

Abstract
AIM: To investigate the ability of histamine to modulate tryptase release from human colon mast cells and the potential mechanisms.
METHODS: Enzymatically dispersed cells from human colons were challenged with histamine, anti-IgE or calcium ionophore A23187 (CI), and the cell supernatants after challenge were collected. Tryptase release was determined with a sandwich ELISA procedure.
RESULTS: Histamine at concentrations from 1 ng/mL was able to induce a “bell” shape dose related release of tryptase from colon mast cells. The maximum release of tryptase was approximately 3.5 fold more than spontaneous release. As little as 10 ng/mL histamine showed a similar potency to 10 μg/mL anti-IgE in induction of tryptase release. Histamine induced release of tryptase initiated at 10 s when histamine (100 ng/mL) was added to cells, gradually increased thereafter, and completed at 5 min. Both pertussis toxin or metabolic inhibitors were able to inhibit histamine induced tryptase release. When histamine and anti-IgE were added to colon mast cells at the same time, the quantity of tryptase released was similar to that induced by anti-IgE alone. The similar results were observed with CI. However, when various concentrations of histamine were incubated with cells for 20 min before adding anti-IgE or CI, the quantity of tryptase released was similar to that was induced by histamine alone.
CONCLUSION: Histamine is a potent activator of human colon mast cells, which represents a novel and pivotal self-amplification mechanism of mast cell degranulation.
INTRODUCTION
It has been reported that mast cells and their inflammatory mediators are closely associated to a number of intestinal diseases including idiopathic inflammatory bowel disease[1], chronic ulcerative colitis[2], Crohn’s disease[3] and collagenous colitis[4]. Through release their proinflammatory mediators including histamine, tryptase, chymase, heparin and some cytokines[5], mast cells actively participate in the pathogenesis of these intestinal diseases.
Tryptase is a tetrameric serine proteinase that constitutes some 20% of the total protein within human mast cells and is stored almost exclusively in the secretory granules of mast cells[6] in a catalytically active form[7]. Relatively higher secretion of tryptase has been detected in ulcerative colitis[8], implicating that this mediator is involved in the pathogenesis of intestinal diseases. Evidence is emerging that tryptase may be a key mediator of allergic inflammation and a promising target for therapeutic intervention[9] as it has been found to be able to induce microvascular leakage in the skin of guinea pig[10], bronchoconstriction[11] in allergic sheep airways, inflammatory cell accumulation in peritoneum of mouse[12] and release of IL-8 from epithelial cells[13]. Moreover, tryptase has long been recognised as a marker of mast cells[14,15], and an indicator of mast cell degranulation in vivo[16,17]. However, the application of tryptase as a unique marker of mast cell degranulation in mast cell challenge study in vitro was only started recently[18]. This was largely due to the lack of adequate assay to detect this mast cell product.
Histamine, on the other hand, has been widely employed as a marker of mast cell degranulation in mast cell challenge studies over the last four decades. But as a activator of mast cells it has hardly been examined. Since increased levels of histamine or enhanced histamine metabolism have been observed in collagenous colitis, food allergy[19], Crohn’s disease[20], ulcerative colitis[20,21] and allergic enteropathy[21] , this proinflammatory mediator is likely to participate in the pathogenesis of these diseases. In the current study, we investigated the potential of histamine to activate human colon mast cells in vitro in order to understand further the role of histamine in inflammatory bowel diseases.
MATERIALS AND METHODS
Reagents
The following compounds were purchased from Sigma (St. Louis, Mo., USA): CI, histamine dihydrochloride, collagenase (type I), hyaluronidase (type I), antimycin A, 2-deoxy-D-glucose, pertussis toxin, bovine serum albumin (BSA, fraction V), penicillin and streptomycin, extravidin peroxidase, o-phenylene diamine, biotin conjugate sheep anti-mouse immunoglobulins, extr-avidin peroxidase. Minimum essential medium (MEM) containing 25 mM N-2-hydroxylethylpiperazine-N’-2-ethane sulphonic acid (HEPES) and FCS was from Gibco (Paisley, Renfrewshire, UK). Goat anti-human IgE (inactivated) was from Serotec (Kidlington, Oxford, UK). A polyclonal antibody and a monoclonal antibody against tryptase were donated by Dr. Andrew F. Walls (University of Southampton, UK). HEPES and all other chemicals were of analytical grade.
