

Abstract
The prevalence of psychiatric disorders which are characterized by cognitive decline is increasing at an alarming rate and account for a significant proportion of the global disease burden. Evidences from human and animal studies indicate that neurocognitive development is influenced by various environmental factors including nutrition. It has been established that nutrition affects the brain throughout life. However, the mechanisms through which nutrition modulates mental health are still not well understood. It has been suggested that the deficiencies of both vitamin B12 and omega-3 fatty acids can have adverse effects on cognition and synaptic plasticity. Studies indicate a need for supplementation of vitamin B12 and omega-3 fatty acids to reduce the risk of cognitive decline, although the results of intervention trials using these nutrients in isolation are inconclusive. In the present article, we provide an overview of vitamin B12 and omega-3 fatty acids, the possible mechanisms and the evidences through which vitamin B12 and omega-3 fatty acids modulate mental health and cognition. Understanding the role of vitamin B12 and omega-3 fatty acids on brain functioning may provide important clues to prevent early cognitive deficits and later neurobehavioral disorders.
Keywords: BDNF, Epigenetics, Homocysteine, Omega-3 fatty acids, One-carbon cycle, Vitamin B12
Background
The escalating prevalence of brain disorders is currently a global health challenge [1] and has emerged as leading contributors to global disease burden [2]. Brain disorders affect neurological and cognitive performance and therefore have lifelong devastating effects on the individual, family and society. However, the underlying causes of mental health problems are poorly understood. Substantial evidence suggests that cognitive impairment can be influenced by number of environmental factors such as nutrition [3]. Nutrition plays a key role in maintaining optimal brain health throughout the lifespan of an individual [4]. In view of this, the studies examining the link between nutrition and mental health have gained widespread attention in recent years.
Nutrients like B vitamins and omega-3 fatty acids have been widely studied in recent years in context of brain development and functioning. In developing countries like India, due to widespread vegetarianism, vitamin B12 deficiency [5] coexists with suboptimal levels of omega-3 fatty acids [6]. Reports suggested that these nutrients are critical for brain health and their deficiencies could influence cognitive performance adversely. It has also been suggested that vegetarians should increase their dietary intake of vitamin B12 and omega-3 fatty acids to reduce increased risk factors for non-communicable diseases [7]. However, the effects of both vitamin B12 and omega-3 fatty acids supplementation together on neurological disorders are relatively unexplored and the underlying mechanisms need to be established.
In this review, we highlight the role of vitamin B12 and omega-3 fatty acids on brain function and proposed mechanisms through which these nutrients influence mental health and cognition.
Review
Vitamin B12
Vitamin B12 is generally found only in foods of animal origin [8]. Thus, the population predominantly consuming a vegetarian diet is deficient in vitamin B12 [5, 9]. Vitamin B12 is a key micronutrient required for proper brain development and is associated with one carbon metabolism that plays a pivotal role in transmethylation reactions. It is involved in the formation of S-adenosylmethionine (SAM), which is an important substrate for epigenetic mechanisms [10]. Vitamin B12 is known to have fundamental roles in the brain function at all ages and also in the prevention of disorders of CNS development, mood disorders and dementias including Alzheimer’s disease and vascular dementia in elderly people [11].
Elevated methylmalonic acid and total homocysteine concentrations are considered as sensitive metabolic markers for vitamin B12 deficiency [12]. Myelopathy and neuropathy are known to be the main clinical manifestations of vitamin B12 deficiency [13]. Symptoms of vitamin B12 deficiency include megaloblastic anaemia, tingling and numbness of the extremities, gait abnormalities, visual disturbances, memory loss and dementia [10]. Reports indicate that low dietary intake of vitamin B12, especially in pregnant women and in the elderly population are associated with developmental and neurological disorders [14, 15].
Studies indicate a need for supplementation of vitamin B12 to improve pregnancy outcome and reduce the risk of neurodevelopmental disorders [9]. Reports indicate a positive association between maternal vitamin B12 status and cognition in the offspring [16]. In contrast, a study in Indian school children at 6–10 years of age found an inverse association of maternal vitamin B12 concentrations with cognitive performance [17]. Our recent animal study has shown that vitamin B12 supplementation (50 μg/kg of diet) was able to maintain the levels of docosahexaenoic acid (DHA) and brain derived neurotrophic factor (BDNF) in the hippocampus and cortex, and sustain cognition in the adult rat offspring as compared to control animals (receiving 25 μg/kg of vitamin B12 in diet) [18]. A review by van de Rest et al. concludes that there are limited studies examining the association of maternal vitamin B12 with cognition and results are inconsistent suggesting a need for more research in this area [19].
Omega-3 fatty acids
The role of omega-3 fatty acids especially DHA in brain development is gaining widespread attention [20]. The dietary sources of omega-3 fatty acids are fish and sea foods only [21] which are the rich sources of DHA. Hence, the vegetarian population particularly Asian Indians are found to be deficient in omega-3 fatty acids [22]. Further, over the past 150 years, the western diet has altered such that the ratio of omega-3 to omega-6 fatty acids has changed from 1:1 to 1:20–25 indicating that this diet is deficient in omega-3 fatty acids and is rich in omega-6 fatty acids [23]. Thus, the deficiency of omega-3 fatty acids and consumption of western diet has been suggested to be associated with cognitive impairment [24, 25].
There is increasing evidence which indicates the importance of omega-3 fatty acids in brain health across the lifespan [26]. DHA, which is the core member of omega-3 fatty acids, is highly concentrated in the brain and the outer segments of retinal rods and cones, constituting around 50 % of the total polyunsaturated fatty acids [27]. DHA participates in a number of neuronal processes including neurogenesis, neuroplasticity, neuron differentiation and survival, membrane integrity and fluidity [28]. A large body of evidence in animals has shown that maternal supplementation of DHA during gestation has neuroprotective effects against prenatal stress-induced brain dysfunction [29], hyperoxic injury [30] and hypoxic ischemic injury [31].
