionally divided into inner and outer hair cells. The inner hair cells detect sound and transmit the information to the brain via the afferent nerves of the cochleovestibular (VIIIth) ganglion, while outer hair cells are important in amplification of sound and receive efferent innervation from the brainstem. Each hair cell is surrounded by dedicated kinds of supporting cells: inner phalangeal and border cells (light green and gray, respectively) surround the inner hair cells, and Deiters’ cell (green) surround the outer hair cells. In addition, an inner and outer pillar cell (yellow) separate these two domains and form the tunnel of Corti (white area between the pillar cells). Immediately next to the Deiters’ cell are the Hensen's cells (sky‐blue), which have been proposed to modulate the interaction between outer hair cells and the tectorial membrane. The tectorial membrane is an acellular sheet secreted largely by the inner sulcus cells (light purple) in the greater epithelial ridge. Next to the Hensen's cells are the Claudius’ cells (light blue). Cells lying beneath Claudius’ cells and the basilar membrane are known as Boettcher's cells (dark blue). These two types of cells have been reported to maintain the microenvironment of the cochlea, such as Na+ absorption or nitric oxide secretion.
A number of cell cycle regulators have been shown to regulate this process in the cochlea. Prior to cell cycle exit, Sox2‐expressing prosensory progenitors appear to require canonical Wnt signaling to maintain their proliferation, and there is some evidence to implicate CyclinD1 in this process (Jacques et al. 2012). Starting at about embryonic day 12–13.5 in the mouse, the cyclin‐dependent kinase inhibitor p27kip1 (Cdkn1b) is upregulated in the cochlea in an apical‐to‐basal gradient that closely follows the gradient of cell cycle exit (Chen & Segil, 1999; Lee et al. 2006). Although p27kip1 is commonly regulated at the post‐transcriptional level, the apical‐to‐basal upregulation of p27kip1 in the cochlea is also regulated transcriptionally (Lee et al. 2006). This apical‐to‐basal pattern of cell cycle exit is disrupted in p27 kip1 mutant mice, with both extra hair cells and supporting cells being produced (Chen & Segil, 1999; Lowenheim et al. 1999; Kanzaki et al. 2006). After cell cycle exit, p27kip1 is most strongly expressed in post‐mitotic hair cells, while other cyclin‐dependent kinase inhibitors such as p19ink4d (Cdkn2d) and p21cip1 (Cdkn1a) are also expressed in hair cells and supporting cells, and their combined action is necessary to maintain the post‐mitotic state and viability of both cell types (Laine et al. 2007; Schimmang & Pirvola, 2013). In addition, members of the pocket protein family (Rb1, Rbl1/p107, Rbl2/p130) also play a role in maintaining the post‐mitotic state of hair cells and supporting cells. (Rocha‐Sanchez & Beisel, 2007). Deletion of Rb1 in hair cells leads to their inappropriate re‐entry into the cell cycle (Sage et al. 2005), which leads to rapid cell death in the cochlea but not the vestibular system (Sage et al. 2006; Weber et al. 2008). Deletion of Rb1 in postnatal supporting cells also permits cell cycle re‐entry (Yu et al. 2010), but again the proliferating supporting cells begin to die after a week. In contrast, germline deletion of Rbl2/p130 causes the appearance of supernumerary hair cells and supporting cells during development (Rocha‐Sanchez et al. 2011), and occasional unscheduled cell cycle re‐entry of supporting cells in the adult. However, in this case, the proliferating cells appear to survive and Rbl2 null mice do not have significantly compromised hearing (Rocha‐Sanchez et al. 2011). This suggests that different members of the Rb family may play different roles during development and maintenance of the post‐mitotic state during cochlear development.
Cell cycle exit and differentiation are typically tightly coupled during development, with differentiation signals frequently driving cells out of the cell cycle. However, in the differentiating organ of Corti, cell cycle exit and differentiation are uncoupled from one another in a striking way. At E13.5, cells in the mid‐basal part of the cochlea duct start to differentiate into hair cells and upregulate the bHLH transcription factor Atoh1 (Chen et al. 2002; Cai et al. 2013). This region of differentiating cells quickly spreads down to the base of the cochlea and also progresses towards the apex over the subsequent 3–4 days. In other words, the first cells to exit the cell cycle at the apex of the cochlea at E12.5 are the last ones to differentiate into hair cells just before birth. In contrast, some of the last cells to exit the cell cycle in the mid‐basal region of the cochlea are the first progenitors to differentiate into hair cells. At present, nothing is known about the signals that initiate and propagate the wave of cell cycle exit in the cochlear duct, or of cellular processes that occur in the post‐mitotic progenitors in the apex of the cochlear duct during their quiescent 5–6‐day period before differentiating.
In the last few years, some of the signals that regulate the basal‐to‐apical gradient of differentiation of the prosensory domain have been characterized. Initially, knockout mouse experiments suggested a role for the transcription factors Neurog1 and Neurod1 in these gradients. First, the cochlear prosensory region of Neurog1 mutant mice shows a precocious exit from the cell cycle about 24–36 h earlier than normal, and hair cell differentiation starts in the apical, not the basal, region of the Neurog1 mutant cochlea (Matei et al. 2005). This disrupted pattern of hair cell differentiation is also seen in Neurod1 mutant mice (Jahan et al. 2010). A clue to the mechanism underlying these defects came from the observation that Neurod1 and Neurog1 are not expressed in the cochlea at detectable levels during differentiation of the prosensory domain. They are, however, expressed in progenitors of the developing spiral ganglion that runs along the length of the cochlear duct, and both Neurod1 and Neurog1 mutant mice have spiral ganglia that are either greatly reduced or completely absent (Ma et al. 1998, 2000; Liu et al. 2000; Kruger et al. 2006). This suggests that signals from the developing spiral ganglion may regulate the timing of differentiation of cochlear hair cells, and that Neurod1 and Neurog1 regulate this process indirectly by regulating spiral ganglion formation.
Several recent studies have proposed that Shh may be one of the signals that regulate differentiation of the cochlear prosensory domain. First, Shh is expressed in the developing spiral ganglion (Liu et al. 2010; Bok et al. 2013) and is downregulated from basal parts of the spiral ganglion at a similar time to the appearance of the first Atoh1‐expressing hair cell progenitors. Second, Shh has been shown to inhibit hair cell formation in vitro (Driver et al. 2008). Third, conditional inactivation of Shh in the spiral ganglion or inactivation of the Shh receptor Smo in the cochlea duct lead to a shorter cochlea in which prosensory cells exit the cell cycle and differentiate prematurely, with both Atoh1 expression and hair cell differentiation following an apical‐to‐basal gradient instead of the normal basal‐to‐apical gradient (Bok et al. 2013; Tateya et al. 2013). Together, these observations indicate that Shh may pay a role in promoting proliferation and preventing premature hair cell differentiation in the cochlear duct, although it is not yet clear whether Shh signaling is acting directly to suppress Atoh1 expression and hair cell differentiation of prosensory progenitors or whether it does so indirectly by regulating an intermediate signal in the cochlear epithelium.
Signals that pattern the cochlear duct to form the organ of Corti
As the cochlear duct begins to elongate from the ventral side of the otocyst, it already contains the molecular signals that will initiate the patterning of the prosensory domain. From E12.5 onward, the asymmetrical distribution of patterning signals is translated into three distinct regions in the cochlear duct (Fig. 3). On the neural side, Jag1 expression becomes restricted to the non‐sensory tissue adjacent to the prosensory domain (Ohyama et al. 2010). In addition to Jag1 these cells also express Lunatic Fringe (LFng) and a gradient of Fgf10 that decreases towards the middle of the cochlea (Cantos et al. 2000; Burton et al. 2004; Ohyama et al. 2010; Wu & Kelley, 2012). These cells form a non‐sensory region adjacent to the organ of Corti, termed Kölliker's organ (Kelley, 2007), and these cells eventually form the inner sulcus of the cochlea. The prosensory cells between Kölliker's organ and the outer sulcus begin to express the cell cycle inhibitor p27 kip1 (Cdkn1b) as they exit the cell cycle (Chen & Segil, 1999; Lee et al. 2006). They also express the bHLH transcriptional repressors Hey1 and Hey2, which are canonical downstream effectors of the Notch pathway (Hayashi et al. 2008a; Li et al. 2008; Doetzlhofer et al. 2009). Flanking the Sox2/Jag1‐positive domain on the abneural side of the cochlea is a narrow strip of non‐sensory cells that express Bmp4. As development progresses, the Bmp4‐positive region expands and these cells will give rise to the future outer sulcus. By E13.5, Kölliker's organ, the prosensory domain and the future outer sulcus are three discrete territories in the ventral wall of the cochlea, with sharp boundaries between them defined by their molecular composition (Ohyama et al. 2010). Sox2 remains strongly expressed by prosensory cells (Kiernan et al. 2005b) and, to a lesser extent, by Jag1‐positive cells, suggesting that non‐sensory cells in Kölliker's organ might retain some generic progenitor capacity.
The formation of all prosensory patches of the bird inner ear is marked by the co‐expression of Sox2, Jag1 and Bmp4, as are the cristae in mammals (Oh et al. 1996; Wu & Oh, 1996; Morsli et al. 1998). In the mammalian maculae, Sox2, Jag1 and Lfng are expressed together, and Jag1 and Sox2 are necessary for the sensory patches to develop properly (Kiernan et al. 2005b, 2006). However, the mammalian organ of Corti is unique in that, as described above, Sox2, Jag1, Bmp4 and Lfng are not co‐expressed by the prosensory cells themselves at the time of hair cell differentiation, but are distributed in the adjacent non‐sensory tissues and their integration in the middle of the cochlear duct results in formation of the prosensory domain. This suggests that the interactions between Sox2, Jag1, BMP4 and FGFs likely have a different role in the cochlea vs. the mouse vestibular organs and all sensory organs of birds. We will next look at the evidence for the involvement of these signals in the specification of the prosensory domain in the mammalian cochlea.
The role of Sox2 in formation of the organ of Corti
Sox2 (SRY‐related HMG box) is a high‐mobility group‐related transcription factor. It is a marker for many types of stem cells and is expressed by neural progenitor cells at different stages of development (Liu et al. 2013; Abdelalim et al. 2014). In the inner ear, Sox2 is one of the earliest markers for all prosensory patches and is expressed in the sensory epithelium of all six sensory regions (Kiernan et al. 2005b; Dabdoub et al. 2008; Kelley et al. 2009). Mice carrying hypomorphic mutations of Sox2 do not develop sensory hair cells or supporting cells, suggesting that Sox2 is necessary for prosensory specification (Kiernan et al. 2005b). Mutations in the human SOX2 locus are also associated with a number of developmental defects including sensorineural hearing loss (Kelberman et al. 2006). Interestingly, forced overexpression of Sox2 in the cochlea does not result in ectopic hair cell formation, but rather in the suppression of hair cell fates and neuronal differentiation (Dabdoub et al. 2008; Puligilla et al. 2010). It has been proposed that this is due to mutual repression between Sox2 and Atoh1, the transcription factor that initiates hair cell differentiation (Dabdoub et al. 2008), and may reflect a role for Sox2 in establishing neural vs. sensory competence in the ear (Ahmed et al. 2012a,b; Raft & Groves, 2015). In all sensory patches, the Notch ligand Jag1 is co‐expressed with Sox2, and it has been suggested that Sox2 expression itself is dependent on Notch signaling (Dabdoub et al. 2008; Pan et al. 2010, 2013). Forced expression of the Notch intracellular domain (NICD) in non‐sensory regions of the cochlea induces ectopic Sox2 expression (Hartman et al. 2010; Pan et al. 2010). However, Sox2 continues to be expressed in the absence of canonical Notch signaling in the cochlea, and some other prosensory markers are present in the cochlea of Notch pathway mutants (Basch et al. 2011). Taken together, these data indicate that Sox2 is necessary for sensory patch induction, and that its expression can be regulated by components of the Notch signaling pathway.
