

Abstract
The inner ear of mammals consists of the cochlea, which is involved with the sense of hearing, and the vestibule and three semicircular canals, which are involved with the sense of balance. Although different regions of the inner ear contribute to different functions, the bony chambers and membranous ducts are morphologically continuous. The gross anatomy of the cochlea that has been related to auditory physiologies includes overall size of the structure, including volume and total spiral length, development of internal cochlear structures, including the primary and secondary bony laminae, morphology of the spiral nerve ganglion, and the nature of cochlear coiling, including total number of turns completed by the cochlear canal and the relative diameters of the basal and apical turns. The overall sizes, shapes, and orientations of the semicircular canals are related to sensitivity to head rotations and possibly locomotor behaviors. Intraspecific variation, primarily in the shape and orientation of the semicircular canals, may provide additional clues to help us better understand form and function of the inner ear.
Keywords: cochlea, inner ear, semicircular canals, vestibule
Introduction
The otic (ear) region is part of the special sensory division of the nervous system of vertebrates. The function of the ear is two‐fold – hearing within the cochlea and balance within the vestibule and semicircular canals. The organs of hearing and balance within the inner ear are minute structures, which are contained within cavities ranging in volume from < 1 mm3 in shrews to over 1000 mm3 in baleen whales (Ekdale, 2013). The inner ear of humans occupies a space of a little over 150 mm3. Although the inner ear organs are small in size, they are quite powerful physiologically. It is amazing that such miniscule structures can cause a myriad of problems from tinnitus to motion sickness to a general lack of balance. This phenomenon, in part, has caused the ear region to be one of the most intensively studied systems of vertebrate anatomy and physiology.
The sense of hearing was the first inner ear function to be recognized by human anatomists and physiologists. The role of the inner ear in hearing was deduced in ancient Egypt from the effects of injuries to the temporal region of the head, but the specific role that the cochlea plays was not determined until the middle of the 17th century (Hawkins, 2004). Sensations of orientation were attributed to the semicircular canals and vestibular system in the late 19th century (Dercum, 1879), and functional similarities of the semicircular canal and lateral line systems in fish and amphibians were recognized around the same time (Lee, 1898). Further advancements in the comparative anatomy of the vertebrate inner ear were made in the middle of the 19th and 20th centuries, through dissections and histological sectioning (Retzius, 1884), corrosion casting of the bony chambers of the inner ear (Hyrtl, 1845), and extraction of the intact membranous inner ear structures as a whole from the surrounding bone (Gray, 1903, 1905, 1907).
From a functional standpoint alone, the auditory and vestibular structures are significant agents in vertebrate biology. Hearing certainly played a role in early tetrapod evolution when newly terrestrial animals moved from detecting water‐borne to air‐borne sounds (Manley, 1972; Clack, 2002). In the case of early mammals, which likely were nocturnal animals (Menaker et al. 1997; Kielan‐Jaworowska et al. 2004; Hall et al. 2012; Gerkema et al. 2013), a reliance on non‐visual senses to navigate the Mesozoic landscape would have been necessary. The hearing capabilities of mammals vary, and mammals can hear across a greater frequency bandwidth than other vertebrates, primarily at high pitches (Echteler et al. 1994; Heffner & Heffner, 2007). For example, elephants are sensitive to very low frequency sound vibrations (14–24 Hz; Payne et al. 1986; Poole et al. 1988) and whales as a group can hear in both infrasonic (below 20 Hz) and ultrasonic (above 20 kHz, up to 180 kHz in some species) ranges (Hall & Johnson, 1972; Ridgway et al. 1981; Ketten & Wartzok, 1990; Ketten, 1997, 2000; Houser et al. 2001; Parks et al. 2007). Microchiropteran bats use ultrasonic echolocation during prey detection and capture (Simmons et al. 1979), and tenrecs and some shrews are known to vocalize in ultrasonic ranges (Gould, 1965; Tomasi, 1979), across which ranges they likely can hear. In contrast, most bony and cartilaginous fishes are only sensitive to sounds below 1 kHz (Fay, 1988; Myrberg, 2001; Casper et al. 2003; Casper & Mann, 2006; Ramcharitar et al. 2006), and upper frequency limits are between 3 and 4 kHz in amphibians (Megela‐Simmons et al. 1985; Fay, 1988) and 1–14 kHz in reptiles, including birds (Christensen‐Dalsgaard & Manley, 2005, 2008; Manley & Kraus, 2010).
Balance was also an important sense in vertebrate history. When the earliest tetrapods left an aquatic environment for land, they faced an array of challenges (Alexander, 2002). Examination of the postcranial skeletons of the earliest tetrapods reveals adaptations to new terrestrial lifestyles (Shubin et al. 2004, 2014; Boisvert, 2005). Balance itself is integral for vertebrate locomotion, and a broad range of mobility is observed within Mammalia (see Spoor et al. 2007). In turn, the anatomy of inner ear compartments correlates to various locomotor behaviors of mammals (e.g. Spoor et al. 1996, 2007; Malinzak et al. 2012).
The morphology of the inner ear is informative for phylogenetic studies at both more‐ and less‐inclusive taxonomic levels. For example, the cochlea completes at least one complete 360° turn in living therian mammals (marsupials and placentals), but less in monotremes and more basally positioned taxa (Gray, 1907, 1908; Rowe, 1988; Ruf et al. 2009, 2013; Luo et al. 2011, 2012). The bony labyrinths of marsupial and placental mammals from the Mesozoic era exhibit ancestral morphologies, such as fusion of the posterior and lateral semicircular canals to form a secondary common crus. The secondary common crus is lost in several clades within crown Theria, including primates, rodents, some carnivorous mammals, and a variety of marsupial clades, and the posterior and lateral canals are separate for their entire lengths in most extant mammals (Meng & Fox, 1995; Schmelzle et al. 2007; Ekdale & Rowe, 2011). Dimensions of the inner ear within Primates differ between great apes and other primates (Spoor & Zonneveld, 1998), as well as between humans and subspecies of chimps (Gunz et al. 2012). Further phylogenetic information can be found within the inner ears of squamate reptiles (Shute & Bellairs, 1953; Schmidt, 1964; Miller, 1966a,b, 1968; Maddin & Sherratt, 2014).
Anatomy and function of the inner ear
The generalized ear of mammals is partitioned into the outer, middle, and inner ears. The outer ear includes the pinna, which funnels sound from the environment into the ear region of the head, and extends from the external surface of the head to the tympanic membrane, or eardrum, via the external acoustic meatus (Fig. 1). The middle ear extends from the tympanic membrane to the lateral surface of the skull and contains the three ear ossicles (malleus, incus, and stapes) within the tympanic cavity of the middle ear. The tympanic cavity itself is enclosed ventrally by the tympanic (auditory) bulla, which forms an often‐bulbous, bony or cartilaginous structure in most mammal taxa. The composition of the tympanic bulla varies among mammal species in the specific bony element or elements that contribute to the structure (van der Klaauw, 1930, 1931; Novacek, 1977, 1993). The middle ear ossicles form a chain connecting the tympanic membrane to the petrosal bone, which contributes to the posterolateral region of the basicranium and surrounds the inner ear cavities. The function of the ossicular chain within the middle ear is to transmit air‐borne sound waves from the tympanic membrane to the fluid filled chambers of the inner ear. The stapes articulates with an opening in the petrosal known as the fenestra vestibuli (oval window) and acts as one of two major areas of communication between the middle and inner ear cavities (Fig. 1). The other opening, which typically is found posteromedial to the fenestra vestibuli, is the fenestra cochleae (round window). The fenestra cochleae is covered by a secondary tympanic membrane that accommodates expansion of the inner ear space during stapedial vibration.
Figure 1.
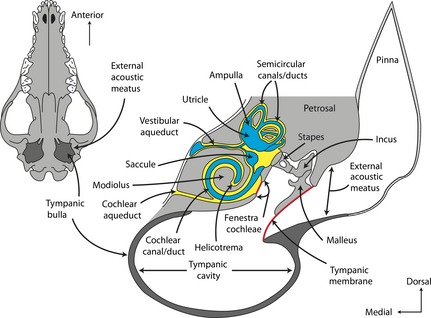
Cross‐section through the head of a domestic dog (Canis familiaris) showing structures of outer, middle, and inner ears (modified from Evans, 1993). Blue indicates structures of the membranous labyrinth, yellow indicates structures of the bony labyrinth, and red indicates the primary (eardrum) and secondary (spanning fenestra cochleae) tympanic membranes.
The inner ear consists of a set of interconnected spaces within the petrosal bone of mammals known as the bony labyrinth (yellow chambers in Fig. 1). Within the bony labyrinth is a series of interconnected soft tissue sacs and ducts known as the membranous labyrinth (blue chambers in Fig. 1). Traditional parlance of the ear region reserves the term ‘canal’ for structures of the bony labyrinth, while the term ‘duct’ refers to the membranous channels. The membranous labyrinth is separated into an inferior division that includes the cochlear duct and the saccule of the membranous vestibule, and a superior division that includes the utricle of the vestibule and the three semicircular ducts and associated ampullae. The organ of Corti (spiral organ of hearing) is housed within the cochlear duct, and receptors that are sensitive to linear motion are found within the saccule. The utricle and semicircular ducts are involved with the sensation of rotational movements of the head. The osseous semicircular canals and cochlea of the bony labyrinth mirror the shape of the membranous ducts within, although the bony canals may not accurately reflect the size of the ducts (Curthoys et al. 1977). Nonetheless, the bony labyrinth often is used to study the form and function of the inner ear in the absence of the membranous ducts (e.g. Fleischer, 1976; Geisler & Luo, 1996).
