

ABSTRACT
The parthenogenetic all-female marbled crayfish is a novel research model and potent invader of freshwater ecosystems. It is a triploid descendant of the sexually reproducing slough crayfish, Procambarus fallax, but its taxonomic status has remained unsettled. By cross-breeding experiments and parentage analysis we show here that marbled crayfish and P. fallax are reproductively separated. Both crayfish copulate readily, suggesting that the reproductive barrier is set at the cytogenetic rather than the behavioural level. Analysis of complete mitochondrial genomes of marbled crayfish from laboratory lineages and wild populations demonstrates genetic identity and indicates a single origin. Flow cytometric comparison of DNA contents of haemocytes and analysis of nuclear microsatellite loci confirm triploidy and suggest autopolyploidisation as its cause. Global DNA methylation is significantly reduced in marbled crayfish implying the involvement of molecular epigenetic mechanisms in its origination. Morphologically, both crayfish are very similar but growth and fecundity are considerably larger in marbled crayfish, making it a different animal with superior fitness. These data and the high probability of a divergent future evolution of the marbled crayfish and P. fallax clusters suggest that marbled crayfish should be considered as an independent asexual species. Our findings also establish the P. fallax–marbled crayfish pair as a novel paradigm for rare chromosomal speciation by autopolyploidy and parthenogenesis in animals and for saltational evolution in general.
KEY WORDS: Marbled crayfish, Autopolyploidy, Parthenogenesis, Epigenetics, Chromosomal speciation, Saltational evolution
Summary: The triploid marbled crayfish is a rare animal example of speciation by autopolyploidisation and parthenogenesis. It seems to be a particularly suitable model to study how much genetic and epigenetic change is necessary to create a new species.
INTRODUCTION
In the last decade, the marbled crayfish (Marmorkrebs) has gained considerable attention in the scientific community and the public because of its obligatory parthenogenetic reproduction, its suitability as a research model and its high potential as an invasive species (Scholtz et al., 2003; Vogt, 2008a, 2011, 2015a; Jones et al., 2009; Chucholl et al., 2012; Chucholl, 2015; Scholtz, 2015; Faulkes, 2015a). Marbled crayfish is the only known obligatory parthenogen among the ∼15,000 decapod crustaceans. It was discovered in 1995 in the German aquarium trade (Vogt, 2008a) and has become a popular pet in Europe and other continents since then (Chucholl, 2014; Faulkes, 2015b). Thriving wild populations have meanwhile developed from releases in several European countries and Madagascar and are feared to threaten native crayfish species by competition and transmission of the crayfish plague (Jones et al., 2009; Kawai et al., 2009; Chucholl et al., 2012; Keller et al., 2014; Chucholl, 2015).
By comparison of morphological traits and molecular markers, Martin et al. (2010) have identified the sexually reproducing slough crayfish Procambarus fallax from Florida and southern Georgia as the mother species of marbled crayfish. However, its taxonomic position remained unsettled. Martin et al. (2010) suggested the provisional name Procambarus fallax forma virginalis, being aware that forma is not a valid category in animal taxonomy. Meanwhile, several important characteristics of marbled crayfish have been described in detail, including morphology (Kawai et al., 2009), embryonic development (Seitz et al., 2005; Alwes and Scholtz, 2006), life history (Vogt et al., 2004; Seitz et al., 2005; Vogt, 2008b, 2010), parthenogenetic reproduction (Scholtz et al., 2003; Martin et al., 2007; Vogt et al., 2008) and a triploid karyotype (Martin et al., 2015).
Speciation in parthenogenetic lineages is a problematic issue because parthenogens do not fit into the classical concepts of speciation, as discussed in detail by Mayr (1963, 1996), Coyne and Orr (2004), Barraclough et al. (2003), Birky and Barraclough (2009) and Martin et al. (2010). However, Barraclough and colleagues emphasised the importance of understanding diversification and speciation in asexual organisms, not least to test theories about the evolutionary advantage of sex (Barraclough et al., 2003; Birky and Barraclough, 2009). They provided a theory on speciation in asexuals, which they named Evolutionary Genetic Species Concept (Birky and Barraclough, 2009). This theory focuses on the criterion that the individuals of the parent species and the neo-species form discrete clusters of very similar genotypes and phenotypes. The new cluster should be of a single origin and both clusters must be separated from each other by reproductive or geographic isolation and a gap of genetic and phenotypic traits so that natural selection can ensure a divergent evolution over time (Barraclough et al., 2003; Birky and Barraclough, 2009; Birky et al., 2010; Tang et al., 2014).