Dispersion of mast cells
The mast cell dispersion procedure was similar to that described previously[21,22]. Human colon tissue was obtained from patients with carcinoma of colon at colectomy. Only macroscopically normal tissue was used for the study. After removal of fat, the tissue was washed and chopped finely with scissors into fragments of 0.5-2.0 mm3, and then incubated with 1.5 mg/mL collagenase and 0.75 mg/mL hyaluronidase in MEM containing 2% fetal calf serum (1 g colon/10 mL buffer) for 70 min at 37 °C. Dispersed cells were separated from the undigested tissue by filtration through a nylon gauze (pore size 100 µm diameter), washed and maintained in MEM (containing 10% FCS, 200 U/mL penicillin, 200 µg/mL streptomycin) on a roller overnight at room temperature. Mast cell purity, as determined by light microscopy after staining by Alcine blue, ranged from 3.6% to 5.8%.
Mast cell challenge
Dispersed cells were resuspended in HEPES buffered salt solution (HBSS, pH 7.4) with 1.8 mM CaCl2 and 0.5 mM MgCl2 (complete HBSS), and 100 µL aliquots containing 4-6 × 103 mast cells was added to 50 µL histamine, anti-IgE, CI or buffer alone and incubated for 15 min at 37 °C. The reaction was terminated by the addition of 150 µL ice cold incomplete HBSS and the tubes were centrifuged immediately (500 g, 10 min, 4 °C). All experiments were performed in duplicate. For the experiments with pertussis toxin, cells were incubated with 0.1 or 1.0 µg/mL pertussis toxin for four hours at 37 °C, and then washed with HBSS before adding stimulus. Similarly, for the experiments with metabolic inhibitors, cells were incubated with 2-deoxy-D-glucose (10 mmol/L) and antimycin A (1 µmol/L) for 40 min at 37 °C before challenged with stimulus. Supernatants were stored at -20 °C until use. As reported previously that 10 µg/mL anti-IgE and 1 µg/mL CI were the optimal concentrations for the induction of tryptase released from human colon mast cells[18], therefore they were selected as the standard concentrations throughout the study.
Tryptase measurement
Tryptase concentrations were measured using a sandwich ELISA procedure with a specific polyclonal antibody against human tryptase as the capture antibody and AA5 a monoclonal antibody specific for human tryptase as the detecting antibody[23].
Statistical analysis
Data were shown as mean ± SEM for the number of experiments (n) indicated, and the paired Student’s t test was applied to evaluate two independent samples. In all analyses, P < 0.05 was taken as statistically significant.
RESULTS
Induction of tryptase release by histamine
Histamine at the concentration of 1 ng/mL was able to induce a ‘bell’ shape dose related release of tryptase from colon mast cells. The maximal release of tryptase was approximately 3.5 fold more than spontaneous release provoked by 100 ng/mL histamine. Relatively less tryptase was released when 1000 ng/mL and 10000 ng/mL histamine were incubated with mast cells. As little as 10 ng/ mL histamine showed a similar potency to 10 μg/mL anti-IgE in induction of tryptase release from colon mast cells (Figure 1).
Figure 1.
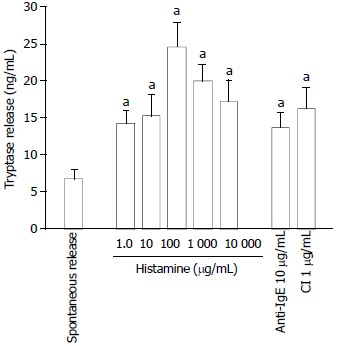
Histamine, anti-IgE and calcium ionophore (CI) in-duced tryptase release from colon mast cells. The values shown are mean ± SEM for six separate experiments. Stimulus or HBSS alone was incubated with cells for 15 min before termination of the reactions. aP < 0.05 compared with spontaneous release group (paired Student’s t test).
Time course for histamine induced tryptase release
Histamine induced release of tryptase initiated at 10 s when histamine (100 ng/mL) was added to cells, gradually increased thereafter, and completed at 5 min (Figure 2).
Figure 2.

Time course for histamine (100 ng/mL) induced re-lease of tryptase from colon mast cells. Data shown are mean ± SEM of five separate experiments.
Effects of pertussis toxin and metabolic inhibitors on histamine induced tryptase release
Tryptase release induced by histamine was reduced to the baseline level by pretreatment of colon mast cells with pertussis toxin or metabolic inhibitors. The same treatment was also able to slightly decrease the spontaneous tryptase release from mast cells (Table 1).
Table 1.