Lymphatic fluid both fills and surrounds the membranous labyrinth. The spaces surrounding the membranous labyrinth within the bony labyrinth are filled with perilymph. The composition of perilymph is similar to extracellular fluid and is rich in sodium but poor in potassium (Sterkers et al. 1988; Echteler et al. 1994). Perilymph originates from the lymphatic system and exits the inner ear via the membranous perilymphatic duct within the bony cochlear aqueduct into the subarachnoid space surrounding the brain (Fig. 1). The cochlear duct and membranous vestibular apparatus are filled with endolymph. In contrast to perilymph, endolymph is rich in potassium ions but poor in sodium ions, with a pH similar to blood plasma, and it has a composition similar to intracellular fluid (Bosher & Warren, 1968; Sterkers et al. 1988; Payan et al. 1997). Endolymph is produced by specialized cells within highly vascularized epithelium along the lateral wall of the cochlear duct called the stria vascularis (see Echteler et al. 1994), which maintains the ionic composition of the endocochlear endolymph as well as its unusual electrical potential (Tasaki & Spyropoulos, 1959; Carlisle et al. 1990; Hibino et al. 2010). The endolymph is resorbed by the endolymphatic sac, which extends from the vestibule via a bony aqueduct and sits within the subdural space around the cerebellum (Fig. 1). There is an apparent heterogeneity of compositions in both the endolymph and perilymph in different regions of the inner ear on account of the irregular shapes of the membranous structures (Sterkers et al. 1988).
Vibrations within the endolymph, either through stapedial vibrations or movements of the head, stimulate auditory receptors in the cochlea and motion receptors in the vestibular system. Although the sensory functions of the cochlea (hearing) and vestibule (balance) differ, endolymph from one region can flow into the other, thereby affecting the function of the other. For example, changes in the pressure and volume of the vestibular endolymph affect both vestibular and auditory functions. An increase in endolymph volume (known as endolymphatic hydrops) has been connected to Ménière's disease, which manifests as a combination of hearing loss and vertigo (Havia et al. 2002; Merchant et al. 2005). Semicircular canal dehiscence, which is a perforation of the bone overlying the semicircular canals, reduces pressure in the vestibule and affects both the hearing and balance of the afflicted patient (Minor et al. 1998, 2001). Interestingly, evidence suggests that the saccule of the vestibule may hold some low frequency acoustic sensitivity in mammals (Todd et al. 2000; Todd, 2001; Jones et al. 2010) and acoustic waves can evoke a vestibular response (Todd & Cody, 2000; Todd et al. 2003). The functional overlap between the systems might be a physiological retention from the vestibular origin of the cochlea during mammal evolution (Todd, 2001). In fact, the saccule mediates hearing in several non‐mammalian vertebrates, including some fish (Furukawa & Ishii, 1967; Saidel & Popper, 1983), frogs (Wever, 1973; Moffat & Capranica, 1976), and potentially turtles (Wever & Vernon, 1956).
Cochlea
The bony cochlea is coiled around a central axis known as the modiolus in all living mammals except for monotremes, in which the canal is bent near its end but does not complete a full turn (Alexander, 1904; Kermack & Mussett, 1983; Jørgenson & Locket, 1995). The center of the modiolus is hollow and transmits the cochlear branch of cranial nerve VIII. The tip of the cochlear spiral is known as the apex, and the basal end originates near the fenestra vestibuli for the stapedial footplate (Fig. 1). The membranous structures within the bony cochlear canal is divided into two compartments or scalae, which are the scala tympani that communicate with the fenestra cochleae and the scala vestibuli that terminates at the fenestra vestibuli. There is no osseous division between the two scalae in monotremes (Alexander, 1904; Zeller, 1989; Luo et al. 2011). However, the two scalae are partially separated in extant therians by a pair of spiral ridges on opposing walls of the cochlear canal – the primary and secondary bony laminae. The primary bony lamina extends for most of the length of the cochlea along the axial (inner) wall, around the modiolus of the cochlea, and it accommodates the spiral ganglion of the auditory nerve (Fig. 2). A secondary bony lamina often extends for a short distance along the radial (outer) wall of the cochlear canal opposite the primary bony lamina, but the two laminae never come into contact. Both bony laminae and the spiral ganglion canal first appear in the fossil record in dryolestids (stem‐therian mammals) and coincide with an increase in nervous structures within the inner ear (Luo et al. 2011, 2012). The evolution of those structures likely indicates the proliferation of auditory profiles in living mammals.
Figure 2.
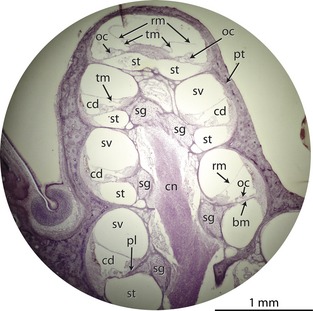
Cross‐section through the cochlea of a guinea pig (Cavia porcellus; Ekdale, personal collection) with internal structures labeled. bm, basilar membrane; cd, cochlear duct (scala media); cn, cochlear branch of cranial nerve VIII within modiolus; oc, organ of Corti; pl, primary bony lamina; pt, petrosal bone (surrounding cochlea); rm, Reissner's membrane; sg, spiral ganglion canal within root of primary bony lamina; st, scala tympani; sv, scala vestibuli; tm, tectorial membrane.
The two bony laminae serve as anchors for a portion of the membranous labyrinth known as the basilar membrane (Fig. 2). The basilar membrane, upon which the organ of Corti sits, defines the tympanal wall of the membranous cochlear duct (often referred to as the scala media). A second soft tissue film known as the vestibular membrane (also known as Reissner's membrane) extends across the width of the cochlear canal to define the vestibular wall of the cochlear duct. The cochlear duct separates the scalae tympani and vestibuli for most of the length of the cochlear spiral, although the two outer scalae communicate at the apex via a small opening known as the helicotrema (Fig. 1).
In general, the basilar membrane is narrow at its base and widens towards the apex (Wever et al. 1971; Webster & Webster, 1980). The stiffness and thickness of the membrane decrease longitudinally from the base to the apex (Naidu & Mountain, 2007), and the membrane is much stiffer radially than longitudinally (Echteler et al. 1994). Vibrations of the stapes within the fenestra vestibuli are transmitted to the basilar membrane via the endolymphatic and perilymphatic fluids, and are propagated across the basilar membrane from the base to the apex. The waves reach a maximum at specific regions of the cochlea according to the frequency of the vibration (Zhang et al. 2007).
The spiral organ of Corti sits upon the vestibular surface of the basilar membrane within the cochlear duct (Fig. 2). Within the organ of Corti, auditory sensations are transduced by cochlear hair cells. A row of flask‐shaped inner hair cells extends longitudinally along the axial border of the basilar membrane. Columnar cells known as outer hair cells occupy a more radial position within the organ of Corti. The outer hair cells contribute over three‐quarters of the sensory cells within the cochlea (Echteler et al. 1994). The inner and outer hair cells differ in morphology and position, but they also have functional differences. Whereas potentials of the inner hair cells are positively correlated with the velocity of basilar membrane vibration, the potentials of the outer hair cells are correlated with the displacement of the basilar membrane (Dallos et al. 1972; Fettiplace & Hackney, 2006). Between the outer and inner hair cells are a series of supporting cells that include inner and outer pillar cells. The pillar cells form the walls of the tunnel of Corti.
The outer and inner hair cells are innervated by branches of cranial nerve VIII, which exits the cranial cavity via the internal acoustic meatus on the endocranial surface of the petrosal. A vestibular branch extends posteriorly to innervate the vestibular end organs, and a cochlear branch descends into the modiolus of the cochlea. The walls of the modiolus are cribriform and form a sieve‐like bony floor for numerous passages of minute nerves from the ganglion for the spiral auditory nerve. The ganglion itself is enclosed by a spiral canal (sometimes referred to as Rosenthal's canal), which is embedded within the axial root (base) of the primary bony lamina (Fig. 2). In contrast to the inner hair cells, which are attached to afferent nerve bundles, the outer hair cells are mostly connected to efferent nerve fibers (Eybalin, 1993; Fettiplace & Hackney, 2006). Myelinated cochlear nerve fibers extend radially from the spiral ganglion canal to the inner hair cells. Unmyelinated nerve fibers cross the lumen of the tunnel of Corti between the inner and outer hair cells (see general review by Raphael & Altschuler, 2003).
An acellular strip of tissue known as the tectorial membrane is positioned vestibular to (above) the organ of Corti (Fig. 2). Two thickened regions that overhang the organ of Corti characterize the tympanic surface of the tectorial membrane. The thickened region along the radial border of the tectorial membrane is known as Hardesty's membrane and attaches directly to the stereocilia of the outer hair cells (Dallos et al. 1972; Hoshino, 1977; Raphael & Altschuler, 2003). In contrast, the stereocilia of the inner hair cells are free in most mammals but may come into contact with the axial thickened region of the cochlea known as Hensen's stripe (Hoshino, 1977; Raphael & Altschuler, 2003). Displacement of the basilar membrane relative to the tectorial membrane leads to the mechanotransduction of a mechanical force to an electrical signal via deflection of the stereocilia of the hair cells (Hudspeth, 1989). Even small displacements of vertebrate hair cells will elicit a nervous response (Rhode & Geisler, 1967; Corey & Hudspeth, 1983), which likely relates to the sensitivity of the cochlea. Functions of the inner and outer hair cells differ as can be inferred from the different nerve fiber types contacting each type of cell (Eybalin, 1993). The inner hair cells likely carry a sensory function only (see review by Fettiplace & Hackney, 2006), but the sensory capability of the outer hair cells is likely low given the paucity of afferent neurons in contact with those cells. Rather, the outer hair cells serve a mechanical feedback function (Dallos, 1992; Fettiplace, 2006).
Vestibular system
The bony vestibule of all jawed vertebrates (cartilaginous fishes, bony fishes, amphibians, reptiles, and mammals) is characterized by three semicircular canals. Jawless vertebrates, including living lampreys and several extinct ‘ostracoderms’, possess two semicircular canals, and extant hagfish only possess one (McVean, 1991; Mazan et al. 2000). The bony semicircular canals arise from a specific chamber of the bony vestibule known as the elliptical recess. The bony elliptical recess, which houses the membranous utricle, is separated from the cochlea by the bony spherical recess, which houses the membranous saccule of the vestibule and communicates with the stapes via the fenestra vestibuli (Fig. 1). The elliptical and spherical recesses themselves are separated by a slight constriction of the bony vestibule known as the vestibular crus and do not accurately represent the shapes of the membranous structures (Fig. 1; Ekdale, 2013). In general, the globular saccule is smaller than the oblong utricle of the membranous vestibule. The two membranous sacs do not communicate directly, but rather ducts leading to the endolymphatic sac bridge the two (Fig. 1; Evans, 1993).