Stimulated by the paper by Martin et al. (2010) there is an ongoing discussion among marbled crayfish experts whether this animal should be treated as a parthenogenetic lineage of P. fallax or as a species in its own right. In order to examine this issue in detail we have tested the above listed operational definitions for asexual species with several experimental and technical approaches. Cross-breeding experiments between marbled crayfish and slough crayfish and parentage analysis by microsatellite markers were performed to test for reproductive isolation. Complete mitochondrial genomes and nuclear microsatellite patterns of marbled crayfish from several laboratory lineages and wild populations were analysed to clarify single origin and to establish its genotypic characteristics. The DNA content of haemocytes, mitochondrial genome sequences and microsatellite patterns were compared between marbled crayfish, P. fallax and the closely related Procambarus alleni to obtain information about the mode of triploidisation of the marbled crayfish. Global DNA methylation was determined to examine the involvement of epigenetic mechanisms in speciation. Finally, taxonomically relevant morphological characters and ecologically and evolutionarily important life history traits were compared to reveal phenotypic differences between the marbled crayfish and P. fallax clusters.
RESULTS
Crossbreeding experiments and parentage analysis
Crossbreeding experiments were performed to investigate whether marbled crayfish and P. fallax can interbreed and produce viable offspring. Behavioural observations revealed that marbled crayfish females and P. fallax males recognise each other as sexual partners. Courtship and mating behaviour included frontal approach, tearing with the chelipeds, intense sweeping with the antennae, sudden turning of the female and mounting by the male (Fig. 1). This courtship behaviour is also typical of other Procambarus species (Gherardi, 2002). P. fallax males copulated with marbled crayfish females in 15 of 21 trials (71%) and with P. fallax females in 6 of 7 trials (86%) (Table 1). In the marbled crayfish x P. fallax pairs, the first contact was often initiated by the marbled crayfish females. Some matings lasted for more than 1 h. P. fallax males can turn significantly larger marbled crayfish females on the back but are not long enough to simultaneously fix the female's chelipeds and insert the gonopods into the annulus ventralis. P. alleni males copulated neither with P. fallax nor with marbled crayfish females (Table 1) suggesting that they did not recognise them as sexual partners.
Fig. 1.

Mating of a marbled crayfish female with a P. fallax male. The male (top) holds the female firmly with the chelipeds and ischial hooks and his gonopods are plugged into the female's spermatheca.
Table 1.
Crossbreeding experiments between marbled crayfish, P. fallax and P. alleni

We obtained a total of ten clutches from the crossbreeding experiments, eight from crosses of three P. fallax males with eight marbled crayfish females and two from crosses of two P. fallax males with two P. fallax females. Four of the P. fallax x marbled crayfish clutches and one P. fallax x P. fallax clutch developed into juveniles whereas the others decayed during embryonic development. In the P. fallax x P. fallax clutch we counted 10 females and 9 males at juvenile stage 7, reflecting the typical 1:1 sex ratio of sexually reproducing crayfish (Reynolds, 2002). In contrast, in the four marbled crayfish x P. fallax batches the 6, 12, 61 and 93 analysed stage 7 offspring were all females indicating reproduction by parthenogenesis.
The progeny of our crossbreeding experiments were also investigated by microsatellite analysis to further clarify parentage. Microsatellite analysis is an established approach to assess parentage and geographic structuring in crayfish populations and to identify clonal lineages, triploids and hybrids (Walker et al., 2002; Williams et al., 2010; Bai et al., 2011; Thielsch et al., 2012). Of the five primer pairs tested, three revealed PCR products that could be used for fragment length determination in marbled crayfish and P. fallax, namely PclG-02, PclG-04 and PclG-26. PclG-02 and PclG-26 were polymorphic and thus suitable for parentage testing. The microsatellite allele combinations in the analysed family groups of marbled crayfish females 1-4 x P. fallax male 1 were identical between mothers and offspring, namely 267 bp/271 bp/303 bp at locus PclG-02 and 189 bp/191 bp at locus PclG-26, but differed from the allele combination of the male that was 255 bp/267 bp and 185 bp/207 bp, respectively (Table 2). All measurements were repeated at least twice, and in the case of the unusual PclG-02 marker up to five times per specimen. Our data indicate that the male did not contribute to the genome of the offspring and that the progeny is the product of apomictic parthenogenesis. The microsatellite patterns were not only identical between mother and offspring but also between the four batches (Table 2) demonstrating clonality of all marbled crayfish from our laboratory.
