

Abstract
Despite renewed interest in the biogeography and evolutionary history of Old World tree frogs (Rhacophoridae), this family still includes enigmatic frogs with ambiguous phylogenetic placement. During fieldwork in four northeastern states of India, we discovered several populations of tree hole breeding frogs with oophagous tadpoles. We used molecular data, consisting of two nuclear and three mitochondrial gene fragments for all known rhacophorid genera, to investigate the phylogenetic position of these new frogs. Our analyses identify a previously overlooked, yet distinct evolutionary lineage of frogs that warrants recognition as a new genus and is here described as Frankixalus gen. nov. This genus, which contains the enigmatic ‘Polypedates’ jerdonii described by Günther in 1876, forms the sister group of a clade containing Kurixalus, Pseudophilautus, Raorchestes, Mercurana and Beddomixalus. The distinctiveness of this evolutionary lineage is also corroborated by the external morphology of adults and tadpoles, adult osteology, breeding ecology, and life history features.
Introduction
The Old World tree frog family Rhacophoridae currently contains over 380 species in 17 genera [1,2], of which eight genera were recognized during the last decade [3–9]. Yet, despite the increasing interest in stabilizing anuran systematics (e.g., [5,6,10–12]), this large neobatrachian radiation still contains several nominal taxa with ambiguous generic placements that have not yet been investigated using molecular phylogenetic analyses. Four of the recently recognized rhacophorid genera—Ghatixalus, Raorchestes, Beddomixalus and Mercurana [5,7,9]—were described from the Western Ghats of India, a region which has witnessed a two-fold increase in the number of known tree frogs (from 21 to 43 species) over the past decade, supported by molecular evidence (e.g., [7,9,13,14]). Although rhacophorids of the Western Ghats have received considerable attention, the Northeast Indian members of this family have not yet been subjected to rigorous phylogenetic studies.
India is a country that contains high biodiversity, with the Western Ghats united in a single biodiversity hotspot with Sri Lanka, and the Northeast Indian region being part of two biodiversity hotspots, the Indo-Burma hotspot and the Himalaya hotspot [15,16]. The eight northeastern states, viz., Arunachal Pradesh, Assam, Manipur, Meghalaya, Mizoram, Nagaland, Sikkim and Tripura are particularly interesting, because they form the transitional zone between the Indian subcontinent and South-East Asia. Despite Northeast India being recognized as one of the major centers of amphibian diversity, so far only the caecilian fauna of this region has been studied within a phylogenetic framework [17,18].
During extensive fieldwork in four Northeast Indian states, we discovered several populations of tree hole breeding frogs. One of these species appeared to be the nominal taxon Polypedates jerdonii, originally described by Günther [19] from “Darjeeling”, West Bengal. Ever since its description, Polypedates jerdonii has been assigned to various widespread genera such as Rhacophorus [20–25], Philautus (Kirtixalus) [26] and Philautus (Philautus) [27]. This taxonomic uncertainty was presumably due to characters such as large snout-vent size and fairly extensive webbing between toes, which is comparable to members of the genera Rhacophorus and Polypedates. However, this species has been retained in the genus Philautus until today, because of “the large unpigmented eggs that strongly suggest that this species might have a direct development” [27].
To clarify the evolutionary position of the Northeast Indian tree hole breeding frogs, we performed phylogenetic analyses of nuclear and mitochondrial gene fragments totaling ~ 3800 characters for 86 taxa representing all major rhacophorid lineages. We demonstrate that these frogs form a distinct evolutionary lineage that warrants recognition as a new genus, and provide a detailed account of their breeding ecology, larval behavior, morphology, and osteology.
Materials and Methods
Ethics statement
This study was conducted with permissions and following guidelines from the responsible authorities in the State Forest Departments (Arunachal Pradesh CWL/G/13(17)/06-07/PT/3878-83 dated 05 June 2009; Manipur 3/22/2006-WL dated 03 March 2009; Meghalaya FWC.G/16 dated 19 February 2009, FWC.G/173 dated 23 March 2009 and Nagaland FOR/GEN-42/2006 dated 29 May 2007, FG-3/28/92 dated 06 April 2009), Ministry of Environment, Forest and Climate Change, Government of India. Our protocols of collection and research complied with the provisions of the Wildlife (Protection) Act 1972, Government of India. Specific methods of collection, euthanasia, tissue sampling and fixation used in our study followed the guidelines for use of live amphibians and reptiles in field research by the American Society of Ichthyologists and Herpetologists (ASIH) (http://www.asih.org/pubs/herpcoll.html; dated 13 March 2006), and were approved by the internal ethical committee of Department of Environmental Studies, University of Delhi.
Field surveys and specimen collection
Field expeditions and sampling were carried out both during day and night in the monsoon months, from June to September of the years 2007–2010, in four Northeast Indian states: Arunachal Pradesh, Manipur, Meghalaya and Nagaland. Life history observations of ‘Polypedates’ jerdonii were done at night (between 18:00–22:00 hours) at Mawphlang Forest, Meghalaya (coordinates 25°26.29'N, 91°45.35'E, 1577 m asl) in June 2009. GPS coordinates were recorded using a Garmin 76CSx. Adult specimens were euthanized in 5% aqueous solution of tricaine methanesulfonate (MS222), fixed in 4% formalin and preserved in 70% ethanol. Immediately after euthanization, tissue samples were taken from the thigh muscle, preserved in 95% ethanol, and stored at –20°C in the Systematics Lab, Department of Environmental Studies, University of Delhi, India (SDBDU). Eggs and tadpoles (euthanized using MS222) were preserved in 3.5% formalin, buffered to pH 7.0 with sodium phosphate, monobasic (NaH2PO4H2O) and sodium phosphate, dibasic (anhydrous) (Na2HPO4). Voucher specimens are deposited in the Bombay Natural History Society Museum (BNHS), Mumbai, and Systematics Lab, University of Delhi (SDBDU).
Taxon sampling and DNA protocols
This study includes 87 taxa, 86 rhacophorid taxa representing 73 species of all genera within the family, with a sampling emphasis on ‘Polypedates’ jerdonii (eight specimens) and an unknown but closely related species (three specimens). A dicroglossid served as the outgroup for phylogenetic analyses. For 62 taxa, sequence data were obtained from GenBank (S1 Table). For the other 25 specimens, genomic DNA was extracted from muscle tissue using a standard extraction protocol by Sambrook et al. [28], or the Qiagen DNeasy blood and tissue kit (Qiagen, Valencia, CA, USA) following the manufacturer’s protocol. Our total data matrix encompasses fragments of two nuclear genes (RHOD: ~ 300 bp and RAG1: ~ 550 bp) and three mitochondrial genes (~ 1400 bp: 12SrRNA, tRNAVAL and 16SrRNA). Primer sets used for RHOD (Rhod1A, Rhod1D), RAG1 (Rag1-C, Rag 1-E), 12SrRNA and tRNAVAL (H3296, L2519), and 16SrRNA (16Sar, 16Sbr) were published by Bossuyt and Milinkovitch [29], Biju and Bossuyt [30], Richards and Moore [31], and Simon et al. [32], respectively. Sequencing was performed on both strands using a BigDye Terminator v3.1 Cycle Sequencing kit and ABI 3730 automated DNA sequencer (Applied Biosystems). Newly generated sequences were checked and assembled in ChromasPro v1.34 (Technelysium Pty Ltd.), and submitted to GenBank under accession numbers KU169932–KU170018 and KU230453–KU230461 (S1 Table).