There are two types of vestibular end organs within the membranous vestibule, which are the cristae within the ampullae of the semicircular ducts and the maculae within the utricle and saccule. Collectively, the maculae of the utricle and saccule are known as the otolithic organs, which detect linear accelerations. The maculae of the saccule detect acceleration along a vertical axis, whereas the maculae of the utricle detect longitudinal acceleration and gravity (Johnsson & Hawkins, 1967). The maculae and cristae possess hair cells that are similar in form and function to those found within the cochlea. Within the maculae, the hair cells are covered with an otolithic membrane that consists of microscopic otoconia embedded within a gelatinous layer. Movements of the head cause the otolithic membrane to shift in position, thereby exciting the hair cells within the maculae (Johnsson & Hawkins, 1967).
While the otolithic organs detect linear accelerations, the semicircular ducts are sensitive to rotations of the head. Within each ear, there is a single horizontal (lateral) semicircular canal and two vertical (anterior and posterior) semicircular canals (Fig. 3). The anterior and posterior semicircular canals, as well as the membranous ducts within, are joined to form a common crus that empties into the posteromedial region of the vestibule. In some mammals, the lateral and posterior semicircular canals join to form a secondary common crus (Macrini et al. 2010; Ekdale, 2013). Each canal opens into a swollen chamber known as an ampulla, which opens into the vestibule (Fig. 1). The membranous ampullae open into the utricle along with the membranous common crus. The ampullae of the vestibular system contain hair cells emerging from a patch of sensory epithelium known as the crista. Hair bundles extend from the crista into the gelatinous cupula. As the head rotates, movement of the endolymph within the semicircular ducts will displace the cupula, thereby exciting the hair cells (Hillman & McLaren, 1979; McLaren & Hillman, 1979; Rabbitt et al. 2009).
Figure 3.
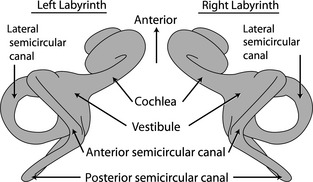
Line drawings of left and right bony labyrinths of a generalized mammal in dorsal view.
The semicircular canals are oriented in general to detect rotations in the pitch (anterior–posterior), roll (left–right on a vertical plane), and yaw (left–right on a horizontal plane) directions. Each canal works with a contralateral partner on the other side of the head. Both left and right lateral semicircular canals work in concert, and each anterior canal works with the opposite posterior canal. Given the orientations of the semicircular canals, each vertical canal is antagonistic with its contralateral counterpart (e.g. right anterior vs. left anterior; Fig. 3). In an ideal physiological model there is symmetry between the left and right vestibular systems so that synergistic canal pairs are parallel and the antagonistic canal pairs are perpendicular (David et al. 2010). However, the planes of ipsilateral canals (on the same side of the head) rarely form right angles in most species and synergistic canal pairs are almost never parallel (Malinzak et al. 2012; Berlin et al. 2013; Ekdale, 2013).
Anatomical and functional variation
Morphological variation is a natural phenomenon that affects all anatomical systems. With regard to the inner ear, morphological variation most often is correlated with auditory and locomotor function. For example, the number of inner and outer hair cell cilia within the cochlea differs among mammalian species, as do the lengths of the cilia (see review by Echteler et al. 1994). The lengths of the outer hair cells in particular are negatively correlated with the frequency at which sensitivity is the greatest so that the outer hair cells increase in length as the upper frequency limit decreases (Echteler et al. 1994: fig. 5.4). The lengthening of the cells also progresses from the base to the apex of the basilar membrane within an individual cochlea, correlating with a downward shift in peak sensitivity along the length of the cochlear organ.
Variations in the cochlea
Building upon the seminal comparative anatomy works of Hyrtl (1845), Gray (1907, 1908), and Fleischer (1973), macroscopic features of the cochlea that are thought to be correlated with hearing physiology in mammals include the total volume of the cochlea (Kirk & Gosselin‐Ildari, 2009; Armstrong et al. 2011), basilar membrane width (Wever et al. 1971) and length (West, 1985; Coleman & Colbert, 2010), especially coupled with the number of turns completed by the cochlear spiral (West, 1985), as well as the graded curvature of the spiral (Manoussaki et al. 2006, 2008) and ‘tightness’ of coiling (Fleischer, 1976).
Cochlear volume has been hypothesized to correspond to auditory physiology in primates in that an increase in the volume of the cochlear cavity relative to body mass indicates a lowering of both low and high frequency sensitivity thresholds (Kirk & Gosselin‐Ildari, 2009). In turn, the cochleae of primates tend to be more voluminous relative to body mass than their close relatives, including rodents, colugos (‘flying lemurs’), and tree shrews (Armstrong et al. 2011). Evidence from the fossil record of primates shows that the increase in cochlear volume and the presumed downward shift in frequency thresholds were relatively late events in primate history in that they occurred at the base of Euprimates, which includes all living primate species, and may have coincided with an expansion of the brain (Armstrong et al. 2011). As of yet there has not been an attempt to extend these observations outside of primates and their closest relatives, but the notion opens the door for future research.
Perhaps the features of the cochlea that have received the greatest attention regarding their relationship to hearing physiology are the dimensions and stiffness of the basilar membrane. The stiffness of the membrane is positively related to the frequencies at which the cochlea is most sensitive (Wever et al. 1971; Pye, 1979), and observations confirm that mammals sensitive to the highest frequency sound waves have the thickest and stiffest basilar membranes (Békésy, 1970; Wever et al. 1971; Echteler et al. 1994; Wartzok & Ketten, 1999). Because the basilar membrane is a soft tissue structure, it does not preserve well in the fossil record. Thus, direct measurements of basilar membrane thickness and stiffness are unavailable for extinct species. However, the width of the basilar membrane, which can be estimated from the distance between the primary and secondary bony spiral laminae (so‐called ‘basilar gap’ of Fleischer, 1976 and ‘laminar gap’ of Geisler & Luo, 1996), relates to the rigidity of the membrane, in that a narrow basilar membrane will be more rigid than a wide membrane. Thus, estimation of the basilar membrane width using the laminar gap as a proxy can be and has been used to reconstruct the physiologies of extinct mammals (Fleischer, 1976; Court, 1992; Luo & Eastman, 1995; Geisler & Luo, 1996; Luo & Marsh, 1996; Ekdale & Rowe, 2011). Animals with well‐developed primary and secondary bony spiral laminae that are narrowly separated will have narrow and stiff basilar membranes and will be sensitive to higher frequency sounds. This pattern is observed in echolocating bats (Ramprashad et al. 1979) and toothed whales (Wever et al. 1971; Fleischer, 1976), but not in elephants and baleen whales, which are sensitive to much lower frequencies (Fleischer, 1973; Ekdale & Racicot, 2015).
Although the primary and secondary bony laminae preserve readily in the fossil record, they are extremely delicate structures and rarely are preserved completely. Caution has been advised in using the laminar gap as a proxy for basilar membrane width, and estimates using the laminae may be off by as little as 25% or much, as over 100% of the actual membrane width for whales (Ketten, 2000). On the other hand, the proximal end of the secondary bony spiral lamina that attaches to the radial wall of the cochlear canal is much more robust than the distal edge that opposes the primary bony lamina, and so the proximal end has a greater preservation potential. The proportion of the cochlear canal through which the base of the secondary lamina extends provides a rough estimate for the overall development of the lamina. It has been demonstrated in whales that the secondary bony lamina is present for a greater proportion of the cochlear canal in species that are sensitive to higher frequencies than other whales adapted to detecting low frequency vibrations (Ketten, 2000; Ekdale & Racicot, 2015). Similar observations have yet to be calculated for many non‐cetacean mammals, but the secondary bony spiral lamina appears to extend for a greater proportion in the echolocating bats Rhinolophus and Tadarida than in other non‐aquatic mammals (Ekdale, 2013: table 2).
As may be expected, variations in the nervous supply of the spiral organ of Corti among various mammals correspond to physiological variations. In particular, the numbers of neurons connected to specific regions of the organ of Corti, as well as the size of the spiral ganglion canal relate to auditory function. For example, the regions responsible for processing echolocation signals in several species of microchiropteran bats that utilize biosonar exhibit a high density of cochlear neurons (Bruns & Schmieszek, 1980; Burda et al. 1988). There is also an increase in nerve cell number coupled with an increase in the cross‐sectional area of the spiral ganglion nerve canal in the region of peak frequency sensitivity in mice (Johnson et al. 2011). This so‐called ‘acoustic fovea’ that is marked by an increase in cochlear neuron density occurs in regions for best frequency sensitivity in several species adapted to low frequency perception (e.g. mole rats; Müller et al. 1992).
Species that are particularly sensitive to high frequency sounds tend to have spiral ganglion canals with large diameters, as has been observed in echolocating bats (Pye, 1966a,b, 1967) and toothed whales (Wever et al. 1971; Fleischer, 1976; Luo & Eastman, 1995; Luo & Marsh, 1996). Conversely, the spiral ganglion canals are relatively narrower in baleen whales and extinct whale ancestors (Geisler & Luo, 1996; Ekdale & Racicot, 2015), as well as non‐echolocating megachiropteran bats (Pye, 1966b). The cross‐sectional area of the canal can be used to estimate the number of spiral ganglion neurons and hair cells within the organ of Corti in mice (Johnson et al. 2011), and the same might apply to other mammals. At the very least, the size of the spiral ganglion canal can serve as an osteological correlate for high vs. low frequency sensitivity.
Perhaps the most obvious variation in cochlear morphology among mammals is the number of turns that the cochlear spiral completes (Fig. 4). Among therian mammals, the cochlea coils between just under a single turn in a few Mesozoic taxa (McKenna et al. 2000; Ekdale & Rowe, 2011) to over four turns in caviomorph rodents such as guinea pigs (Gray, 1907, 1908; Ekdale, 2013). Correlations have been found between the number of turns completed by the cochlea, especially if that value is multiplied by the absolute length of the cochlear canal (West, 1985; Manoussaki et al. 2008). In particular, a large product (number of turns multiplied by cochlear length) is indicative of mammals with sensitivity to low frequency sounds, such as elephants and cows. Tonotopical mapping indicates that low frequencies are detected in the apical regions of the basilar membrane and high frequencies are detected near the base (Liberman, 1982; Müller, 1996) following variations in outer hair cell dimensions; thus a lengthening of the cochlea would expand the area of low frequency sensitivity. This may also correspond to the expansion of frequency bandwidths at which mammals are sensitive over other vertebrates (Echteler et al. 1994).