A recent study has reported an inverse association between intake of omega-3 fatty acids and depression [32]. DHA is well-implicated in synaptic transmission, synaptogenesis, learning and memory processes [33]. The density of dendritic spine has shown to be increased in the hippocampus of the animals with the oral supplementation of DHA [34]. Several evidences from animal and human studies have shown a positive association between DHA and cognitive development [35–38]. Thus, the role of omega-3 fatty acids in influencing brain health and wellbeing is well established however further investigation is required to better understand the underlying mechanisms and also to develop therapeutic targets for neurological disorders.
Possible mechanisms of the effects of vitamin B12 and omega-3 fatty acids on brain development
The combined deficiency of both vitamin B12 and omega-3 fatty acids could impair brain function and increase the risk for neurological and developmental disorders. The deficiency of both these nutrients can affect neural function by mechanisms [Fig. 1] discussed below.
Fig. 1.
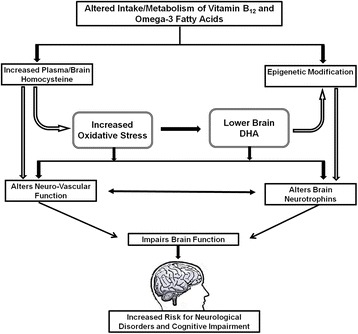
Proposed mechanisms through which vitamin B12 and omega-3 fatty acids affects brain functioning. Alerted intake/metabolism of vitamin B12 and omega-3 fatty acids affects brain function through multiple pathways. Increased homocysteine levels and altered epigenetic modification influences brain neurotrophins and neuro-vascular function directly or through increased oxidative stress and lower brain omega-3 fatty acid levels which may increase the risk for neurological disorders and cognitive impairment
Homocysteine- induced oxidative stress
Elevated levels of homocysteine have been associated with vitamin B12 deficiency [5]. Vitamin B12 is an important component of the one-carbon metabolism where it acts as a cofactor for the enzyme methionine synthase, which converts homocysteine to methionine [39]. Hence, if there is a deficiency of vitamin B12, remethylation of homocysteine via the enzyme methionine synthase is reduced and the plasma levels of homocysteine are elevated [40]. It has also been observed that elevated homocysteine levels are a sign of disturbed remethylation of homocysteine [41].
It has been reported that homocysteine activates glutamate receptors by acting as an agonist at the glutamate binding site of the N-methyl-D-aspartate receptor [42, 43]. Thus, overactivation of the glutamate receptors leads to increased intracellular calcium levels and further activation of signaling kinases resulting in neurodegeneration and neuronal damage through a process called excitotoxicity [44, 45]. Homocysteine also increases reactive oxygen species generation and induce neuronal DNA damage, triggering apoptosis and affects synaptic and glial function [46, 47].
It is known that the brain is highly susceptible to oxidative cellular damage due to high metabolic load and poor antioxidant defense system [48]. Reports suggest that omega-3 fatty acids are susceptible for degradation due to increased oxidative stress [49]. Our earlier animal study has demonstrated increased plasma homocysteine levels in the offspring as a consequence of maternal vitamin B12 deficiency [50]. Further, reduced plasma levels of vitamin B12 and DHA and increased homocysteine levels were also observed in schizophrenic patients [51] suggesting their role in the psychological abnormality underlying the disease. We have also observed a negative association between maternal plasma homocysteine and erythrocyte DHA levels in pregnancy complications like preeclampsia [52].
Hyperhomocysteinemia through the mediation of oxidative stress produces changes in structure and function of cerebral blood vessels [53]. Animal studies have also reported that high levels of homocysteine cause damage and leakage to hippocampal microvasculature [54, 55] leading to vascular remodeling which could disrupt the blood–brain barrier [56]. Reports have also suggested that homocysteine inhibits angiogenesis through the inhibition of vascular endothelial growth factor (VEGF) and its downstream signaling pathway as demonstrated in cultured human umbilical vein endothelial cells [57]. A recent study has demonstrated beneficial effects of omega-3 fatty acid supplementation against cerebral ischemia and has been shown to enhance cerebral angiogenesis [58]. Thus, supplementation of vitamin B12 and omega-3 fatty acids together may help to protect against homocysteine-induced adverse neurodegenerative effects.
Altered neurotrophins
Vitamin B12 has been implicated in the maintenance of equilibrium between neurotrophic and neurotoxic factors in the central nervous system [59]. Neurotrophins are growth factors that influence the proliferation, differentiation, survival and death of neuronal and non-neuronal cells. A series of our animal studies have demonstrated reduced levels of neurotrophins like NGF (nerve growth factor) and BDNF in the brain as a consequence of vitamin B12 deficiency [50, 60, 61]. The reduction in the levels of neurotrophins could be attributed to increased oxidative stress and decreased DHA levels [62].
Reduced levels of BDNF have been widely implicated in the pathophysiology of various psychiatric disorders like schizophrenia [63] Alzheimer’s [64] and Parkinson’s disease [65] and Huntington’s disease [66]. Studies have also reported lower serum NGF levels in the schizophrenic patients [67]. Lower levels of BDNF level in the schizophrenic patients has been associated with cognitive impairment [68]. It has been suggested that high neurotrophin expression in the brain may act as neuroprotective against neurological diseases [69].
Experimental evidence suggests that omega-3 fatty acids act as neuroprotective agent against neurological insults through the BDNF signaling pathways [70, 71]. It has been demonstrated that DHA supplementation in aged mice improved cognitive dysfunction through increased BDNF levels [72]. DHA is suggested to increase neurotrophins in the brain by increasing membrane fluidity, reducing oxidative stress, through neuroprotection D1 [38]. A recent study reported by us has demonstrated that combined supplementation of both vitamin B12 and omega-3 fatty acids together increases the levels of BDNF in the cortex and hippocampus region of the brain [18]. Thus, based on all above facts, altered neurotrophins and their downstream signaling pathway could be one of the possible mechanisms affected by the deficiency of vitamin B12 and omega-3 fatty acids.