Effects of pertussis toxin (1.0 μg/mL) and metabolic inhibitors on histamine induced tryptase release from colon mast cells
| Treatment |
Tryptase release (ng/mL) |
||
| Hist | Hist | Buffer alone | |
| 100 μg/mL | 1000 μg/mL | ||
| Untreated | 32 ± 5.5 | 30.5 ± 3.9 | 20.5 ± 2.8 |
| Pertussis toxin | 17.1 ± 1.8a | 16.7 ± 1.7a | 17.6 ± 2.7 |
| Metabolic inhibitors | 18.8 ± 3.5a | 17.5 ± 1.9a | 16.3 ± 1.9 |
The values shown are mean ± SEM of five separate experiments performed in duplicate. aP < 0.05 compared with the untreated group (paired Student’s t test). Metabolic inhibitors = 1 mM antimycin A + 10 mM 2-deoxy-D-glucose. Hist = histamine.
Effect of histamine on anti-IgE and CI induced tryptase release
When 100 ng/mL or 1000 ng/mL histamine and anti-IgE were added to colon mast cells at the same time, tryptase release was significantly less than that induced by addition of the corresponding concentration of histamine alone. The similar results were observed when the same concentrations of histamine were added to cells at the same time with CI, though there was no significant difference between histamine alone and histamine plus CI (Figure 3). However, addition of various concentrations of histamine to colon mast cells 20 min before placing anti-IgE or CI, tryptase release was similar to that induced by the corresponding histamine alone, except for 10 ng/mL histamine with CI, indicating a synergistic action between them (Figure 4).
Figure 3.

Effect of co-addition of histamine (hist) and anti-IgE or calcium ionophore (CI) on tryptase release from colon mast cells. The values shown are mean ± SEM for six separate experiments. Various concentrations of histamine and anti-IgE or CI were added to cells at the same time, and then incubated with cells for 15 min before termination of the reactions. aP < 0.05 compared with the corresponding histamine alone group (paired Student’s t test).
Figure 4.

Effect of histamine (hist) on anti-IgE or calcium iono-phore (CI) induced tryptase release from colon mast cells. The values shown are mean ± SEM for six separate experiments. Various concentrations of histamine were incubated with cells for 20 min before addition of anti-IgE or CI. aP < 0.05 compared with the corresponding histamine alone group (paired Student’s t test). Pred.hist + anti-IgE or CI = the sum of tryptase release induced by histamine alone and anti-IgE or CI alone.
DISCUSSION
The finding that histamine is a potent activator of human colon mast cells demonstrated a novel and pivotal self-amplification mechanism of human mast cell degranulation, that is mast cells release histamine upon degranulation, and then the released histamine activates the adjacent mast cells. Thus, there are at least two self-amplification mechanisms in human mast cells upon degranulation, including tryptase and PAR-2 feedback process previously reported[22,23] and histamine inducing tryptase release.
Histamine appeared a potent secretogogue of tryptase, as little as 10 ng/mL histamine could share a similar potency of 10 μg/mL anti-IgE. This was a surprising result, but it clearly demonstrate a novel self-amplification mechanism of mast cell degranulation as the concentration of histamine inside the mast cell granules was estimated over 100 mmol/mL[24]. Interestingly, histamine concentration at 1000 ng/mL or above was able to induce less tryptase release from mast cells than 100 ng/mL, which may represent a novel self-protection mechanism of mast cells. Histamine induced tryptase release started slower than that elicited by anti-IgE, CI[18] or tc- LIGRLO[23], but completed 1 min earlier than that induced by anti-IgE and CI, indicating that it has its own activation-degranulation passway in mast cells. Pretreatment of cells with metabolic inhibitors antimycin A which blocks the oxidative phosphorylation process of cells, and 2-deoxy-D-glucose which blocks anaerobic metabolism pathway in cells, abolished the action of histamine, indicating that histamine induced release of tryptase is a non-cytotoxic process depending on cell energy supply. Tryptase release provoked by histamine was also inhibited by pretreatment of cells with pertussis toxin, suggesting that the degranulation process is associated with the activation of G-protein coupled receptors[25].