Figure 4.
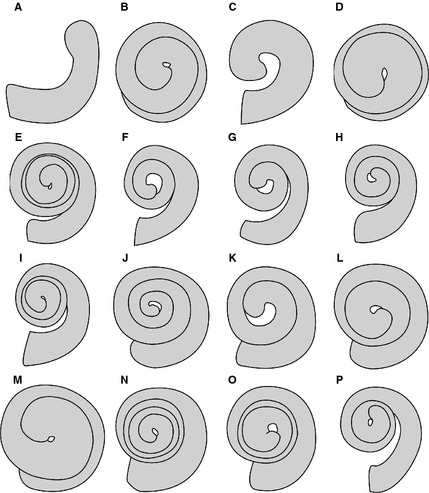
Line drawings of right cochleae in vestibular for a diversity of mammals. (A) Platypus (Ornithorhynchus anatinus); (B) opossum (Didelphis virginiana); (C) Mesozoic eutherian (Kulbeckia kulbecke); (D) elephant shrew (Macroscelides proboscideus); (E) pig (Sus scrofa); (F) humpback whale (Megaptera novaeangliae); (G) bottlenose dolphin (Tursiops truncatus); (H) horse (Equus caballus); (I) cat (Felis catus); (J) greater horseshoe bat (Rhinolophus ferrumequinum); (K) flying fox (Pteropus lylei); (L) cottontail rabbit (Sylvilagus floridanus); (M) house mouse (Mus musculus); (N) guinea pig (Cavia porcellus); (O) tree shrew (Tupaia glis); (P) human (Homo sapiens). The cochlea of the platypus was drawn following digital reconstructions from a figure by Luo et al. (2011: fig. 3b); all others were drawn from digital reconstructions described by Ekdale (2013).
It has been hypothesized that coiling of the cochlea is an adaptation to overcome a packaging problem associated with elongation of the basilar membrane in mammals (Meyer, 1907), although there is not a strong correlation between number of turns and length of the cochlea (West, 1985; Ekdale, 2013). Recent research on cochlear mechanics has revealed that the curved nature of the canal affects pressure distributions across the basilar membrane (Zhang et al. 2007) and might enhance a stimulus response to low frequencies, and even sound localization (Manoussaki et al. 2006; Huang et al. 2012). In particular, the graded curvature of the cochlea, or change in diameter between the basal and apical turns, appears to correlate with low frequency sensitivity. Terrestrial mammals that are sensitive to low frequencies, such as cows, elephants, and humans, tend to have greater ratios between the basal and apical cochlear turns compared with mammals with relatively higher low frequency thresholds, such as rats and mice (Manoussaki et al. 2008). The initial observations were made across a broad range of placental mammal clades and lifestyles, but there has yet to be a thorough study investigating the connection in groups of closely related organisms or for really specialized hearers. The major limiting factor is the availability of audiogram data for most mammal species, although the body of data is growing (e.g. a large number of audiograms are reported by Fay, 1988). Nonetheless, the correlation has been used to estimate the hearing physiologies of extinct ungulates (Orliac et al. 2012; Macrini et al. 2013) and cetaceans (Ekdale & Racicot, 2015).
An additional form–function connection that has been proposed, which may be related somewhat to the graded curvature of the cochlea, is the ‘tightness’ of cochlear coiling, or the distance between successive turns of the cochlea. Through a comparison of the cochleae of extinct and extant whales, Fleischer (1976) noted that the separation between the first (basal) and second turns of the cochlea was relatively greater in toothed whales sensitive to high frequencies than baleen whales. A similar pattern was observed in a greater sampling of baleen whales (Ekdale & Racicot, 2015), but the phenomenon has yet to be investigated thoroughly in other mammal clades. Given that the distributions of wave pressure differ through the cochlea per its coiled nature, the difference in cochlear morphology beyond the simple number of turns (Fig. 4) likely affect the function of the auditory organs of the inner ear.
Fleischer (1976) also described an expansion of the scala tympani in baleen whales that he termed the ‘tympanal recess’. An undivided perilymphatic foramen in which the fenestra cochleae is confluent with the cochlear aqueduct in elephants and manatees (Court, 1990, 1992; Fischer, 1990; Ekdale et al. 2011) may be an adaptation for low frequency hearing (Court, 1994). The confluence of those openings, which also characterizes early non‐therian mammals (Zeller, 1985, 1989; Wible, 1990), would expand the scala tympani in the same region as the tympanal recess observed in baleen whales by Fleischer (1976) and Ekdale & Racicot (2015). Interestingly, both a tympanal recess and an undivided perilymphatic foramen are present in gray whales and some juvenile fin whales (Geisler & Luo, 1996; Ekdale et al. 2011), both of which are thought to be sensitive to low frequency sounds. It may be that the presence of both features indicates a further downward shift in the low frequency thresholds of those mysticetes relative to odontocetes and the common ancestor of crown Cetacea, but there are no audiograms available for any baleen whale species that could be used to test such a hypothesis. Given the apparent plasticity and potential reversal to an ancestral mammalian morphology, further study of the functional implications of the undivided perilymphatic foramen as well as its taxonomic distribution would shed some light on the evolution of form and function of the inner ear in several mammal groups.
As has been shown, there is some extreme between‐species variation in the cochlea, but some aspects of the cochlea vary within species as well. These include absolute length of the canal and number of turns (degree of coiling). However, it is unclear how those intraspecific morphological variations relate to intraspecific variations in physiology. For example, the length of the cochlear canal is positively correlated with body mass (Ekdale, 2013), although it is unclear if larger bodied individuals are sensitive to a broader or narrower bandwidth of frequencies than smaller bodied individuals of the same species are. Contrary to cochlear length, the number of turns completed by the cochlea does not exhibit a strong correlation with body mass, nor does the degree of coiling vary much in most mammals (Ekdale & Rowe, 2011). One exception is a range of a nearly one quarter turn (90°) that was calculated for a population of captive opossums Monodelphis domestica (Ekdale, 2010). Unfortunately, audiograms for the individuals investigated by Ekdale (2010) were not recorded. The cochleae of two gray whale individuals of different maturities (juvenile and adult) were compared, and although the number of cochlear turns, cochlear canal length, and graded curvature of the cochlea varied slightly between the individuals, the low frequency limits estimated for each individual were comparable (Ekdale & Racicot, 2015).
Variations in the semicircular canals
A lot of recent attention has been directed towards variations within the vestibular system. The connection between the semicircular canals and rotational sensations has been known for nearly a century and a half (Dercum, 1879), and the connections between canal anatomy, orientation, sensitivity, behavior, and phylogeny is an active field of investigation in evolutionary biology. Several studies have used the size and shape of the semicircular canals to infer the agilities and locomotor abilities of numerous extinct and extant mammals, such as primates, whales, ungulates, and sloths (Spoor et al. 2002, 2007; Silcox et al. 2009; Macrini et al. 2010, 2013; Orliac et al. 2012; Billet et al. 2013). For example, the arc of the anterior semicircular canal tends to be more elliptical in marine carnivorans such as pinnipeds than in terrestrial species such as canids (Fig. 5; Georgi, 2008; Ekdale, 2013). Similar correlates between semicircular morphology and ecology also have been found for other non‐mammalian vertebrates (Clack, 2002; Witmer et al. 2003; Clarke, 2005; Georgi & Sipla, 2008; Georgi et al. 2013). However, phylogeny has a strong effect on the evolutionary development of semicircular canal morphology, so ecological comparisons among distantly related taxa may not be appropriate (Georgi & Sipla, 2008).
Figure 5.
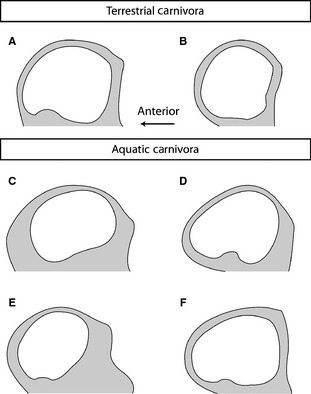
Line drawings of the anterior semicircular canal for terrestrial (A,B) and aquatic (C‐F) carnivorans. (A) Domestic dog (Canis familiaris; redrawn from Ekdale, 2013); (B) wolf (Canis lupus; redrawn from Georgi, 2008); (C) Stellar's sea lion (Eumetopias jubatus; redrawn from Ekdale, 2013); (D) Galapagos sea lion (Arctocephalus galapagoensis; redrawn from Georgi, 2008); (E) California sea lion (Zalophus californianus; redrawn from Georgi, 2008); (F) leopard seal (Hydrurga leptonyx; redrawn from Georgi, 2008).
There has yet to be a comprehensive and systematic study of variations in semicircular canal shape as they relate to locomotor differences across a broad array of mammals representing a spectrum of behaviors. Nor has the shape of a semicircular canal arc been linked directly to the canal's sensitivity. However, neurophysiological data support a connection between the size of the semicircular canal circuit, which is typically expressed as the radius of the arc of the semicircular canal (Jones & Spells, 1963; Spoor & Zonneveld, 1995; Spoor et al. 2007), and the sensitivity of the canal (Yang & Hullar, 2007). The general pattern is that large semicircular canals are more sensitive to rotations in space than are small canals. Interestingly, slow‐moving animals tend to have smaller canals (relative to body mass) than faster‐moving animals, which suggests that there is a connection between semicircular canal size, agility, and even locomotor behaviors (Spoor et al. 2007; Silcox et al. 2009; Berlin et al. 2013).
