

Abstract
Purpose
Previous studies have demonstrated that melatonin has an important role in the modulation of photoreceptor viability during aging and may be involved in the pathogenesis of age-related macular degeneration.This hormone exerts its influence by binding to G-protein coupled receptors named melatonin receptor 1 (MT1) and 2 (MT2). Melatonin receptors 1 and 2 activate a wide variety of signaling pathways.
Methods
Melatonin-proficient mice (C3H/f+/+) and melatonin-proficient mice lacking MT1 or MT2 receptors (MT1−/− and MT2−/−) were used in this study. Mice were killed at the ages of 3 and 18 months, and photoreceptor viability was determined by counting nuclei number in the outer nuclear layer (ONL). Cones were identified by immunohistochemistry using peanut agglutinin (PNA) and green/red and blue opsin antibodies. Protein kinase B (AKT) and forkhead box O (FOXO1) were assessed by Western blotting and immunohistochemistry.
Results
The number of nuclei in the ONL was significantly reduced in C3Hf+/+, MT1−/−, and MT2−/− mice at 18 months of age with respect to 3-month-old animals. In 18-month-old MT1−/− and MT2−/− mice, but not in C3H/f+/+, the number of cones was significantly reduced with respect to young MT1−/− and MT2−/− mice or age-matched C3H/f+/+. In C3H/f+/+, activation of the AKT-FOXO1 pathway in the photoreceptors showed a significant difference between night and day.
Conclusions
Our data indicate that disruption of MT1/MT2 heteromer signaling induces a reduction in the number of photoreceptors during aging and also suggest that the AKT-FOXO1 survival pathway may be involved in the mechanism by which melatonin protects photoreceptors.
Keywords: melatonin, viability, photoreceptors
Retinal melatonin is synthesized by photoreceptors of many vertebrate species via a well-defined biosynthetic pathway.1,2 The amount of melatonin produced by the retina is much smaller than that produced by the pineal gland, and retinal melatonin is thought to act as a local neuromodulator within the eye.3 Melatonin synthesis in the retina, as in the pineal gland, occurs primarily during the night in darkness, and, thus, melatonin levels are high during the night and low during the day.4 Melatonin synthesis is controlled by a circadian clock located within the photoreceptors.5–8 Transcriptional and posttranslational mechanisms ensure that melatonin levels are maintained at extremely low levels in the presence of light.9,10 Such tight control of retinal melatonin levels suggests that high melatonin levels during the light-phase may be deleterious for the photoreceptor cells.11
Melatonin exerts its influence by binding to G protein-coupled receptors (GPCRs) named melatonin receptor type 1 (MT1) and type 2 (MT2). Both receptors are present in the vertebrate retina and activate a wide variety of signaling pathways.12–15 In the photoreceptors MT1 and MT2 form heteromeric complexes, as demonstrated previously in vitro16,17 and recently confirmed in vivo.12
Melatonin modulates the sensitivity of photoreceptors and second-order neurons at night when photopic input is low.18 In some species, melatonin can affect glutamatergic transmission from cones to cone-driven bipolar cells19 and may potentiate responses of ON bipolar cells to rod signals.20 In Xenopus laevis, melatonin directly stimulates the responsiveness of rod photoreceptors.21 Administration of exogenous melatonin in fishes and amphibians can increase the amplitude of the a- and b-wave of the scotopic electroretinogram.20,21 In mice, administration of exogenous melatonin increases the amplitudes of a- and b- waves and lowers the scotopic threshold response (i.e., visual sensitivity) to levels observed at night under controlled conditions; removal of MT1 and/or MT2 receptors abolishes these effects.12,22 Finally, several studies have shown that melatonin has a key role in the regulation of retinal circadian rhythms.23,24
Earlier studies also have demonstrated that melatonin can exert beneficial and detrimental effects on photoreceptors: on one hand it may have an important role in protecting them from oxidative stress,25 preventing apoptosis26 and promotes photoreceptor viability during aging.22 On the other hand, it also can sensitize photoreceptors to light-induced damage.27 A few studies also have implicated melatonin in the pathogenesis of age-related macular degeneration (ARMD). Age-related macular degeneration is a slow and progressive disease of the macula, that is, the central part of the retina where cone density is maximal, and constitutes the leading cause of irreversible visual loss in the Western world.28 Yi et al.29 reported that daily administration of melatonin (3 mg) may protect the retina and delay the progression of ARMD, while Rosen et al.30 described that production of melatonin is decreased in ARMD patients with respect to age-matched controls. These findings suggest that a deficiency in melatonin may have a role in the progression of ARMD.
To gather more insight into the role of melatonin during aging, we investigated the effect of MT1 and MT2 deletion on the viability of cone photoreceptors and the potential role of melatonin receptors signaling on the survival pathway involving a serine/threonine protein kinase (AKT) and the Forkhead-related family of mammalian transcription factor (FOXO1).31
Materials and Methods
Animals
We used C3H/f+/+, C3H/f+/+MT1−/−, and C3H/f+/+MT2−/− mice in this study.12,22 All the experimental procedures were done in accordance with the Association for Assessment of Laboratory Animal Care policies and approved by the Morehouse School of Medicine Animal Care and Use Committee and in accordance with the ARVO Statement for the Use of Animals in Ophthalmic and Vision Research.
Outer Nuclear Layer Cell Counting
Photoreceptor nuclei in the ONL were counted in a 10-μm microscopic field that was centered at 300 μm above the edge of the optic nerve head. For each sample, we counted the number of photoreceptor cells in 10 different locations within each of 3 adjacent sections (see the study of Baba et al.22 for further details).