Altered angiogenic factors
It has been demonstrated that neurotrophins like BDNF and NGF are involved in the regulation of angiogenic markers in the brain [73, 74]. Studies also indicate that neurotrophin activation of tyrosine kinase receptors stimulates an increase in vascular endothelial growth factor (VEGF) transcription in neuronal tissue [75]. VEGF plays a key role in promoting and coordinating angiogenesis during development and adulthood [76]. However, both in vitro and in vivo experiments indicated the diverse roles of VEGF-A in the brain including neuronal survival and migration [77]. The neurotrophin mediated increase in VEGF in neuronal cells is shown to be accompanied by an increase in the hypoxia inducible factor-1 alpha (HIF-1 alpha) levels which is dependent on tropomyosin receptor kinase (Trk)/ phosphoinositide 3-kinase (PI-3kinase)/ serine/threonine-specific protein kinase (AKT)/ mammalian target of rapamycin (mTOR) pathway [78]. HIF-1α expression is known to be regulated by the mTOR signaling pathway [79]. Activation of mTOR leads to the phosphorylation of two downstream effectors: ribosomal protein S6 kinase (p70S6K) and eukaryotic initiation factor 4E-binding protein-1 (4E-BP1) [80, 81]. Phosphorylation activates p70S6K and inactivates 4E-BP1 which in turn known to regulate HIF-1α expression at the translational level [79] (Fig. 2).
Fig. 2.
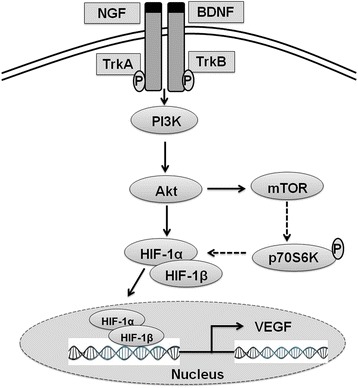
Interaction between neurotrophins and VEGF. Binding of neurotrophins like BDNF (brain derived growth factor) and NGF (nerve growth factor) to their respective receptors TrkB/A triggers PI-3kinase/AKT/mTOR pathway and leads to increased expression of HIF-1α protein expression. Activated mTOR leads to the phosphorylation and activation of ribosomal protein S6 kinase (p70S6K) which in turn known to regulate HIF-1α expression at the translational level. HIF-1α then dimerizes with HIF-1β to form HIF complex which translocate to the nucleus and binds to hypoxia response elements and leads to the increased transcription of VEGF (vascular endothelial growth factor) in the brain. NGF: Nerve Growth Factor: BDNF: Brain Derived Growth Factor; TrkB/TrkA: Tropomyosin receptor kinase B/A; PI3K: phosphoinositide 3-kinase; Akt: serine/threonine-specific protein kinase also called as protein kinase B; mTOR: mammalian target of rapamycin; HIF-1α: Hypoxia Inducible Factor-α; HIF-1β: Hypoxia Inducible Factor-β; p70S6K: ribosomal protein S6 kinase; VFGF: Vascular Endothelial Growth Factor
Thus, it is clear that there is an interaction between neurotrophins and VEGF in the brain. Our recent study demonstrates that maternal vitamin B12 and omega-3 fatty acids influence the levels and expression of VEGF and NGF in the pup brain [82].
Altered one-carbon metabolism and epigenetic regulation
The dysregulation of the one-carbon metabolism is well implicated in brain disorders like schizophrenia, bipolar disorder, autism and depression [83]. Vitamin B12 is important cofactor in one carbon cycle and is involved in the formation of S-adenosyl methionine (SAM). SAM is a universal methyl donor for important methylation reactions including methylation of DNA, neurotransmitters and phospholipids. Phospholipids utilize methyl groups for the conversion of phosphatidylethanolamine (PE) to phosphatidylcholine (PC). The conversion of PE to PC in biological membranes is critical for mobilization of DHA from liver to plasma and brain [84, 85]. A study in the patients of Alzheimer disease demonstrated that the high levels of circulating homocysteine and decreased mobilization of DHA from the liver into plasma and peripheral tissues may contribute to cerebrovascular and neurodegenerative changes [86].
The one-carbon metabolism is known to influence epigenetic modifications which in turn produce long-term changes in the brain affecting memory, learning, cognition and behavior [87]. Epigenetics induces changes in the chromatin without disrupting the basic DNA sequence [88]. DNA methylation is the most widely studied form of epigenetic modification which occurs through one-carbon metabolism. DNA methylation/demethylation plays an important role in learning and memory as suppression of DNA methylation has been associated with impaired long term potentiation [89] suggesting a critical role for epigenetic modifiers in neurodevelopment [90]. Studies have demonstrated an association of memory with changes in DNA methylation in the BDNF gene [91]. It has been demonstrated that DNA methylation can also control BDNF expression during development of the forebrain in mice [92].
It is reported that adequate supply of nutrients which are the source of methyl groups to the brain is necessary for proper functioning [93]. Vitamin B12 is an important modifier of epigenetics being involved as a cofactor in the one-carbon cycle. It has been indicated that omega-3 fatty acids are also known to influence epigenetic mechanisms regulating gene expression [94]. We have demonstrated altered global methylation patterns in the brain of the offspring as a consequence of imbalanced (excess folate and vitamin B12 deficient) maternal micronutrients in animals. We also demonstrate the important role of prenatal omega-3 fatty acids in reversing methylation patterns thereby highlighting its contribution in neuroprotection and cognition [95].
Studies suggest that the brain has significantly higher levels of methylated DNA in comparison to tissues like the placenta [96]. The presence of methylated CpG dinucleotides is specific for each brain region and for each stage of development [97]. In spite of this, limited studies have examined the association of nutrients, especially those that are part of the one carbon cycle, and methylation changes in the brain of the offspring.
Conclusion
Inadequate nutrition can increase the risk of developing neurodevelopmental and cognitive deficits. There are number of studies and reviews which have evaluated the neuroprotective benefits of vitamin B12 and omega-3 fatty acids in isolation, however, the combination of these nutrients are not reported. Thus, there is a need to study whether there exists any synergistic or antagonistic effects between these nutrients. Further research is recommended to investigate the optimal dose required to demonstrate preventive effects on cognitive function.