Table 2.
Parentage analysis in crossbreeds of marbled crayfish xP. fallax
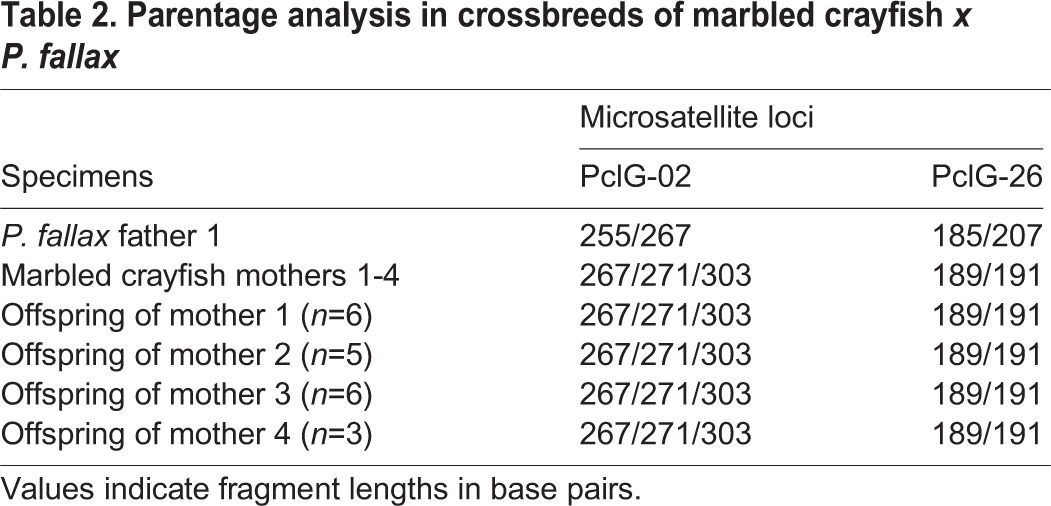
The P. fallax male 1 x P. fallax female 1 family was used as a positive control. Analysis of locus PclG-26 revealed the allele combinations 185 bp/207 bp in the father, 179 bp/185 bp in the mother and 179 bp/185 bp (2×), 179 bp/207 bp (4×), 185 bp/185 bp (4×) and 185 bp/207 bp (4×) in the 14 offspring. These data indicate Mendelian segregation and demonstrate that both parents contributed equally to the genome of the offspring, as is expected for sexually reproducing species.
Single origin and clonality of marbled crayfish populations
For a more detailed genetic analysis of marbled crayfish, we established complete mitochondrial genome sequences of specimens from our Heidelberg and Petshop lineages and from wild populations of Lake Moosweiher (Germany) and Madagascar by high-coverage shotgun sequencing and sequence mapping. Remarkably, these mitochondrial genome sequences were completely identical (Fig. 2), thus confirming the clonal nature of the tested populations and their single origin. Comparison of our sequences with the mitochondrial genome sequence of marbled crayfish published earlier by Shen et al. (2013) revealed six scattered mismatches and major differences in one fragment ranging from position 4600 to 5500. These differences are probably related to technical issues because Shen et al. (2013) used PCR-based methods and primer walking single/double strands sequencing whereas we used next-generation sequencing with a sequencing coverage per nucleotide of >100×.
Fig. 2.

Comparison of complete mitochondrial genomes of marbled crayfish, P. fallax and P. alleni. The sequences of marbled crayfish from two laboratory populations (Heidelberg, Petshop) and two wild populations (Moosweiher, Madagascar) are completely identical. In contrast, the sequences of P. fallax and P. alleni differ in 144 and 1165 SNPs (vertical lines) from marbled crayfish, respectively. Purple bars indicate positions of 12S rRNA and cytochrome oxidase subunit I (COI) fragments that were used for an earlier phylogenetic analysis (Martin et al., 2010).
We also established complete mitochondrial genome sequences for P. fallax and P. alleni. Analysis of the mitochondrial 12S rRNA, 16S rRNA and cytochrome oxidase subunit I genes have earlier indicated a close relationship between marbled crayfish and these species (Scholtz et al., 2003; Jones et al., 2009; Martin et al., 2010). P. alleni occurs sympatrically with P. fallax in many locations in Florida (Hendrix and Loftus, 2000) and was therefore regarded as a candidate that might have contributed to the origination of marbled crayfish by hybridisation with P. fallax (Martin, 2015). Sequence comparison revealed 144 single nucleotide polymorphisms (SNPs) between marbled crayfish and P. fallax but 1165 SNPs between marbled crayfish and P. alleni (Fig. 2). Interestingly, these SNPs were not evenly distributed over the mitochondrial genome, which explains why in the study by Martin et al. (2010) small genetic differences between marbled crayfish and P. fallax were detected in the cytochrome oxidase subunit I gene but not in the 12S rRNA gene. Our results confirm the close genetic relationship between marbled crayfish and P. fallax and a greater distance towards P. alleni.