Immunohistochemical Analysis of Phosphorylated-AKT, Phosphorylated-FOXO1, and FOXO1
After euthanasia, eyes were collected and fixed with 4% paraformaldehyde (PAF) in PBS for 12 hours. Subsequent to fixation, the eyeballs were rinsed in PBS, transferred to sucrose 30% overnight, and embedded (Tissue-Tek CRYO-OCT Compound; Fisher Healthcare, Pittsburgh, PA, USA). Cryostat sections (12 μm thick) were mounted on SuperFrost*Plus slides (VWR, Atlanta, GA, USA). The retinas sections were permeabilized with 0.1% Triton X-100 for 5 minutes and the endogenous peroxidase activity was quenched with PBS + 1% H202 during 1 hour. Sections then were incubated in blocking buffer 1 hour (1% BSA in PBS) and in primary antibody overnight at 4°C. The primary antibodies were as follows: monoclonal anti-rabbit phosphorylated (P) AKT (Ser473, #4060; Cell Signaling, Boston, MA, USA), dilution 1:200; polyclonal anti-rabbit P-FOXO1 (Ser256, #9461; Cell Signaling), 1:500; monoclonal anti-rabbit FOXO1 (detects P and unphosphorylated [UnP] FOXO1; #2880; Cell Signaling), 1:200. After extensive wash in PBS, retinas were incubated 1 hour in HRP conjugated secondary antibody (Tyramide Signal Amplification Kit #12; Molecular Probes, Life Technologies, Carlsbad, CA, USA), dilution 1:100. Retinal sections again were washed thoroughly and incubated 10 minutes with labeled tyramide (Tyramide Signal Amplification Kit #12; Molecular Probes, Life Technologies), dilution 1:100. After several washes, retinal sections were mounted with mounting medium (Prolong; Molecular Probes, Life Technologies) before being examined with a confocal microscope (Zeiss LSM700).
Immunofluorescence Analysis of AKT, Red/Green Opsin, Blue Opsin, and Peanut Agglutinin
The samples were treated as described previously. For AKT immunostaining, after fixation, the eyecups were incubated in a retrieval solution (10 mM sodium citrate, pH6) overnight at 4°C. The eyes were immersed in boiling retrieval solution 3 minutes and immediately placed in cold sucrose 30%.
The sections were permeabilized with Triton X-100 (0.1% in PBS) for 5 minutes and then saturated with blocking buffer (3% BSA, 0.05% Tween-20, 0.1% Triton X-100, 0.1% sodium azide in PBS) during 2 hours. Sections were incubated 2 hours with rhodamine-labeled peanut agglutinin (PNA, RL-1072; Vector Laboratories, Burlingame, CA, USA), dilution 1:500; or overnight with primary antibody diluted in blocking buffer: polyclonal anti-rabbit red/green opsin (medium/long-wavelength opsin, AB5405; Chemicon International, Millipore, Billerica, MA, USA), 1:1000; polyclonal anti-rabbit blue opsin (short-wavelength opsin, AB5407; Chemicon International, Millipore), 1:500; monoclonal anti-mouse AKTpan (detects P and UnP AKT, #2920; Cell Signaling), 1:50. Secondary antibody incubation was performed at room temperature for 2 hours with Alexa488 conjugated goat anti-rabbit or Alexa568 conjugated goat anti-mouse (Molecular Probes, Thermo Fisher, Rockford, IL, USA), dilution 1:500. Retinas were washed thoroughly, mounted with Prolong medium (Prolong, Molecular Probes, Life Technologies) and observed with a confocal microscope (Zeiss LSM700).
Pictures were obtained of the central, middle, and peripheral sections of the retina (six regions in total). The number of cones was counted at the level of cone outer segment (OS) in a 140 μm length area for each region of the retina. The number of cones in the retina represents the average of the six values and was determined for each animal.
Western Blotting
Retinas were sonicated in cell extraction buffer (Novex; Invitrogen, Camarillo, CA, USA) and briefly centrifuged. The protein content of the supernatants were determined by Micro BCA Protein Assay Kit (Thermo Fisher Scientific Scientific). Proteins (10 μg/lane) were separated according to the method of Laemmli and transferred to Immobilon-P transfer membrane (Millipore). Membranes were briefly washed with Tris-buffered saline (20 mM Tris-HCl, pH 7.6; 150 mM NaCl) containing 0.1% Tween20 (TBS-Tween). Then membranes were blocked for 1 hour in TBS-Tween containing 5% nonfat milk powder or BSA at room temperature. Membranes were washed with TBS-Tween, and then incubated 16 hours at 4°C with primary antibodies: P-AKT (Ser473), P-AKT (Thr308, #2965; Cell Signaling), AKTpan, P-FOXO1 (Ser256), FOXO1, diluted 1:5000-1:80000 in TBS-Tween containing 1 % nonfat milk or BSA. Membranes then were washed four times for 10 minutes with TBS-Tween previous incubation with HRP conjugated donkey anti-rabbit IgG or anti-mouse IgG diluted 1:1000 to 1:5000 (Cell Signaling) in TBS-Tween containing 1% nonfat milk. Membranes finally were washed 4 times for 10 minutes with TBS-Tween. Super Signal West FemtoChemiluminescent Substrate (Thermo Fisher Scientific) was used to detect the antigen.
Statistical Analysis
Data are given as the mean ± SEM of n = 3 to 6 animals. Statistical analysis was performed with 1-way ANOVA using the Sigma plot software. The significant level was set at P < 0.05. For cell counting, ANOVA were followed by post hoc Holm-Sidak test. For Western blotting, ANOVA were followed by the post hoc Tukey test.
Results
Effect of MT1 and MT2 Deletion on Photoreceptor Cell Viability During Aging
No significant differences were observed among young (3 months of age) C3H/f+/+, C3H/f+/+MT1−/−, and C3H/f+/+MT2−/− mice (P > 0.05, 1-way ANOVA; Fig. 1A), whereas in older C3H/f+/+MT1−/−, and C3H/f+/+MT2−/− mice (18 months of age) we observed a significant reduction in the number of photoreceptors with respect to C3H/f+/+ at the same age (P < 0.05, 1-way ANOVA, Holm-Sidak test; Fig. 1B). We also observed a small, but significant, reduction in the number of photoreceptor nuclei in the ONL of older C3H/f+/+ (18 months) with respect to the number of cells of young C3H/f+/+ (3 months, P < 0.05, 1-way ANOVA, Holm-Sidak test; Figs. 1A, 1B).
Figure 1.

Melatonin receptor 1 and MT2 deletion reduce photoreceptor viability during aging. Number of cell nuclei in the ONL of C3H/f+/+, C3H/f+/+MT1−/−, and C3H/f+/+MT2−/− mice at 3 (A) and 18 months (B) of age in the central retina. A significant change in the ONL cell number occurs between C3H/f+/+MT1−/−, C3H/f+/+MT2−/−, and C3H/f+/+ at 18 months of age (*P < 0.05). The number of cells in each genotype also is affected by aging (#P < 0.05). Error bars: Mean ± SEM; n = 4 to 6. #P < 0.05, 1-way ANOVA, followed by Holm-Sidak test. WT, C3H/f+/+; MT1−/−, C3H/f+/+MT1−/−; MT2−/−, C3H/f+/+MT2−/−.