The anterior semicircular canal tends to have the greatest radius of curvature in most mammals, which suggests that most mammals are most sensitive to pitch rotations (anterior–posterior) of the head (Curthoys et al. 1977; Calabrese & Hullar, 2006; Spoor et al. 2007; Ekdale, 2013). However, variation in the largest canal arc among species may indicate functional differences. For example, the lateral semicircular canal is the most sensitive (greatest arc radius of curvature) in some marine mammals, including sea lions, manatees, and bottlenose dolphins (Ekdale, 2013), which might indicate that sensitivity to yaw rotations (left–right along a horizontal plane) is an adaptation for a secondary aquatic lifestyle in those animals. However, a recent investigation of the inner ears of extinct and extant cetaceans indicates that most whales follow the pattern of terrestrial mammals with large anterior semicircular canals relative to the others (Ekdale & Racicot, 2015), and thus a greater sensitivity to pitch rotations than yaw.
An overall reduction of the vestibular system might indicate fully aquatic lifestyles in marine mammals. For example, it is well known that the semicircular canals of cetaceans are small with respect to the cochlea (Yamada & Yoshizaki, 1959; Fleischer, 1976), and this pattern can be traced across cetacean evolution in the fossil record (Spoor et al. 2002; Ekdale & Racicot, 2015). The semicircular canal reduction in comparison with terrestrial mammals has been related to an obligate aquatic lifestyle, and in particular an adaptation for rapid head rotations in an aquatic environment (Spoor et al. 2002). Two or more cervical vertebrae are fused in many extant cetaceans (Barnes & McLeod, 1984; Geisler & Sanders, 2003), thereby limiting movement at the neck, and the vestibulo‐colic and vestibulo‐ocular reflexes that stabilize the head and eyes during rapid body rotations becomes ineffective (Calabrese & Hullar, 2006). In such situations, large and sensitive semicircular canals could not compensate for agile movements when the head is unable to rotate at the neck. Thus, a reduced vestibular system would reduce semicircular canal sensitivity and lessen any negative effects of an agile aquatic lifestyle with fused cervical vertebrae.
However, it is unclear whether the heads of cetaceans are rotating at greater velocities than those of mammals with mobile necks that can stabilize their heads. Rotational ratemeters were attached to captive‐born bottlenose dolphins (Tursiops truncatus) and a rodeo bull (Bos taurus) to test whether the acrobatic dolphins rotated their heads at a greater angular velocity than seen in an equally agile and closely related terrestrial relative (Kandel & Hullar, 2010). Interestingly, the head movements of the bull always exceeded those of the dolphins, thereby refuting the hypothesis that the reduced semicircular canals of cetaceans are adaptations to counteract increases in head rotations. Those results indicate that simplified models, such as correlations using semicircular canal arc radius of curvature, can limit inferences of head motions and behaviors. The motion and behavior could be ascertained using in vivo data for correlations to semicircular canal arc radii (Malinzak et al. 2012). Nonetheless, a reduced vestibular system may characterize the inner ears of aquatic mammals (Ekdale, 2013); however, as with semicircular canal shape, there has yet to be a systematic study of the issue.
An additional relationship that has been hypothesized to be related to behavior, and specifically aquatic behavior, is a dimensionless ratio between the radius and the length of the unampullated (slender) portion of the same semicircular canal (Boyer & Georgi, 2007). Changes in either dimension would relate to changes in frequency ranges transduced from head rotations among the taxa. In that study, ratios were calculated for two extinct eutherian mammals that presumably inhabited terrestrial habitats and a third extinct species thought to have been aquatic. There appeared to be a separation between the two terrestrial taxa and the aquatic species, although there is not a large body of data for aquatic mammals and close terrestrial relatives to test the significance of the observed pattern.
The size of a semicircular canal certainly is related to its sensitivity, but orientations of the canal planes likely affect the sensitivity of the canal system and perhaps also provide a behavioral signal (Malinzak et al. 2012; Berlin et al. 2013). The usefulness of the arc radius of curvature of semicircular canals in the interpretation of locomotion behaviors has been questioned recently, owing in large part to the influence of phylogeny on inner ear shapes and dimensions (Lebrun et al. 2010; Knoll et al. 2012; Benoit et al. 2013). Furthermore, the size of the inner ear labyrinth is strongly correlated with body mass, which can be problematic when attempting to ascertain the behaviors of extinct animals. In contrast, orientations of the planes of the semicircular canals corrected for phylogeny are independent of body mass (Malinzak et al. 2012).
In the simplest model of the vestibular system, the planes of the semicircular canals are parallel to the three orthogonal planes of the head. However, this is almost never the case (see discussion of Berlin et al. 2013). Because most ipsilateral semicircular canal pairs do not form right angles, more than one semicircular canal will elicit a response during head rotations. Different locomotion behaviors will cause the head to rotate in different fashions, and so the shapes and orientations of the canals should correlate with behavior. A general pattern has been observed among primates, in that species that move their heads rapidly tend to have semicircular canal pairs that are closer to orthogonality, or close to 90° (Malinzak et al. 2012). Furthermore, regressions between angular velocity magnitudes and the deviation of canal pairs from 90° (‘90var’ of Malinzak et al. 2012) separate fast leapers from slow quadrupeds. There is some suggestion that those patterns observed for primates may be extended to non‐primate mammals.
An interesting, and potentially important pattern that recently has emerged involves the intraspecific variation in the shapes and orientations of vestibular elements. In a study investigating the bony labyrinths of xenarthrans, it was found that the slow‐moving three‐toed sloths exhibited greater intraspecific variation in semicircular canal orientation than did faster moving species such as tamanduas and armadillos (Billet et al. 2012). Furthermore, the sizes of the semicircular canals of mice with different locomotor behaviors and agility (individuals bred for high voluntary wheel‐running vs. a non‐selected control group) do not differ, but the shapes of the canals do (Schutz et al. 2014). The implication is that there is intraspecific plasticity in the shape of the canals that relates to differences in locomotion, but not the size of the canals themselves. Therefore, variations in semicircular canal shape might indicate sensitivity variations even among very closely related individuals (within the same species) of very similar body size. The results of both studies accentuate the need for increased explorations of the intraspecific variation of semicircular canal morphology and orientations in order to fully elucidate the connection between form and function of the vestibular system.
Concluding remarks
The inner ear is one of the best‐studied regions of the nervous systems in vertebrates. The general form and function of the inner ear appear to be straightforward – the coiled cochlea is involved with hearing and the loops of the semicircular canals are involved with balance. Variations in the gross morphology among different species almost certainly coincide with the vast diversity of auditory and locomotion physiologies observed across Mammalia, and vertebrate morphologists are gaining a greater understanding about the amount of morphological variation between species. However, additional work is needed in the realm of intraspecific variation, including the degree and nature of coiling within the cochlear spiral, and shapes and orientations of the semicircular canals. In particular, there may be interesting connections between how interspecific form–function relationships translate to intraspecific variations in anatomy and physiology.
Acknowledgements
I thank M. Mason and A. Tucker (editors), as well as A. Ekdale, Z.‐X. Luo and an anonymous reviewer for helpful and constructive comments during the development and review process of the manuscript. The photograph of the cochlea in cross‐section was taken by N. Zellmer.
References
- Alexander G (1904) Entwichlung und Bau des innerens Gehörorgans von Echidna aculeala . Dekschr Mednaturwiss Ges Jena 3, 1–118. [Google Scholar]
- Alexander RM (2002) Stability and maneuverability of terrestrial vertebrates. Integr Comp Biol 42, 158–164. [DOI] [PubMed] [Google Scholar]
- Armstrong SD, Bloch JI, Houde P, et al. (2011) Cochlear labyrinth volume in Euarchontoglires: implications for the evolution of hearing in primates. Anat Rec 294, 263–266. [DOI] [PubMed] [Google Scholar]
- Barnes LG, McLeod SA (1984) The fossil record and phyletic relationships of gray whales In: The Gray Whale: Eschrichtius robustus. (eds Jones ML, Swartz SL, Leatherwood S.), pp. 3–32, Orlando: Academic Press. [Google Scholar]
- Békésy G (1970) Travelling wave as frequency analyzers in the cochlea. Nature 225, 1207–1209. [DOI] [PubMed] [Google Scholar]