The role of FGF signaling in formation of the organ of Corti
Like Sox2 and Jag1, Fgf10 is also expressed in all vestibular sensory patches and is necessary for their specification (Pirvola et al. 2000; Pauley et al. 2003). Targeted mutations of Fgfr1 or Fgf10 result in severe dysgenesis of the vestibular labyrinth and truncation of the semicircular canals (Pirvola et al. 2000; Pauley et al. 2003). In addition, Fgfr1 mutants have only a rudimentary cochlea (Pirvola et al. 2000). Because of its early asymmetrical localization on the abneural side of the cochlea where it forms a sharp border with the prosensory domain, and its requirement for vestibular sensory development, Fgf10 is a potential candidate for cochlear prosensory specification. However, Fgf10 mutants do not display obvious defects in cochlear hair cell or supporting cell formation (Pauley et al. 2003), suggesting that the effect observed in the Fgfr1 mutants is mediated by other members of the Fgf family. Recent work has shown that Fgf20 is expressed in the prosensory region of the cochlea overlapping Sox2 expression (Hayashi et al. 2008b; Huh et al. 2012). The time and place of Fgf20 expression are consistent with a role in prosensory specification (Groves & Fekete, 2012). Using specific function‐blocking antibodies against Fgf20, Hayashi and colleagues observed a phenotype reminiscent of the Fgfr1 mutants: a disruption and severe reduction in the formation of hair cells and supporting cells (Hayashi et al. 2008b; Munnamalai et al. 2012). The fact that hair cells and supporting cells still form after blocking Fgf20 seems to indicate that either the effect of the antibody is partial, or that a prosensory domain is specified but perhaps reduced in size. More recent evidence suggests that the latter may be the case; Fgf20 knockout mice develop a subset of sensory cells along the cochlea (Huh et al. 2012). Instead of the normal pattern consisting of one row of inner hair cells and three rows of outer hair cells, these mice exhibit a reduction in the number of outer hair cells and their associated supporting cells, suggesting this signal is preferentially required for the differentiation of the outer hair cell compartment (Huh et al. 2012). Interspersed in the patches of missing hair cells are Sox2‐positive cells that the authors suggest to be undifferentiated prosensory progenitors. Interestingly, there is a short window of time where these mutants can be rescued by the addition of exogenous Fgfs, and this time window (between E13.5 and E14.5) is consistent with the onset of hair cell differentiation (Huh et al. 2012).
The role of BMP signaling in formation of the organ of Corti
Bone morphogenetic proteins are a family of secreted molecules that belong to the TGFβ superfamily. BMPs can act as morphogens generating a graded response over small distances (Affolter & Basler, 2007; Plouhinec et al. 2011). The downstream effectors of BMPs are the SMAD proteins, and one of the readouts of BMP activity is the detection of phosphorylated SMADs (Fig. 3). Bmp4 is expressed in all sensory patches in birds (Wu & Oh, 1996), and it is necessary for the patterning of the sensory and non‐sensory portions of each crista ampullaris at the base of the mammalian semicircular canals (Morsli et al. 1998; Chang et al. 2008). Unfortunately, Bmp4 mutant mice die too early to analyze the cochlear phenotype, and inner ear‐specific conditional Bmp4 mutants that have been generated thus far fail to remove the protein from the non‐sensory regions of the cochlea where it is expressed (Chang et al. 2008). Nevertheless, Bmp4 expression suggests that it could play a pivotal role in the specification of the cochlear prosensory domain. As stated earlier, Bmp4 is one of the early proteins asymmetrically expressed as the cochlear duct begins to grow (Ohyama et al. 2010). As Bmp4 expression extends throughout the future outer sulcus flanking the prosensory domain on its abneural side, a gradient of phosphorylated SMAD1/5/8 can be detected extending from the border of the outer sulcus towards the neural side of the cochlear duct (Ohyama et al. 2010). When the levels of BMP are reduced throughout the cochlea by conditional deletion of the BMP type I receptors Alk3 and Alk6, the cells on the abneural side are now exposed to low levels of BMP and therefore acquire a fate corresponding to Kölliker's organ (Ohyama et al. 2010). Moreover, culture experiments where cochlear explants were exposed to varying concentrations of BMP4 suggest that this gradient can specify sensory and non‐sensory regions of the cochlea in a dose‐dependent manner (Ohyama et al. 2010). Intermediate levels of BMP signals are responsible for the specification of prosensory fates, whereas low doses specify Kölliker's organ and high doses generate outer sulcus and inhibit Kölliker's organ gene expression (Ohyama et al. 2010).
The role of Notch‐Jagged signaling in formation of the organ of Corti
The Notch ligand Jagged1 is expressed in all sensory patches in the inner ear together with Sox2 (Brooker et al. 2006; Kiernan et al. 2006). However, in the mammalian cochlea, Jag1 protein expression quickly restricts to the non‐sensory domain on the neural side of the cochlea and is not co‐expressed with markers of the prosensory domain (Ohyama et al. 2010). This expression pattern is consistent with a Notch‐mediated lateral induction signal that could specify the adjacent prosensory domain. Conditional Jag1 mutant mice have dramatic vestibular phenotypes with varying degrees of truncation in the semicircular canals, and reduction or absence of the cristae and maculae (Brooker et al. 2006; Kiernan et al. 2006). The cochlea of Jag1 conditional mice have reduced numbers of hair cells and supporting cells, with basal regions of the cochlea lacking both cell types and apical regions containing a partial row of inner hair cells (Brooker et al. 2006; Kiernan et al. 2006). Activation of the canonical Notch signaling pathway by ectopic expression of the Notch1 intracellular domain (N1ICD) is able to generate ectopic sensory tissue in both birds and mammals (Daudet & Lewis, 2005; Daudet et al. 2007; Hartman et al. 2010; Pan et al. 2010). Ectopic expression of JAG1 in the chick otocyst is also able to stabilize SOX2 expression, although it appears unable to induce it de novo (Neves et al. 2011). However, activation of Notch signaling in the E13.5 mouse cochlea is not sufficient to elicit hair cell or supporting cell fates in the non‐sensory regions of the mammalian cochlea (Pan et al. 2010, 2013). Although forced expression of N1ICD does upregulate the expression of Sox2 (Pan et al. 2010, 2013), this is not enough to induce the expression of other prosensory markers (Dabdoub et al. 2008; Basch et al. 2011). The strongest argument against the requirement for Jag1 or Notch signaling in the specification of the cochlear prosensory domain derives from the observation that in RBPJk mutants, where the canonical downstream effector of the Notch pathway has been conditionally deleted, hair cells and supporting cells continue to form (Basch et al. 2011; Yamamoto et al. 2011). Furthermore, the patterning of the prosensory region and the non‐sensory regions of the cochlea is not disrupted in these mutants (Basch et al. 2011). These results again seem to suggest a unique mode of induction of the organ of Corti distinct from other mammalian ear sensory organs, with Jag1‐Notch signaling appearing to be necessary for induction of the vestibular organs in whose prosensory patches Jag1 continues to be expressed, but is not necessary for the induction of the cochlear prosensory domain, from which Jag1 is excluded by the time the first specific prosensory markers are induced.
Regulation of hair cell differentiation in the organ of Corti
The differentiation of hair cells and supporting cells in the organ of Corti begins between E13 and E14 starting close to the base of the cochlea. It quickly extends towards the base of the cochlea, and over the next 4 days it spread towards the apex (Li & Ruben, 1979; Lim & Anniko, 1985; Chen et al. 2002; Montcouquiol & Kelley, 2003). The bHLH transcription factor Atoh1 (Jarman & Groves, 2013) is one of the earliest markers to be expressed in differentiating hair cells (Woods et al. 2004; Mulvaney & Dabdoub, 2012; Cai et al. 2013). Atoh1 null mice fail to develop hair cells in the inner ear (Bermingham et al. 1999; Chen et al. 2002; Pan et al. 2011), whereas overexpression of Atoh1 is sufficient to induce ectopic hair cells in cochlear cultures (Zheng & Gao, 2000). The initial reports of Atoh1 expression in the cochlea were somewhat dependent on the methods used to visualize it. A LacZ reporter that replaced the Atoh1 coding region indicated a broad expression of Atoh1 throughout the cochlea at E13, followed by a restriction of expression to the differentiating hair cells (Woods et al. 2004), whereas in situ hybridization and antibody staining suggested the initial expression of Atoh1 was in differentiating hair cells at the mid‐base of the cochlea at E14 (Chen et al. 2002). More recently, the use of EGFP‐tagged knock‐in mouse lines that recapitulate endogenous Atoh1 protein expression have shown its initial expression precedes the differentiation of hair cells (Cai et al. 2013; Cai & Groves, 2014). More precisely, it starts just before differentiation begins and it is restricted to a subset of progenitors in the prosensory domain, rather than being expressed generally throughout the cochlea (Cai et al. 2013). Consistent with the timing of its initial expression, lineage analysis using an Atoh1 CRE line revealed that progenitor cells expressing Atoh1 are able to generate both hair cells and supporting cells (Yang et al. 2010). However, the variable number of supporting cells labeled in different inner ear sensory patches in these experiments, and the lack of a clear pattern to the labeled supporting cells, suggests that the onset of Atoh1 expression happens shortly before hair cell progenitors commit to a hair cell fate but that cell–cell interactions (likely through Notch signaling; see below) can re‐direct some of these progenitors towards a supporting cell fate (Driver et al. 2013). Given the precise matching of hair cell and supporting cell numbers in the cochlea compared with other sensory organs of the inner ear, it is likely that the conversion of hair cell progenitors to supporting cells would occur more frequently in the organ of Corti than in other inner ear sensory organs. Consistent with this idea, the largest number of Atoh1 CRE‐labeled supporting cells can be found in the cochlea (Yang et al. 2010; reviewed in Cai & Groves, 2014).
The signals responsible for the onset of Atoh1 expression are an active area of investigation (Mulvaney & Dabdoub, 2012). Wnts and BMPs have been shown to regulate Atoh1 gene or protein expression in a variety of tissues (Ebert et al. 2003; Tsuchiya et al. 2007; Aragaki et al. 2008; Zhao et al. 2008; Kamaid et al. 2010; Shi et al. 2010), or to regulate hair cell production in the cochlea (Jacques et al. 2012, 2014), and functional Lef/Tcf‐binding sites have been found in the 3′ autoregulatory enhancer of Atoh1 (Shi et al. 2010). The regulation of Atoh1 expression by the Notch signaling pathway in the inner ear is also well documented (Kiernan, 2013). Two canonical Notch effectors, Hey1 and Hey2, are expressed in the prosensory domain of the cochlea, and their expression becomes restricted to a subset of supporting cells as hair cell differentiation proceeds (Hayashi et al. 2008a,b; Li et al. 2008; Doetzlhofer et al. 2009). Hey1 and Hey2 encode bHLH transcriptional factors that belong to the Hes/Hey family of transcriptional repressors. These genes are known to repress the hair cell differentiation program in progenitor cells (Li et al. 2008; (Tateya et al. 2011). These observations led to the idea that Notch signaling could be responsible for the early onset of Atoh1 expression (Hayashi et al. 2008a,b). However, the timing of Atoh1 expression in the cochlea is not affected in mice where canonical Notch signaling has been conditionally inactivated (Basch et al. 2011). Furthermore, Hey1 and Hey2 continue to be expressed in the prosensory domain of these mice, suggesting that Notch signaling is dispensable for the initiation of Atoh1 expression, and also that other signals are contributing to the regulation of Hey1 and Hey2 expression (Basch et al. 2011).