Phylogenetic analyses
Sequences were aligned using ClustalX 1.64 [33]. Ambiguous sections were identified by eye for non-coding DNA and by comparison with amino acid sequences for coding DNA using MacClade v4.0 [34]. Phylogenetic estimations were obtained under Maximum Likelihood (ML) criteria and in a Bayesian framework. Maximum Likelihood analyses were performed with the concatenated dataset under the GTR+CAT model using RAxML 8.0 [35] on the Cipres Science Gateway [36]. The RAxML search was performed specifying 200 alternative runs on distinct random starting trees. Clade support was assessed by 1,000 rapid bootstrap replicates.
Bayesian analyses were performed with MrBayes 3.1.2 [37], using a GTR+G+I mixed model according to a gene-based data partition. Two parallel runs of four Markov chain Monte Carlo (MCMC) chains each were executed for 10 million generations, with a sampling interval of 500 generations and a burn-in corresponding to the first five million generations. Convergence of the parallel runs was confirmed by split frequency standard deviations (<0.01), and by potential scale reduction factors (~1.0) for all model parameters using Tracer v1.3 [38]. A Bayesian consensus phylogram and Bayesian Posterior Probabilities (BPP) were inferred from the last 10,000 sampled trees of both runs. Additional Bayesian analyses, using the above parameters, were done for the nuclear and mitochondrial datasets separately.
Adult morphology
Determination of sex and maturity were done by examining the gonads through a small ventral incision. Measurements and associated terminology follow Biju et al. [39]; webbing formulae follow Savage and Heyer [40] as modified by Myers and Duellman [41]. The amount of webbing relative to subarticular tubercles is described by numbering the tubercles 1–3, starting from the toe discs. The term shank is used here to refer to the lower part of the leg containing the tibia, and thigh is used for the upper part containing the femur. Measurements of all specimens were taken by using a digital slide-caliper, or a binocular microscope with a micrometer ocular, to the nearest 0.1 mm. All measurements provided in the taxonomy section and S2 Table are in millimeters. See S1A File for abbreviations used in this section, and S2 File for details of specimens examined in the study.
Adult Osteology
We used five osteological characters to compare the internal morphology of ‘Polypedates’ jerdonii with three relevant genera (Gracixalus, Kurixalus and Philautus) representing the evolutionary transitions in the backbone of the tree. These characters are: sphenethmoid; premaxillary teeth and structure of premaxillae; shape of the frontoparietals, vomerines, epicoracoidal bridge (= “base of the omosternum”, Liem [42]); sternum (= “metasternum”, Liem [42]); and carpal elements. The latter four characters were also used by Liem [42].
Osteological characterization was done for adults of ‘Polypedates’ jerdonii, Philautus aurifasciatus, Gracixalus gracilipes and Kurixalus eiffingeri. Two neutral buffered formalin preserved specimens of ‘Polypedates’ jerdonii (SDBDU 2009.1163 and SDBDU 2009.312a), and three 70% alcohol preserved specimens belonging to Philautus aurifasciatus (AMNH A24559), Gracixalus gracilipes (AMNH A163893) and Kurixalus eiffingeri (AMNH A14498) (the type species of the respective genera) were cleared and double stained following the procedure by Taylor and Van Dyke [43]. Initial dehydration was done in 100% ethanol, followed by submersion in alcian blue for cartilage staining. Excessive musculature was digested using an infusion of borax and trypsin, and the specimens were subsequently stained in alizarin red for bone visualization. Preparations were photographed and scored for bones and cartilage within 2–3 days after the staining procedure was completed. Osteological terminologies follow Trueb [44], Duellman and Trueb [45], Maglia et al., [46] Pugener and Maglia [47].
Larval morphology
Morphological studies are based upon examination of four tadpoles. Two tadpoles BNHS 5975 and SDBDU 2009.1295 (stage 36) were examined with light microscopy for external morphology. Prior to morphological examination, preserved tadpoles were transferred via a series of 35%, 55% and 70% alcohol and kept in each of these for at least 24 hours [43]. Larvae were staged according to Gosner [48]. External larval morphological terminology and measurements were adapted from McDiarmid and Altig [49] and Bowatte and Meegaskumbura [50]. Measurements were rounded to the nearest 0.01 mm using a stereomicroscope with a micrometer ocular or a digital slide-caliper. Oral morphology and margins of papillae were visualized using blue ink. In two specimens (stage 36), incisions were made up to 2 mm deep on the sides of the mouth opening to observe the jaw sheaths. See S1B File for abbreviations used in this section. For museums and other frequently used terms, see abbreviations in S1C File.
Nomenclatural acts
The electronic edition of this article conforms to the requirements of the amended International Code of Zoological Nomenclature (ICZN), and hence the new names contained herein are available under that Code from the electronic edition of this article. This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN. The ZooBank LSIDs (Life Science Identifiers) can be resolved and the associated information viewed through any standard web browser by appending the LSID to the prefix "http://zoobank.org/". The LSID for this publication is: urn:lsid:zoobank.org:pub:2EBBC122-024A-4678-9B94-27AE2A873889. The electronic edition of this work was published in a journal with an ISSN, which has been archived and is available from the following digital repositories: PubMed Central, LOCKSS.
Results and Discussion
1. Phylogenetic analyses reveal a distinct evolutionary lineage of tree hole breeding frogs
Alignment of our DNA sequences resulted in a dataset of 3811 bp, 3174 of which could be aligned unambiguously. Maximum Likelihood analyses of the total dataset produced a single tree, which is very similar to the Bayesian consensus phylogram (Fig 1 and S1 Fig). The majority of the relationships obtained are in agreement with previous studies (e.g., [51,52]). In contrast with previous studies, however, Theloderma moloch, an adult from Namdapha, Arunachal Pradesh, which is close to the type locality (“Upper Renging”, Arunachal Pradesh), groups together with the other Theloderma species. The previously ambiguously assigned “Theloderma moloch” (larva voucher 6225Rao) from Li et al. [53] nested within the newly described genus (Fig 1).
Fig 1. Bayesian consensus phylogram showing phylogenetic relationships among 86 taxa representing all known rhacophorid genera and one outgroup species.

Numbers above the branches represent Bayesian Posterior Probabilities, numbers below the branches represent Maximum Likelihood bootstrap values. Clade representing Frankixalus gen. nov. is shown in red. The specimen that was assigned to “Theloderma moloch” by Li et al. [53] is indicated by an asterisk. Colors of taxa labels represent the reproductive modes: blue, terrestrial foam-nesting, exotrophic tadpoles; orange, terrestrial gel-nesting, exotrophic tadpoles; green, terrestrial direct-developing, endotrophic tadpoles; cyan, aquatic gel-nesting, exotrophic tadpoles. The new genus Frankixalus is also a terrestrial gel-nesting form.
Interestingly, our tree hole breeding frogs (represented by ‘Polypedates’ jerdonii and close relatives) form a distinct lineage that is the sister clade of a group containing the genera Pseudophilautus, Raorchestes, Mercurana, Beddomixalus and Kurixalus. Based on this phylogenetic evidence, we propose to allocate all frogs of this lineage to a new genus.
Taxonomic treatment
Amphibia Linnaeus, 1758
Anura Fischer von Waldheim, 1813
Rhacophoridae Hoffman, 1932
Rhacophorinae Hoffman, 1932
Frankixalus gen. nov. urn:lsid:zoobank.org:act:9DB458C3-9D5A-4058-842A-CEA7D41BA79F
Fig 5. Geographic distribution of Frankixalus in Northeast India and China.