The current article discusses various mechanisms through which vitamin B12 and omega-3 fatty acids can support brain function. Initially, animal and cell-culture studies are recommended which will help to better understand the mechanisms involved. This may provide important insights into the etiology of various neurodevelopmental disorders.
Acknowledgement
The authors acknowledge Indian Council of Medical Research for awarding Fellowship to one of the authors (Richa Rathod) for pursuing her PhD work.
Abbreviations
- Akt
Protein kinase B
- BDNF
Brain derived neurotrophic factor
- CNS
Central nervous system
- DHA
Docosahexaenoic acid
- HIF-1 alpha
Hypoxia inducible factor-1 alpha
- mTOR
mammalian target of rapamycin
- NGF
Nerve growth factor
- PC
Phosphatidylcholine
- PE
Phosphatidylethanolamine
- PI3K
Phosphoinositide 3-kinase
- SAM
S-adenosylmethionine
- TrkB/TrkA
Tropomyosin receptor kinase B/A
- VEGF
Vascular endothelial growth factor
Footnotes
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
RR wrote the paper and prepared the figures, AK gave inputs for writing the paper and SJ conceptualized and revised the final version of the manuscript. All authors read and approved the final manuscript.
References
- 1.Wittchen HU, Jacobi F, Rehm J, Gustavsson A, Svensson M, Jönsson B, et al. The size and burden of mental disorders and other disorders of the brain in Europe 2010. Eur Neuropsychopharmacol. 2011;21:655–79. doi: 10.1016/j.euroneuro.2011.07.018. [DOI] [PubMed] [Google Scholar]
- 2.Baxter AJ, Patton G, Scott KM, Degenhardt L, Whiteford HA. Global epidemiology of mental disorders: what are we missing? PLoS One. 2013;8 doi: 10.1371/journal.pone.0065514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Nyaradi A, Li J, Hickling S, Foste J, Oddy WH. The role of nutrition in children’s neurocognitive development, from pregnancy through childhood. Front Hum Neurosci. 2011;7:97. doi: 10.3389/fnhum.2013.00097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Dauncey MJ. Genomic and epigenomic insights into nutrition and brain disorders. Nutrients. 2013;5:887–914. doi: 10.3390/nu5030887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Yajnik CS, Deshpande SS, Lubree HG, Naik SS, Bhat DS, Uradey BS, et al. Vitamin B12 deficiency and hyperhomocysteinemia in rural and urban Indians. J Assoc Physicians India. 2006;54:775–82. [PubMed] [Google Scholar]
- 6.Muthayya S, Dwarkanath P, Thomas T, Ramprakash S, Mehra R, Mhaskar A, et al. The effect of fish and omega-3 LCPUFA intake on low birth weight in Indian pregnant women. Eur J Clin Nutr. 2009;63:340–6. doi: 10.1038/sj.ejcn.1602933. [DOI] [PubMed] [Google Scholar]
- 7.Li D. Effect of the vegetarian diet on non-communicable diseases. J Sci Food Agric. 2014;941:169–73. doi: 10.1002/jsfa.6362. [DOI] [PubMed] [Google Scholar]
- 8.Simpson JL, Bailey LB, Pietrzik K, Shane B, Holzgreve W. Micronutrients and women of reproductive potential: required dietary intake and consequences of dietary deficiency or excess. Part I-Folate, Vitamin B12, Vitamin B6. J Matern Fetal Neonatal Med. 2010;23:1323–43. doi: 10.3109/14767051003678234. [DOI] [PubMed] [Google Scholar]
- 9.Pawlak R, Parrott SJ, Raj S, Cullum-Dugan D, Lucus D. How prevalent is vitamin B (12) deficiency among vegetarians? Nutr Rev. 2013;71:110–7. doi: 10.1111/nure.12001. [DOI] [PubMed] [Google Scholar]
- 10.Gröber U, Kisters K, Schmidt J. Neuroenhancement with vitamin B12-underestimated neurological significance. Nutrients. 2013;5:5031–45. doi: 10.3390/nu5125031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Reynolds E. Vitamin B12, folic acid, and the nervous system. Lancet Neurol. 2006;5:949–60. doi: 10.1016/S1474-4422(06)70598-1. [DOI] [PubMed] [Google Scholar]
- 12.Herrmann W, Schorr H, Bodis M, Knapp JP, Müller A, Stein G, et al. Role of homocysteine, cystathionine and methylmalonic acid measurement for diagnosis of vitamin deficiency in high-aged subjects. Eur J Clin Invest. 2000;30:1083–9. doi: 10.1046/j.1365-2362.2000.00746.x. [DOI] [PubMed] [Google Scholar]
- 13.Cetin I, Berti C, Calabrese S. Role of micronutrients in the periconceptional period. Hum Reprod Update. 2010;16:80–95. doi: 10.1093/humupd/dmp025. [DOI] [PubMed] [Google Scholar]
- 14.Dror DK, Allen LH. Effect of vitamin B12 deficiency on neurodevelopment in infants: current knowledge and possible mechanisms. Nutr Rev. 2008;66:250–5. doi: 10.1111/j.1753-4887.2008.00031.x. [DOI] [PubMed] [Google Scholar]
- 15.Wang ZP, Shang XX, Zhao ZT. Low maternal vitamin B (12) is a risk factor for neural tube defects: a meta-analysis. J Matern Fetal Neonatal Med. 2012;25:389–94. doi: 10.3109/14767058.2011.580800. [DOI] [PubMed] [Google Scholar]
- 16.Bhate V, Deshpande S, Bhat D, Joshi N, Ladkat R, Watve S, et al. Vitamin B12 status of pregnant Indian women and cognitive function in their 9-year-old children. Food Nutr Bull. 2008;29:249–54. doi: 10.1177/156482650802900401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Eilander A, Muthayya S, van der Knaap H, Srinivasan K, Thomas T, Kok FJ. Undernutrition, fatty acid and micronutrient status in relation to cognitive performance in Indian school children: a cross-sectional study. Br J Nutr. 2010;103:1056–64. doi: 10.1017/S000711450999273X. [DOI] [PubMed] [Google Scholar]