As described above, recent evidence points to the Hedgehog signaling pathway as a potential candidate for regulation Atoh1 expression and hair cell differentiation. Components of the Hedgehog pathway are expressed at the right place and time (Driver et al. 2008), and truncating mutants of the transcription factor Gli3, a downstream effector of HH, result in an increase in the size of the sensory epithelium (Driver et al. 2008). Conversely, treatment of cochlear explants with exogenous SHH inhibits the formation of hair cells (Driver et al. 2008). Conditional inactivation of Shh using different spiral ganglion‐specific Cre drivers resulted in a reversal of the differentiation gradient in the cochlea, with premature differentiation of hair cells starting at the apex (Bok et al. 2013; Tateya et al. 2013). Interestingly, Hey1/2 double mutants exhibit premature hair cell differentiation and an increased number of hair cells, reminiscent of the Shh mutant phenotype (Benito‐Gonzalez & Doetzlhofer, 2014). Moreover, cochlear culture experiments where SHH was added or inhibited revealed that SHH is required to maintain Hey1 and Hey2 expression (Benito‐Gonzalez & Doetzlhofer, 2014). In addition, it was established that FGF signaling mediates the regulation of Hey1 and Hey2 expression by SHH (Benito‐Gonzalez & Doetzlhofer, 2014). Taken together, these data suggest that Hedgehog signaling prevents premature upregulation of Atoh1 in the differentiating organ of Corti by indirectly maintaining expression of Hey1 and Hey2 through regulation of Fgf signaling.
Generating the pattern of hair cells and supporting cells in the organ of Corti
The cellular mosaic of the avian basilar papilla can be explained through a relatively simple model of Notch‐mediated lateral inhibition (Lewis, 1991, 1998; Adam et al. 1998; Eddison et al. 2000). As differentiation begins, a progenitor cell from the prosensory domain begins to express the Notch ligands Serrate2 and Delta1 in response to an as‐yet unidentified signal. This cell will differentiate into a hair cell (the primary cell fate) and will signal to its neighboring cells. As a consequence of Notch activation, the signal‐receiving cell will express Hey and Hes transcriptional repressors, which block the hair cell differentiation program and the neighboring cell adopts a secondary cell fate to become a supporting cell. The repetition of this process throughout the basilar papilla will result in the generation of a regular mosaic of hair cells and supporting cells (Eddison et al. 2000; Daudet & Lewis, 2005; Daudet et al. 2009), although the precise ratio of hair cells and surrounding supporting cells varies in different regions of the basilar papilla (Goodyear et al. 1995; Goodyear & Richardson, 1997).
Over the past decade, many groups have shown that Notch signaling also plays a pivotal role in patterning the organ of Corti (Lanford et al. 1999, 2000; Kiernan et al. 2005a; Brooker et al. 2006; Doetzlhofer et al. 2009; Kiernan, 2013). However, the asymmetrical cellular complexity in the mammalian cochlea cannot be explained simply through lateral inhibition. Instead of a regular mosaic of alternating hair cells and supporting cells, two pillar cells separate the inner hair cell region from the outer hair cells, and specific supporting cell types surround each individual hair cell, with Deiters’ cells adjacent to outer hair cells, and border cells and inner phalangeal cells surrounding the inner hair cells. The different types of supporting cells in the organ of Corti have morphological distinctions that correspond to positional and functional characteristics. In addition, each type of supporting cell can be assigned a molecular identity, which is defined by a specific combination of Hey and Hes gene expression. For example, Deiters’ cells express Hes5, Hey1 and HeyL, while Hensen's cells express Hes1 and Hey1 (Zheng et al. 2000; Zine et al. 2001; Hayashi et al. 2008a; Li et al. 2008; Doetzlhofer et al. 2009; Hartman et al. 2009; Murata et al. 2009). The different combinations of Hey and Hes genes in supporting cells are expressed in response to Notch signaling, and they repress the expression of Atoh1, preventing the onset of hair cell differentiation (Doetzlhofer et al. 2009; Tateya et al. 2011). According to the lateral inhibition model, one could predict that the absence of Notch signaling would result in all progenitor cells adopting a primary fate and becoming hair cells at the expense of supporting cells. Consistent with this idea, conditional deletion of Notch1 in the inner ear or deletion of Delta1 and Jag2, two Notch ligands expressed in hair cells, leads to an overproduction of hair cells (Kiernan et al. 2005a,b; Brooker et al. 2006). However, not all supporting cells adopt a hair cell fate as the model would predict – for example, pillar cells remain present in the organ of Corti after loss of Notch‐mediated lateral inhibition (Kiernan et al. 2005a; Doetzlhofer et al. 2009). The identity of pillar cells is dependent on expression of Hey2: in the absence of Notch signaling, Hey2 remains present in pillar cells, and only on deletion of Hey2 do pillar cells convert into hair cells when Notch signaling is blocked (Doetzlhofer et al. 2009). These data suggest that other signals are involved in maintaining Hey2 expression and preventing pillar cells from acquiring a hair cell fate. Fgf8 is expressed by inner hair cells adjacent to pillar cells, and loss of Fgfr3 or pharmacological inhibition of FGF receptors leads to abnormal pillar cell development (Hayashi et al. 2007; Jacques et al. 2007; Puligilla et al. 2007). Fgf signaling therefore seems a good candidate to maintain pillar cell identity. However, blocking FGF signaling in cochlear explants does not lead to a reduction of Hey2, or a loss of pillar cell identity. Rather, inhibition of both FGF and Notch signaling results in loss of Hey2 expression and pillar cell identity, suggesting that both pathways act to maintain these supporting cells, perhaps as a way to protect this evolutionary novel cell type through redundant mechanisms (Doetzlhofer et al. 2009).
Development of tonotopy in the mammalian cochlea
As shown in Fig. 1, the hearing organs of many terrestrial vertebrates have elongated during the course of evolution to permit hearing over a wide range of sound frequencies. This ability reaches its apogee in mammals, where different species can hear as low as a few tens of Hz to ultrasonic ranges of over 100 kHz in some bats. The ability to sample a wide sound spectrum is reflected in a tonotopic gradient of sensitivity and response along the length of the cochlear duct, where hair cells in the base of the cochlea respond to sounds of high frequency, while hair cells in the apex respond best to sounds of lower frequency. A number of factors act in concert to tune the response of hair cells so exquisitely: these include gradations in the mechanical properties of the basilar membrane and the tectorial membrane of the cochlear duct, gradients of size and shape of hair cells and their stereociliary bundles, and differences in gene and protein expression in hair cells at different points along the tonotopic axis (Mann & Kelley, 2011). When cochlear development is complete, these properties of the cochlear duct and its hair cells vary systematically along its length and allow discrimination of sounds of different frequencies (Fig. 5).
Figure 5.
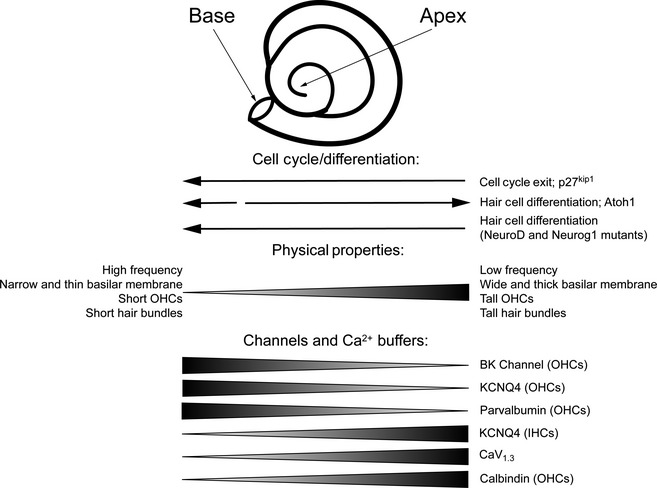
Basal‐to‐apical differences in patterns of cell cycle exit, differentiation and tonotopic properties of the cochlea. Prosensory progenitors exit the cell cycle at the base of the cochlea and express p27kip1 protein in an apical‐to‐basal gradient. However, differentiation of hair cell progenitors begins close to the base of the cochlear duct and spreads down to the apex over the next 4–5 days. This uncoupling of cell cycle exit and differentiation is abolished in a variety of mouse mutants. The physical properties of the cochlear duct and hair cells also vary along the tonotopic axis (basal: high‐frequency; apical: low‐frequency), and tonotopic gradients of ion channels and calcium buffers have also been observed, either in all hair cells or in inner or outer hair cells (IHCs, OHCs).
The classic experiments of von Bekesy demonstrated that the cochlear duct vibrates optimally in response to different frequencies at different positions along its length (von Békésy, 1960). This frequency discrimination is due to the passive properties of the cochlea and can still be observed in dead specimens. It is refined to a great degree by the active properties of hair cells (Hudspeth, 2008, 2014), which in mammals are predominantly exhibited by outer hair cells that employ motor protein‐based amplification (Dallos, 2008; Dallos et al. 2008). The basilar membrane on which the organ of Corti sits changes its mechanical properties along the length of the cochlea, being stiff, thin and narrow at the base, and thicker, wider and more compliant at the apex (Dallos, 1996). The primary components of the basilar membrane are type II and IV collagens, keratin sulfate proteoglycans and a variety of extracellular matrix glycoproteins including fibronectin, tenascin and emilin2 (Cosgrove et al. 1996; Tsuprun & Santi, 1999; Dreiling et al. 2002a,b; Amma et al. 2003) . At present, very little is known about how the distribution and packing of these proteins is regulated along the tonotopic axis to change the thickness, width and stiffness of the basilar membrane. The tectorial membrane that lies above the organ of Corti and contacts outer hair cell stereocilia has a similar morphological variation along the cochlear duct to the basilar membrane, being thin and narrow at the base and wide and thick at the apex. Collagens are again a major component of the tectorial membrane, although several ear‐specific glycoproteins such as α‐ and β‐tectorin and otogelin also contribute to the matrix of glycoproteins (Goodyear & Richardson, 2002), and mutations in these genes can cause deafness in humans and mice (Legan et al. 2000; Simmler et al. 2000; Pfister et al. 2004; Plantinga et al. 2006; Russell et al. 2007; Xia et al. 2010; Gueta et al. 2011; Schraders et al. 2012; Yariz et al. 2012). The organization of collagen fibers varies along the length of the tectorial membrane, although it is not clear to what extent these contribute to changes in tectorial membrane stiffness (Gueta et al. 2006, 2007; Richter et al. 2007).
In addition to the properties of the membranes that lie above and below them, hair cells also show morphological variation along the tonotopic axis of the cochlear duct, for example, in the size of outer hair cells (Engel et al. 2006). The stereocilia of individual hair cells form a precise staircase‐like pattern in each hair bundle, with adjacent rows of stereocilia connected to each other by tip and ankle links (Nayak et al. 2007). The height of each stereociliary bundle is precisely regulated along the tonotopic axis of the cochlea, with hair cells at the base of the cochlea having short stereocilia and the height of the hair bundles increasing (sometimes as much as threefold) in a gradient towards the apex of the cochlea (Wright, 1984; Lim, 1986; Kaltenbach et al. 1994). This trend is also seen in other amniotes (Tilney & Saunders, 1983). The length of hair cell stereocilia is constantly maintained by a dynamic process of actin filament stabilization and growth (Belyantseva et al. 2005; Manor & Kachar, 2008; Peng et al. 2009). This suggests that hair cell bundle length could be regulated by the fine tuning of actin regulatory proteins at either end of the actin filament core in stereocilia (e.g. espin, radixin, Triobp or twinfilin; Manor & Kachar, 2008) or by members of the group of myosins that are present in hair bundles (Myosin1c, 3a, 6, 7a and 15a; Manor & Kachar, 2008). For example, mouse mutants of Myosin15a, which localizes to the tips of stereocilia, have extremely short hair bundles (Anderson et al. 2000; Belyantseva et al. 2005). However, it is not clear how the activity of these actin regulatory proteins is fine‐tuned to create such precise variations in stereocilium length in a given hair bundle and between hair cell bundles at different positions along the basilar membrane. It is also possible that external influences may help set the height of hair bundles, for example, the fact that the tectorial membrane touches the tips of the highest stereocilia in each outer hair cell (Fettiplace & Hackney, 2006) suggests that either signaling or direct physical force could regulate stereocilia height. Alternatively, because mechanotransduction channels are located close to the tips of stereocilia (Beurg et al. 2009; Schwander et al. 2010), calcium entry during mechanotransduction could also interact with and modulate actin regulatory proteins at stereocilia tips (see Manor & Kachar, 2008 for additional discussion).