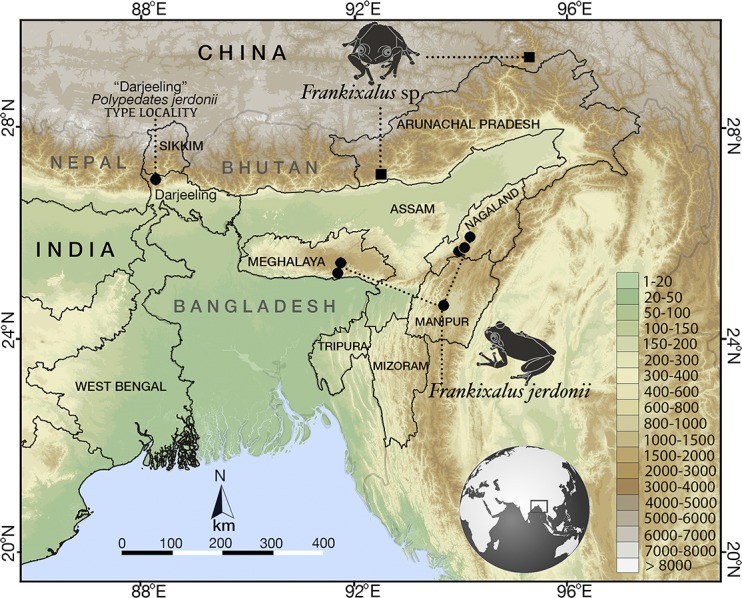
Circle = Frankixalus jerdonii, square = Frankixalus sp.
Etymology
The genus is named after Prof. Franky Bossuyt of the Vrije Universiteit Brussel (Belgium), as a token of appreciation for his contribution to amphibian research and herpetology education, and in particular for the valuable role he played in the scientific career of SDB and IVB. The generic epithet is derived from the name ‘Franky’ (used as a noun in the nominative singular) in conjugation with the genus name ‘Ixalus’ Duméril & Bibron, 1841, often used as a suffix in rhacophorid generic names. For the purposes of nomenclature, the gender of this genus is male.
Suggested common name
Franky’s tree frogs
Type species
Polypedates jerdonii Günther, 1876
Diagnosis
We consider Frankixalus to consist of the most inclusive clade that contains Frankixalus jerdonii comb. nov. but not Kurixalus eiffingeri. Frankixalus currently contains two species, F. jerdonii (Günther, 1876) and a currently unidentified species.
Frankixalus can be distinguished from the other rhacophorid genera by the combination of the following characters: medium-sized adults (male SVL 37.1–42.1 mm, N = 11; female SVL 46.8 mm, N = 1), webbing medium (foot webbing: I2––2+II1+–21/4III1+–11/2IV11/2−1+V; hand webbing: I1–1+II1+–2+III2––1+IV); creamy-white, gel-encapsulated eggs without pigmentation are laid in tree holes (phytotelm-breeding) where they also undergo development. The tadpole is oophagous and lacks keratinized tooth rows. The two currently included species are geographically restricted to high altitudes (approximately 1100–1600 m asl) in Northeast India and adjoining regions in China.
Comparison
Frankixalus differs from other rhacophorid genera as follows: Breeding, oviposition and developmental strategies (Fig 2): differs from Philautus, Pseudophilautus and Raorchestes by its free-living (exotrophic) tadpoles (vs. direct developing [endotrophic]); differs from Chiromantis, Ghatixalus, Polypedates, Rhacophorus and Taruga by its gel-encapsulated eggs (vs. eggs laid in foam nests); differs from Beddomixalus, Buergeria, Feihyla, Liuixalus and Mercurana by its phytotelm-breeding behavior, and oviposition on walls of tree holes (vs. terrestrial breeding and aquatic eggs in Beddomixalus, Buergeria and Liuixalus; leaf-breeding and oviposition on leaves in Feihyla; and terrestrial breeding and laid eggs mixed with mud in shallow pits in Mercurana). However, Frankixalus shares its breeding, oviposition and reproductive behavior with four other rhacophorid genera, namely Gracixalus, Nyctixalus, Theloderma and Kurixalus. External morphology (Fig 2): differs from all known rhacophorids by the combination of the following characters: outline of snout truncate in dorsal view, semi-circular in ventral view, vertical in lateral view (vs. not a combination of these characters), snout not protruding (vs. protruding). Tadpole morphology (Fig 3): differs from all known rhacophorids with exotrophic larvae by its absence of tooth rows (vs. present, except in Rhacophorus vampyrus) and presence of a sinistral, ventrally positioned spiracle (vs. sinistral, laterally positioned spiracle, except in Rhacophorus vampyrus). Presence of submarginal and marginal papillae shared with Polypedates, Taruga, Beddomixalus, Mercurana, Gracixalus and Mercurana (vs. absent in Rhacophorus vampyrus).
Fig 2. A–C, Frankixalus jerdonii in life.

(A) dorsolateral view of an adult male (BNHS 5976), (B) an adult male (SDBDU 2009.271) emerging from a tree hole, (C) frontal view of an adult male (BNHS 5977). D–H, A composite showing the breeding habitat of Frankixalus jerdonii. (D) Evergreen forest at Mawphlang in East Khasi Hills district of Meghalaya state, (E) close-up of a tree hole opening located 3.4 meters above the ground, (F) oviposition site with eggs adhered to the inner vertical walls of the tree hole above the water level, and arrow pointing towards an adult female found submerged about 1 cm below the water surface, (G) unpigmented gel-encapsulated eggs, (H) premetamorphic larva inside the water-filled tree hole.
Fig 3. Various life history stages of Frankixalus jerdonii.

(A) lateral, (B) dorsal, (C) ventral views of a preserved stage 36 tadpole, (D) unfertilised “nutritive” eggs found inside the dissected larval gut (mean diameter = 1.0 mm), (E) oral disc with papillae demarcating its margins, shown in frontal view of a stage 36 tadpole, (F) dextral vent tube, in ventral view of a stage 26 tadpole, (G) sinistral spiracular tube, in ventral view of a stage 36 tadpole, (H) gel-encapsulated eggs (mean diameter = 2.0 mm) found on the inside wall of a tree hole, (I) dorsolateral view of a stage 44 tadpole, (J) oral disc of a live stage 36 tadpole having a bifurcated muscular tongue, shown in frontal view, (K) dorsal, (L) ventral views of a live stage 35 tadpole, (M) serrated, inverted upper jaw of a stage 37 tadpole in ventral view, (N) serrated, V-shaped lower jaw of a stage 37 tadpole in ventral view.
Adult members of the new genus Frankixalus are morphologically most similar to members of the genus Kurixalus due to comparable adult snout-vent size, SVL 37.1–42.1 mm, male (N = 11); SVL 46.8 mm, female (N = 1) (Kurixalus SVL 23.0–43.0 mm, male; 25.0–45 mm, female [54,55]); basal webbing on fingers, moderate webbing between toes. The breeding ecology of Kurixalus also resembles Frankixalus, especially the type species Kurixalus eiffingeri, which lays eggs on the inner walls of tree hollows or bamboo stumps with small water accumulations [56]. Frankixalus can be differentiated from Kurixalus by the following morphological characters: snout not protruding (vs. protruding); outline of snout rather truncate in dorsal view, semi-circular in ventral view (vs. pointed to rounded), dermal fringes absent (vs. dermal fringes present on forearm and tarsus [57–59] (S2 Fig). The differences between these genera are however most noticeable in osteology and tadpole morphology.
Adult osteology (Fig 4A–4E): Tri-radiate extension of the sphenethmoid develops surpassing the posterior margins of the nasal capsules in Frankixalus (Fig 4C). However, this is not apparent in Gracixalus, Kurixalus and Philautus, where the anterior margin of the sphenethmoid ends in a concave shape without extending beyond the nasal capsules. Premaxillary teeth are present in Gracixalus, Philautus and Frankixalus, but appear to be absent in Kurixalus (Fig 4A). Pars dentalis (of premaxillae) of Kurixalus is slightly medially depressed, but straight in the other three. Frankixalus has trapezoidal shaped frontoparietals, which are not fused medially, a condition also observed in Gracixalus and Kurixalus, but half oval-shaped frontoparietals were observed in Philautus (Fig 4B). The parieto-squamosal arch was absent in all four, which was an important feature highlighted by Liem [42] in distinguishing rhacophorid genera.