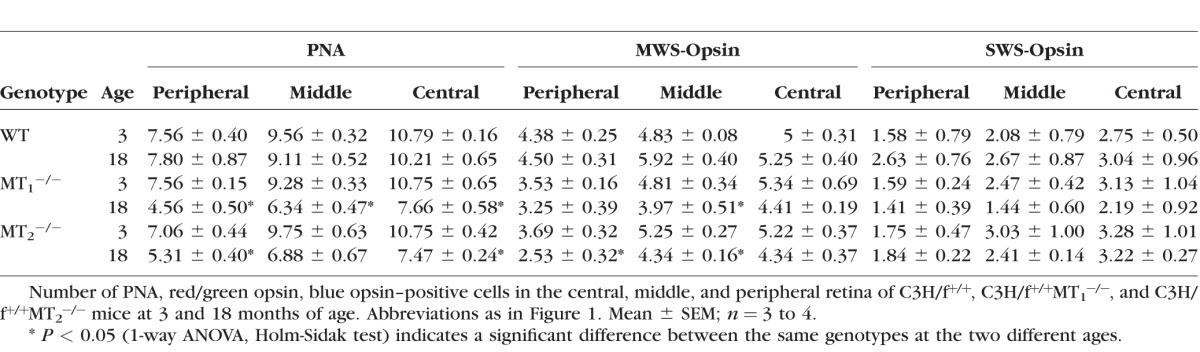
Retinal sections obtained from C3H/f+/+, C3H/f+/+MT1−/−, and C3H/f+/+MT2−/− mice at 3 and 18 months of age (Figs. 2A, 2B) were labeled with PNA to identify cones. No differences were observed in 3-month-old mice of the different genotypes (P > 0.05, 1-way ANOVA; Fig. 2C), whereas at 18 months, C3H/f+/+MT1−/− and C3H/f+/+MT2−/− mice showed a significant decrease (approximately 30%) in the number of PNA-labeled cones in the retina with respect to C3H/f+/+ at the same age (P < 0.05, 1-way ANOVA, Holm-Sidak test; Fig. 2D). Each region of the retina (peripheral, middle, central) exhibited a significant decrease in the number of PNA-positive cells for C3H/f+/+MT1−/− mice (P < 0.05, 1-way ANOVA, Holm-Sidak test; see Table), while only peripheral and central retina of C3H/f+/+MT2−/− mice showed significant decreases (P < 0.05, 1-way ANOVA, Holm-Sidak test; see Table). To explore possible differences in cone subpopulations, we labeled red/green (medium/long wavelength opsin, Figs. 2E, 2F) and blue cones (short wavelength opsin, Figs. 2I, 2J) with specific antibodies. No significant differences were observed between the three genotypes at 3 months of age for either red/green (P > 0.05, 1-way ANOVA; Fig. 2G) or blue cones (P > 0.05, 1-way ANOVA; Fig. 2K). On the other hand, there was a significant reduction (27%–30%) in the number of red/green cones in the entire retina of C3H/f+/+MT1−/− and C3H/f+/+MT2−/− mice with respect to C3H/f+/+at the same age (P < 0.05, 1-way ANOVA, Holm-Sidak test; Fig. 2H). As shown in the Table, there was a significant reduction in the number of red/green cones in the middle retina of C3H/f+/+MT1−/− and C3H/f+/+MT2−/− mice, and in the peripheral retina of C3H/f+/+MT2−/− mice (P < 0.05, 1-way ANOVA, Holm-Sidak test; see Table). The number of blue cones was not different among the different parts of the retina, for all genotypes and ages (P > 0.05, 1-way ANOVA; Fig. 2L, see Table).
Figure 2.

Melatonin receptor 1 and MT2 deletion reduce cones viability during aging. Peanut agglutinin immunoreactivity in the central retina of C3H/f+/+, C3H/f+/+MT1−/−, and C3H/f+/+MT2−/− mice at 3 months (A) and 18 months of age (B). Quantification of PNA-labeled cones at the photoreceptor OS level of C3H/f+/+, C3H/f+/+MT1−/−, and C3H/f+/+MT2−/− mice retina at 3 months (C) and 18 months of age (D). A significant change in the number of PNA-positive cells is observed between C3H/f+/+ and C3H/f+/+MT1−/−, C3H/f+/+MT2−/− at 18 months. Red/green cone (red/green opsin) localization in the central retina of 3 months (E) and 18 months (F) C3H/f+/+, C3H/f+/+MT1−/−, and C3H/f+/+MT2−/− mice. Quantification of red/green cone-positive cells in C3H/f+/+, C3H/f+/+MT1−/−, and C3H/f+/+MT2−/− mice retina at 3 months (G) and 18 months of age (H). A significant change in the number of red/green cones occurs between 18 months old C3H/f+/+ and C3H/f+/+MT1−/−, C3H/f+/+MT2−/− mice. Blue cones (blue opsin) immunoreactivity in the central retina of C3H/f+/+, C3H/f+/+MT1−/−, and C3H/f+/+MT2−/− mice at 3 months (I) and 18 months of age (J). Quantification of blue cone–positive cells in the retina of the three genotypes of mice at 3 months (K) and 18 months of age (L). The number of blue cones does not change between 18 months old C3H/f+/+ and C3H/f+/+MT1−/−, C3H/f+/+MT2−/− mice. Error bars: Mean ± SEM; n = 3 to 4. P < 0.05, 1-way ANOVA, Holm-Sidak test. Scale bar: 20 μm.
Table.
MT1 and MT2 Deletion Reduce Cone Cell Viability During Aging
Effect of MT1 and MT2 Deletion on the AKT-FOXO1 Pathway
To investigate the signaling pathway by which melatonin may affect cone viability, we determined the levels of P-AKTSer473/AKT and P-AKTThr308/AKT in the retina of C3H/f+/+, C3H/f+/+MT1−/−, and C3H/f+/+MT2−/− mice over 24 hours. In C3H/f+/+ mice, P-AKTSer473/AKT levels showed significant changes during the light/dark cycle (P < 0.01, 1-way ANOVA, Tukey test) with a peak at ZT22 (Fig. 3D). P-AKTThr308/AKT exhibited similar profile over 24 hours (P < 0.05, 1-way ANOVA, Tukey test, Fig. 3J). In C3H/f+/+MT1−/− and C3H/f+/+MT2−/− mice, P-AKTSer473/AKT (Figs. 3E, 3F) and P-AKTThr308/AKT (Figs. 3K, 3L) levels were not significantly different during the 24-hour period (P > 0.05 in each case, 1-way ANOVA). We also assayed the P-FOXO1/FOXO1 levels in the retina of the three genotypes (Fig. 4). Once more, the levels of P-FOXO1/FOXO1 exhibited a significant change in the retina of C3H/f+/+ over 24 hours with a peak at ZT22 (P < 0.05 1-way ANOVA, Tukey test; Fig. 4D). No changes in P-FOXO1/FOXO1 level were observed over the 24 hours in the retinas of C3H/f+/+MT1−/− and C3H/f+/+MT2−/− mice (P > 0.05 in both case, 1-way ANOVA; Figs. 4E, 4F).