The fine‐grained changes in hair cell morphology along the tonotopic axis are accompanied by changes in the electrophysiological properties of hair cells (Beurg et al. 2006), and concomitant gradients of protein and mRNA expression of genes that regulate the electrophysiological or calcium‐buffering properties of hair cells. Calcium buffers such as calretinin, calbindin and parvalbumin are expressed in a number of gradients, for example, parvalbumin expression is highest in basal inner hair cells, but lowest in basal outer hair cells (Pack & Slepecky, 1995; Hackney et al. 2005). A number of ion channels are also expressed in gradients in hair cells along the cochlear duct, such as the BK calcium‐activated potassium channel (Engel et al. 2006), the CaV1.3 voltage‐gated calcium channel (Engel et al. 2006) and the KCNQ4 voltage‐gated potassium channel (Beisel et al. 2000, 2005).
When considering all these observed tonotopic differences in hair cells, the outstanding question remains of how such minute cell‐to‐cell changes in size, hair bundle morphology or protein expression could be precisely calibrated along the cochlear duct. As discussed above, external physical forces may play a role, but it is also possible that gradients of signaling molecules, receptors or transcription factors or co‐factors may determine the tonotopic properties of hair cells. Two recent studies searched for differentially expressed genes along the proximal–distal axis of the chicken basilar papilla using RNA‐seq and microarrays (Mann et al. 2014; Thiede et al. 2014). In birds, the proximal region of the basilar papilla responses to higher frequencies similar to the basal part of the mammalian organ of Corti, while distal regions respond to lower frequencies, similar to the apical organ of Corti. These two studies suggested that RA and BMP signaling may play a role in specifying morphology and gene expression of hair cells. BMP7 is enriched in the distal end of the basilar papilla, and addition of BMP7 to proximal portions of the basilar papilla is sufficient to produce changes in cell density, calbindin expression and stereocilia number and length consistent with their adopting a more proximal hair cell identity (Mann et al. 2014). Conversely, blocking BMP activity by addition of the BMP7 antagonist Chordin‐like 1 (CHRDL1) to the distal section of the basilar papilla induced a proximal hair cell phenotype (Mann et al. 2014). Interestingly, BMP7 and CHRDL1 are expressed in opposing gradients in the basilar papilla, suggesting that the sharply varying gradients of BMP signaling might result from the superposition of a BMP signal and its antagonist. In mammals, Bmp7 has also been shown to be expressed in the mouse otocyst and later in the cochlea (Ohyama et al. 2010). However, no inner ear defects were detected in Bmp7 mutant mice (Dudley et al. 1995; Luo et al. 1995) although because these mice exhibit early perinatal death, tonotopic or other ear defects may not have been apparent. Conditional deletion of Bmp7 in the cochlea may help to address this question. In further support of the role of BMP signaling in mammalian tonotopic axis formation, a recent microarray study in the mouse cochlea suggested a possible role of Fst (Follistatin), an antagonist of TGF‐β/BMP signaling, in setting up the base to apex gradient (Son et al. 2012). Unlike in chick, where the CHRDL1 BMP inhibitor is present in the proximal (high‐frequency) region of the basilar papillae, Fst is expressed more strongly in the apex (low‐frequency) region of the mouse cochlea.
Retinoic acid signaling was also shown to regulate the proximal–distal patterning and gene expression in the chick basilar papilla (Thiede et al. 2014). Like the BMP signaling pathway, a complementary expression pattern of source and sink genes establishes a gradient of retinoid signaling along the proximal–distal axis of the basilar papilla. The RA‐synthesizing enzyme RALDH3 is expressed strongly in the proximal region of the basilar papilla, and the RA‐degrading enzyme CYP26C1 is expressed strongly in the distal region of the papilla, although these gradients change and ultimately reverse during the course of embryonic development and after hatching (Thiede et al. 2014). Like BMP7, treatment of distal regions of the basilar papilla with RA is able to transform the hair cells in these regions to an identity more reminiscent of proximal parts of the papilla, while RA receptor antagonists have the opposite effect. It is of note that RA treatment was also able to suppress expression of BMP7 in these experiments, and that BMP7 treatment could downregulate RALDH3 expression (Thiede et al. 2014), suggesting a reciprocal interaction between these two pathways during the establishment of tonotopic organization. In mice, the RA signaling pathway has also been shown to be important for the development of the otocyst and cochlea (Kelley et al. 1993; Raz & Kelley, 1999; Bok et al. 2011), and can regulate the expression of certain hair cell proteins such as prestin (Gross et al. 2011). However, it is not yet clear whether it has a role in tonotopic organization of the mammalian organ of Corti.
Future directions in cochlear development research
The last 10 years have witnessed a flurry of studies that have significantly advanced our understanding of how the cochlea develops. However, each of the topics discussed in this review make clear that much still needs to be discovered. Below we briefly mention some areas that are ripe for further exploration.
Cochlear evolution
Understanding the molecular changes that have accompanied the individuation and enlargement of the basilar papilla during amniote evolution is a fascinating problem. It remains an open question as to whether this enlargement involved the co‐option of existing transcription and growth factor expression domains or whether entirely new patterns of transcription factor expression arose in the amniote lineage in response to new inducing signals (as seen in the emergence of otx1 expression in the lateral semicircular canal during its addition in the transition from lampreys to fish; Hammond & Whitfield, 2006).
Cochlear outgrowth
While the role of the planar cell polarity pathway has been well established in driving the processes of convergent extension and radial intercalation in cochlear development (Kelly & Chen, 2007; Goodrich & Strutt, 2011; Chacon‐Heszele et al. 2012), we know very little of the signals that initiate the outgrowth of the cochlear duct, and almost nothing about the spatial regulation of proliferation and cell shape that give the cochlear duct its characteristic coiled shape. The advent of transgenic mice expressing nuclear‐ and membrane‐bound forms of fluorescent protein, together with advances in tissue clearing (Vogt, 2015) and high‐resolution fluorescent imaging (Udan et al. 2014) will allow a much greater understanding of the cellular processes that regulate cochlear shape.
Cell cycle exit and differentiation in the cochlea
As discussed above, the first prosensory cells to exit the cell cycle at the apex of the cochlear duct are the last ones to differentiate into hair cells and supporting cells (Lee et al. 2006). At present, we know nothing about the transcriptional or post‐transcriptional changes (if any) that occur in these quiescent cells while they are waiting to be induced to form hair cells and supporting cells.
Cochlear patterning
It is now well established that localized sources of FGFs, BMPs, Wnts and Shh are integrated to set up developmental gradients across the developing cochlear duct (Groves & Fekete, 2012). However, it is much less clear how these combined gradients are interpreted, first to give broad developmental domains (e.g. the prosensory domain vs. the non‐sensory epithelium of the roof of the cochlear duct), and second to set sharp boundaries between these developmental territories. Moreover, although the signals that specify the prosensory domain (and subsequently the organ of Corti) are beginning to be characterized, we know far less about signals that induce equally important non‐sensory regions of the cochlear duct, such as the lateral wall (including the stria vascularis), which is crucial for regulation of endolymph composition, or Reissner's membrane, which separates the scala media of the cochlear duct from the scala vestibuli that lies above it (Kelly & Chen, 2009). Interestingly, the formation of the scala vestibuli and scala tympani from mesenchyme that surrounds the cochlear duct follows the same basal‐to‐apical gradient as the differentiation of the cochlea itself (Sher, 1971), but nothing is known about how this differentiation is coordinated with cochlear patterning.
Hair cell and supporting cell identity
As described above, recent evidence has identified some signaling pathways, such as FGF20, that can specify different regions of the organ of Corti (Huh et al. 2012). However, with the exception of pillar cells (Mueller et al. 2002; Jacques et al. 2007; Puligilla et al. 2007; Doetzlhofer et al. 2009), the signals that specify the individual types of hair cells and supporting cell types seen in the organ of Corti (Fig. 4) are far less clear. It is also clear that some plasticity between these cell types can persist into the postnatal animal, for example, the modulation of FGF signaling can cause a conversion of Deiters’ cells into pillar cells (Shim et al. 2005; Mansour et al. 2013), but the molecular mechanisms underlying this transformation are unknown. Recent advances in purification of particular cell populations from the cochlea, together with the increasing ease of deep sequencing on very small populations of cells, means that it will soon become possible to assemble a complete transcriptome of each cell type in the organ of Corti. Such technology can also be used to better interrogate the underlying mechanisms of tonotopic development in the inner ear.
Acknowledgements
The writing of this review was supported in part by NIH grants DC006185 and DC013072 (AKG), and an NIH Diversity Supplement DC013072‐S1 (RMB). The authors declare they have no conflicts of interest.