- 18.Rathod R, Khaire A, Kemse N, Kale A, Joshi S. Maternal omega-3 fatty acid supplementation on vitamin B12 rich diet improves brain omega-3 fatty acids, neurotrophins and cognition in the Wistar rat offspring. Brain Dev. 2014;36:853–63. doi: 10.1016/j.braindev.2013.12.007. [DOI] [PubMed] [Google Scholar]
- 19.van de Rest O, van Hooijdonk LW, Doets E, Schiepers OJ, Eilander A, de Groot LC. B vitamins and n-3 fatty acids for brain development and function: review of human studies. Ann Nutr Metab. 2012;60:272–92. doi: 10.1159/000337945. [DOI] [PubMed] [Google Scholar]
- 20.Parlettaa N, Milteb CM, Meyerc BJ. Nutritional modulation of cognitive function and mental health. J Nutr Biochem. 2013;24:725–43. doi: 10.1016/j.jnutbio.2013.01.002. [DOI] [PubMed] [Google Scholar]
- 21.Innis SM. Perinatal biochemistry and physiology of long chain polyunsaturated fatty acids. J Pediatr. 2003;143(Suppl 4):1–8. doi: 10.1067/S0022-3476(03)00396-2. [DOI] [PubMed] [Google Scholar]
- 22.Manav M, Su J, Hughes K, Lee HP, Ong CN. Omega-3 fatty acids and selenium as coronary heart disease risk modifying factors in Asian Indian and Chinese males. Nutrition. 2004;20:967–73. doi: 10.1016/j.nut.2004.08.003. [DOI] [PubMed] [Google Scholar]
- 23.Simopoulos AP. Evolutionary aspects of diet: the omega-6/omega-3 ratio and the brain. Mol Neurobiol. 2011;44:203–15. doi: 10.1007/s12035-010-8162-0. [DOI] [PubMed] [Google Scholar]
- 24.Jump DB. The biochemistry of n-3 polyunsaturated fatty acids. J Biol Chem. 2002;277:8755–8. doi: 10.1074/jbc.R100062200. [DOI] [PubMed] [Google Scholar]
- 25.Kanoski SE, Davidson TL. Western diet consumption and cognitive impairment: links to hippocampal dysfunction and obesity. Physiol Behav. 2011;103:59–68. doi: 10.1016/j.physbeh.2010.12.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Sinn N, Milte C, Howe PR. Oiling the brain: a review of randomized controlled trials of omega-3 fatty acids in psychopathology across the lifespan. Nutrients. 2010;2:128–70. doi: 10.3390/nu2020128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Wainwright PE. Dietary essential fatty acids and brain function: a developmental perspective on mechanisms. Proc Nutr Soc. 2002;61:61–9. doi: 10.1079/PNS2001130. [DOI] [PubMed] [Google Scholar]
- 28.Hashimoto M. Omega-3 fatty acids and cognition. Nihon Rinsho. 2014;72:648–56. [PubMed] [Google Scholar]
- 29.Feng Z, Zou X, Jia H, Li X, Zhu Z, Liu X, et al. Maternal docosahexaenoic acid feeding protects against impairment of learning and memory and oxidative stress in prenatally stressed rats: possible role of neuronal mitochondria metabolism. Antioxid Redox Signal. 2012;16:275–89. doi: 10.1089/ars.2010.3750. [DOI] [PubMed] [Google Scholar]
- 30.Tuzun F, Kumral A, Ozbal S, Dilek M, Tugyan K, Duman N, et al. Maternal prenatal omega-3 fatty acid supplementation attenuates hyperoxia-induced apoptosis in the developing rat brain. Int J Dev Neurosci. 2012;30:315–23. doi: 10.1016/j.ijdevneu.2012.01.007. [DOI] [PubMed] [Google Scholar]
- 31.Suganuma H, Arai Y, Kitamura Y, Hayashi M, Okumura A, Shimizu T. Maternal docosahexaenoic acid-enriched diet prevents neonatal brain injury. Neuropathology. 2010;30:597–605. doi: 10.1111/j.1440-1789.2010.01114.x. [DOI] [PubMed] [Google Scholar]
- 32.Wilczyńska A. Fatty acids in treatment and prevention of depression. Psychiatr Pol. 2013;47:657–66. [PubMed] [Google Scholar]
- 33.Cao D, Kevala K, Kim J, Moon HS, Jun SB, Lovinger D, et al. Docosahexaenoic acid promotes hippocampal neuronal development and synaptic function. J Neurochem. 2009;111:510–21. doi: 10.1111/j.1471-4159.2009.06335.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Sakamoto T, Cansev M, Wurtman RJ. Oral supplementation with docosahexaenoic acid and uridine-5′-monophosphate increases dendritic spine density in adult gerbil hippocampus. Brain Res. 2007;1182:50–9. doi: 10.1016/j.brainres.2007.08.089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Chung WL, Chen JJ, Su HM. Fish oil supplementation of control and (n-3) fatty acid-deficient male rats enhances reference and working memory performance and increases brain regional docosahexaenoic acid levels. J Nutr. 2008;138:1165–71. doi: 10.3945/jn.108.089631. [DOI] [PubMed] [Google Scholar]
- 36.Helland IB, Smith L, Blomén B, Saarem K, Saugstad OD, Drevon CA. Effect of supplementing pregnant and lactating mothers with n-3 very-long-chain fatty acids on children’s IQ and body mass index at 7 years of age. Pediatrics. 2008;122:e472–9. doi: 10.1542/peds.2007-2762. [DOI] [PubMed] [Google Scholar]