- Benoit J, Essid EM, Marzougui W, et al. (2013) New insights into the ear region anatomy and cranial blood supply of advanced stem Strepsirhini: evidence from three primate petrosals from the Eocene of Chambi, Tunisia. J Hum Evol 65, 551–572. [DOI] [PubMed] [Google Scholar]
- Berlin JC, Kirk EC, Rowe TB (2013) Functional implications of ubiquitous semicircular canal non‐orthogonality in mammals. PLoS ONE 8, e79585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Billet G, Hautier L, Asher RJ, et al. (2012) High morphological variation of vestibular system accompanies slow and infrequent locomotion in three‐toed sloths. Proc R Soc B 279, 3932–3939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Billet G, Germain D, Ruf I, et al. (2013) The inner ear of Megatherium and the evolution of the vestibular system in sloths. J Anat 223, 557–567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boisvert CA (2005) The pelvic fin and girdle of Panderichthys and the origin of tetrapod locomotion. Nature 438, 1145–1147. [DOI] [PubMed] [Google Scholar]
- Bosher SK, Warren RL (1968) Observations on the electrochemistry of the cochlear endolymph of the rat: a quantitative study of its electrical potential and ionic composition as determined by means of flame spectrophotometry. Proc R Soc B 171, 227–247. [DOI] [PubMed] [Google Scholar]
- Boyer DM, Georgi JA (2007) Cranial morphology of a pantolestid eutherian mammal from the Eocene Bridger Formation, Wyoming, USA: implications for relationships and habitat. J Mamm Evol 14, 239–380. [Google Scholar]
- Bruns V, Schmieszek E (1980) Cochlear innervation in the greater horseshoe bat: demonstration of an acoustic fovea. Hear Res 3, 27–43. [DOI] [PubMed] [Google Scholar]
- Burda H, Fiedler J, Brums V (1988) The receptor and neuron distribution in the cochlea of the bat, Taphozous kachhensis . Hear Res 32, 131–135. [DOI] [PubMed] [Google Scholar]
- Calabrese DR, Hullar TE (2006) Planar relationships of the semicircular canals in two strains of mice. J Assoc Res Otolaryngol 7, 151–159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carlisle L, Steel K, Forge A (1990) Endocochlear potential generation is associated with intracellular communication in the stria vascularis: structural analysis in the variable dominant spotting mouse mutant. Cell Tissue Res 262, 329–337. [DOI] [PubMed] [Google Scholar]
- Casper BM, Mann DA (2006) Evoked potential audiograms of the nurse shark (Ginglymostoma cirratum) and the yellow stingray (Urobatis jamaicensis). Environ Biol Fish 76, 101–108. [Google Scholar]
- Casper BM, Lobel PS, Yan HY (2003) The hearing sensitivity of the little skate, Raja erinacea: a comparison of two methods. Environ Biol Fish 68, 371–379. [Google Scholar]
- Christensen‐Dalsgaard J, Manley GA (2005) Directionality of the lizard ear. J Exp Biol 208, 1209–1217. [DOI] [PubMed] [Google Scholar]
- Christensen‐Dalsgaard J, Manley GA (2008) Acoustical coupling of lizard eardrums. J Assoc Res Otolaryngol 9, 407–416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clack JA (2002) Patterns and processes in the early evolution of the tetrapod ear. J Neurobiol 53, 251–264. [DOI] [PubMed] [Google Scholar]
- Clarke AH (2005) On the vestibular labyrinth of Brachiosaurus brancai . J Vestib Res 15, 65–71. [PubMed] [Google Scholar]
- Coleman MN, Colbert MW (2010) Correlations between auditory structures and hearing sensitivity in non‐human primates. J Morphol 271, 511–532. [DOI] [PubMed] [Google Scholar]
- Corey DP, Hudspeth AJ (1983) Kinetics of the receptor current in bullfrog saccular hair cells. J Neurosci 3, 962–976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Court N (1990) Periotic anatomy of Arsinotherium (Mammalia, Embrithopoda) and its phylogenetic implications. J Vertebr Paleontol 10, 170–182. [Google Scholar]
- Court N (1992) Cochlea anatomy of Numidotherium koholense: auditory acuity in the oldest known proboscidean. Lethaia 25, 211–215. [Google Scholar]
- Court N (1994) The periotic of Moeritherium (Mammalia, Proboscidea): homology or homoplasy in the ear region of Tethytheria McKenna, 1975? Zool J Linn Soc 112, 13–28. [Google Scholar]
- Curthoys IS, Markham CH, Curthoys EJ (1977) Semicircular duct and ampulla dimensions in cat, guinea pig and man. J Morphol 151, 17–34. [DOI] [PubMed] [Google Scholar]
- Dallos P (1992) The active cochlea. J Neurosci 12, 4575–4585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dallos P, Billone MC, Durrant JD, et al. (1972) Cochlear inner and outer hair cells: functional differences. Science 177, 356–358. [DOI] [PubMed] [Google Scholar]
- David R, Droulez J, Allain R, et al. (2010) Motion from the past. A new method to infer vestibular capacities of extinct species. CR Palevol 9, 397–410. [Google Scholar]
- Dercum F (1879) On the morphology of the semicircular canals. Am Nat 13, 366–374. [Google Scholar]
- Echteler SM, Fay RR, Popper AN (1994) Structure of the mammalian cochlea In: Comparative Hearing: Mammals. (eds Fay RR, Popper AN.), pp. 134–171, New York: Springer‐Verlag. [Google Scholar]
- Ekdale EG (2010) Ontogenetic variation in the bony labyrinth of Monodelphis domestica (Mammalia: Marsupialia) following ossification of the inner ear cavities. Anat Rec 293, 1896–1912. [DOI] [PubMed] [Google Scholar]
- Ekdale EG (2013) Comparative anatomy of the bony labyrinth (inner ear) of placental mammals. PLoS ONE 8, e66624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ekdale EG, Racicot RA (2015) Anatomical evidence for low frequency sensitivity in an archaeocete whale: comparison of the inner ear of Zygorhiza kochii with that of crown Mysticeti. J Anat 226, 22–39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ekdale EG, Rowe T (2011) Morphology and variation within the bony labyrinth of zhelestids (Mammalia, Eutheria) and other therian mammals. J Vertebr Paleontol 31, 658–675. [Google Scholar]
- Ekdale EG, Berta A, Deméré TA (2011) The comparative osteology of the petrotympanic complex (ear region) of extant baleen whales (Cetacea: Mysticeti). PLoS One 6, e21311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evans H (1993) The ear In: Miller's Anatomy of the Dog, 3rd edn (ed. Evans HE.), pp. 988–1008, Philadelphia: Saunders. [Google Scholar]
- Eybalin M (1993) Neurotransmitters and neuromodulators of the mammalian cochlea. Phys Rev 73, 309–373. [DOI] [PubMed] [Google Scholar]
- Fay RR (1988) Hearing in Vertebrates: A Psychophysics Databook. Winnetka: Hill‐Fay Associates. [Google Scholar]
- Fettiplace R (2006) Active hair bundle movements in auditory hair cells. J Physiol 576, 29–36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fettiplace R, Hackney CM (2006) The sensory and motor roles of auditory hair cells. Nat Rev Neurosci 7, 19–29. [DOI] [PubMed] [Google Scholar]
- Fischer M (1990) Un trait unique de l'oreille des éléphants et des Sirérniens (Mammalia): un paradoze phylogénétique. C R Acad Sci III‐Vie 311, 157–162. [Google Scholar]
- Fleischer G (1973) Studien am Skelett des Gehörorgans der Säugetiere einschließlich des Menschen. Säugetierkunlihe Mirreilungen 21, 131–239. [Google Scholar]
- Fleischer G (1976) Hearing in extinct cetaceans as determined by cochlear structure. J Paleontol 50, 133–152. [Google Scholar]
- Furukawa T, Ishii Y (1967) Neurophysiological studies on hearing in goldfish. J Neurophysiol 30, 1377–1403. [DOI] [PubMed] [Google Scholar]
- Geisler JH, Luo Z‐X (1996) The petrosal and inner ear of Herpetocetus sp. (Mammalia: Cetacea) and their implications for the phylogeny and hearing of archaic mysticetes. J Paleontol 70, 1045–1066. [Google Scholar]
- Geisler JH, Sanders AE (2003) Morphological evidence for the phylogeny of Cetacea. J Mamm Evol 10, 23–129. [Google Scholar]
- Georgi JA (2008) Semicircular canal morphology as evidence of locomotor environment in amniotes. PhD Thesis. Stony Brook: Stony Brook University. [Google Scholar]
- Georgi JA, Sipla JS (2008) Comparative and functional anatomy of balance in aquatic reptiles and birds In: Sensory Evolution on the Threshold: Adaptations in Secondarily Aquatic Vertebrates. (eds Thewissen JGM, Nummela S.), pp. 233–256, Berkeley: University of California Press. [Google Scholar]
- Georgi JA, Sipla JS, Forster CA (2013) Turning semicircular canal function on its head: dinosaurs and a novel vestibular analysis. PLoS One 8, e59517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gerkema MP, Davies WIL, Foster RG, et al. (2013) The nocturnal bottleneck and the evolution of activity patterns in mammals. Proc R Soc B 280:2013.0508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gould E (1965) Evidence for echolocation in the Tenrecidae of Madagascar. Proc Am Philos Soc 109, 352–360. [Google Scholar]
- Gray AA (1903) On a method of preparing the membranous labyrinth. J Anat Physiol 37, 379–381. [PMC free article] [PubMed] [Google Scholar]
- Gray AA (1905) A demonstration of specimens prepared by the author's method to show the membranous labyrinth in man and lower animals. Trans Otol Soc UK 6, 9–13. [Google Scholar]
- Gray AA (1907) The Labyrinth of Animals: Including Mammals, Birds, Reptiles and Amphibians, Vol. 1 London: J. and A. Churchill. [Google Scholar]
- Gray AA (1908) The Labyrinth of Animals: Including Mammals, Birds, Reptiles and Amphibians, Vol. 2 London: J. and A. Churchill. [Google Scholar]
- Gunz P, Ramsier M, Kuhrig M, et al. (2012) The mammalian bony labyrinth reconsidered, introducing a comprehensive geometric morphometric approach. J Anat 220, 529–543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hall JD, Johnson CS (1972) Auditory thresholds of a killer whale Orcinus orca Linnaeus. J Acoust Soc Am 51, 515–517. [Google Scholar]
- Hall MI, Kamilar JM, Kirk EC (2012) Eye shape and the nocturnal bottleneck of mammals. Proc R Soc B 279, 4962–4968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Havia M, Kentala E, Pyykkö I (2002) Hearing loss and tinnitus in Meniere's disease. Auris Nasus Larynx 29, 115–119. [DOI] [PubMed] [Google Scholar]