When histamine and anti-IgE were placed to mast cells at the same time, the quantity of released tryptase was similar to that elicited by anti-IgE alone, much less than that provoked by histamine alone, implicating that tryptase released from colon mast cells was mainly induced by anti-IgE, rather than by histamine. The reason for this was that the initiation of anti-IgE induced tryptase release was quicker than that induced by histamine, and mast cells only accepted one type of stimulation at a time[26,27]. Similar phenomena were observed with CI, and CI provoked mast cell degranulation apparently faster than that induced by histamine. In contrast, addition of various concentrations of histamine to colon mast cells 20 min before placing anti-IgE or CI, tryptase release was similar to that induced by the corresponding histamine alone, which proved further that colon mast cells were only able to respond to one optimal activation at a time, and the desensitized mast cells did not respond to any further stimulation. This behavior is an effective self-protection mechanism of mast cells. However, a synergistic action between 10 ng/mL histamine and CI, but not anti-IgE was observed, suggesting that mast cells experiencing a non-optimal stimulation may still have ability to respond to a further stimulation. In conclusion, histamine is a potent activator of human colon mast cells, and represents a novel and pivotal self-amplification mechanism of mast cell degranulation.
Footnotes
Supported by the National Natural Science Foundation of China, No. 30140023, and the Li Ka Shing Foundation, Hong Kong, China, No. C0200001
Edited by Wang XL
References
- 1.Fox CC, Lichtenstein LM, Roche JK. Intestinal mast cell responses in idiopathic inflammatory bowel disease. Histamine release from human intestinal mast cells in response to gut epithelial proteins. Dig Dis Sci. 1993;38:1105–1112. doi: 10.1007/BF01295728. [DOI] [PubMed] [Google Scholar]
- 2.Stoyanova II, Gulubova MV. Mast cells and inflammatory mediators in chronic ulcerative colitis. Acta Histochem. 2002;104:185–192. doi: 10.1078/0065-1281-00641. [DOI] [PubMed] [Google Scholar]
- 3.Nishida Y, Murase K, Isomoto H, Furusu H, Mizuta Y, Riddell RH, Kohno S. Different distribution of mast cells and macrophages in colonic mucosa of patients with collagenous colitis and inflammatory bowel disease. Hepatogastroenterology. 2002;49:678–682. [PubMed] [Google Scholar]
- 4.Schwab D, Raithel M, Hahn EG. Evidence for mast cell activation in collagenous colitis. Inflamm Res. 1998;47 Suppl 1:S64–S65. doi: 10.1007/s000110050276. [DOI] [PubMed] [Google Scholar]
- 5.Walls AF, He SH, Buckley MG, McEuen AR. Roles of the mast cell and basophil in asthma. Clin Exp Allergy Rev. 2001;1:68–72. [Google Scholar]
- 6.Abraham WM. Tryptase: potential role in airway inflammation and remodeling. Am J Physiol Lung Cell Mol Physiol. 2002;282:L193–L196. doi: 10.1152/ajplung.00429.2001. [DOI] [PubMed] [Google Scholar]
- 7.McEuen AR, He S, Brander ML, Walls AF. Guinea pig lung tryptase. Localisation to mast cells and characterisation of the partially purified enzyme. Biochem Pharmacol. 1996;52:331–340. doi: 10.1016/0006-2952(96)00211-0. [DOI] [PubMed] [Google Scholar]
- 8.Raithel M, Winterkamp S, Pacurar A, Ulrich P, Hochberger J, Hahn EG. Release of mast cell tryptase from human colorectal mucosa in inflammatory bowel disease. Scand J Gastroenterol. 2001;36:174–179. doi: 10.1080/003655201750065933. [DOI] [PubMed] [Google Scholar]
- 9.Walls AF. Structure and function of human mast cell tryptase. In: Marone G, editor. Mast cells and basophils. New York: Academic Press; 2000. pp. p. 291–309. [Google Scholar]
- 10.He S, Walls AF. Human mast cell tryptase: a stimulus of microvascular leakage and mast cell activation. Eur J Pharmacol. 1997;328:89–97. doi: 10.1016/s0014-2999(97)83033-6. [DOI] [PubMed] [Google Scholar]
- 11.Molinari JF, Scuri M, Moore WR, Clark J, Tanaka R, Abraham WM. Inhaled tryptase causes bronchoconstriction in sheep via histamine release. Am J Respir Crit Care Med. 1996;154:649–653. doi: 10.1164/ajrccm.154.3.8810600. [DOI] [PubMed] [Google Scholar]