Figure 3.

Melatonin receptor 1 and MT2 deletion effects of P-AKT/AKT levels. Western blotting analysis of P-AKT and AKT (P and UnP) levels in retinas at different Zeitgeber Time (ZT) in 3 months C3H/f+/+ (A, D, G, J), C3H/f+/+MT1−/− (B, E, H, K), and C3H/f+/+MT2−/− mice (C, F, I, L). P-AKT (Thr308 and Ser473) levels were significantly higher in the late night (ZT19-22; P < 0.05, 1-way ANOVA, followed by Tukey test) in C3H/f+/+ but not in C3H/f+/+MT1−/− and C3H/f+/+MT2−/− mice. Densitometric quantification of P-AKT (Thr308 and Ser473) and AKT levels were performed on three independent retinal samples for each ZT. AKT: serine/threonine protein kinase.
Figure 4.

Melatonin receptor 1 and MT2 deletion affects the of P-FOXO1/FOXO1 levels. Western blotting analysis of P-FOXO1 and FOXO1 levels (P and UnP) in retinas at different Zeitgeber Time (ZT) in 3-month-old C3H/f+/+ (A, D), C3H/f+/+MT1−/− (B, E), and C3H/f+/+MT2−/− mice (C, F). P-FOXO1 levels were significantly higher in the late night in C3H/f+/+ (ZT19-22, P < 0.05, 1-way ANOVA, followed by Tukey test) but not in C3H/f+/+MT1−/− and C3H/f+/+MT2−/− mice. Densitometric quantification of P-FOXO1 and FOXO1 levels were performed on three independent retinal samples for each ZT. FOXO1: Forkhead-related family of mammalian transcription factor 1.
Localization of AKT and FOXO1 in Photoreceptors
To localize AKT in the retina of C3H/f+/+, C3H/f+/+MT1−/−, and C3H/f+/+MT2−/− mice we performed immunohistochemistry using antibodies against AKT and P-AKTSer473, at ZT22 (when the level of P-AKT is highest in C3H/f+/+ mice; Figs. 3D, 3J) and ZT1 (when the level of P-AKT is lowest in C3H/f+/+ mice; Figs. 3D, 3J). In C3H/f+/+, AKTpan was widespread and immunoreactivity was observed in the OS and inner segment (IS, Supplementary Figs. S1A, S1A') as well as in the outer plexiform layer (OPL), inner plexiform layer (IPL) and ganglion cell layer (GCL, Supplementary Figs. S1A). At ZT22, P-AKTSer473 immunoreactivity was localized in OS and GCL of C3H/f+/+ (Figs. 5A, 5A'), while at ZT1 P-AKTSer473 immunoreactivity was weaker in these locations (Figs. 5B, 5B'). In C3H/f+/+MT1−/− and C3H/f+/+MT2−/− mice, P-AKTSer473 immunoreactivity was detected in the OS and in GCL at ZT22 (Figs. 5C, 5E) and at ZT1 (Figs. 5D, 5F). There were no clear changes in the signal intensity between the two time points. We then investigated the cellular localization of P-AKTSer473 in the OS by performing double immunofluorescence with PNA (Fig. 6A'). In C3H/f+/+ mice, P-AKTSer473 immunoreactivity was observed in the OS of rods and cones (Fig. 6A'').
Figure 5.

P-AKT is localized within photoreceptors and the GCL. P-AKT localization by fluorescence immunohistochemistry on the retina sections of 3 months old C3H/f+/+, C3H/f+/+MT1−/−, and C3H/f+/+MT2−/− mice at ZT22 and ZT1. At ZT22 (nighttime), P-AKT staining is intense in OS and GCL of C3H/f+/+ mice (A). P-AKT staining is moderate in OS and GCL of C3H/f+/+MT1−/− and C3H/f+/+MT2−/− mice, respectively (C, E). Enlargement of boxed area of C3H/f+/+ mice at ZT22 (A'). Staining is present in one specific structure in OS (arrowhead, [A']). Enlargement of boxed area of C3H/f+/+MT1−/− and C3H/f+/+MT2−/− mice retinas, respectively, confirm P-AKT expression in OS (C', E'). At ZT1, P-AKT staining is low in OS and GCL of C3H/f+/+ (B). P-AKT staining is present in OS and GCL of C3H/f+/+MT1−/− and C3H/f+/+MT2−/− mice, respectively (D, F). Enlargement of boxed area of C3H/f+/+ mice retinas at ZT1 confirm low P-AKT staining in OS (B'). Enlargement of boxed area of C3H/f+/+MT1−/− and C3H/f+/+MT2−/− mice retinas, respectively, show moderate staining in OS (D', F'). Control without primary antibody (G). Scale bars: 50 μm (A–G), 20 μm (A'–F').
Figure 6.
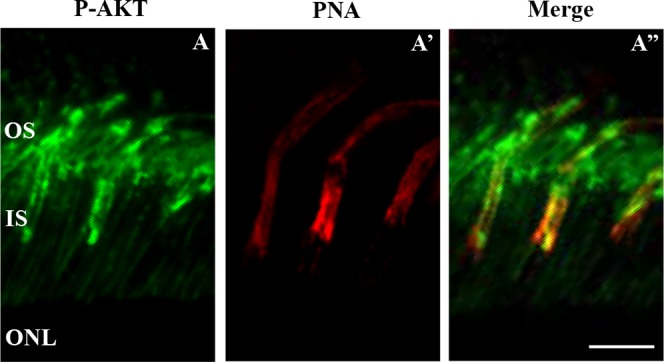
P-AKT is present in rods and cones. Fluorescence immunohistochemistry of C3H/f+/+ mice retinal sections at 3 months. Images were obtained at the level of the photoreceptor OS at ZT22. P-AKT (green) is seen in one specific structure (A). Cones are stained with PNA (red) while rods are unstained and visible as dark profiles (A'). In the merge, P-AKT is present in rod and cones OS (A''). Scale bar: 10 μm.