References
- Abdelalim EM, Emara MM, Kolatkar PR (2014) The SOX transcription factors as key players in pluripotent stem cells. Stem Cells Dev 23, 2687–2699. [DOI] [PubMed] [Google Scholar]
- Adam J, Myat A, Le Roux I, et al. (1998) Cell fate choices and the expression of Notch, Delta and Serrate homologues in the chick inner ear: parallels with Drosophila sense‐organ development. Development 125, 4645–4654. [DOI] [PubMed] [Google Scholar]
- Affolter M, Basler K (2007) The Decapentaplegic morphogen gradient: from pattern formation to growth regulation. Nat Rev Genet 8, 663–674. [DOI] [PubMed] [Google Scholar]
- Ahmed M, Wong EY, Sun J, et al. (2012a) Eya1‐Six1 interaction is sufficient to induce hair cell fate in the cochlea by activating Atoh1 expression in cooperation with Sox2. Dev Cell 22, 377–390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ahmed M, Xu J, Xu PX (2012b) EYA1 and SIX1 drive the neuronal developmental program in cooperation with the SWI/SNF chromatin‐remodeling complex and SOX2 in the mammalian inner ear. Development 139, 1965–1977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alsina B, Abello G, Ulloa E, et al. (2004) FGF signaling is required for determination of otic neuroblasts in the chick embryo. Dev Biol 267, 119–134. [DOI] [PubMed] [Google Scholar]
- Amma LL, Goodyear R, Faris JS, et al. (2003) An emilin family extracellular matrix protein identified in the cochlear basilar membrane. Mol Cell Neurosci 23, 460–472. [DOI] [PubMed] [Google Scholar]
- Anderson DW, Probst FJ, Belyantseva IA, et al. (2000) The motor and tail regions of myosin XV are critical for normal structure and function of auditory and vestibular hair cells. Hum Mol Genet 9, 1729–1738. [DOI] [PubMed] [Google Scholar]
- Aragaki M, Tsuchiya K, Okamoto R, et al. (2008) Proteasomal degradation of Atoh1 by aberrant Wnt signaling maintains the undifferentiated state of colon cancer. Biochem Biophys Res Commun 368, 923–929. [DOI] [PubMed] [Google Scholar]
- Basch ML, Ohyama T, Segil N, et al. (2011) Canonical Notch signaling is not necessary for prosensory induction in the mouse cochlea: insights from a conditional mutant of RBPjkappa. J Neurosci 31, 8046–8058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beisel KW, Nelson NC, Delimont DC, et al. (2000) Longitudinal gradients of KCNQ4 expression in spiral ganglion and cochlear hair cells correlate with progressive hearing loss in DFNA2. Brain Res Mol Brain Res 82, 137–149. [DOI] [PubMed] [Google Scholar]
- Beisel KW, Rocha‐Sanchez SM, Morris KA, et al. (2005) Differential expression of KCNQ4 in inner hair cells and sensory neurons is the basis of progressive high‐frequency hearing loss. J Neurosci 25, 9285–9293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- von Békésy G (1960) Experiments in Hearing. New York, NY: McGraw. [Google Scholar]
- Belyantseva IA, Boger ET, Naz S, et al. (2005) Myosin‐XVa is required for tip localization of whirlin and differential elongation of hair‐cell stereocilia. Nat Cell Biol 7, 148–156. [DOI] [PubMed] [Google Scholar]
- Benito‐Gonzalez A, Doetzlhofer A (2014) Hey1 and Hey2 control the spatial and temporal pattern of mammalian auditory hair cell differentiation downstream of Hedgehog signaling. J Neurosci 34, 12 865–12 876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bermingham NA, Hassan BA, Price SD, et al. (1999) Math1: an essential gene for the generation of inner ear hair cells. Science 284, 1837–1841. [DOI] [PubMed] [Google Scholar]
- Beurg M, Evans MG, Hackney CM, et al. (2006) A large‐conductance calcium‐selective mechanotransducer channel in mammalian cochlear hair cells. J Neurosci 26, 10 992–11 000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beurg M, Fettiplace R, Nam JH, et al. (2009) Localization of inner hair cell mechanotransducer channels using high‐speed calcium imaging. Nat Neurosci 12, 553–558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bok J, Bronner‐Fraser M, Wu DK (2005) Role of the hindbrain in dorsoventral but not anteroposterior axial specification of the inner ear. Development 132, 2115–2124. [DOI] [PubMed] [Google Scholar]
- Bok J, Chang W, Wu DK (2007a) Patterning and morphogenesis of the vertebrate inner ear. Int J Dev Biol 51, 521–533. [DOI] [PubMed] [Google Scholar]
- Bok J, Dolson DK, Hill P, et al. (2007b) Opposing gradients of Gli repressor and activators mediate Shh signaling along the dorsoventral axis of the inner ear. Development 134, 1713–1722. [DOI] [PubMed] [Google Scholar]
- Bok J, Raft S, Kong KA, et al. (2011) Transient retinoic acid signaling confers anterior‐posterior polarity to the inner ear. Proc Natl Acad Sci USA 108, 161–166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bok J, Zenczak C, Hwang CH, et al. (2013) Auditory ganglion source of Sonic hedgehog regulates timing of cell cycle exit and differentiation of mammalian cochlear hair cells. Proc Natl Acad Sci USA 110, 13 869–13 874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brigande JV, Iten LE, Fekete DM (2000a) A fate map of chick otic cup closure reveals lineage boundaries in the dorsal otocyst. Dev Biol 227, 256–270. [DOI] [PubMed] [Google Scholar]
- Brigande JV, Kiernan AE, Gao X, et al. (2000b) Molecular genetics of pattern formation in the inner ear: do compartment boundaries play a role? Proc Natl Acad Sci USA 97, 11 700–11 706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brooker R, Hozumi K, Lewis J (2006) Notch ligands with contrasting functions: Jagged1 and Delta1 in the mouse inner ear. Development 133, 1277–1286. [DOI] [PubMed] [Google Scholar]
- Brown AS, Epstein DJ (2011) Otic Ablation of Smoothened Reveals Direct and Indirect Requirements for Hedgehog Signaling in Inner Ear Development. Development 138, 3967–3976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown AS, Rakowiecki SM, Li JY, et al. (2015) The cochlear sensory epithelium derives from Wnt responsive cells in the dorsomedial otic cup. Dev Biol 399, 177–187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burton Q, Cole LK, Mulheisen M, et al. (2004) The role of Pax2 in mouse inner ear development. Dev Biol 272, 161–175. [DOI] [PubMed] [Google Scholar]
- Cai T, Groves AK (2014) The role of atonal factors in mechanosensory cell specification and function. Mol Neurobiol DOI: 10.1007/s12035‐014‐8925‐0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cai T, Seymour ML, Zhang H, et al. (2013) Conditional deletion of Atoh1 reveals distinct critical periods for survival and function of hair cells in the organ of Corti. J Neurosci 33, 10 110–10 122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cantos R, Cole LK, Acampora D, et al. (2000) Patterning of the mammalian cochlea. Proc Natl Acad Sci USA 97, 11 707–11 713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chacon‐Heszele MF, Ren D, Reynolds AB, et al. (2012) Regulation of cochlear convergent extension by the vertebrate planar cell polarity pathway is dependent on p120‐catenin. Development 139, 968–978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang W, Lin Z, Kulessa H, et al. (2008) Bmp4 is essential for the formation of the vestibular apparatus that detects angular head movements. PLoS Genet 4, e1000050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen P, Segil N (1999) p27(Kip1) links cell proliferation to morphogenesis in the developing organ of Corti. Development 126, 1581–1590. [DOI] [PubMed] [Google Scholar]
- Chen P, Johnson JE, Zoghbi HY, et al. (2002) The role of Math1 in inner ear development: uncoupling the establishment of the sensory primordium from hair cell fate determination. Development 129, 2495–2505. [DOI] [PubMed] [Google Scholar]
- Chen Z, Montcouquiol M, Calderon R, et al. (2008) Jxc1/Sobp, encoding a nuclear zinc finger protein, is critical for cochlear growth, cell fate, and patterning of the organ of corti. J Neurosci 28, 6633–6641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choo D, Ward J, Reece A, et al. (2006) Molecular mechanisms underlying inner ear patterning defects in kreisler mutants. Dev Biol 289, 308–317. [DOI] [PubMed] [Google Scholar]
- Cole LK, Le Roux I, Nunes F, et al. (2000) Sensory organ generation in the chicken inner ear: contributions of bone morphogenetic protein 4, serrate1, and lunatic fringe. J Comp Neurol 424, 509–520. [DOI] [PubMed] [Google Scholar]
- Cosgrove D, Kornak JM, Samuelson G (1996) Expression of basement membrane type IV collagen chains during postnatal development in the murine cochlea. Hear Res 100, 21–32. [DOI] [PubMed] [Google Scholar]
- Dabdoub A, Puligilla C, Jones JM, et al. (2008) Sox2 signaling in prosensory domain specification and subsequent hair cell differentiation in the developing cochlea. Proc Natl Acad Sci USA 105, 18 396–18 401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dallos P (1996) Overview: cochlear neurobiology In: The Cochlea . (eds Dallos P, Popper AN, Fay RR.), pp. 1–43. New York, NY: Springer. [Google Scholar]
- Dallos P (2008) Cochlear amplification, outer hair cells and prestin. Curr Opin Neurobiol 18, 370–376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dallos P, Wu X, Cheatham MA, et al. (2008) Prestin‐based outer hair cell motility is necessary for mammalian cochlear amplification. Neuron 58, 333–339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daudet N, Lewis J (2005) Two contrasting roles for Notch activity in chick inner ear development: specification of prosensory patches and lateral inhibition of hair‐cell differentiation. Development 132, 541–551. [DOI] [PubMed] [Google Scholar]
- Daudet N, Ariza‐McNaughton L, Lewis J (2007) Notch signalling is needed to maintain, but not to initiate, the formation of prosensory patches in the chick inner ear. Development 134, 2369–2378. [DOI] [PubMed] [Google Scholar]
- Daudet N, Gibson R, Shang J, et al. (2009) Notch regulation of progenitor cell behavior in quiescent and regenerating auditory epithelium of mature birds. Dev Biol 326, 86–100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deol MS (1964) The abnormalities of the inner ear in Kreisler mice. J Embryol Exp Morphol 12, 475–490. [PubMed] [Google Scholar]
- Doetzlhofer A, White P, Lee YS, et al. (2006) Prospective identification and purification of hair cell and supporting cell progenitors from the embryonic cochlea. Brain Res 1091, 282–288. [DOI] [PubMed] [Google Scholar]
- Doetzlhofer A, Basch ML, Ohyama T, et al. (2009) Hey2 regulation by FGF provides a Notch‐independent mechanism for maintaining pillar cell fate in the organ of Corti. Dev Cell 16, 58–69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dreiling FJ, Henson MM, Henson OW Jr (2002a) Immunolabeling type II collagen in the basilar membrane, a pre‐embedding approach. Hear Res 166, 181–191. [DOI] [PubMed] [Google Scholar]
- Dreiling FJ, Henson MM, Henson OW Jr (2002b) The presence and arrangement of type II collagen in the basilar membrane. Hear Res 166, 166–180. [DOI] [PubMed] [Google Scholar]
- Driver EC, Pryor SP, Hill P, et al. (2008) Hedgehog signaling regulates sensory cell formation and auditory function in mice and humans. J Neurosci 28, 7350–7358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Driver EC, Sillers L, Coate TM, et al. (2013) The Atoh1‐lineage gives rise to hair cells and supporting cells within the mammalian cochlea. Dev Biol 376, 86–98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dudley AT, Lyons KM, Robertson EJ (1995) A requirement for bone morphogenetic protein‐7 during development of the mammalian kidney and eye. Genes Dev 9, 2795–2807. [DOI] [PubMed] [Google Scholar]
- Ebert PJ, Timmer JR, Nakada Y, et al. (2003) Zic1 represses Math1 expression via interactions with the Math1 enhancer and modulation of Math1 autoregulation. Development 130, 1949–1959. [DOI] [PubMed] [Google Scholar]
- Eddison M, Le Roux I, Lewis J (2000) Notch signaling in the development of the inner ear: lessons from Drosophila. Proc Natl Acad Sci USA 97, 11 692–11 699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Engel J, Braig C, Ruttiger L, et al. (2006) Two classes of outer hair cells along the tonotopic axis of the cochlea. Neuroscience 143, 837–849. [DOI] [PubMed] [Google Scholar]