- 12.He S, Peng Q, Walls AF. Potent induction of a neutrophil and eosinophil-rich infiltrate in vivo by human mast cell tryptase: selective enhancement of eosinophil recruitment by histamine. J Immunol. 1997;159:6216–6225. [PubMed] [Google Scholar]
- 13.Cairns JA, Walls AF. Mast cell tryptase is a mitogen for epithelial cells. Stimulation of IL-8 production and intercellular adhesion molecule-1 expression. J Immunol. 1996;156:275–283. [PubMed] [Google Scholar]
- 14.Proud D, Bailey GS, Naclerio RM, Reynolds CJ, Cruz AA, Eggleston PA, Lichtenstein LM, Togias AG. Tryptase and histamine as markers to evaluate mast cell activation during the responses to nasal challenge with allergen, cold, dry air, and hyperosmolar solutions. J Allergy Clin Immunol. 1992;89:1098–1110. doi: 10.1016/0091-6749(92)90293-b. [DOI] [PubMed] [Google Scholar]
- 15.He SH, Li P, Buckley MG, Walls AF. Identification of mast cell subtypes by double labeling immunohistochemistry technique. Chin J Pathol. 2000;29:383–384. [Google Scholar]
- 16.Schwartz LB, Metcalfe DD, Miller JS, Earl H, Sullivan T. Tryptase levels as an indicator of mast-cell activation in systemic anaphylaxis and mastocytosis. N Engl J Med. 1987;316:1622–1626. doi: 10.1056/NEJM198706253162603. [DOI] [PubMed] [Google Scholar]
- 17.Laroche D, Vergnaud MC, Sillard B, Soufarapis H, Bricard H. Biochemical markers of anaphylactoid reactions to drugs. Comparison of plasma histamine and tryptase. Anesthesiology. 1991;75:945–949. doi: 10.1097/00000542-199112000-00004. [DOI] [PubMed] [Google Scholar]
- 18.He SH, Xie H, He YS. Induction of tryptase and histamine release from human colon mast cells by IgE dependent or independent mechanisms. World J Gastroenterol. 2004;10:319–322. doi: 10.3748/wjg.v10.i3.319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Schwab D, Hahn EG, Raithel M. Enhanced histamine metabolism: a comparative analysis of collagenous colitis and food allergy with respect to the role of diet and NSAID use. Inflamm Res. 2003;52:142–147. doi: 10.1007/s000110300064. [DOI] [PubMed] [Google Scholar]
- 20.Winterkamp S, Weidenhiller M, Otte P, Stolper J, Schwab D, Hahn EG, Raithel M. Urinary excretion of N-methylhistamine as a marker of disease activity in inflammatory bowel disease. Am J Gastroenterol. 2002;97:3071–3077. doi: 10.1111/j.1572-0241.2002.07028.x. [DOI] [PubMed] [Google Scholar]
- 21.Raithel M, Matek M, Baenkler HW, Jorde W, Hahn EG. Mucosal histamine content and histamine secretion in Crohn's disease, ulcerative colitis and allergic enteropathy. Int Arch Allergy Immunol. 1995;108:127–133. doi: 10.1159/000237129. [DOI] [PubMed] [Google Scholar]
- 22.He S, Gaça MD, Walls AF. A role for tryptase in the activation of human mast cells: modulation of histamine release by tryptase and inhibitors of tryptase. J Pharmacol Exp Ther. 1998;286:289–297. [PubMed] [Google Scholar]
- 23.He SH, He YS, Xie H. Activation of human colon mast cells through proteinase activated receptor-2. World J Gastroenterol. 2004;10:327–331. doi: 10.3748/wjg.v10.i3.327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Schwartz LB, Irani AM, Roller K, Castells MC, Schechter NM. Quantitation of histamine, tryptase, and chymase in dispersed human T and TC mast cells. J Immunol. 1987;138:2611–2615. [PubMed] [Google Scholar]
- 25.Piliponsky AM, Gleich GJ, Nagler A, Bar I, Levi-Schaffer F. Non-IgE-dependent activation of human lung- and cord blood-derived mast cells is induced by eosinophil major basic protein and modulated by the membrane form of stem cell factor. Blood. 2003;101:1898–1904. doi: 10.1182/blood-2002-05-1488. [DOI] [PubMed] [Google Scholar]
- 26.Shalit M, Levi-Schaffer F. Challenge of mast cells with increasing amounts of antigen induces desensitization. Clin Exp Allergy. 1995;25:896–902. doi: 10.1111/j.1365-2222.1995.tb00033.x. [DOI] [PubMed] [Google Scholar]
- 27.Rubinchik E, Shalit M, Levi-Schaffer F. Responsiveness of human skin mast cells to repeated activation: an in vitro study. Allergy. 1998;53:14–19. doi: 10.1111/j.1398-9995.1998.tb03768.x. [DOI] [PubMed] [Google Scholar]