- 37.Pérez MÁ, Terreros G, Dagnino-Subiabre A. Long-term ω-3 fatty acid supplementation induces anti-stress effects and improves learning in rats. Behav Brain Funct. 2013;9:25. doi: 10.1186/1744-9081-9-25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Wu A, Ying Z, Gomez-Pinilla F. Docosahexaenoic acid dietary supplementation enhances the effects of exercise on synaptic plasticity and cognition. Neuroscience. 2008;155:751–9. doi: 10.1016/j.neuroscience.2008.05.061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.James SJ, Cutler P, Melnyk S, Jernigan S, Janak L, Gaylor DW, et al. Metabolic biomarkers of increased oxidative stress and impaired methylation capacity in children with autism. Am J Clin Nutr. 2004;80:1611–7. doi: 10.1093/ajcn/80.6.1611. [DOI] [PubMed] [Google Scholar]
- 40.de Jager CA. Critical levels of brain atrophy associated with homocysteine and cognitive decline. Neurobiol Aging. 2014;35:S35–9. doi: 10.1016/j.neurobiolaging.2014.03.040. [DOI] [PubMed] [Google Scholar]
- 41.Geisel J, Schorr H, Bodis M, Isber S, Hübner U, Knapp JP, et al. The vegetarian lifestyle and DNA methylation. Clin Chem Lab Med. 2005;43:1164–9. doi: 10.1515/CCLM.2005.202. [DOI] [PubMed] [Google Scholar]
- 42.Lipton SA, Kim WK, Choi YB, Kumar S, D’Emilia DM, Rayudu PV, et al. Neurotoxicity associated with dual actions of homocysteine at the N-methyl-D aspartate receptor. Proc Natl Acad Sci U S A. 1997;94:5923–8. doi: 10.1073/pnas.94.11.5923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Ziemińska E, Stafiej A, Łazarewicz JW. Role of group I metabotropic glutamate receptors and NMDA receptors in homocysteine-evoked acute neurodegeneration of cultured cerebellar granule neurons. Neurochem Int. 2003;43:481–92. doi: 10.1016/S0197-0186(03)00038-X. [DOI] [PubMed] [Google Scholar]
- 44.Poddar R, Paul S. Homocysteine-NMDA receptor-mediated activation of extracellular signal-regulated kinase leads to neuronal cell death. J Neurochem. 2009;110:1095–106. doi: 10.1111/j.1471-4159.2009.06207.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Obeid R, Herrmann W. Mechanisms of homocysteine neurotoxicity in neurodegenerative diseases with special reference to dementia. FEBS Lett. 2006;580:2994–3005. doi: 10.1016/j.febslet.2006.04.088. [DOI] [PubMed] [Google Scholar]
- 46.Mattson MP, Shea TB. Folate and homocysteine metabolism in neural plasticity and neurodegenerative disorders. Trends Neurosci. 2003;26:137–46. doi: 10.1016/S0166-2236(03)00032-8. [DOI] [PubMed] [Google Scholar]
- 47.Tyagi N, Sedoris KC, Steed M, Ovechkin AV, Moshal KS, Tyagi SC. Mechanisms of homocysteine-induced oxidative stress. Am J Physiol Heart Circ Physiol. 2005;289:H2649–56. doi: 10.1152/ajpheart.00548.2005. [DOI] [PubMed] [Google Scholar]
- 48.Halliwell B. Role of free radicals in the neurodegenerative diseases: therapeutic implications for antioxidant treatment. Drugs Aging. 2001;18:685–716. doi: 10.2165/00002512-200118090-00004. [DOI] [PubMed] [Google Scholar]
- 49.Song JH, Fujimoto K, Miyazawa T. Polyunsaturated (n-3) fatty acids susceptible to peroxidation are increased in plasma and tissue lipids of rats fed docosahexaenoic acid-containing oils. J Nutr. 2000;130:3028–33. doi: 10.1093/jn/130.12.3028. [DOI] [PubMed] [Google Scholar]
- 50.Sable PS, Kale AA, Joshi SR. Prenatal omega-3 fatty acid supplementation to a micronutrient imbalanced diet protects brain neurotrophins both in the cortex and hippocampus in the adult rat offspring. Metabolism. 2013;62:1607–22. doi: 10.1016/j.metabol.2013.06.002. [DOI] [PubMed] [Google Scholar]
- 51.Kale A, Naphade N, Sapkale S, Kamaraju M, Pillai A, Joshi S, et al. Reduced folic acid, vitamin B12 and docosahexaenoic acid and increased homocysteine and cortisol in never-medicated schizophrenia patients: implications for altered one-carbon metabolism. Psychiatry Res. 2010;175:47–53. doi: 10.1016/j.psychres.2009.01.013. [DOI] [PubMed] [Google Scholar]
- 52.Kulkarni A, Mehendale S, Pisal H, Kilari A, Dangat K, Salunkhe S, et al. Association of omega-3 fatty acids and homocysteine concentrations in pre-eclampsia. Clin Nutr. 2011;30:60–4. doi: 10.1016/j.clnu.2010.07.007. [DOI] [PubMed] [Google Scholar]
- 53.Faraci FM, Lentz SR. Hyperhomocysteinemia, oxidative stress, and cerebral vascular dysfunction. Stroke. 2004;35:345–7. doi: 10.1161/01.STR.0000115161.10646.67. [DOI] [PubMed] [Google Scholar]
- 54.Lominadze D, Roberts AM, Tyagi N, Moshal KS, Tyagi SC. Homocysteine causes cerebrovascular leakage in mice. Am J Physiol Heart Circ Physiol. 2006;290:H1206–13. doi: 10.1152/ajpheart.00376.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Troen AM, Shea-Budgell M, Shukitt-Hale B, Smith DE, Selhub J, Rosenberg IH. B-vitamin deficiency causes hyperhomocysteinemia and vascular cognitive impairment in mice. Proc Natl Acad Sci U S A. 2008;105:12474–9. doi: 10.1073/pnas.0805350105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Kamath AF, Chauhan AK, Kisucka J, Dole VS, Loscalzo J, Handy DE, et al. Elevated levels of homocysteine compromise blood–brain barrier integrity in mice. Blood. 2006;107:591–3. doi: 10.1182/blood-2005-06-2506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Zhang Q, Li Q, Chen Y, Huang X, Yang IH, Cao L, et al. Homocysteine-impaired angiogenesis is associated with VEGF/VEGFR inhibition. Front Biosci. 2012;4:2525–35. doi: 10.2741/E563. [DOI] [PubMed] [Google Scholar]