At ZT22, FOXO1 immunoreactivity was weak and widespread (Supplementary Figs. S2A). P-FOXO1 immunoreactivity was localized to the OS, IS and GCL of C3H/f+/+mice at ZT22 (Fig. 7A). As for P-AKT, P-FOXO1 signal in the OS and IS was stronger at ZT22 (Fig. 7A') than at ZT1 (Figs. 7B, 7B'). In C3H/f+/+MT1−/− and C3H/f+/+MT2−/− mice, P-FOXO1 was weakly detected in OS and IS at ZT22 (Figs. 7C, 7E) and ZT1 (Figs. 7D, 7F). The pattern of P-FOXO1 immunoreactivity in OS and IS was similar (slight signal and no difference between ZT22 and ZT1; Figs. 7C', 7D', 7E', 7F') in C3H/f+/+MT1−/− and C3H/f+/+MT2−/− mice. Also in this case, we performed double immunofluorescence with P-FOXO1 antibody and PNA. P-FOXO1 staining was stronger in the cones, but the signal also was present in rods (Fig. 8A'').
Figure 7.

P-FOXO1 is localized in photoreceptors and the GCL. P-FOXO1 localization by fluorescence immunohistochemistry performed on retinal sections of 3 months old C3H/f+/+, C3H/f+/+MT1−/−, and C3H/f+/+MT2−/− mice at ZT22 and ZT1. At ZT22, P-FOXO1 is localized in OS, IS and GCL (arrowhead) of C3H/f+/+ (A). Enlargement of boxed area shows the staining mainly in one specific structure in IS and OS (arrowhead, [A']). At ZT1, P-FOXO1 is not detected in the retina of C3H/f+/+ (B). Enlargement of the boxed area confirms no staining detected in OS and IS of C3H/f+/+ (B'). P-FOXO1 staining is very weak in OS, IS, and GCL of C3H/f+/+MT1−/− at ZT22 (C) and ZT1 (D) as in OS, IS, and GCL of C3H/f+/+MT2−/− mice at ZT22 (E) and ZT1 (F). Enlargement of boxed area confirms a very low level of P-FOXO1 in OS and IS of C3H/f+/+ MT1−/− and C3H/f+/+ MT2−/− mice (C'–F'). Control without primary antibody (G). Scale bars: 50 μm (A–F), 20 μm (A'–F').
Figure 8.
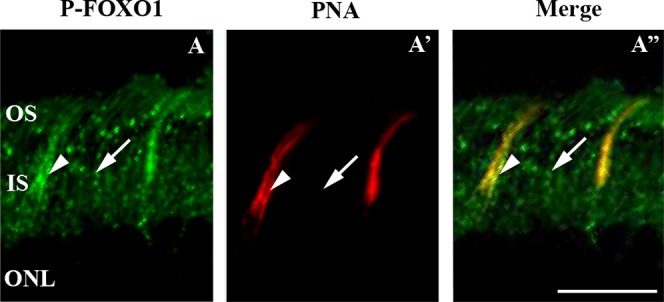
P-FOXO1 is present in rods and in cones. Fluorescence immunohistochemistry at the level of photoreceptor OS and IS of C3H/f+/+mice (3 months old) at ZT22. P-FOXO1 (green) is seen in two structures (arrow, arrowhead, [A]). Cones are stained with PNA in red (A'). Merged image reveals P-FOXO1 labeling mostly in cone (arrowhead), with weaker staining in the surrounding rod OS and IS (arrow, [A'']). Scale bar: 20 μm.
Discussion
The data presented in this study indicated that melatonin signaling via its G-protein coupled receptors increases photoreceptor viability during aging. C3H/f+/+MT1−/− and C3H/f+/+MT2−/− mice lose photoreceptors at 18 months compared to C3H/f+/+ mice at the same age (Fig. 1). The observation that similar results were obtained in C3H/f+/+MT1−/− and C3H/f+/+MT2−/− mice suggests that also in this case the action of melatonin is mediated by MT1/MT2 heteromers.12 We used PNA marker to determine the effect of MT1 and MT2 receptor deletion on cone number during aging. In the mouse, cones represent a small percentage (2%–3%) of the total number of photoreceptors with the red/green being the most prevalent type and with a wide distribution within the retina, while the blue cones are low in number and with inferior-superior gradient.32–34 Furthermore, it also has been reported that both opsins are present in the same cone.32,35 As shown in the Table, we did not detect any significant changes in the number of PNA-labeled cones in C3H/f+/+ mice during aging, whereas removal of melatonin receptors led to a decrease of PNA-labeled cones during aging. The decline in the number of red/green cones during aging is more pronounced in the middle retina of C3H/f+/+MT1−/− and C3H/f+/+MT2−/− (see Table). However, it is worth mentioning that in MT2−/− mice we also detected a significant reduction in the number of red/green cones in the periphery (see Table). This result is somewhat puzzling, since we believe that the action of melatonin on the photoreceptors is mediated by MT1/MT2 signaling.12 Further studies will be needed to determine whether the red/green cones viability in the periphery are, indeed, negatively affected by MT2 signaling removal or such a result is due to other factors (e.g., biological variability and/or counting procedures). We did not detect a significant change in the number of blue cones in C3H/f+/+, C3H/f+/+MT1−/−, and C3H/f+/+MT2−/− mice (see Table) possibly due to their unequal topography and their low density across the entire retina.33 In agreement with a previous study,33 we observed a centro-peripheral density gradient of PNA, red/green, and blue labeled cones in C3H/f+/+, C3H/f+/+MT1−/−, and C3H/f+/+MT2−/− mice at 3 and 18 months (see Table).
It is worth mentioning that the vast majority of mouse strains are genetically incapable of synthesizing melatonin.36 Indeed, most of laboratory strains (e.g., C57/BL6, Balb/C, SV129) are considered melatonin-deficient mice, whereas only CBA and C3H are considered melatonin-proficient strains. Interestingly, melatonin-deficient mice strains also show a significant decrease in cone numbers during aging37,38 further supporting the notion that melatonin receptors signaling may be an important signal to promote cones viability during aging.