Fig 4. Comparison of osteological characters of four closely-related generic type species: Gracixalus gracilipes (AMNH A163893, an adult female, SVL 29.6 mm, from Vietnam), Frankixalus jerdonii (SDBDU 2009.1163, an adult male, SVL 41.6 mm, from Mawphlang), Kurixalus eiffengeri (AMNH A14498, an adult female, SVL 33.0 mm, from Taiwan) and Philautus aurifasciatus (AMNH A24559, an adult female, SVL 22.2 mm, from Indonesia).

(A) shape of the premaxillae, (B) shape and development of frontoparietals, (C) structure of vomers, (D) ossified elements and shape of the pectoral girdles, (E) anatomy of the carpal elements. Abbreviations: FP, frontoparietal; OS, omosternum; PM, premaxilla; PP, prepollex; SP, sphenethmoid; ST, sternum; VM, vomer. All scale bars represent 2 mm.
The structure of the dentary process (= “vomerine odontophore”) [42] of the vomers and presence/absence of vomerine teeth also showed differences and similarities between the four genera (Fig 4C). Frankixalus and Kurixalus have dentary processes extended towards the mid portion of the sphenethmoid; contrastingly, Kurixalus have vomerine teeth (2/3 blunt teeth on each); Philautus and Gracixalus lack a vomerine dentary process as well as vomerine teeth.
Considering the differences of omosternum and sternum (= “Metasternum”, Liem [42]), Frankixalus is the only genus among these genera to have a cartilaginous epicoracoidal bridge at the base of the omosternum, giving it a forked appearance (Fig 4D). Philautus, Kurixalus and Gracixalus possess omosternums with unforked, ossified bases. In all four genera, the bony stylus of the sternum extends posteriorly and ends in cartilaginous dilated distal ends. This is wider than its length in Frankixalus and has an irregular margin. Comparatively, Kurixalus and Philautus have smaller sized, cartilaginous distal ends, but this is fork-shaped in Gracixalus (Fig 4D).
Carpals (= “carpale/carpal elements”, Liem [42]) show slight differences among the four species (Fig 4E). Radiale and centrale, having a similar structure, are not different among the four. However, the prehallux shows considerable differentiation in developing into a distal prehallical element in Gracixalus, Frankixalus and Philautus, but not in Kurixalus (Fig 4E).
Tadpole morphology (Fig 3A–3M): Tadpoles of Frankixalus with relatively long bodies are moderately depressed in a lateral profile, which is similar to Kurixalus [60]. Bulbous medium-sized eyes are placed dorsally in Frankixalus and dorsolaterally in Kurixalus. In tree hole dwelling Frankixalus, a sinistral tubular spiracle is located ventrally and is clearly distinguishable as a protrusion from the ventral body wall with the spiracular opening located closer to the vent tube than the snout. However, in Kurixalus, a laterally placed sinistral spiracle is present (no tubular portion) with a rounded aperture [60].
Oral discs of Frankixalus and Kurixalus are elliptical and demarcated by rows of papillae. A major feature that differentiates these two genera is the existence of tooth rows. On Frankixalus the oral disc lacks tooth rows, but Kurixalus possesses three to five rows, depending on its developmental stage [60]. Keratinized lower and upper jaw sheaths are present in both Frankixalus and Kurixalus. In both, the upper jaw sheath (upper beak) is inverted U-shaped. However, the V-shaped lower jaw sheath is medially separated in Frankixalus (Fig 3M and 3N).
The tadpole of Philautus (= Gracixalus) cf. carinensis described by Wassersug et al. [61] appears to be most similar to Frankixalus jerdonii tadpoles by external morphology, but differs from Frankixalus by its denticle rows I/O and upper row closer to the margin of upper lip.
Tadpole oophagy: The presence of eggs in Frankixalus jerdonii larval intestines suggests that these tadpoles are oophagous in nature (see detailed notes under Frankixalus jerdonii). Three rhacophorid species where oophagous tadpoles have been reported so far are Kurixalus eiffingeri [56,60,62–63], Philautus (= Gracixalus) cf. carinensis [61] and Rhacophorus vampyrus [64,65].
2. Description of the name-bearing type
Frankixalus jerdonii (Günther, 1876) comb. nov.
Common name
Jerdon’s tree frog
Comments
Günther [19] described Polypedates jerdonii from “Darjeeling”, West Bengal, India on the basis of two syntypes [(NHM 1947.2.7.84 (ex BMNH 72.4.17.189), and NHM 1947.2.7.85 (ex BMNH 72.4.17.190)]. Since this species was first described, no additional specimens have been reported. Dubois [26] designated NHM 1947.2.7.84 (ex BMNH 1872.4.17.189) as the lectotype, without a formal description. We studied this specimen but found it to be poorly preserved (S3 Fig). Our extensive field surveys in the type locality “Darjeeling” and vicinities were unsuccessful in finding this species from the place of its original description. New collections of the nominal taxon in the present study are from East Khasi Hills district (Meghalaya state), Churachandarpur district (Manipur state) and Kohima district (Nagaland state). These specimens are morphologically similar to the type series.
A detailed description of the lectotype NHM 1947.2.7.84 (ex BMNH 1872.4.17.189) is provided in S2A File and also included in the morphometric measurement table (S2 Table).
Name-bearing type
Lectotype, NHM 1947.2.7.84 (ex BMNH 72.4.17.189), an adult female [26].
Type locality
“Darjeeling”, West Bengal, India.
Material examined
Measurements of thirteen specimens, collected from three states of Northeast India as part of this study and two museum specimens from “Darjeeling” (including the lectotype) are provided in S2 Table and S2 File.
Genetic divergence
Pairwise comparisons of the sequenced mtDNA 16S rRNA gene fragments of Frankixalus jerdonii and Frankixalus sp. show average uncorrected genetic divergence of 8.9% (8.4–9.5%, N = 16). Average uncorrected intraspecific genetic distance within Frankixalus jerdonii populations was 0.8% (0–1.7%, N = 8).
Color in life
BNHS 5976: Dorsally reddish-brown with a dark brownish-black X-shaped marking on the back (Fig 2A); BNHS 5977: Dorsum brownish-grey with a pair of brownish-black concave stripes that extend from behind the eyes to the vent, uniting in the middle and forming an X-shaped marking on the back; loreal and tympanic region dark brownish-grey with a red tinge; broad dark stripe on sides of the head; flanks light yellow; fore and hind limbs light brownish-grey with brownish-black cross-bands; iris reddish-brown, encircled with thin light blue outer ring; ventral surface yellowish-brown (Fig 2C); SDBDU 2009.1163: dorsum with yellowish-grey stripe between the eyes, a light reddish-brown triangular marking on the snout; iris golden brown, encircled with thin blue outer ring.
Secondary sexual characters
Male (NHM 1947.2.7.85): nuptial pad present on finger I, yellowish-white in color, vocal sac present, a pair of internal gular slits near the base of the lower jaw. Female (NMH 1947.2.7.84): a tube-like dermal extension of the cloaca present, eggs creamy-white, unpigmented (diameter 1.8 ± 0.8 mm, N = 10).
Intraspecific variation
Measurements representing morphological variations among eight specimens from different localities are provided in S2 Table.