- Fekete DM, Wu DK (2002) Revisiting cell fate specification in the inner ear. Curr Opin Neurobiol 12, 35–42. [DOI] [PubMed] [Google Scholar]
- Fettiplace R, Hackney CM (2006) The sensory and motor roles of auditory hair cells. Nat Rev Neurosci 7, 19–29. [DOI] [PubMed] [Google Scholar]
- Fritzsch B (1987) Inner ear of the coelacanth fish Latimeria has tetrapod affinities. Nature 327, 153–154. [DOI] [PubMed] [Google Scholar]
- Fritzsch B (1992) The water‐to‐land transition: evolution of the tetrapod basilar papilla, middle ear and auditory nuclei In: The Evolutionary Biology of Hearing . (eds Webster DB, Fay RR, Popper AN.), pp. 351–375. New York, NY: Springer. [Google Scholar]
- Fritzsch B (2003) The ear of Latimeria chalumnae revisited. Zoology (Jena) 106, 243–248. [DOI] [PubMed] [Google Scholar]
- Fritzsch B, Straka H (2014) Evolution of vertebrate mechanosensory hair cells and inner ears: toward identifying stimuli that select mutation driven altered morphologies. J Comp Physiol A Neuroethol Sens Neural Behav Physiol 200, 5–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fritzsch B, Jahan I, Pan N, et al. (2011) Dissecting the molecular basis of organ of Corti development: where are we now? Hear Res 276, 16–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fritzsch B, Pan N, Jahan I, et al. (2013) Evolution and development of the tetrapod auditory system: an organ of Corti‐centric perspective. Evol Dev 15, 63–79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giraldez F (1998) Regionalized organizing activity of the neural tube revealed by the regulation of lmx1 in the otic vesicle. Dev Biol 203, 189–200. [DOI] [PubMed] [Google Scholar]
- Gleich O, Fischer FP, Köppl C, et al. (2004) Hearing organ evolution and specialization: archosaurs In: Evolution of the Vertebrate Auditory System . (eds Manley GA, Popper AN, Fay RR.), pp. 224–255. New York: Springer. [Google Scholar]
- Goodrich LV, Strutt D (2011) Principles of planar polarity in animal development. Development 138, 1877–1892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goodyear R, Richardson G (1997) Pattern formation in the basilar papilla: evidence for cell rearrangement. J Neurosci 17, 6289–6301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goodyear RJ, Richardson GP (2002) Extracellular matrices associated with the apical surfaces of sensory epithelia in the inner ear: molecular and structural diversity. J Neurobiol 53, 212–227. [DOI] [PubMed] [Google Scholar]
- Goodyear R, Holley M, Richardson G (1995) Hair and supporting‐cell differentiation during the development of the avian inner ear. J Comp Neurol 351, 81–93. [DOI] [PubMed] [Google Scholar]
- Grocott T, Tambalo M, Streit A (2012) The peripheral sensory nervous system in the vertebrate head: a gene regulatory perspective. Dev Biol 370, 3–23. [DOI] [PubMed] [Google Scholar]
- Gross J, Stute K, Fuchs J, et al. (2011) Effects of retinoic acid and butyric acid on the expression of prestin and Gata‐3 in organotypic cultures of the organ of corti of newborn rats. Dev Neurobiol 71, 650–661. [DOI] [PubMed] [Google Scholar]
- Groves AK (2005) The induction of the otic placode In: Development of the Inner Ear . (eds Popper AN, Kelley MW, Wu DK.), pp. 10–42. New York: Springer. [Google Scholar]
- Groves AK, Bronner‐Fraser M (2000) Competence, specification and commitment in otic placode induction. Development 127, 3489–3499. [DOI] [PubMed] [Google Scholar]
- Groves AK, Fekete DM (2012) Shaping sound in space: the regulation of inner ear patterning. Development 139, 245–257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Groves AK, LaBonne C (2014) Setting appropriate boundaries: fate, patterning and competence at the neural plate border. Dev Biol 389, 2–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gueta R, Barlam D, Shneck RZ, et al. (2006) Measurement of the mechanical properties of isolated tectorial membrane using atomic force microscopy. Proc Natl Acad Sci USA 103, 14 790–14 795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gueta R, Tal E, Silberberg Y, et al. (2007) The 3D structure of the tectorial membrane determined by second‐harmonic imaging microscopy. J Struct Biol 159, 103–110. [DOI] [PubMed] [Google Scholar]
- Gueta R, Levitt J, Xia A, et al. (2011) Structural and mechanical analysis of tectorial membrane Tecta mutants. Biophys J 100, 2530–2538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hackney CM, Mahendrasingam S, Penn A, et al. (2005) The concentrations of calcium buffering proteins in mammalian cochlear hair cells. J Neurosci 25, 7867–7875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haddon C, Lewis J (1996) Early ear development in the embryo of the zebrafish, Danio rerio. J Comp Neurol 365, 113–128. [DOI] [PubMed] [Google Scholar]
- Hammond KL, Whitfield TT (2006) The developing lamprey ear closely resembles the zebrafish otic vesicle: otx1 expression can account for all major patterning differences. Development 133, 1347–1357. [DOI] [PubMed] [Google Scholar]
- Hammond KL, Whitfield TT (2011) Fgf and Hh Signalling Act on a Symmetrical Pre‐Pattern to Specify Anterior and Posterior Identity in the Zebrafish Otic Placode and Vesicle. Development, 138, 3977–3987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hammond KL, Loynes HE, Folarin AA, et al. (2003) Hedgehog signalling is required for correct anteroposterior patterning of the zebrafish otic vesicle. Development 130, 1403–1417. [DOI] [PubMed] [Google Scholar]
- Hammond KL, van Eeden FJ, Whitfield TT (2010) Repression of Hedgehog signalling is required for the acquisition of dorsolateral cell fates in the zebrafish otic vesicle. Development 137, 1361–1371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hartman BH, Basak O, Nelson BR, et al. (2009) Hes5 expression in the postnatal and adult mouse inner ear and the drug‐damaged cochlea. J Assoc Res Otolaryngol 10, 321–340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hartman BH, Reh TA, Bermingham‐McDonogh O (2010) Notch signaling specifies prosensory domains via lateral induction in the developing mammalian inner ear. Proc Natl Acad Sci USA 107, 15 792–15 797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayashi T, Cunningham D, Bermingham‐McDonogh O (2007) Loss of Fgfr3 leads to excess hair cell development in the mouse organ of Corti. Dev Dyn 236, 525–533. [DOI] [PubMed] [Google Scholar]
- Hayashi T, Kokubo H, Hartman BH, et al. (2008a) Hesr1 and Hesr2 may act as early effectors of Notch signaling in the developing cochlea. Dev Biol 316, 87–99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayashi T, Ray CA, Bermingham‐McDonogh O (2008b) Fgf20 is required for sensory epithelial specification in the developing cochlea. J Neurosci 28, 5991–5999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heller N, Brandli AW (1999) Xenopus Pax‐2/5/8 orthologues: novel insights into Pax gene evolution and identification of Pax‐8 as the earliest marker for otic and pronephric cell lineages. Dev Genet 24, 208–219. [DOI] [PubMed] [Google Scholar]
- Hudspeth AJ (2008) Making an effort to listen: mechanical amplification in the ear. Neuron 59, 530–545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hudspeth AJ (2014) Integrating the active process of hair cells with cochlear function. Nat Rev Neurosci 15, 600–614. [DOI] [PubMed] [Google Scholar]
- Huh SH, Jones J, Warchol ME, et al. (2012) Differentiation of the lateral compartment of the cochlea requires a temporally restricted FGF20 signal. PLoS Biol 10, e1001231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jacques BE, Montcouquiol ME, Layman EM, et al. (2007) Fgf8 induces pillar cell fate and regulates cellular patterning in the mammalian cochlea. Development 134, 3021–3029. [DOI] [PubMed] [Google Scholar]
- Jacques BE, Puligilla C, Weichert RM, et al. (2012) A dual function for canonical Wnt/beta‐catenin signaling in the developing mammalian cochlea. Development 139, 4395–4404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jacques BE, Montgomery WHT, Uribe PM, et al. (2014) The role of Wnt/beta‐catenin signaling in proliferation and regeneration of the developing basilar papilla and lateral line. Dev Neurobiol, 74, 438–456. [DOI] [PubMed] [Google Scholar]
- Jahan I, Pan N, Kersigo J, et al. (2010) Neurod1 suppresses hair cell differentiation in ear ganglia and regulates hair cell subtype development in the cochlea. PLoS ONE 5, e11661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jarman AP, Groves AK (2013) The role of Atonal transcription factors in the development of mechanosensitive cells. Semin Cell Dev Biol 24, 438–447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jayasena CS, Ohyama T, Segil N, et al. (2008) Notch signaling augments the canonical Wnt pathway to specify the size of the otic placode. Development 135, 2251–2261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaltenbach JA, Falzarano PR, Simpson TH (1994) Postnatal development of the hamster cochlea. II. Growth and differentiation of stereocilia bundles. J Comp Neurol 350, 187–198. [DOI] [PubMed] [Google Scholar]
- Kamaid A, Neves J, Giraldez F (2010) Id gene regulation and function in the prosensory domains of the chicken inner ear: a link between Bmp signaling and Atoh1. J Neurosci 30, 11 426–11 434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kanzaki S, Beyer LA, Swiderski DL, et al. (2006) p27(Kip1) deficiency causes organ of Corti pathology and hearing loss. Hear Res 214, 28–36. [DOI] [PubMed] [Google Scholar]
- Kelberman D, Rizzoti K, Avilion A, et al. (2006) Mutations within Sox2/SOX2 are associated with abnormalities in the hypothalamo‐pituitary‐gonadal axis in mice and humans. J Clin Invest 116, 2442–2455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kelley MW (2007) Cellular commitment and differentiation in the organ of Corti. Int J Dev Biol 51, 571–583. [DOI] [PubMed] [Google Scholar]
- Kelley MW, Xu XM, Wagner MA, et al. (1993) The developing organ of Corti contains retinoic acid and forms supernumerary hair cells in response to exogenous retinoic acid in culture. Development 119, 1041–1053. [DOI] [PubMed] [Google Scholar]
- Kelley MW, Driver EC, Puligilla C (2009) Regulation of cell fate and patterning in the developing mammalian cochlea. Curr Opin Otolaryngol Head Neck Surg 17, 381–387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kelly M, Chen P (2007) Shaping the mammalian auditory sensory organ by the planar cell polarity pathway. Int J Dev Biol 51, 535–547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kelly MC, Chen P (2009) Development of form and function in the mammalian cochlea. Curr Opin Neurobiol 19, 395–401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khatri SB, Groves AK (2013) Expression of the Foxi2 and Foxi3 transcription factors during development of chicken sensory placodes and pharyngeal arches. Gene Expr Patterns 13, 38–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khatri SB, Edlund RK, Groves AK (2014) Foxi3 Is Necessary for the Induction of the Chick Otic Placode in Response to Fgf Signaling. Dev Biol 391, 158–169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kiernan AE (2013) Notch signaling during cell fate determination in the inner ear. Semin Cell Dev Biol 24, 470–479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kiernan AE, Cordes R, Kopan R, et al. (2005a) The Notch ligands DLL1 and JAG2 act synergistically to regulate hair cell development in the mammalian inner ear. Development 132, 4353–4362. [DOI] [PubMed] [Google Scholar]
- Kiernan AE, Pelling AL, Leung KK, et al. (2005b) Sox2 is required for sensory organ development in the mammalian inner ear. Nature 434, 1031–1035. [DOI] [PubMed] [Google Scholar]
- Kiernan AE, Xu J, Gridley T (2006) The Notch ligand JAG1 is required for sensory progenitor development in the mammalian inner ear. PLoS Genet 2, e4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koundakjian EJ, Appler JL, Goodrich LV (2007) Auditory neurons make stereotyped wiring decisions before maturation of their targets. J Neurosci 27, 14 078–14 088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kruger M, Schmid T, Kruger S, et al. (2006) Functional redundancy of NSCL‐1 and NeuroD during development of the petrosal and vestibulocochlear ganglia. Eur J Neurosci 24, 1581–1590. [DOI] [PubMed] [Google Scholar]