Previous studies have reported that the number of photoreceptors decreases during aging and that rods and cones are both affected.22,37,39,40 However, it is believed that rods dye first and cones later;39,41 thus, suggesting that rods may produce a trophic factor necessary for cone survival.42,43 However, the variability in cone survival patterns in different rod-initiated degenerations suggests this cannot be the only explanation.44 A more recent work has suggested that the death of rods leads to development of a hyperoxygenated microenvironment that increases oxidative stress and promotes cone death.45 In our study, we detected a reduction in the number of rods (Fig. 1) and cones (Fig. 2; Table). Since MT1 and MT2 receptors are localized in both photoreceptor types,12,46 we believe that rods and cones may be directly affected by removal of melatonin receptors signaling. Additional studies are required to determine whether rods or cones die first in mice lacking melatonin receptors signaling.
Previous studies also have shown that melatonin may affect the phosphatidylinositol 3 kinase (PI3K)-AKT pathway.47 AKT has a critical role in controlling the balance between apoptosis and cell survival in response to extra- and intracellular signaling. The principal role of AKT is to facilitate growth factor-mediated cell survival and to block apoptotic cell death, which is achieved by phosphorylating and deactivating proapoptotic factors. Protein kinase B activity is the result of a balance between two phosphorylation sites on Thr308 and Ser473.48 Several studies have shown that AKT can directly phosphorylate FOXO, thus promoting cell survival.31 When AKT is inactive, FOXOs are localized within the nucleus where it induces the expression of FasL mRNA.49 On the contrary, when AKT is activated, it phosphorylates FOXOs. FOXOs then are translocated from the nucleus to the cytoplasm, thus inhibiting the expression of proapoptotic genes.49,50 Experimental evidence indicates that inactivation of AKT-FOXO protein inhibits the survival pathway in the photoreceptors.51
The results obtained with the Western blotting indicate that melatonin receptors signaling have a significant effect on the P-AKT/AKT and P-FOXO1/FOXO1 levels (Figs. 3, 4). In C3H/f+/+ mice the levels of P-AKTSer473/AKT, P-AKTThr308/AKT, and P-FOXO1/FOXO1 showed a significant increase during the night with a peak at night (ZT22, i.e., when melatonin levels are high, Figs. 3, 4). These results are consistent with previous study showing a circadian regulation of AKT phosphorylation on Thr308 that peaked at night in chick retina.52 Indeed, PI3K-AKT signaling has been shown to contribute to the circadian phase-dependent modulation of L-type voltage-gated calcium channels (L-VGCCs) in photoreceptors suggesting that PI3K-AKT signaling serves as a circadian output in the retina (52). P-AKTSer473/AKT, P-AKTThr308/AKT and P-FOXO1/FOXO1 nocturnal peaks were lost in mice lacking melatonin receptors. Such a result suggests that melatonin receptors signaling are necessary for the activation of the AKT-FOXO1 pathway at night in the retina. Although the data obtained with Western blotting were promising, they did not indicate whether the changes in the level of P-AKT/AKT and P-FOXO1/FOXO1 observed in C3H/f+/+ was occurring in the photoreceptors or in other parts of the retina. To address this question we decided to investigate the expression of P-AKT and P-FOXO1 in the retina using immunocytochemistry (Figs. 5–8). Consistent with previous reports.51,53,54 we localized P-AKT and P-FOXO1 within photoreceptors (rods and cones) and GCL. Moreover, the pattern of AKT and FOXO1 phosphorylation in the photoreceptors is affected by removal of melatonin receptors signaling (Figs. 5, 7) since no differences in the levels were observed in mice lacking melatonin receptors (Figs. 5, 7). Our data suggested that the melatonin activation of AKT-FOXO1 pathway at night may be important for the rod and cone survival during aging.
In conclusion, we believe that melatonin signaling via MT1/MT2 heteromers protects photoreceptors during aging and we propose the following working model to explain the protective action of melatonin on the photoreceptors. In photoreceptors, nocturnal melatonin binds to MT1/MT2 heteromers to elicit the recruitment of PI3K to the plasma membrane. The catalytic subunit of PI3K generates the phosphoinositide-phosphates PIP2 and PIP3, which, in turn, phosphorylate the phosphoinositide-dependent kinase-1 (PDK1). Phosphoinositide-dependent kinase-1 activates AKT at Thr308, but AKT also can be phosphorylated at Ser473 by a PDK1 independent mechanism.49,55 Once AKT is activated, FOXOs are phosphorylated; 14-3-3 protein binds to phosphorylated FOXO to export it to the cytoplasm. This leads to the suppression of proapoptotic genes transcription56 (Fig. 9).
Figure 9.

Proposed mechanism to explain the protective action of melatonin on photoreceptors. Left: during the night, once AKT is activated by melatonin receptors signaling, FOXO1 is phosphorylated. 14-3-3 protein binds to phosphorylated FOXO1 and exported it to the cytoplasm. When FOXO1 is localized in the cytoplasm, the transcription of proapoptotic genes is suppressed. Right: in the absence of melatonin receptors signaling AKT is inactive and FOXO1 is localized in the nucleus where it activates the transcription of proapoptotic genes. G, G protein; PIP2, phosphoinositide-phosphates 2; PIP3, phosphoinositide-phosphates 3; PDK1, phosphoinositide-dependent kinase-1; FasL, Fas ligand.
Supplementary Material
Acknowledgments
The authors thank the animal facility of Morehouse School of Medicine for the care and breeding of mice.
Supported by National Institutes of Health (Bethesda, MD, USA) Grants EY022216 and EY020821 (GT), and by Grants 5U54NS083932, S21MD000101, G12-RR03034, U54RR026137 to Morehouse School of Medicine.