Geographic distribution
Frankixalus jerdonii is widely distributed in three Northeast Indian states (Meghalaya, Manipur and Nagaland), and in the “Darjeeling” region of West Bengal. Meghalaya: East Khasi Hills district, Wahlynkien (Marai Kaphon), Cherrapunjee (25°16.673'N, 91°43.075'E, 1337 m asl), and Mawphlang forest (25°26.292'N, 91°45.348'E, 1577 m asl); Manipur: Churachandarpur district, Zaraengtung, Raenghzaeng village (24°38.790'N, 93°42.983'E, 1392 m asl); Nagaland: Kohima district, Sechüma village, Zubza (25°41.333'N, 94°01.767'E, 1470 m asl), Meriema village (25°43.0'N, 94°05.25'E, 1425 m asl), Seukwehii, Tseminyu village (25°55.541'N, 94°13.066'E, 1340 m asl); West Bengal: Darjeeling district, “Darjeeling” (27°03.617'N, 88°15.667'E, 1600 m asl) (Fig 5).
3. Natural history and breeding ecology
All males in our study were found on arboreal vegetation in montane evergreen forest (Mawphlang, Meghalaya), or secondary forests (Zaraengtung, Manipur and Zubza, Nagaland); males from Zubza were collected from inside bamboo poles with slits. Breeding activities of Frankixalus jerdonii take place between May–August. Males of F. jerdonii were heard calling at night (between 18:00–22:00 hours) from tree holes located at heights ranging from 0.8–5.5 m at Mawphlang forest, Meghalaya (in June 2009) soon after sporadic rain showers. The habitat at this locality is composed of an evergreen forest with sparse undergrowth, consisting of scattered shrubs and herbs (Fig 2D). The trunks of hardwood trees in these montane evergreen forests usually have large growths of bryophytes (Fig 2E). Amplexus was not observed. Freshly laid egg clutches (unpigmented, gel-encapsulated) were found adhering on the inner walls of a tree hole about 5 m above the ground (Fig 2F and 2G). In total, nine nest sites were observed in tree hollows, with tree diameters of about 10–30 cm (measured at the height of the hole). Occupied tree holes had openings oriented both horizontally (N = 5) and vertically (N = 4), usually with narrow openings, and contained water that ranged in depth from about 5–50 cm (volume of water contained ranging from 30–160 ml, N = 3). A deep layer of organic debris was observed at the bottom of some tree holes, and at two nest sites a dormant male was found submerged under water (Fig 2F). When disturbed, one male tightly wedged itself into a crevice in the bottom of the hollow. At another nest site, a female (not collected) was observed submerged in water. Eggs were observed between 0.3–10 cm above the water surface, were round, diameter measuring 2.0 ± 0.1 mm, N = 18, with a thick jelly layer of about 0.2–0.4 mm. Clutch size varied from 16–30 eggs per mass (2.5–5.6 cm, N = 7). During repeated surveys at the same site between 27–29 June 2009, we also observed tadpoles of various sizes (stages 10–44) inside the water-filled tree holes (Fig 2H).
4. Oophagy in tadpoles
At Mawphlang forest, we recorded 4–15 tadpoles inside each tree hole (N = 4), either resting motionless at the surface or at the bottom, but rapidly surfacing and diving occasionally to apparently gulp air. On being disturbed, they immediately hid under debris at the bottom of the cavity. Close morphological examination of tadpoles collected from the tree holes (stages 34–44) showed that they contained 3–19 eggs inside their intestines. Two individuals were further studied by making a small ventral incision: one (stage 36) contained at least 13 eggs (Fig 3D), and the other (stage 43) contained 19 eggs within their guts. Several of the eggs inside the abdomen were found intact (N = 10) and seem to have been eaten shortly before tadpole collection.
Oophagy in phytotelm-breeding frogs is an adaptation to live in nutrient deficient environments; our study shows that Frankixalus jerdonii tadpoles are indeed oophagous in nature. However, there is no direct evidence for the fact that these eggs are conspecific i.e. field observations of a female laying unfertilized eggs in the tree holes with tadpoles were not made. Nevertheless, suggestive evidence is available from (i) the presence of eggs in larval guts (Fig 3C), and (ii) females possessing an extension of the cloacal skin (like a tube) that might facilitate egg laying one at a time [60]. Future field studies are needed to gain additional knowledge on this matter.
5. Tadpole morphology
Descriptions of Frankixalus jerdonii tadpoles are based on stage 36 (N = 4) (Fig 3) and are compared with tadpoles of Kurixalus. Frankixalus tadpoles show similarities with other rhacophorid exotrophic larvae in several aspects (well-developed tail musculature, depressed snouts, elongated bodies, dextral vent tubes), except for a few significant characters in oral morphology (see the section ‘comparison‘).
The body is relatively long (BL 31.4% of TL) with clearly distinguishable wider anterior region and a narrower posterior region, ventrally flattened in a lateral profile. Bulbous medium-sized eyes (ED 20.7% of BW) are placed dorsally, located close to one another with a medial separation of 26.4% of BW. Nasolacrimal duct and elygium are absent. Snout appears depressed. Non-perforated, laterally placed narial depressions are present, with pigmentation visible around the pits. The narial depressions are located closer to the eyes than to the snout. Sinistral spiracular tube is ventrally positioned. The myotomic muscle masses, which are divided by V shaped septa, construct the larval tail region. The tail has unequal tail membranes, extending to the tip of the tail, which is rounded. The well-developed tail musculature is placed near the narrow posterior part of the body, and extends almost up to the tail tip. The tail fins arise at the distal end of the body, widen medially, and narrow again towards the tip. A dextral, well-developed vent tube with a medial opening is present.
The oral disc is elliptical and is demarcated by rows of submarginal and marginal papillae. The marginal papillae are continuous and possess both short and long blunt papillae, whereas the long ones are more concentrated along the lower labium. Lateral emarginations are present and the demarcation is apparent due to the presence of dense small papillae on the lateral sides of the oral disc. Blunt submarginal papillae are small in size, observed in 2–3 rows, and appear continuous throughout the lower labium. Tooth rows are absent, but keratinized lower and upper jaw sheaths represent the keratinized structures of the oral disc. Both upper and lower jaws have serrated margins on their posterior and anterior margins, respectively.
Similar to the case of Kurixalus eiffingeri [60], we assume that large amounts of yolk provide nourishment to the tadpoles even after hatching, until they develop mouthparts that facilitate oophagy.
Measurements of Frankixalus jerdonii tadpoles (stage 36, N = 4) are as follows: BH, 3.22; BL, 9.81; BW, 5.52; ED, 1.14; NN, 2.30; NP, 1.16; PP, 2.60; RN, 2.16; SS, 6.32; SU, 10.83; SVL, 10.40; TL, 31.28; VT, 17.98; TMH, 2.44; TMW, 2.21. Preserved specimens are deposited in SDBDU.
6. Conservation
The major threat for amphibians in Northeast India is disturbance of primary and secondary forests by ‘jhumming’ (slash and burn) with the purpose of cultivating crops. Several localities where Frankixalus jerdonii is reported to occur are highly disturbed and fragmented habitats. The population at Cheerapunjee in Meghalaya state was recorded from a secondary forest adjacent to a highly polluted Wahlynkien stream, individuals from Manipur were from tree stumps within a jhum field, and those from Nagaland were from a secondary forest. These threats are alarming, especially for species that have very specialized habitat requirements, such as availability of small water collections in tree holes that are crucial for their survival and reproductive success.
Conclusion
Multiple lines of evidence from our study highlight the unique evolutionary position and life history features of Frankixalus. The description of this enigmatic lineage from the relatively unexplored northeast region of India not only emphasizes that part of this region’s biodiversity still remains poorly studied, but also underscores the need to replicate similar studies in other animal groups within this globally recognized biodiversity hotspot.