- Hawkins JE (2004) Sketches of otohistory. Part 1: otoprehistory: how it all began. Audiol Neurootol 9, 66–71. [DOI] [PubMed] [Google Scholar]
- Heffner HE, Heffner RS (2007) Hearing ranges of laboratory animals. J Am Assoc Lab Anim Sci 46, 11–13. [PubMed] [Google Scholar]
- Hibino H, Nin F, Tsuzuki C, et al. (2010) How is the highly positive endocochlear potential formed? The specific architecture of the stria vascularis and the roles of the ion‐transport apparatus. Pflugers Arch 459, 521–533. [DOI] [PubMed] [Google Scholar]
- Hillman DE, McLaren JW (1979) Displacement configuration of semicircular canal cupulae. Neuroscience 4, 1980–2000. [DOI] [PubMed] [Google Scholar]
- Hoshino T (1977) Contact between the tectorial membrane and the cochlear sensory hairs in the human and the monkey. Arch Otorhinolaryngol 217, 53–60. [DOI] [PubMed] [Google Scholar]
- Houser DS, Helweg DA, Moore PWB (2001) A bandpass filter‐bank model of auditory sensitivity in the humpback whale. Aquat Mamm 27, 82–91. [Google Scholar]
- Huang X, Xu C, Bai L (2012) Is the cochlea coiled to provide sound localization? Europhys Lett 98, 58002. [Google Scholar]
- Hudspeth AJ (1989) How the ear's works work. Nature 341, 397–404. [DOI] [PubMed] [Google Scholar]
- Hyrtl J (1845) Verleichende‐Anatomische Untersuchungen über das innere Gehörorgan des Menschen und der Säugethiere. Prague: Friedrich Ehrlich. [Google Scholar]
- Johnson SB, Schmitz HM, Santi PA (2011) TSLIM imaging and a morphometric analysis of the mouse spiral ganglion. Hear Res 278, 34–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnsson L‐G, Hawkins JE (1967) Otolithic membranes of the saccule and utricle in man. Science 157, 1454–1456. [DOI] [PubMed] [Google Scholar]
- Jones GM, Spells KE (1963) A theoretical and comparative study of the functional dependence of the semicircular canal upon its physical dimensions. Proc R Soc Lond B 157, 403–419. [DOI] [PubMed] [Google Scholar]
- Jones GP, Lukashkina VA, Russell IJ, et al. (2010) The vestibular system mediates sensation of low‐frequency sounds in mice. J Assoc Res Otolaryngol 11, 725–732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jørgenson JM, Locket NA (1995) The inner ear of the echidna Tachyglossius aculeatus: the vestibular sensory organs. Proc Biol Sci 260, 183–189. [DOI] [PubMed] [Google Scholar]
- Kandel BM, Hullar TE (2010) The relationship of head movements to semicircular canal size in cetaceans. J Exp Biol 213, 1175–1181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kermack KA, Mussett F (1983) The ear in mammal‐like reptiles and early mammals. Acta Palaeontol Pol 28, 147–158. [Google Scholar]
- Ketten DR (1997) Structure and function in whale ears. Bioacoustics 8, 103–135. [Google Scholar]
- Ketten DR (2000) Cetacean ears In: Hearing by Whales and Dolphins. (eds Au WWL, Popper AN, Ray RR.), pp. 43–108, New York: Springer‐Verlag. [Google Scholar]
- Ketten DR, Wartzok D (1990) Three‐dimensional reconstructions of the dolphin ear In: Sensory Abilities of Cetaceans. (eds Thomas J, Kastelein R.), pp. 81–105, New York: Plenum. [Google Scholar]
- Kielan‐Jaworowska Z, Cifelli R, Luo Z‐X (2004) Mammals from the Age of the Dinosaurs. New York: Columbia University Press. [Google Scholar]
- Kirk EC, Gosselin‐Ildari AD (2009) Cochlear labyrinth volume and hearing abilities in primates. Anat Rec 292, 765–776. [DOI] [PubMed] [Google Scholar]
- Knoll F, Witmer LM, Ortega F, et al. (2012) The braincase of the basal sauropod dinosaur Spinophorosaurus and 3D reconstructions of the cranial endocast and inner ear. PLoS One 7, e30060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lebrun R, de Léon MP, Tafforeau P, et al. (2010) Deep evolutionary roots of strepsirrhine primate labyrinthine morphology. J Anat 216, 368–380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee FS (1898) Functions of the ear and the lateral line in fishes. Am J Physiol 1, 128–144. [Google Scholar]
- Liberman MC (1982) The cochlear frequency map for the cat: labeling auditory‐nerve fibers of known characteristic frequency. J Acoust Soc Am 72, 1441–1449. [DOI] [PubMed] [Google Scholar]
- Luo Z‐X, Eastman ER (1995) Petrosal and inner ear of a squalodontoid whale: implications for evolution of hearing in odontocetes. J Vertebr Paleontol 15, 431–442. [Google Scholar]
- Luo Z‐X, Marsh KK (1996) Petrosal (periotic) and inner ear of a Pliocene kogiine whale (Kogiinae, Odontoceti): implications on relationships and hearing evolution of toothed whales. J Vertebr Paleontol 16, 328–348. [Google Scholar]
- Luo Z‐X, Ruf I, Schultz JA, et al. (2011) Fossil evidence on evolution of inner ear cochlea in Jurassic mammals. Proc R Soc B 278, 28–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo Z‐X, Ruf I, Martin T (2012) The petrosal and inner ear of the Late Jurassic cladotherian mammal Dryolestes leiriensis and implications for ear evolution in therian mammals. Zool J Linn Soc 166, 433–463. [Google Scholar]
- Macrini TE, Flynn JJ, Croft DA, et al. (2010) Inner ear of a notoungulare placental mammal: anatomical description and examination of potentially phylogenetically informative characters. J Anat 216, 600–610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Macrini TE, Flynn JJ, Ni X, et al. (2013) Comparative study of notoungulate (Placentalia, Mammalia) bony labyrinths and new phylogenetically informative inner ear characters. J Anat 223, 442–461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maddin HC, Sherratt E (2014) Influence of fossoriality on inner ear morphology: insights from caecilian amphibians. J Anat 225, 83–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malinzak MD, Kay RF, Hullar TE (2012) Locomotor head movements and semicircular canal morphology in primates. Proc Natl Acad Sci U S A 109, 17914–17919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manley GA (1972) A review of some current concepts of the functional evolution of the ear in terrestrial vertebrates. Evolution 26, 608–621. [DOI] [PubMed] [Google Scholar]
- Manley GA, Kraus JEM (2010) Exceptional high‐frequency hearing and matched vocalizations in Australian pygopod geckos. J Exp Biol 213, 1876–1885. [DOI] [PubMed] [Google Scholar]
- Manoussaki D, Dimitriadis EK, Chadwick RS (2006) Cochlea's graded curvature effect on low frequency waves. Phys Rev Lett 96, 1–4. [DOI] [PubMed] [Google Scholar]
- Manoussaki D, Chadwick RS, Ketten DR, et al. (2008) The influence of cochlear shape on low‐frequency hearing. Proc Natl Acad Sci U S A 105, 6162–6166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mazan S, Jaillard D, Baratte B, et al. (2000) Otx1 gene‐controlled morphogenesis of the horizontal semicircular canal and the origin of the gnathostome characteristics. Evol Dev 2, 186–193. [DOI] [PubMed] [Google Scholar]
- McKenna MC, Kielan‐Jaworowska Z, Meng J (2000) Earliest eutherian mammal skull, from the Late Cretaceous (Coniacian) of Uzbekistan. Acta Palaeontol Pol 45, 1–54. [Google Scholar]
- McLaren JW, Hillman DE (1979) Displacement of the semicircular canal cupula during sinusoidal rotation. Neuroscience 4, 2001–2008. [DOI] [PubMed] [Google Scholar]
- McVean A (1991) The semicircular canals of the hagfish Myxine glutinosa . J Zool 224, 213–222. [Google Scholar]
- Megela‐Simmons A, Moss CF, Daniel KM (1985) Behavioral audiograms of the bullfrog (Rana catesbiana) and the green tree frog (Hyla cinerea). J Acoust Soc Am 78, 1236–1244. [DOI] [PubMed] [Google Scholar]
- Menaker M, Moreira LF, Tosini G (1997) Evolution of circadian organization in vertebrates. Braz J Med Biol Res 30, 305–313. [DOI] [PubMed] [Google Scholar]
- Meng J, Fox RC (1995) Osseous inner ear structures and hearing in early marsupials and placentals. Zool J Linn Soc 115, 47–71. [Google Scholar]
- Merchant SN, Adams JC, Nadol JB (2005) Pathophysiology of Ménière's syndrome: are symptoms caused by endolymphatic hydrops? Otol Neurotol 26, 74–81. [DOI] [PubMed] [Google Scholar]
- Meyer M (1907) An introduction to the mechanics of the inner ear. Univ Missouri Stud Sci Ser 11, 1–139. [Google Scholar]
- Miller MR (1966a) The cochlear duct of lizards. Proc Calif Acad Sci 33, 255–359. [Google Scholar]
- Miller MR (1966b) The cochlear ducts of Lanthonotus and Anelytropsis with remarks on the familial relationship between Anelytropsis and Dibamus . Occ Pap Calif Acad Sci 60, 1–15. [Google Scholar]
- Miller MR (1968) The cochlear duct of snakes. Proc Calif Acad Sci 35, 425–475. [Google Scholar]
- Minor L, Solomon D, Zinreich JS, et al. (1998) Sound‐ and/or pressure‐induced vertigo due to bone dehiscence of the superior semicircular canal. Arch Otolaryngol Head Neck Surg 124, 249–258. [DOI] [PubMed] [Google Scholar]
- Minor L, Cremer PD, Carey JP, et al. (2001) Symptoms and signs in superior canal dehiscence syndrome. Ann N Y Acad Sci 942, 259–273. [DOI] [PubMed] [Google Scholar]
- Moffat AJM, Capranica RR (1976) Auditory sensitivity of the saccule in the American toad (Bufo americanus). J Comp Physiol 105, 1–8. [Google Scholar]
- Müller M (1996) The cochlear place‐frequency map of the adult developing Mongolian gerbil. Hear Res 94, 148–156. [DOI] [PubMed] [Google Scholar]
- Müller M, Laube Burda H, Bruns V (1992) Structure and function of the cochlea in the African mole rat (Cryptomys hottentotus): evidence for a low frequency acoustic fovea. J Comp Physiol A 171, 469–476. [DOI] [PubMed] [Google Scholar]
- Myrberg AA (2001) The acoustical biology of elasmobranchs. Environ Biol Fish 60, 31–45. [Google Scholar]
- Naidu RC, Mountain DC (2007) Basilar membrane tension calculations for the gerbil cochlea. J Acoust Soc Am 121, 994–1002. [DOI] [PubMed] [Google Scholar]