Disclosure: C. Gianesini, None; S. Hiragaki, None; V. Laurent, None; D. Hicks, None; G. Tosini, None
References
- 1. Klein DC,, Coon SL,, Roseboom PH,, et al. The melatonin rhythm-generating enzyme: molecular regulation of serotonin N-acetyltransferase in the pineal gland. Recent Prog Horm Res. 1997; 52: 307–357. [PubMed] [Google Scholar]
- 2. Iuvone PM,, Tosini G,, Pozdeyev N,, Haque R,, Klein DC,, Chaurasia SS. Circadian clocks, clock networks, arylalkylamine N-acetyltransferase, and melatonin in the retina. Prog Retin Eye Res. 2005; 24: 433–456. [DOI] [PubMed] [Google Scholar]
- 3. Cahill GM,, Besharse JC. Retinal melatonin is metabolized within the eye of xenopus laevis. Proc Natl Acad Sci U S A. 1989; 86: 1098–1102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Tosini G,, Baba K,, Hwang CK,, Iuvone PM. Melatonin: an underappreciated player in retinal physiology and pathophysiology. Exp Eye Res. 2012; 103: 82–89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Cahill GM,, Besharse JC. Circadian clock functions localized in Xenopus retinal photoreceptors. Neuron. 1993; 10: 573–577. [DOI] [PubMed] [Google Scholar]
- 6. Tosini G,, Menaker M. The clock in the mouse retina: melatonin synthesis and photoreceptor degeneration. Brain Res. 1998; 789: 221–228. [DOI] [PubMed] [Google Scholar]
- 7. Tosini G,, Davidson AJ,, Fukuhara C,, Kasamatsu M,, Castanon-Cervantes O. Localization of a circadian clock in mammalian photoreceptors. FASEB J. 2007; 21: 3866–3871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Zawilska JB,, Iuvone PM. Melatonin synthesis in chicken retina: effect of kainic acid-induced lesions on the diurnal rhythm and D2-dopamine receptor-mediated regulation of serotonin N-acetyltransferase activity. Neurosci Letters. 1992; 135: 71–74. [DOI] [PubMed] [Google Scholar]
- 9. Fukuhara C,, Dirden JC,, Tosini G. Photic regulation of melatonin in rat retina and the role of proteasomal proteolysis. Neuroreport. 2001; 12: 3833–3837. [DOI] [PubMed] [Google Scholar]
- 10. Fukuhara C, Liu C, Ivanova TN,et al. . Gating of the cAMP signaling cascade and melatonin synthesis by the circadian clock in mammalian retina. J Neurosci. 2004; 24: 1803–1811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Wiechmann AF,, O'Steen WK. Melatonin increases photoreceptor susceptibility to light-induced damage. Invest Ophthalmol Vis Sci. 1992; 33: 1894–1902. [PubMed] [Google Scholar]
- 12. Baba K,, Benleulmi-Chaachoua A,, Journé A-S,, et al. Heteromeric MT1/MT2 Melatonin receptors modulate photoreceptor function. Sci Signal. 2013; 6:ra89. [DOI] [PMC free article] [PubMed]
- 13. Scher J,, Wankiewicz E,, Brown GM,, Fujieda H. MT(1) melatonin receptor in the human retina: expression and localization. Invest Ophthalmol Vis Sci. 2002; 43: 889–897. [PubMed] [Google Scholar]
- 14. Savaskan E,, Jockers R,, Ayoub M,, et al. The MT2 melatonin receptor subtype is present in human retina and decreases in Alzheimer's disease. Curr Alzheimer Res. 2007; 4: 47–51. [DOI] [PubMed] [Google Scholar]
- 15. Fujieda H,, Hamadanizadeh SA,, Wankiewicz E,, Pang SF,, Brown GM. Expression of mt1 melatonin receptor in rat retina: evidence for multiple cell targets for melatonin. Neuroscience. 1999; 93: 793–799. [DOI] [PubMed] [Google Scholar]
- 16. Ayoub MA,, Levoye A,, Delagrange P,, Jockers R. Preferential formation of MT1/MT2 melatonin receptor heterodimers with distinct ligand interaction properties compared with MT2 homodimers. Mol Pharmacol. 2004; 66: 312–321. [DOI] [PubMed] [Google Scholar]
- 17. Ayoub MA,, Couturier C,, Lucas-Meunier E,, et al. Monitoring of ligand-independent dimerization and ligand-induced conformational changes of melatonin receptors in living cells by bioluminescence resonance energy transfer. J Biol Chem. 2002; 277: 21522–21528. [DOI] [PubMed] [Google Scholar]
- 18. Wiechmann AF,, Yang XL,, Wu SM,, Hollyfield JG. Melatonin enhances horizontal cell sensitivity in salamander retina. Brain Res. 1988; 453: 377–380. [DOI] [PubMed] [Google Scholar]
- 19. Huang H,, Lee S-C,, Yang X-L. Modulation by melatonin of glutamatergic synaptic transmission in the carp retina. J Phyiol. 2005; 569: 857–871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Ping Y,, Huang H,, Zhang X-J,, Yang X-L. Melatonin potentiates rod signals to ON type bipolar cells in fish retina. J Physiol. 2008; 586: 2683–2694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Wiechmann AF,, Vrieze MJ,, Dighe R,, Hu Y. Direct modulation of rod photoreceptor responsiveness through a Mel 1c melatonin receptor in transgenic Xenopus laevis retina. Invest Ophthalmol Vis Sci. 2003; 44: 4522–4531. [DOI] [PubMed] [Google Scholar]
- 22. Baba K,, Pozdeyev N,, Mazzoni F,, et al. Melatonin modulates visual function and cell viability in the mouse retina via the MT1 melatonin receptor. Proc Natl Acad Sci U S A. 2009; 106: 15043–15048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Hiragaki S,, Baba K,, Coulson E,, Kunst S,, Spessert R,, Tosini G. Melatonin signaling modulates clock genes expression in the mouse retina. PLoS One. 2014; 9: e106819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. McMahon DG,, Iuvone PM,, Tosini G. Circadian organization of the mammalian retina: from gene regulation to physiology and diseases. Prog Retin Eye Res. 2014; 39: 58–76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Marchiafava PL,, Longoni B. Melatonin as an antioxidant in retinal photoreceptors. J Pineal Res. 1999; 26: 184–189. [DOI] [PubMed] [Google Scholar]
- 26. Liang FQ,, Aleman TS, ZaixinYang, Cideciyan AV, Jacobson SG, Bennett J. Melatonin delays photoreceptor degeneration in the rds/rds mouse. Neuroreport. 2001; 12: 1011–1014. [DOI] [PubMed] [Google Scholar]
- 27. Sugawara T,, Sieving PA,, Iuvone PM,, Bush RA. The melatonin antagonist luzindole protects retinal photoreceptors from light damage in the rat. Invest Ophthalmol Vis Sci. 1998; 39: 2458–2465. [PubMed] [Google Scholar]