The description of this new rhacophorid genus adds to our knowledge on reproductive diversification in one of the most specious groups of neobatrachian amphibians. Such information is essential in understanding the evolution of reproductive strategies that allowed amphibians to occupy a broad variety of ecological niches (e.g., [11,30,52,66–69]).
Supporting Information
Numbers above and below the branches represent Bayesian Posterior Probabilities obtained for the nuclear and mitochondrial datasets, respectively.
(PDF)
A–C, K. eiffingeri (AMNH A14498, an adult female from Taiwan). (A) rounded in dorsal view, (B) rounded and protruding in ventral view, (C) rounded in lateral view; D–F, Frankixalus jerdonii (SDBDU 2009.1163). (D) truncate in dorsal view, (E) non-protruding semi-circular in ventral view, (F) vertical in lateral view.
(PDF)
(A) dorsal view, (B) ventral view, (C) lateral view of head, (D) ventral view of hand, (E) ventral view of foot, (F) schematic illustration of webbing on foot.
(PDF)
(A) Material and methods adult morphology, (B) Material and methods larval morphology, (C) Museum and people.
(DOC)
(A) Description of lectotype (all measurements in mm), (B) Specimens of Frankixalus jerdonii examined.
(DOC)
(DOC)
Status of specimens is given after the Museum number: LT- Lectotype, PL-Paralectotype, RS- Referred specimens.
(DOC)
Acknowledgments
The authors thank Yeong-Choy Kam and Chuang Ming-Feng (Tunghai University, Taiwan) and Jodi JL Rowley (Australian Museum) for sharing unpublished data on mouthparts of Kurixalus and Gracixalus species, respectively; David A Kizirian, Curatorial Associate, and Lauren Vonnahme, Scientific Assistant (AMNH, USA) for specimen loan; Nancy E Karraker and Yik Hei Sung for providing Liuixalus romeri images used in Fig 1; Mark Wilkinson and David Gower (NHM, London), K. Venkataraman (ZSI, Kolkata) for museum support; Darrel Frost (AMNH, USA) for fruitful discussions and advice; Tambor Lyngdoh, Robin Suyesh, Tengbat Sangma, Vetzo Koza, Gaigongdin Panmei, Jenghemlung Panmei, Landuanlung Kamei, Machunlung G Kamei, for assistance in field work and logistics; State Forest Departments of Arunachal Pradesh, Meghalaya, Manipur and Nagaland for providing study permissions; IVB is supported by a post-doctoral fellowship from FWO-Vlaanderen; MM is supported by the National Research Council of Sri Lanka (# NRC 11–124); SG is supported by a research fellowship from the Council for Scientific and Industrial Research, India (CSIR No. 9/45(1381)/2015-EMR-I); RGK is supported by Marie Skłodowska-Curie Fellowship (PIIF-GA-2013-625870).
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This study was supported by the following grants to SDB: DU/DST Purse Grant ‘2009/868’, Department of Science and Technology, Ministry of Science and Technology, Government of India; ‘BT/PR7833/NDB/51/153/2006’, Department of Biotechnology, Ministry of Science and Technology, Government of India; University of Delhi Research and Development Grants ‘2007/130’, ‘2007/858’, ‘2008/302’, ‘2014/110’. IVB is supported by a post-doctoral fellowship from FWO-Vlaanderen. MM is supported by the National Research Council of Sri Lanka (# NRC 11-124); SG is supported by a research fellowship from the Council for Scientific and Industrial Research, India (CSIR No. 9/45(1381)/2015-EMR-I); RGK is supported by Marie Skłodowska-Curie Fellowship (PIIF-GA-2013-625870). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Frost DR. Amphibian Species of the World: an Online Reference, Version 6.0. Electronic Database. 2015. Available: http://researchamnhorg/vz/herpetology/amphibia/.
- 2.AmphibiaWeb. Information on amphibian biology and conservation (web application). 2015. Berkeley, California: AmphibiaWeb. Available: http://amphibiaweb.org.
- 3.Delorme M, Dubois A, Grosjean S, Ohler A. Une nouvelle classification générique et subgénérique de la tribu des Philautini (Amphibia, Anura, Ranidae, Rhacophorinae). Bull Mens Soc Linn Lyon. 2005; 74(5):165–171. [Google Scholar]
- 4.Frost DR, Grant T, Faivovich J, Bain RH, Haas A, Haddad CFB, et al. The amphibian tree of life. Bull Am Mus Nat Hist. 2006; 297:1–370. 10.1206/0003-0090(2006)297[0001:TATOL]2.0.CO;2 [DOI] [Google Scholar]
- 5.Biju SD, Roelants K, Bossuyt F. Phylogenetic position of the montane treefrog Polypedates variabilis Jerdon, 1853 (Anura: Rhacophoridae), and description of a related species. Org Divers Evol. 2008; 8(4):267–276. 10.1016/j.ode.2007.11.004 [DOI] [Google Scholar]
- 6.Li JT, Che J, Bain RH, Zhao EM, Zhang YP. Molecular phylogeny of Rhacophoridae (Anura): A framework of taxonomic reassignment of species within the genera Aquixalus, Chiromantis, Rhacophorus and Philautus. Mol Phylogenet Evol. 2008; 48(1):302–312. 10.1016/j.ympev.2008.03.023 . [DOI] [PubMed] [Google Scholar]
- 7.Biju SD, Shouche YS, Dubois A, Dutta SK, Bossuyt F. A ground-dwelling rhacophorid frog from the highest mountain peak of the Western Ghats of India. Curr Sci. 2010; 98(8):1119–1125. [Google Scholar]
- 8.Meegaskumbura M, Meegaskumbura S, Bowatte G, Manamendra-Arachchi K, Pethiyagoda R, Hanken J, et al. Taruga (Anura: Rhacophoridae), a new genus of foam-nesting tree frogs endemic to Sri Lanka. Ceylon J Sci Biol Sci. 2010; 39(2):75–94. 10.4038/cjsbs.v39i2.2995 [DOI] [Google Scholar]
- 9.Abraham RK, Pyron RA, Ansil BR, Zachariah A, Zachariah A. Two novel genera and one new species of treefrog (Anura: Rhacophoridae) highlight cryptic diversity in the Western Ghats of India. Zootaxa. 2013; 3640(2):177–189. 10.11646/zootaxa.3640.2.3. [DOI] [PubMed] [Google Scholar]
- 10.Li JT, Li Y, Murphy RW, Rao DQ, Zhang YP. Phylogenetic resolution and systematics of the Asian tree frogs, Rhacophorus (Rhacophoridae, Amphibia). Zool Scr. 2012; 41(6):557–570. 10.1111/j.1463-6409.2012.00557.x [DOI] [Google Scholar]