- 58.Wang J, Shi Y, Zhang L, Zhang F, Hu X, Zhang W, et al. Omega-3 polyunsaturated fatty acids enhance cerebral angiogenesis and provide long-term protection after stroke. Neurobiol Dis. 2014;68:91–103. doi: 10.1016/j.nbd.2014.04.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Scalabrino G, Buccellato FR, Veber D, Mutti E. New basis of the neurotrophic action of vitamin B12. Clin Chem Lab Med. 2003;41:1435–7. doi: 10.1515/CCLM.2003.220. [DOI] [PubMed] [Google Scholar]
- 60.Sable P, Dangat K, Kale A, Joshi S. Altered brain neurotrophins at birth: consequence of imbalance in maternal folic acid and vitamin B12 metabolism. Neuroscience. 2011;190:127–34. doi: 10.1016/j.neuroscience.2011.05.010. [DOI] [PubMed] [Google Scholar]
- 61.Sable PS, Dangat KD, Joshi AA, Joshi SR. Maternal omega-3 fatty acid supplementation during pregnancy to a micronutrient imbalanced diet protects postnatal reduction of brain neurotrophins in the rat offspring. Neuroscience. 2012;217:46–55. doi: 10.1016/j.neuroscience.2012.05.001. [DOI] [PubMed] [Google Scholar]
- 62.Bhatia S, Agrawal R, Sharma S, Huo YX, Ying Z, Gomez-Pinilla F. Omega-3 fatty acid deficiency during brain maturation reduces neuronal and behavioral plasticity in adulthood. PLoS One. 2011;6 doi: 10.1371/journal.pone.0028451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Sotiropoulou M, Mantas C, Bozidis P, Marselos M, Mavreas V, Hyphantis T, et al. BDNF serum concentrations in first psychotic episode drug-naïve schizophrenic patients: associations with personality and BDNF Val66Met polymorphism. Life Sci. 2013;92:305–10. doi: 10.1016/j.lfs.2013.01.008. [DOI] [PubMed] [Google Scholar]
- 64.Laske C, Stransky E, Leyhe T, Eschweiler GW, Wittorf A, Richartz E, et al. Stage-dependent BDNF serum concentrations in Alzheimer’s disease. J Neural Transm. 2006;113:1217–24. doi: 10.1007/s00702-005-0397-y. [DOI] [PubMed] [Google Scholar]
- 65.Scalzo P, Kümmer A, Bretas TL, Cardoso F, Teixeira AL. Serum levels of brain derived neurotrophic factor correlate with motor impairment in Parkinson’s disease. J Neurol. 2010;257:540–5. doi: 10.1007/s00415-009-5357-2. [DOI] [PubMed] [Google Scholar]
- 66.Baydyuk M, Xu B. BDNF signaling and survival of striatal neurons. Front Cell Neurosci. 2014;8:254. doi: 10.3389/fncel.2014.00254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Xiong P, Zeng Y, Zhu Z, Tan D, Xu F, Lu J, et al. Reduced NGF serum levels and abnormal P300 event-related potential in first episode schizophrenia. Schizophr Res. 2010;119:34–9. doi: 10.1016/j.schres.2010.02.1063. [DOI] [PubMed] [Google Scholar]
- 68.Zhang XY, Liang J, Chen da C, Xiu MH, Yang FD, Kosten TA, et al. Low BDNF is associated with cognitive impairment in chronic patients with schizophrenia. Psychopharmacology (Berl) 2012;222:277–84. doi: 10.1007/s00213-012-2643-y. [DOI] [PubMed] [Google Scholar]
- 69.Cai J, Hua F, Yuan L, Tang W, Lu J, Yu S, et al. Potential therapeutic effects of neurotrophins for acute and chronic neurological diseases. Biomed Res Int. 2014;2014:601084. doi: 10.1155/2014/601084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Balanzá-Martínez V, Fries GR, Colpo GD, Silveira PP, Portella AK, Tabarés-Seisdedos R, et al. Therapeutic use of omega-3 fatty acids in bipolar disorder. Expert Rev Neurother. 2011;11:1029–47. doi: 10.1586/ern.11.42. [DOI] [PubMed] [Google Scholar]
- 71.Wu A, Ying Z, Gomez-Pinilla F. Dietary omega-3 fatty acids normalize BDNF levels, reduce oxidative damage, and counteract learning disability after traumatic brain injury in rats. J Neurotrauma. 2004;21:457–67. doi: 10.1089/neu.2004.21.1457. [DOI] [PubMed] [Google Scholar]
- 72.Jiang LH, Shi Y, Wang LS, Yang ZR. The influence of orally administered docosahexaenoic acid on cognitive ability in aged mice. J Nutr Biochem. 2009;20:735–41. doi: 10.1016/j.jnutbio.2008.07.003. [DOI] [PubMed] [Google Scholar]
- 73.Kermani P, Hempstead B. Brain-derived neurotrophic factor: a newly described mediator of angiogenesis. Trends Cardiovasc Med. 2007;17:140–3. doi: 10.1016/j.tcm.2007.03.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Nico B, Mangieri D, Benagiano V, Crivellato E, Ribatti D. Nerve growth factor as an angiogenic factor. Microvasc Res. 2008;75:135–41. doi: 10.1016/j.mvr.2007.07.004. [DOI] [PubMed] [Google Scholar]
- 75.Calza L, Giardino L, Giuliani A, Aloe L, Levi-Montalcini R. Nerve growth factor control of neuronal expression of angiogenetic and vasoactive factors. Proc Natl Acad Sci U S A. 2001;8:4160–5. doi: 10.1073/pnas.051626998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Licht T, Goshen I, Avital A, Kreisel T, Zubedat S, Eavri R, et al. Reversible modulation of neuronal plasticity by VEGF. Proc Natl Acad Sci U S A. 2011;108:5081–6. doi: 10.1073/pnas.1007640108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Mackenzie F, Ruhrberg C. Diverse roles for VEGF-A in the nervous system. Development. 2012;139:1371–80. doi: 10.1242/dev.072348. [DOI] [PubMed] [Google Scholar]