- 28. Coleman HR,, Chan C-C,, Ferris FL,, Chew EY. Age-related macular degeneration. Lancet. 2008; 372: 1835–1845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Yi C,, Pan X,, Yan H,, Guo M,, Pierpaoli W. Effects of melatonin in age-related macular degeneration. Ann N Y Acad Sci. 2005; 1057: 384–392. [DOI] [PubMed] [Google Scholar]
- 30. Rosen R,, Hu D,, Perez V,, et al. Urinary 6-sulfatoxymelatonin level in age-related macular degeneration patients. Mol Vis. 2009; 15: 1673–1679. [PMC free article] [PubMed] [Google Scholar]
- 31. Brunet A,, Bonni A,, Zigmond MJ,, et al. Akt promotes cell survival by phosphorylating and inhibiting a forkhead transcription factor. Cell. 1999; 96: 857–868. [DOI] [PubMed] [Google Scholar]
- 32. Röhlich P,, Van Veen T,, Szél Á. Two different visual pigments in one retinal cone cell. Neuron. 1994; 13: 1159–1166. [DOI] [PubMed] [Google Scholar]
- 33. Peichl L. Diversity of mammalian photoreceptor properties: adaptations to habitat and lifestyle? Anat Rec. 2005; 287: 1001–1012. [DOI] [PubMed] [Google Scholar]
- 34. Ait-Hmyed O,, Felder-Schmittbuhl M-P,, Garcia-Garrido M,, et al. Mice lacking Period 1 and Period 2 circadian clock genes exhibit blue cone photoreceptor defects. Eur J Neurosci. 2013; 37: 1048–1060. [DOI] [PubMed] [Google Scholar]
- 35. Lukáts Á,, Szabó A,, Röhlich P,, Vígh B,, Szél Á. Photopigment coexpression in mammals: comparative and developmental aspects. Histol Histopathol. 2005; 20: 551–574. [DOI] [PubMed] [Google Scholar]
- 36. Tosini G,, Owino S,, Guillaume JL,, Jockers R. Understanding melatonin receptor pharmacology: latest insights from mouse models, and their relevance to human disease. BioEssays. 2014; 36: 778–787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Gresh J,, Goletz PW,, Crouch RK,, Rohrer B. Structure-function analysis of rods and cones in juvenile, adult, and aged C57bl/6 and Balb/c mice. Vis Neurosci. 2003; 20: 211–220. [DOI] [PubMed] [Google Scholar]
- 38. Fernandez-Sanchez L, de Sevilla Muller LP, Brecha NC, Cuenca N. Loss of outer retinal neurons and circuitry alterations in the DBA/2J mouse. Invest Ophthalmol Vis Sci. 2014; 55: 6059–6072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Curcio CA,, Millican CL,, Allen KA,, Kalina RE. Aging of the human photoreceptor mosaic: evidence for selective vulnerability of rods in central retina. Invest Ophthalmol Vis Sci. 1993; 34: 3278–3296. [PubMed] [Google Scholar]
- 40. Cunea A,, Jeffery G. The ageing photoreceptor. Vis Neurosci. 2007; 24: 151–155. [DOI] [PubMed] [Google Scholar]
- 41. Hicks D,, Sabel J. The implications of rod-dependent cone survival for basic and clinical research. Invest Ophthalmol Vis Sci. 1999; 40: 3071–3074. [PubMed] [Google Scholar]
- 42. Mohand-Said S,, Deudon-Combe A,, Hicks D,, et al. Normal retina releases a diffusible factor stimulating cone survival in the retinal degeneration mouse. Neurobiology. 1998; 95: 8357–8362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Léveillard T,, Mohand-Saïd S,, Lorentz O,, et al. Identification and characterization of rod-derived cone viability factor. Nat Genet. 2004; 36: 755–759. [DOI] [PubMed] [Google Scholar]
- 44. Marc RE,, Jones BW,, Watt CB,, Strettoi E. Neural remodeling in retinal degeneration. Prog Retin Eye Res. 2003; 22: 607–655. [DOI] [PubMed] [Google Scholar]
- 45. Komeima K,, Rogers BS,, Lu L,, Campochiaro PA. Antioxidants reduce cone cell death in a model of retinitis pigmentosa. Proc Natl Acad Sci U S A. 2006; 103: 11300–11305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Sengupta A,, Baba K,, Mazzoni F,, et al. Localization of melatonin receptor 1 in mouse retina and its role in the circadian regulation of the electroretinogram and dopamine levels. PLoS One. 2011; 6: e24483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Hardeland R. Melatonin: signaling mechanisms of a pleiotropic agent. BioFactors. 2009; 35: 183–192. [DOI] [PubMed] [Google Scholar]
- 48. Beaulieu J-M,, Tirotta E,, Sotnikova TD,, et al. Regulation of Akt signaling by D2 and D3 dopamine receptors in vivo. J Neurosci. 2007; 27: 881–885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Brunet A,, Datta SR,, Greenberg ME. Transcription-dependent and -independent control of neuronal survival by the PI3K – Akt signaling pathway. Curr Opin Surg. 2001; 11: 297–305. [DOI] [PubMed] [Google Scholar]
- 50. Daitoku H,, Sakamaki JI,, Fukamizu A. Regulation of FoxO transcription factors by acetylation and protein-protein interactions. Biochim Biophys Acta. 2011; 1813: 1954–1960. [DOI] [PubMed] [Google Scholar]
- 51. Jomary C,, Cullen J,, Jones SE. Inactivation of the Akt survival pathway during photoreceptor apoptosis in the retinal degeneration mouse. Invest Ophthalmol Vis Sci. 2006; 47: 1620–1629. [DOI] [PubMed] [Google Scholar]
- 52. Ko ML,, Jian K,, Shi L,, Ko GY-P. Phosphatidylinositol 3 kinase-Akt signaling serves as a circadian output in the retina. J Neurochem. 2009; 108: 1607–1620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Reiter CEN,, Sandirasegarane L,, Wolpert EB,, et al. Characterization of insulin signaling in rat retina in vivo and ex vivo. Am J Physiol Endocr Metab. 2003; 285: E763–E764. [DOI] [PubMed] [Google Scholar]
- 54. Li G,, Anderson RE,, Tomita H,, et al. Nonredundant role of Akt2 for neuroprotection of rod photoreceptor cells from light-induced cell death. J Neurosci. 2007; 27: 203–211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Franke TF,, Hornik CP,, Segev L,, Shostak GA,, Sugimoto C. PI3K/Akt and apoptosis: size matters. Oncogene. 2003; 22: 8983–8998. [DOI] [PubMed] [Google Scholar]
- 56. Brunet A,, Kanai F,, Stehn J,, et al. 14-3-3 transits to the nucleus and participates in dynamic nucleocytoplasmic transport. J Cell Biol. 2002; 156: 817–828. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.