- Ladhams A, Pickles JO (1996) Morphology of the monotreme organ of Corti and macula lagena. J Comp Neurol 366, 335–347. [DOI] [PubMed] [Google Scholar]
- Laine H, Doetzlhofer A, Mantela J, et al. (2007) p19(Ink4d) and p21(Cip1) collaborate to maintain the postmitotic state of auditory hair cells, their codeletion leading to DNA damage and p53‐mediated apoptosis. J Neurosci 27, 1434–1444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lanford PJ, Lan Y, Jiang R, et al. (1999) Notch signalling pathway mediates hair cell development in mammalian cochlea. Nat Genet 21, 289–292. [DOI] [PubMed] [Google Scholar]
- Lanford PJ, Shailam R, Norton CR, et al. (2000) Expression of Math1 and HES5 in the cochleae of wildtype and Jag2 mutant mice. J Assoc Res Otolaryngol 1, 161–171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee YS, Liu F, Segil N (2006) A morphogenetic wave of p27Kip1 transcription directs cell cycle exit during organ of Corti development. Development 133, 2817–2826. [DOI] [PubMed] [Google Scholar]
- Legan PK, Lukashkina VA, Goodyear RJ, et al. (2000) A targeted deletion in alpha‐tectorin reveals that the tectorial membrane is required for the gain and timing of cochlear feedback. Neuron 28, 273–285. [DOI] [PubMed] [Google Scholar]
- Lewis J (1991) Rules for the production of sensory cells. Ciba Found Symp, 160, 25–39; discussion 40–53. [PubMed] [Google Scholar]
- Lewis J (1998) Notch signalling and the control of cell fate choices in vertebrates. Semin Cell Dev Biol 9, 583–589. [DOI] [PubMed] [Google Scholar]
- Li CW, Ruben RJ (1979) Further study of the surface morphology of the embryonic mouse cochlear sensory epithelia. Otolaryngol Head Neck Surg 87, 479–485. [DOI] [PubMed] [Google Scholar]
- Li CW, Van De Water TR, Ruben RJ (1978) The fate mapping of the eleventh and twelfth day mouse otocyst: an in vitro study of the sites of origin of the embryonic inner ear sensory structures. J Morphol, 157, 249–267. [DOI] [PubMed] [Google Scholar]
- Li S, Mark S, Radde‐Gallwitz K, et al. (2008) Hey2 functions in parallel with Hes1 and Hes5 for mammalian auditory sensory organ development. BMC Dev Biol 8, 20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liang JK, Bok J, Wu DK (2010) Distinct contributions from the hindbrain and mesenchyme to inner ear morphogenesis. Dev Biol 337, 324–334. [DOI] [PubMed] [Google Scholar]
- Liem KF Jr, Tremml G, Roelink H, et al. (1995) Dorsal differentiation of neural plate cells induced by BMP‐mediated signals from epidermal ectoderm. Cell 82, 969–979. [DOI] [PubMed] [Google Scholar]
- Liem KF Jr, Jessell TM, Briscoe J (2000) Regulation of the neural patterning activity of sonic hedgehog by secreted BMP inhibitors expressed by notochord and somites. Development 127, 4855–4866. [DOI] [PubMed] [Google Scholar]
- Lim DJ (1986) Functional structure of the organ of Corti: a review. Hear Res 22, 117–146. [DOI] [PubMed] [Google Scholar]
- Lim DJ, Anniko M (1985) Developmental morphology of the mouse inner ear. A scanning electron microscopic observation. Acta Otolaryngol Suppl 422, 1–69. [PubMed] [Google Scholar]
- Lin Z, Cantos R, Patente M, et al. (2005) Gbx2 is required for the morphogenesis of the mouse inner ear: a downstream candidate of hindbrain signaling. Development 132, 2309–2318. [DOI] [PubMed] [Google Scholar]
- Litsiou A, Hanson S, Streit A (2005) A balance of FGF, BMP and WNT signalling positions the future placode territory in the head. Development 132, 4051–4062. [DOI] [PubMed] [Google Scholar]
- Liu M, Pereira FA, Price SD, et al. (2000) Essential role of BETA2/NeuroD1 in development of the vestibular and auditory systems. Genes Dev 14, 2839–2854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu Z, Owen T, Zhang L, et al. (2010) Dynamic expression pattern of Sonic hedgehog in developing cochlear spiral ganglion neurons. Dev Dyn 239, 1674–1683. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu K, Lin B, Zhao M, et al. (2013) The multiple roles for Sox2 in stem cell maintenance and tumorigenesis. Cell Signal 25, 1264–1271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lowenheim H, Furness DN, Kil J, et al. (1999) Gene disruption of p27(Kip1) allows cell proliferation in the postnatal and adult organ of corti. Proc Natl Acad Sci USA 96, 4084–4088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo G, Hofmann C, Bronckers AL, et al. (1995) BMP‐7 is an inducer of nephrogenesis, and is also required for eye development and skeletal patterning. Genes Dev 9, 2808–2820. [DOI] [PubMed] [Google Scholar]
- Ma Q, Chen Z, del Barco Barrantes I, et al. (1998) Neurogenin1 is essential for the determination of neuronal precursors for proximal cranial sensory ganglia. Neuron 20, 469–482. [DOI] [PubMed] [Google Scholar]
- Ma Q, Anderson DJ, Fritzsch B (2000) Neurogenin 1 null mutant ears develop fewer, morphologically normal hair cells in smaller sensory epithelia devoid of innervation. J Assoc Res Otolaryngol 1, 129–143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maier EC, Whitfield TT (2014) RA and FGF signalling are required in the zebrafish otic vesicle to pattern and maintain ventral otic identities. PLoS Genet 10, e1004858. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manley GA (2004) The lizard basilar papilla and its evolution In: Evolution of the Vertebrate Auditory System . (eds Manley GA, Popper AN, Fay RR.), pp. 200–223. New York, NY: Springer. [Google Scholar]
- Manley GA (2012) Evolutionary paths to mammalian cochleae. J Assoc Res Otolaryngol 13, 733–743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mann ZF, Kelley MW (2011) Development of tonotopy in the auditory periphery. Hear Res 276, 2–15. [DOI] [PubMed] [Google Scholar]
- Mann ZF, Thiede BR, Chang W, et al. (2014) A gradient of Bmp7 specifies the tonotopic axis in the developing inner ear. Nat Commun 5, 3839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manor U, Kachar B (2008) Dynamic length regulation of sensory stereocilia. Semin Cell Dev Biol 19, 502–510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mansour SL, Li C, Urness LD (2013) Genetic rescue of Muenke syndrome model hearing loss reveals prolonged FGF‐dependent plasticity in cochlear supporting cell fates. Genes Dev 27, 2320–2331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mark M, Lufkin T, Vonesch JL, et al. (1993) Two rhombomeres are altered in Hoxa‐1 mutant mice. Development 119, 319–338. [DOI] [PubMed] [Google Scholar]
- Matei V, Pauley S, Kaing S, et al. (2005) Smaller inner ear sensory epithelia in Neurog 1 null mice are related to earlier hair cell cycle exit. Dev Dyn 234, 633–650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McKay IJ, Lewis J, Lumsden A (1996) The role of FGF‐3 in early inner ear development: an analysis in normal and kreisler mutant mice. Dev Biol 174, 370–378. [DOI] [PubMed] [Google Scholar]
- Montcouquiol M, Kelley MW (2003) Planar and vertical signals control cellular differentiation and patterning in the mammalian cochlea. J Neurosci 23, 9469–9478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morsli H, Choo D, Ryan A, et al. (1998) Development of the mouse inner ear and origin of its sensory organs. J Neurosci 18, 3327–3335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morsli H, Tuorto F, Choo D, et al. (1999) Otx1 and Otx2 activities are required for the normal development of the mouse inner ear. Development 126, 2335–2343. [DOI] [PubMed] [Google Scholar]
- Mueller KL, Jacques BE, Kelley MW (2002) Fibroblast growth factor signaling regulates pillar cell development in the organ of corti. J Neurosci 22, 9368–9377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mulvaney J, Dabdoub A (2012) Atoh1, an essential transcription factor in neurogenesis and intestinal and inner ear development: function, regulation, and context dependency. J Assoc Res Otolaryngol 13, 281–293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Munnamalai V, Hayashi T, Bermingham‐McDonogh O (2012) Notch prosensory effects in the Mammalian cochlea are partially mediated by Fgf20. J Neurosci 32, 12 876–12 884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murata J, Ohtsuka T, Tokunaga A, et al. (2009) Notch‐Hes1 pathway contributes to the cochlear prosensory formation potentially through the transcriptional down‐regulation of p27Kip1. J Neurosci Res 87, 3521–3534. [DOI] [PubMed] [Google Scholar]
- Nayak GD, Ratnayaka HS, Goodyear RJ, et al. (2007) Development of the hair bundle and mechanotransduction. Int J Dev Biol 51, 597–608. [DOI] [PubMed] [Google Scholar]
- Neves J, Parada C, Chamizo M, et al. (2011) Jagged 1 regulates the restriction of Sox2 expression in the developing chicken inner ear: a mechanism for sensory organ specification. Development 138, 735–744. [DOI] [PubMed] [Google Scholar]
- Nissen RM, Yan J, Amsterdam A, et al. (2003) Zebrafish foxi one modulates cellular responses to Fgf signaling required for the integrity of ear and jaw patterning. Development 130, 2543–2554. [DOI] [PubMed] [Google Scholar]
- Oh SH, Johnson R, Wu DK (1996) Differential expression of bone morphogenetic proteins in the developing vestibular and auditory sensory organs. J Neurosci 16, 6463–6475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohyama T, Mohamed OA, Taketo MM, et al. (2006) Wnt signals mediate a fate decision between otic placode and epidermis. Development 133, 865–875. [DOI] [PubMed] [Google Scholar]
- Ohyama T, Groves AK, Martin K (2007) The first steps towards hearing: mechanisms of otic placode induction. Int J Dev Biol 51, 463–472. [DOI] [PubMed] [Google Scholar]
- Ohyama T, Basch ML, Mishina Y, et al. (2010) BMP signaling is necessary for patterning the sensory and nonsensory regions of the developing mammalian cochlea. J Neurosci 30, 15 044–15 051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ozaki H, Nakamura K, Funahashi J, et al. (2004) Six1 controls patterning of the mouse otic vesicle. Development 131, 551–562. [DOI] [PubMed] [Google Scholar]
- Pack AK, Slepecky NB (1995) Cytoskeletal and calcium‐binding proteins in the mammalian organ of Corti: cell type‐specific proteins displaying longitudinal and radial gradients. Hear Res 91, 119–135. [DOI] [PubMed] [Google Scholar]
- Pan W, Jin Y, Stanger B, et al. (2010) Notch signaling is required for the generation of hair cells and supporting cells in the mammalian inner ear. Proc Natl Acad Sci USA 107, 15 798–15 803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pan N, Jahan I, Kersigo J, et al. (2011) Conditional deletion of Atoh1 using Pax2‐Cre results in viable mice without differentiated cochlear hair cells that have lost most of the organ of Corti. Hear Res 275, 66–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pan W, Jin Y, Chen J, et al. (2013) Ectopic expression of activated notch or SOX2 reveals similar and unique roles in the development of the sensory cell progenitors in the mammalian inner ear. J Neurosci 33, 16 146–16 157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pauley S, Wright TJ, Pirvola U, et al. (2003) Expression and function of FGF10 in mammalian inner ear development. Dev Dyn 227, 203–215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pauley S, Lai E, Fritzsch B (2006) Foxg1 is required for morphogenesis and histogenesis of the mammalian inner ear. Dev Dyn 235, 2470–2482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peng AW, Belyantseva IA, Hsu PD, et al. (2009) Twinfilin 2 regulates actin filament lengths in cochlear stereocilia. J Neurosci 29, 15 083–15 088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pfeffer PL, Gerster T, Lun K, et al. (1998) Characterization of three novel members of the zebrafish Pax2/5/8 family: dependency of Pax5 and Pax8 expression on the Pax2.1 (noi) function. Development 125, 3063–3074. [DOI] [PubMed] [Google Scholar]