- 11.Hertwig ST, Das I, Schweizer M, Brown R, Haas A. Phylogenetic relationships of the Rhacophorus everetti-group and implications for the evolution of reproductive modes in Philautus (Amphibia: Anura: Rhacophoridae). Zool Scr. 2012; 41(1):29–46. 10.1111/j.1463-6409.2011.00499.x [DOI] [Google Scholar]
- 12.Yu G, Zang M, Yang J. Molecular evidence for taxonomy of Rhacophorus appendiculatus and Kurixalus species from northern Vietnam, with comments on systematics of Kurixalus and Gracixalus (Anura: Rhacophoridae). Biochem Syst Ecol. 2013; 47:31–37. 10.1016/j.bse.2012.09.023 [DOI] [Google Scholar]
- 13.Biju SD, Bossuyt F. Systematics and phylogeny of Philautus Gistel, 1848 (Anura, Rhacophoridae) in the Western Ghats of India, with descriptions of 12 new species. Zool J Linn Soc. 2009; 155(2):374–444. 10.1111/j.1096-3642.2008.00466.x [DOI] [Google Scholar]
- 14.Vijayakumar SP, Dinesh KP, Mrugank VP, Kartik S. Lineage delimitation and description of nine new species of bush frogs (Anura: Raorchestes, Rhacophoridae) from the Western Ghats Escarpment. Zootaxa. 2014; 3893(4):451–488. 10.11646/zootaxa.3893.4.1 [DOI] [PubMed] [Google Scholar]
- 15.Myers N, Mittermeier RA, Mittermeier CG, da Fonseca GAB, Kent J. Biodiversity hotspots for conservation priorities. Nature. 2000; 403(6772):853–858. 10.1038/35002501 [DOI] [PubMed] [Google Scholar]
- 16.Mittermeier RA, Gil PR, Hoffmann M, Pilgrim J, Brooks T, Mittermeier CG, et al. Hotspots Revisited Earth's Biologically Richest and Most Endangered Terrestrial Eco-regions Conservation International, Sierra Madre and University of Virginia; 2004. [Google Scholar]
- 17.Kamei RG, Mauro DS, Gower DJ, Bocxlaer IV, Sherratt E, Thomas A, et al. Discovery of a new family of amphibians from northeast India with ancient links to Africa. Proc R Soc Lond B Biol Sci. 2012; 279(1737):2396–2401. 10.1098/rspb.2012.0150 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kamei RG, Gower DJ, Wilkinson M, Biju SD. Systematics of the caecilian family Chikilidae (Amphibia: Gymnophiona) with the description of three new species of Chikila from northeast India. Zootaxa. 2013; 3666(4):401–435. 10.11646/zootaxa.3666.4.1 [DOI] [PubMed] [Google Scholar]
- 19.Günther ACLG. Third report on collections of Indian reptiles obtained by the British Museum. Proc Zool Soc. 1876; 1875:567–577. [Google Scholar]
- 20.Boulenger GA. Catalogue of the Batrachia Salientia s Ecaudata in the Collection of the British Museum. 2nd ed. London: Taylor and Francis; 1882. [Google Scholar]
- 21.Amphibia Ahl E., Anura III, Polypedatidae Das Tierreich 55. Leipzig und Berlin: Walter de Gruyter & Co.; 1931. [Google Scholar]
- 22.Wolf S. Revision der Untergattung Rhacophorus (ausschliesslich der Madagaskar-Formen). Raffles Bull Zool. 1936; 12:137–217. [Google Scholar]
- 23.Dutta SK. Amphibians of India and Sri Lanka (Checklist and Bibliography) Bhubaneswar: Odyssey Publishing House; 1997. [Google Scholar]
- 24.Das I, Dutta SK. Checklist of the amphibians of India, with English common names. Hamadryad. 1998; 23:63–68. [Google Scholar]
- 25.Sarkar A, Ray S. Fauna of Arunachal Pradesh In: State Fauna Series 13 (Part I). Calcutta: Zoological Survey of India; 2006. p. 285–316. [Google Scholar]
- 26.Dubois A. Miscellanea taxonomica batrachologica (I). Alytes. 1987; 5(1–2):7–95. [Google Scholar]
- 27.Bossuyt F, Dubois A. A review of the frog genus Philautus Gistel, 1848 (Amphibia, Anura, Ranidae, Rhacophorinae). Zeylanica. 2001; 6:1–112. [Google Scholar]
- 28.Sambrook J, Fritschi EF, Maniatis T. Molecular cloning: a laboratory manual, New York: Cold Spring Harbor Laboratory Press; 1989. [Google Scholar]
- 29.Bossuyt F, Milinkovitch MC. Convergent adaptive radiations in Madagascan and Asian ranid frogs reveal covariation between larval and adult traits. Proc Natl Acad Sci U S A. 2000; 97(12):6585–6590. 10.1073/pnas.97.12.6585 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Biju SD, Bossuyt F. New frog family from India reveals an ancient biogeographical link with the Seychelles. Nature. 2003; 425(6959):711–714. 10.1038/nature02019 [DOI] [PubMed] [Google Scholar]
- 31.Richards CM, Moore WS. A phylogeny for the African treefrog family Hyperoliidae based on mitochondrial rDNA. Mol Phylogenet Evol. 1996; 5(3):522–532. 10.1006/mpev.1996.0047 [DOI] [PubMed] [Google Scholar]
- 32.Simon C, Frati F, Beckenbach A, Crespi B, Liu H, Flook P. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann Entomol Soc Am. 1994; 87(6):651–701. 10.1093/aesa/87.6.651 [DOI] [Google Scholar]
- 33.Thompson JD. The clustal-x windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997; 25(24):4876–4882. 10.1093/nar/25.24.4876 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Maddison DR, Maddison WP. MacClade: Analysis of phylogeny and character evolution, version 4.0 Sunderland, Massachusetts: Sinauer Associates; 2000. [Google Scholar]
- 35.Stamatakis A, Hoover P, Rougemont J. A rapid bootstrap algorithm for the RAxML Web Servers. Syst Biol. 2008; 57(5):758–771. 10.1080/10635150802429642 [DOI] [PubMed] [Google Scholar]
- 36.Miller MA, Pfeiffer W, Schwartz T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees Gateway Computing Environments Workshop (GCE), New Orleans, LA; 2010. 10.1109/GCE.2010.5676129 [DOI] [Google Scholar]
- 37.Ronquist F, Teslenko M, van der Mark P, Ayres DL, Darling A, Höhna S, et al. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst Biol. 2012; 61(3):539–542. 10.1093/sysbio/sys029 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Rambaut A, Suchard MA, Xie D, Drummond AJ. Tracer v1.6. Computer program and documentation distributed by the author. 2014. Available: http://beast.bio.ed.ac.uk/Tracer.