- 78.Nakamura K, Tan F, Li Z, Thiele CJ. NGF activation of TrkA induces vascular endothelial growth factor expression via induction of hypoxia-inducible factor-1α. Mol Cell Neurosci. 2011;46:498–506. doi: 10.1016/j.mcn.2010.12.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.García-Maceira P, Mateo J. Silibinin inhibits hypoxia-inducible factor-1alpha and mTOR/p70S6K/4E-BP1 signalling pathway in human cervical and hepatoma cancer cells: implications for anticancer therapy. Oncogene. 2009;28:313–24. doi: 10.1038/onc.2008.398. [DOI] [PubMed] [Google Scholar]
- 80.Bjornsti MA, Houghton PJ. The TOR pathway: a target for cancer therapy. Nat Rev Cancer. 2004;4:335–48. doi: 10.1038/nrc1362. [DOI] [PubMed] [Google Scholar]
- 81.van den Beucken T, Koritzinsky M, Wouters BG. Translational control of gene expression during hypoxia. Cancer Biol Ther. 2006;5:749–55. doi: 10.4161/cbt.5.7.2972. [DOI] [PubMed] [Google Scholar]
- 82.Rathod RS, Khaire AA, Kale AA, Joshi SR. Maternal omega-3 fatty acid supplementation to a vitamin B12 deficient diet normalizes angiogenic markers in the pup brain at birth. Int J Dev Neurosci. 2015;43:43–9. doi: 10.1016/j.ijdevneu.2015.04.006. [DOI] [PubMed] [Google Scholar]
- 83.Sudgen C. One-carbon metabolism in psychiatric illness. Nutr Res Rev. 2006;19:117–36. doi: 10.1079/NRR2006119. [DOI] [PubMed] [Google Scholar]
- 84.Umhau JC, Dauphinais KM, Patel SH, Nahrwold DA, Hibbeln JR, Rawlings RR, et al. The relationship between folate and docosahexaenoic acid in men. Eur J Clin Nutr. 2006;60:352–7. doi: 10.1038/sj.ejcn.1602321. [DOI] [PubMed] [Google Scholar]
- 85.van Wijk N, Watkins CJ, Hageman RJ, Sijben J, Kamphuis PG, Wurtman RJ, et al. Combined dietary folate vitamin B-12 and vitamin B-6 intake influences plasma docosahexaenoic acid concentration in rats. Nutr Metab (Lond) 2012;9:49. doi: 10.1186/1743-7075-9-49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Selley ML. A metabolic link between S-adenosyl homocysteine and polyunsaturated fatty acid metabolism in Alzheimer’s disease. Neurobiol Aging. 2007;28:1834–9. doi: 10.1016/j.neurobiolaging.2006.08.003. [DOI] [PubMed] [Google Scholar]
- 87.Schaevitz L, Berger-Sweeney J, Ricceri L. One-carbon metabolism in neurodevelopmental disorders: using broad-based nutraceutics to treat cognitive deficits in complex spectrum disorders. Neurosci Biobehav Rev. 2014;46:270–84. doi: 10.1016/j.neubiorev.2014.04.007. [DOI] [PubMed] [Google Scholar]
- 88.Koukoura O, Sifakis S, Spandidos DA. DNA methylation in the human placenta and fetal growth (review) Mol Med Rep. 2012;5:883–9. doi: 10.3892/mmr.2012.763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Sui L, Wang Y, Ju LH, Chen M. Epigenetic regulation of reelin and brain-derived neurotrophic factor genes in long-term potentiation in rat medial prefrontal cortex. Neurobiol Learn Mem. 2012;97:425–40. doi: 10.1016/j.nlm.2012.03.007. [DOI] [PubMed] [Google Scholar]
- 90.Gapp K, Woldemichael BT, Bohacek J, Mansuy IM. Epigenetic regulation in neurodevelopment and neurodegenerative diseases. Neuroscience. 2014;264:99–111. doi: 10.1016/j.neuroscience.2012.11.040. [DOI] [PubMed] [Google Scholar]
- 91.Lester BM, Tronick E, Nestler E, Abel T, Kosofsky B, Kuzawa CW, et al. Behavioral epigenetics. Ann N Y Acad Sci. 2011;1226:14–33. doi: 10.1111/j.1749-6632.2011.06037.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Dennis KE, Levitt P. Regional expression of brain derived neurotrophic factor (BDNF) is correlated with dynamic patterns of promoter methylation in the developing mouse forebrain. Brain Res Mol Brain Res. 2005;140:1–9. doi: 10.1016/j.molbrainres.2005.06.014. [DOI] [PubMed] [Google Scholar]
- 93.Pogribny IP, Karpf AR, James SR, Melnyk S, Han T, Tryndyak VP. Epigenetic alterations in the brains of Fisher 344 rats induced by long-term administration of folate/methyl-deficient diet. Brain Res. 2008;1237:25–34. doi: 10.1016/j.brainres.2008.07.077. [DOI] [PubMed] [Google Scholar]
- 94.Benatti P, Peluso G, Nicolai R, Calvani M. Polyunsaturated fatty acids: biochemical, nutritional and epigenetic properties. J Am Coll Nutr. 2004;23:281–302. doi: 10.1080/07315724.2004.10719371. [DOI] [PubMed] [Google Scholar]
- 95.Sable P, Randhir K, Kale A, Chavan-Gautam P, Joshi S. Maternal micronutrients and brain global methylation patterns in the offspring. Nutr Neurosci. 2015;18:30–6. doi: 10.1179/1476830513Y.0000000097. [DOI] [PubMed] [Google Scholar]
- 96.Costello JF. DNA methylation in brain development and gliomagenesis. Front Biosci. 2003;8:s175–84. doi: 10.2741/1027. [DOI] [PubMed] [Google Scholar]
- 97.Sharma RP, Gavin DP, Grayson DR. CpG methylation in neurons: message, memory, or mask? Neuropsychopharmacology. 2010;35:2009–20. doi: 10.1038/npp.2010.85. [DOI] [PMC free article] [PubMed] [Google Scholar]