- Pfister M, Thiele H, Van Camp G, et al. (2004) A genotype–phenotype correlation with gender‐effect for hearing impairment caused by TECTA mutations. Cell Physiol Biochem 14, 369–376. [DOI] [PubMed] [Google Scholar]
- Pirvola U, Spencer‐Dene B, Xing‐Qun L, et al. (2000) FGF/FGFR‐2(IIIb) signaling is essential for inner ear morphogenesis. J Neurosci 20, 6125–6134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plantinga RF, de Brouwer AP, Huygen PL, et al. (2006) A novel TECTA mutation in a Dutch DFNA8/12 family confirms genotype–phenotype correlation. J Assoc Res Otolaryngol 7, 173–181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plouhinec JL, Zakin L, De Robertis EM (2011) Systems control of BMP morphogen flow in vertebrate embryos. Curr Opin Genet Dev 21, 696–703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Popper AN, Fay RR (1999) The auditory periphery in fishes In: Comparative Hearing: Fish and Amphibians . (eds Popper AN, Fay RR.), pp. 43–100. New York: Springer. [Google Scholar]
- Puligilla C, Feng F, Ishikawa K, et al. (2007) Disruption of fibroblast growth factor receptor 3 signaling results in defects in cellular differentiation, neuronal patterning, and hearing impairment. Dev Dyn 236, 1905–1917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Puligilla C, Dabdoub A, Brenowitz SD, et al. (2010) Sox2 induces neuronal formation in the developing mammalian cochlea. J Neurosci 30, 714–722. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Radosevic M, Robert‐Moreno A, Coolen M, et al. (2011) Her9 represses neurogenic fate downstream of Tbx1 and retinoic acid signaling in the inner ear. Development 138, 397–408. [DOI] [PubMed] [Google Scholar]
- Raft S, Groves AK (2015) Segregating neural and mechanosensory fates in the developing ear: patterning, signaling, and transcriptional control. Cell Tissue Res 359, 315–332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raft S, Nowotschin S, Liao J, et al. (2004) Suppression of neural fate and control of inner ear morphogenesis by Tbx1. Development 131, 1801–1812. [DOI] [PubMed] [Google Scholar]
- Raft S, Koundakjian EJ, Quinones H, et al. (2007) Cross‐regulation of Ngn1 and Math1 coordinates the production of neurons and sensory hair cells during inner ear development. Development 134, 4405–4415. [DOI] [PubMed] [Google Scholar]
- Raz Y, Kelley MW (1999) Retinoic acid signaling is necessary for the development of the organ of Corti. Dev Biol 213, 180–193. [DOI] [PubMed] [Google Scholar]
- Riccomagno MM, Martinu L, Mulheisen M, et al. (2002) Specification of the mammalian cochlea is dependent on Sonic hedgehog. Genes Dev 16, 2365–2378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Riccomagno MM, Takada S, Epstein DJ (2005) Wnt‐dependent regulation of inner ear morphogenesis is balanced by the opposing and supporting roles of Shh. Genes Dev 19, 1612–1623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richter CP, Emadi G, Getnick G, et al. (2007) Tectorial membrane stiffness gradients. Biophys J 93, 2265–2276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rocha‐Sanchez SM, Beisel KW (2007) Pocket proteins and cell cycle regulation in inner ear development. Int J Dev Biol 51, 585–595. [DOI] [PubMed] [Google Scholar]
- Rocha‐Sanchez SM, Scheetz LR, Contreras M, et al. (2011) Mature mice lacking Rbl2/p130 gene have supernumerary inner ear hair cells and supporting cells. J Neurosci 31, 8883–8893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruben RJ (1967) Development of the inner ear of the mouse: a radioautographic study of terminal mitoses. Acta Otolaryngol Suppl 220, 1–44. [PubMed] [Google Scholar]
- Russell IJ, Legan PK, Lukashkina VA, et al. (2007) Sharpened cochlear tuning in a mouse with a genetically modified tectorial membrane. Nat Neurosci 10, 215–223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sage C, Huang M, Karimi K, et al. (2005) Proliferation of functional hair cells in vivo in the absence of the retinoblastoma protein. Science 307, 1114–1118. [DOI] [PubMed] [Google Scholar]
- Sage C, Huang M, Vollrath MA, et al. (2006) Essential role of retinoblastoma protein in mammalian hair cell development and hearing. Proc Natl Acad Sci USA 103, 7345–7350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schimmang T, Pirvola U (2013) Coupling the cell cycle to development and regeneration of the inner ear. Semin Cell Dev Biol 24, 507–513. [DOI] [PubMed] [Google Scholar]
- Schraders M, Ruiz‐Palmero L, Kalay E, et al. (2012) Mutations of the gene encoding otogelin are a cause of autosomal‐recessive nonsyndromic moderate hearing impairment. Am J Hum Genet 91, 883–889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwander M, Kachar B, Muller U (2010) Review series: the cell biology of hearing. J Cell Biol 190, 9–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sher AE (1971) The embryonic and postnatal development of the inner ear of the mouse. Acta Otolaryngol Suppl 285, 1–77. [PubMed] [Google Scholar]
- Shi F, Cheng YF, Wang XL, et al. (2010) Beta‐catenin up‐regulates Atoh1 expression in neural progenitor cells by interaction with an Atoh1 3′ enhancer. J Biol Chem 285, 392–400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shim K, Minowada G, Coling DE, et al. (2005) Sprouty2, a mouse deafness gene, regulates cell fate decisions in the auditory sensory epithelium by antagonizing FGF signaling. Dev Cell 8, 553–564. [DOI] [PubMed] [Google Scholar]
- Simmler MC, Zwaenepoel II, Verpy E, et al. (2000) Twister mutant mice are defective for otogelin, a component specific to inner ear acellular membranes. Mamm Genome 11, 961–966. [DOI] [PubMed] [Google Scholar]
- Smotherman M, Narins P (2004) Evolution of the amphibian ear In: Evolution of the Vertebrate Auditory System . (eds Manley GA, Popper AN, Fay RR.), pp. 164–199. New York: Springer. [Google Scholar]
- Solomon KS, Kudoh T, Dawid IB, et al. (2003) Zebrafish foxi1 mediates otic placode formation and jaw development. Development 130, 929–940. [DOI] [PubMed] [Google Scholar]
- Son EJ, Wu L, Yoon H, et al. (2012) Developmental gene expression profiling along the tonotopic axis of the mouse cochlea. PLoS ONE 7, e40735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Streit A (2007) The preplacodal region: an ectodermal domain with multipotential progenitors that contribute to sense organs and cranial sensory ganglia. Int J Dev Biol 51, 447–461. [DOI] [PubMed] [Google Scholar]
- Tateya T, Imayoshi I, Tateya I, et al. (2011) Cooperative functions of Hes/Hey genes in auditory hair cell and supporting cell development. Dev Biol 352, 329–340. [DOI] [PubMed] [Google Scholar]
- Tateya T, Imayoshi I, Tateya I, et al. (2013) Hedgehog signaling regulates prosensory cell properties during the basal‐to‐apical wave of hair cell differentiation in the mammalian cochlea. Development 140, 3848–3857. [DOI] [PubMed] [Google Scholar]
- Thiede BR, Mann ZF, Chang W, et al. (2014) Retinoic acid signalling regulates the development of tonotopically patterned hair cells in the chicken cochlea. Nat Commun 5, 3840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tilney LG, Saunders JC (1983) Actin filaments, stereocilia, and hair cells of the bird cochlea. I. Length, number, width, and distribution of stereocilia of each hair cell are related to the position of the hair cell on the cochlea. J Cell Biol 96, 807–821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsuchiya K, Nakamura T, Okamoto R, et al. (2007) Reciprocal targeting of Hath1 and beta‐catenin by Wnt glycogen synthase kinase 3beta in human colon cancer. Gastroenterology 132, 208–220. [DOI] [PubMed] [Google Scholar]
- Tsuprun V, Santi P (1999) Ultrastructure and immunohistochemical identification of the extracellular matrix of the chinchilla cochlea. Hear Res 129, 35–49. [DOI] [PubMed] [Google Scholar]
- Udan RS, Piazza VG, Hsu CW, et al. (2014) Quantitative imaging of cell dynamics in mouse embryos using light‐sheet microscopy. Development 141, 4406–4414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vater M, Meng J, Fox RC (2004) Hearing organ evolution and specialization: early and later mammals In: Evolution of the Vertebrate Auditory System . (eds Manley GA, Popper AN, Fay RR.), pp. 256–288. New York: Springer. [Google Scholar]
- Vitelli F, Morishima M, Taddei I, et al. (2002) Tbx1 mutation causes multiple cardiovascular defects and disrupts neural crest and cranial nerve migratory pathways. Hum Mol Genet 11, 915–922. [DOI] [PubMed] [Google Scholar]
- Vogt N (2015) Transparency in large tissue samples. Nat Methods 12, 11. [DOI] [PubMed] [Google Scholar]
- Waldman EH, Castillo A, Collazo A (2007) Ablation studies on the developing inner ear reveal a propensity for mirror duplications. Dev Dyn 236, 1237–1248. [DOI] [PubMed] [Google Scholar]
- Weber T, Corbett MK, Chow LM, et al. (2008) Rapid cell‐cycle reentry and cell death after acute inactivation of the retinoblastoma gene product in postnatal cochlear hair cells. Proc Natl Acad Sci USA 105, 781–785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whitfield TT, Riley BB, Chiang MY, et al. (2002) Development of the zebrafish inner ear. Dev Dyn 223, 427–458. [DOI] [PubMed] [Google Scholar]
- Woods C, Montcouquiol M, Kelley MW (2004) Math1 regulates development of the sensory epithelium in the mammalian cochlea. Nat Neurosci 7, 1310–1318. [DOI] [PubMed] [Google Scholar]
- Wright A (1984) Dimensions of the cochlear stereocilia in man and the guinea pig. Hear Res 13, 89–98. [DOI] [PubMed] [Google Scholar]
- Wright TJ, Mansour SL (2003) Fgf3 and Fgf10 are required for mouse otic placode induction. Development 130, 3379–3390. [DOI] [PubMed] [Google Scholar]
- Wu DK, Kelley MW (2012) Molecular mechanisms of inner ear development. Cold Spring Harb Perspect Biol 4, a008409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu DK, Oh SH (1996) Sensory organ generation in the chick inner ear. J Neurosci 16, 6454–6462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu DK, Nunes FD, Choo D (1998) Axial Specification for Sensory Organs Versus Non‐Sensory Structures of the Chicken Inner Ear. Development 125, 11–20. [DOI] [PubMed] [Google Scholar]
- Xia A, Gao SS, Yuan T, et al. (2010) Deficient forward transduction and enhanced reverse transduction in the alpha tectorin C1509G human hearing loss mutation. Dis Model Mech 3, 209–223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamamoto N, Chang W, Kelley MW (2011) Rbpj regulates development of prosensory cells in the mammalian inner ear. Dev Biol 353, 367–379. [DOI] [PubMed] [Google Scholar]
- Yang H, Xie X, Deng M, et al. (2010) Generation and characterization of Atoh1‐Cre knock‐in mouse line. Genesis 48, 407–413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yariz KO, Duman D, Seco CZ, et al. (2012) Mutations in OTOGL, encoding the inner ear protein otogelin‐like, cause moderate sensorineural hearing loss. Am J Hum Genet 91, 872–882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu Y, Weber T, Yamashita T, et al. (2010) In vivo proliferation of postmitotic cochlear supporting cells by acute ablation of the retinoblastoma protein in neonatal mice. J Neurosci 30, 5927–5936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao H, Ayrault O, Zindy F, et al. (2008) Post‐transcriptional down‐regulation of Atoh1/Math1 by bone morphogenic proteins suppresses medulloblastoma development. Genes Dev 22, 722–727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zheng JL, Gao WQ (2000) Overexpression of Math1 induces robust production of extra hair cells in postnatal rat inner ears. Nat Neurosci 3, 580–586. [DOI] [PubMed] [Google Scholar]
- Zheng JL, Shou J, Guillemot F, et al. (2000) Hes1 is a negative regulator of inner ear hair cell differentiation. Development 127, 4551–4560. [DOI] [PubMed] [Google Scholar]
- Zheng W, Huang L, Wei ZB, et al. (2003) The role of Six1 in mammalian auditory system development. Development 130, 3989–4000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zine A, Aubert A, Qiu J, et al. (2001) Hes1 and Hes5 activities are required for the normal development of the hair cells in the mammalian inner ear. J Neurosci 21, 4712–4720. [DOI] [PMC free article] [PubMed] [Google Scholar]