- Novacek MJ (1977) Aspects of the problem of variation, origin and evolution of the eutherian auditory bulla. Mamm Rev 7, 131–149. [Google Scholar]
- Novacek MJ (1993) Patterns of diversity in the mammalian skull In: The Skull, Vol. 2 (eds Hanken J, Hall BK.), pp. 438–545, Chicago: University of Chicago Press. [Google Scholar]
- Orliac MJ, Benoit J, O'Leary MA (2012) The inner ear of Diacodexis, the oldest artiodactyl mammal. J Anat 221, 417–426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parks SE, Ketten DR, O'Malley JT, et al. (2007) Anatomical predictions of hearing in the North Atlantic right whale. Anat Rec 290, 734–744. [DOI] [PubMed] [Google Scholar]
- Payan P, Kossmann H, Watrin A, et al. (1997) Ionic composition of endolymph in teleosts: origin and importance of endolymph alkalinity. J Exp Biol 200, 1905–1912. [DOI] [PubMed] [Google Scholar]
- Payne KB, Langbauer WR Jr, Thomas EM (1986) Infrasonic calls of the Asian elephant (Elephas maximus). Behav Ecol Sociobiol 18, 297–301. [Google Scholar]
- Poole JH, Payne KB, Langbauer WR Jr, et al. (1988) Social contexts of some very low frequency calls of African elephants. Behav Ecol Sociobiol 22, 385–392. [Google Scholar]
- Pye A (1966a) The structure of the cochlea in Chiroptera I. Microchiroptera: Emballonuroidea and Rhinolophoidea. J Morphol 118, 495–510. [DOI] [PubMed] [Google Scholar]
- Pye A (1966b) The megachiroptera and vespertilionoidea of the microchiroptera. J Morphol 119, 101–120. [DOI] [PubMed] [Google Scholar]
- Pye A (1967) The structure of the cochlea in Chiroptera III. Microchiroptera: Phyllostomatoidea. J Morphol 121, 241–254. [DOI] [PubMed] [Google Scholar]
- Pye A (1979) The structure of the cochlea in some mammals. J Zool 187, 39–53. [Google Scholar]
- Rabbitt RD, Breneman KD, King C, et al. (2009) Dynamic displacement of normal and detached semicircular canal cupula. J Assoc Res Otolaryngol 10, 497–509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramcharitar J, Gannon DP, Popper AN (2006) Bioacoustics of fishes of the Family Sciaenidae (corakers and drums). Trans Am Fish Soc 135, 1409–1431. [Google Scholar]
- Ramprashad F, Landolt JP, Money KE, et al. (1979) A morphometric study of the cochlea of the little brown bat (Myotis lucifugus). J Morphol 160, 345–358. [DOI] [PubMed] [Google Scholar]
- Raphael Y, Altschuler RA (2003) Structure and innervation of the cochlea. Brain Res Bull 60, 397–422. [DOI] [PubMed] [Google Scholar]
- Retzius G (1884) Das Gehörorgan der Wirbelthiere: morphologisch‐histologische Studien. II. Das Gehörorgan der Reptilien, der Vögel und der Säugethiere. Stockholm: Samson and Wallin. [Google Scholar]
- Rhode WS, Geisler CD (1967) Model of the displacement between opposing points on the tectorial membrane and reticular lamina. J Acoust Soc Am 42, 185–190. [DOI] [PubMed] [Google Scholar]
- Ridgway SH, Bullock TH, Carder DA, et al. (1981) Auditory brainstem response in dolphins. Proc Natl Acad Sci U S A 78, 1943–1947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rowe T (1988) Definition, diagnosis, and origin of Mammalia. J Vertebr Paleontol 8, 241–264. [Google Scholar]
- Ruf I, Luo Z‐X, Wible JR, et al. (2009) Petrosal anatomy and inner ear structures of the Late Jurassic Henkelotherium (Mammalia, Cladotheria, Dryolestoidea): insight into the early evolution of the ear region in cladotherium mammals. J Anat 214, 679–693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruf I, Luo Z‐X, Martin T (2013) Reinvestigation of the basicranium of Haldanodon exspectatus (Mammaliaformes, Docodonta). J Vertebr Paleontol 33, 382–400. [Google Scholar]
- Saidel WM, Popper AN (1983) The saccule may be the transducer for directional hearing of nonostariophysine teleosts. Exp Brain Res 50, 149–152. [DOI] [PubMed] [Google Scholar]
- Schmelzle T, Sánchez‐Villagra MR, Maier W (2007) Vestibular labyrinth diversity in diprotodontian marsupial mammals. Mamm Study 32, 83–97. [Google Scholar]
- Schmidt RS (1964) Phylogenetic significance of lizard cochlea. Copeia 1964, 542–549. [Google Scholar]
- Schutz H, Jamniczky HA, Hallgrímsson B, et al. (2014) Shape‐shift: semicircular canal morphology responds to selective breeding for increased locomotor activity. Evolution 68, 3184–3198. [DOI] [PubMed] [Google Scholar]
- Shubin NH, Daeschler EB, Coates MI (2004) The early evolution of the tetrapod humerus. Science 304, 90–93. [DOI] [PubMed] [Google Scholar]
- Shubin NH, Daeschler EB, Jenkins FA (2014) Pelvic girdle and fin of Tiktaalik roseae . Proc Natl Acad Sci U S A 111, 893–899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shute CCD, Bellairs AD'A (1953) The cochlear apparatus of Geckonidae and Pygopodidae and its bearing on the affinities of these groups of lizards. Proc Zool Soc Lond 123, 695–709. [Google Scholar]
- Silcox MT, Bloch JI, Boyer DM, et al. (2009) Semicircular canal system in early primates. J Hum Evol 56, 315–327. [DOI] [PubMed] [Google Scholar]
- Simmons JA, Fenton MB, O'Farrell MJ (1979) Echolocation and pursuit of prey by bats. Science 203, 16–21. [DOI] [PubMed] [Google Scholar]
- Spoor F, Zonneveld F (1995) Morphometry of the primate bony labyrinth: a new method based on high‐resolution computed tomography. J Anat 186, 271–286. [PMC free article] [PubMed] [Google Scholar]
- Spoor F, Zonneveld F (1998) Comparative review of the human bony labyrinth. Yearb Phys Antropol 41, 211–251. [DOI] [PubMed] [Google Scholar]
- Spoor F, Wood B, Zonneveld F (1996) Evidence for a link between human semicircular canal size and bipedal behaviour. J Hum Evol 30, 183–187. [Google Scholar]
- Spoor F, Bajpai S, Hussain ST, et al. (2002) Vestibular evidence for the evolution of aquatic behaviour in ear cetaceans. Nature 417, 163–166. [DOI] [PubMed] [Google Scholar]
- Spoor F, Garland T Jr, Krovitz G, et al. (2007) The primate semicircular canal system and locomotion. Proc Natl Acad Sci U S A 104, 10808–10812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sterkers O, Ferrary E, Amiel C (1988) Production of inner ear fluids. Physiol Rev 68, 1083–1128. [DOI] [PubMed] [Google Scholar]
- Tasaki I, Spyropoulos CS (1959) Stria vascularis as source of endocochlear potential. J Neurophysiol 22, 149–155. [DOI] [PubMed] [Google Scholar]
- Todd N (2001) Evidence for a behavioral significance of saccular acoustic sensitivity in humans. J Acoust Soc Am 110, 380–390. [DOI] [PubMed] [Google Scholar]
- Todd NPA, Cody FW (2000) Vestibular responses to loud dance music: a physiological basis of the ‘rock and roll threshold’? J Acoust Soc Am 107, 496–500. [DOI] [PubMed] [Google Scholar]
- Todd NPA, Cody FWJ, Banks JR (2000) A saccular origin of frequency tuning in myogenic vestibular evoked potentials?: implications for human responses to loud noises. Hear Res 141, 180–188. [DOI] [PubMed] [Google Scholar]
- Todd NPA, Rosengren SM, Colebatch JG (2003) A short latency vestibular evoked potential (VsEP) produced by bone‐conducted acoustic stimulation. J Acoust Soc Am 114, 3264–3272. [DOI] [PubMed] [Google Scholar]
- Tomasi TE (1979) Echolocation by the short‐tailed shrew Blarina brevicauda . J Mammal 60, 751–759. [PubMed] [Google Scholar]
- van der Klaauw CJ (1930) On mammalian auditory bullae showing an indistinctly complex structure in the adult. J Mammal 11, 55–60. [Google Scholar]
- van der Klaauw CJ (1931) The auditory bulla in some mammals, with a general introduction to this region of the skull. Bull Am Mus Nat Hist 57, 1–352. [Google Scholar]
- Wartzok D, Ketten DR (1999) Marine mammal sensory systems In: Biology of Marine Mammals. (eds Reynolds J, Rommel S.), pp. 117–175, Washington, DC: Smithsonian Institution Press. [Google Scholar]
- Webster DB, Webster M (1980) Morphological adaptations of the ear in the rodent family Heteromyidae. Am Zool 20, 247–254. [Google Scholar]
- West CD (1985) The relationship of the spiral turns of the cochlea and length of the basilar membrane to range of audible frequencies in ground dwelling mammals. J Acoust Soc Am 77, 1091–1101. [DOI] [PubMed] [Google Scholar]
- Wever EG (1973) The ear and hearing in the frog, Rana pipiens . J Morphol 141, 461–478. [DOI] [PubMed] [Google Scholar]
- Wever EG, Vernon JA (1956) Sound transmission in the turtle ear. Proc Natl Acad Sci U S A 42, 292–299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wever EG, McCormick JG, Palin J, et al. (1971) Cochlea of the dolphin, Tursiops truncatus: the basilar membrane. Proc Natl Acad Sci U S A 68, 2708–2711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wible JR (1990) Petrosals of Late Cretaceous marsupials from North America, and a cladistic analysis of the petrosal in therian mammals. J Vertebr Paleontol 10, 183–205. [Google Scholar]
- Witmer LM, Chatterjee S, Franzosa J, et al. (2003) Neuroanatomy of flying reptiles and implications for flight, posture, and behaviour. Nature 425, 950–953. [DOI] [PubMed] [Google Scholar]
- Yamada M, Yoshizaki F (1959) Osseous labyrinth of Cetacea. Sci Rep Whales Res Inst 14, 291–304. [Google Scholar]
- Yang A, Hullar TE (2007) Relationship of semicircular canal size to vestibular‐nerve afferent sensitivity in mammals. J Neurophysiol 98, 3197–3205. [DOI] [PubMed] [Google Scholar]
- Zeller U (1985) The morphogenesis of the fenestra rotunda in mammals. Fortschr Zool 30, 153–157. [Google Scholar]
- Zeller U (1989) Die Enwicklung und Morphologie des Schädels von Ornithorhynchus anatinus (Mammalia: Prototheria: Monotremata). Abh Senckenberg Naturforsch Ges 545, 1–188. [Google Scholar]
- Zhang Y, Ck Kim, Lee K‐J‐B, et al. (2007) Resultant pressure distribution pattern along the basilar membrane in the spiral shaped cochlea. J Biol Phys 33, 195–211. [DOI] [PMC free article] [PubMed] [Google Scholar]