- 39.Biju SD, Garg S, Mahony S, Wijayathilaka N, Senevirathne G, Meegaskumbura M. DNA barcoding, phylogeny and systematics of Golden-backed frogs (Hylarana, Ranidae) of the Western Ghats-Sri Lanka biodiversity hotspot, with the description of seven new species. Contrib Zool. 2014; 83(4):269–335. http://www.contributionstozoology.nl/vol83/nr04/a04 [Google Scholar]
- 40.Savage JM, Heyer WR. Variation and distribution in the tree-frog genus Phyllomedusa in Costa Rica, Central America: With 6 figures. Stud Neotrop Fauna Environ. 1967; 5(2):111–131. [Google Scholar]
- 41.Myers CW, Duellman WE. A New Species of Hyla from Cerro Colorado, and other Tree frog records and geographical notes from Western Panama. Am Mus Novit. 1982; 2752:1–32. [Google Scholar]
- 42.Liem SS. The morphology, systematics, and evolution of the Old World treefrogs (Rhacophoridae and Hyperoliidae). Fieldiana Zool. 1970; 57:1–145. [Google Scholar]
- 43.Taylor WR, Van Dyke GC. Revised procedures for staining and clearing small fishes and other vertebrates for bone and cartilage study. Cybium. 1985; 9(2):107–119. [Google Scholar]
- 44.Trueb L. Bones, frogs, and evolution In: Vial JL, editor. Evolutionary biology of the anurans, contemporary research on major problems. Columbia, MO: University of Missouri Press; 1973. pp 65–132. [Google Scholar]
- 45.Duellman WE, Trueb L. Biology of the Amphibians New York: McGraw-Hill; 1986. [Google Scholar]
- 46.Maglia AM, Pugener LA, Mueller JM . Skeletal morphology and postmetamorphic ontogeny of Acris crepitans (Anura: Hylidae): A case of miniaturization in frogs. J Morphol. 2007; 268(3):194–223. 10.1002/jmor.10508 [DOI] [PubMed] [Google Scholar]
- 47.Pugener LA, Maglia AM. Skeletal morphogenesis of the vertebral column of the miniature hylid frog Acris crepitans, with comments on anomalies. J Morphol. 2009; 270(1):52–69. 10.1002/jmor.10665 [DOI] [PubMed] [Google Scholar]
- 48.Gosner KL. A simplified table for staging anuran embryos and larvae with notes on identification. Herpetologica. 1960; 16(3):183–190. [Google Scholar]
- 49.McDiarmid RW, Altig R. Tadpoles: The Biology of Anuran larvae Chicago: The University of Chicago Press; 1999. [Google Scholar]
- 50.Bowatte G, Meegaskumbura M. Morphology and ecology of tadpoles of Ramanella obscura (Anura: Microhylidae). Ceylon J Sci Biol Sci. 2011; 40(2):109–120. 10.4038/cjsbs.v40i2.3927 [DOI] [Google Scholar]
- 51.Li JT, Li Y, Klaus S, Rao DQ, Hillis DM, Zhang Y. Diversification of rhacophorid frogs provides evidence for accelerated faunal exchange between India and Eurasia during the Oligocene. Proc Natl Acad Sci U S A. 2013; 110(9):3441–3446. 10.1073/pnas.1300881110] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Meegaskumbura M, Senevirathne G, Biju SD, Garg S, Meegaskumbura S, Pethiyagoda R, et al. Patterns of reproductive-mode evolution in Old World tree frogs (Anura, Rhacophoridae). Zool Scr. 2015; 44(5):509–522. 10.1111/zsc.12121 [DOI] [Google Scholar]
- 53.Li JT, Che J, Murphy RW, Zhao H, Zhao EM, Rao DQ, et al. New insights to the molecular phylogenetics and generic assessment in the Rhacophoridae (Amphibia: Anura) based on five nuclear and three mitochondrial genes, with comments on the evolution of reproduction. Mol Phylogenet Evol. 2009; 53(2):509–522. 10.1016/j.ympev.2009.06.023 [DOI] [PubMed] [Google Scholar]
- 54.Nguyen TT, Matsui M, Duc HM. A new tree frog of the genus Kurixalus (Anura: Rhacophoridae) from Vietnam. Curr Herpetol. 2014; 33(2):101–111. 10.5358/hsj.33.101 [DOI] [Google Scholar]
- 55.Nguyen TT, Matsui M, Eto K. Mitochondrial phylogeny of an Asian tree frog genus Theloderma (Anura: Rhacophoridae). Mol Phylogenet Evol. 2015; 85:59–67. 10.1016/j.ympev.2015.02.003 [DOI] [PubMed] [Google Scholar]
- 56.Kam YC, Chuang ZS, Yen CF. Reproduction, oviposition-site selection, and tadpole oophagy of an arboreal nester, Chirixalus eiffingeri (Rhacophoridae), from Taiwan. J Herpetol. 1996; 30(1):52–59. 10.2307/1564706 [DOI] [Google Scholar]
- 57.Taylor EH. The amphibian fauna of Thailand. University of Kansas Science Bulletin. 1962; 43(1):255–599. [Google Scholar]
- 58.Matsui M, Orlov NL. A new species of Chirixalus from Vietnam (Anura: Rhacophoridae). Zoolog Sci. 2004; 21(6):671–676. [DOI] [PubMed] [Google Scholar]
- 59.Yu G, Zhang M, Yang J. A species boundary within the Chinese Kurixalus odontotarsus species group (Anura: Rhacophoridae): New insights from molecular evidence. Mol Phylogenet Evol. 2010; 56(3):942–950. 10.1016/j.ympev.2010.05.008 [DOI] [PubMed] [Google Scholar]
- 60.Kuramoto M, Wang C. A new rhacophorid treefrog from Taiwan, with comparisons to Chirixalus eiffingeri (Anura, Rhacophoridae). Copeia. 1987; 1987(4):931–942. [Google Scholar]
- 61.Wassersug RJ, Frogner KJ, Inger RF. Adaptations for life in tree holes by rhacophorid tadpoles from Thailand. J Herpetol. 1981; 15(1):41–52. 10.2307/1563645 [DOI] [Google Scholar]
- 62.Chiu CT, Kam YC. Growth of oophagous tadpoles (Chirixalus eiffingeri: Rhacophoridae) after nest displacement: implications for materal care and nest homing. Behaviour. 2006; 143(1):123–139. 10.1163/156853906775133605 [DOI] [Google Scholar]
- 63.Chen YH, Su YJ, Lin YS, Kam YC. Inter- and intra clutch competition among oophagous tadpoles of the Taiwanese tree frog, Chirixalus eiffingeri (Anura: Rhacophoridae). Herpetologica. 2001; 57(4):438–448. [Google Scholar]
- 64.Rowley JJL, Tran DTA, Le DTT, Hoang HD, Altig R. The strangest tadpole: the oophagous, tree-hole dwelling tadpole of Rhacophorus vampyrus (Anura: Rhacophoridae) from Vietnam. J Nat Hist. 2012; 46(47–48): 2969–2978. 10.1080/00222933.2012.732622 [DOI] [Google Scholar]
- 65.Vassilieva AB, Galoyan EA, Poyarkov NA Jr. Rhacophorus vampyrus (Anura: Rhacophoridae) reproductive biology: a new type of oophagous tadpole in Asian Treefrogs. J Herpetol. 2013; 47(4):607–614. 10.1670/12-180 [DOI] [Google Scholar]
- 66.Haddad CFB, Prado CPA. Reproductive modes in frogs and their unexpected diversity in the Atlantic forest of Brazil. Bioscience. 2005; 55(3):207–217. 10.1641/0006-3568(2005)055[0207:RMIFAT]2.0.CO;2 [DOI] [Google Scholar]
- 67.Wells KD. The ecology and behavior of amphibians Chicago: The University of Chicago Press; 2007. [Google Scholar]
- 68.Grosjean S, Delorme M, Dubois A, Ohler A. Evolution of reproduction in the Rhacophoridae (Amphibia, Anura). J Zool Syst Evol Res. 2008; 46(2):169–176. 10.1111/j.1439-0469.2007.00451.x [DOI] [Google Scholar]
- 69.Gomez-Mestre I, Pyron RA, Wiens JJ. Phylogenetic analyses reveal unexpected patterns in the evolution of reproductive modes in frogs. Evolution. 2012; 66(12):3687–3700. 10.1111/j.1558-5646.2012.01715.x [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Numbers above and below the branches represent Bayesian Posterior Probabilities obtained for the nuclear and mitochondrial datasets, respectively.
(PDF)
A–C, K. eiffingeri (AMNH A14498, an adult female from Taiwan). (A) rounded in dorsal view, (B) rounded and protruding in ventral view, (C) rounded in lateral view; D–F, Frankixalus jerdonii (SDBDU 2009.1163). (D) truncate in dorsal view, (E) non-protruding semi-circular in ventral view, (F) vertical in lateral view.
(PDF)
(A) dorsal view, (B) ventral view, (C) lateral view of head, (D) ventral view of hand, (E) ventral view of foot, (F) schematic illustration of webbing on foot.
(PDF)
(A) Material and methods adult morphology, (B) Material and methods larval morphology, (C) Museum and people.
(DOC)
(A) Description of lectotype (all measurements in mm), (B) Specimens of Frankixalus jerdonii examined.
(DOC)
(DOC)
Status of specimens is given after the Museum number: LT- Lectotype, PL-Paralectotype, RS- Referred specimens.
(DOC)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.