The single origin and clonality of marbled crayfish from the laboratory and the wild was further confirmed by the analysis of microsatellite loci PclG-02, PclG-04 and PclG-26 in 24 specimens from our laboratory lineages (see parentage analysis), six specimens from a stable wild population in Lake Moosweiher (Chucholl and Pfeiffer, 2010) and one specimen from Madagascar (Jones et al., 2009). All these marbled crayfish showed the same microsatellite patterns, namely the allele associations 267 bp/271 bp/303 bp at locus PclG-02, 159 bp at PclG-04 and 189 bp/191 bp at PclG-26. The fragment lengths of the alleles of locus PclG-02 overlapped in marbled crayfish (267-303 bp) and P. fallax (239-267 bp) but were longer in P. alleni (329-384 bp) and shorter in P. clarkii (211-228 bp). Marbled crayfish shared two of six alleles with P. fallax, namely 267 bp at locus PclG-02 and 159 bp at locus PclG-04, but none with the other species thus confirming the particularly close relationship between P. fallax and marbled crayfish.
Ploidy status of marbled crayfish
Martin et al. (2015) recently used karyological analysis to demonstrate that marbled crayfish has a triploid genome. Our microsatellite analysis confirms this finding. Marbled crayfish generally have the allele association 267 bp/271 bp/303 bp at locus PclG-02 (Fig. 3A), whereas P. fallax, P. alleni and P. clarkii have one or two alleles at this locus, which is consistent with diploidy and sexual reproduction. In an earlier paper, Martin et al. (2007) have also analysed locus PclG-02 and reported only two alleles of 267 bp and 271 bp. However, a recent re-examination of their material confirmed the presence of the third 303 bp allele (G. Scholtz, personal communication).
Fig. 3.
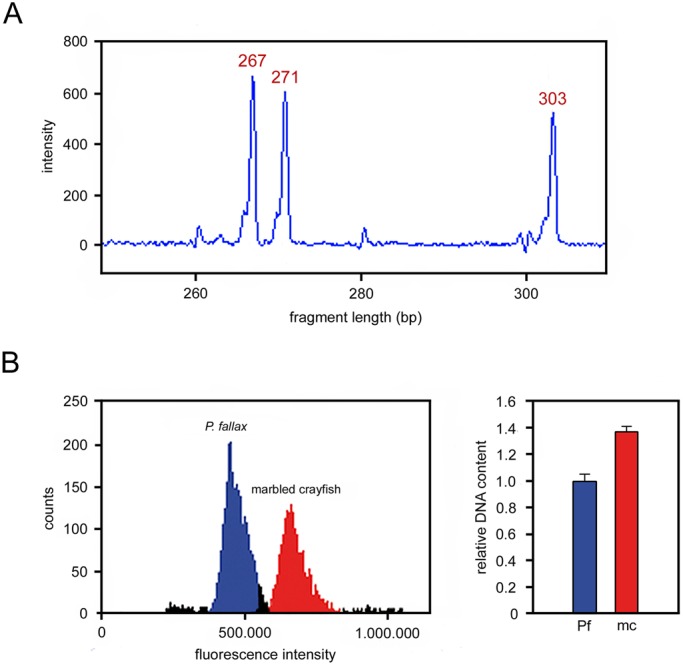
Ploidy status of the marbled crayfish genome. (A) Microsatellite locus PclG-02 in marbled crayfish showing a combination of three alleles of 267 bp, 271 bp and 303 bp fragment length. (B) Flow cytometry of haemocytes of P. fallax (Pf) and marbled crayfish (mc) revealing an approximately 1.4 fold increased DNA content in marbled crayfish. The right panel shows the means±standard deviations (s.d.) of two biological and three technical replicates. Differences are highly significant (P=1.33×10−7, Welsh two-sided t-test).
We further corroborated triploidy in marbled crayfish by flow cytometric measurement of the DNA content of haemocytes in marbled crayfish and P. fallax. Haemocytes are particularly suitable for this purpose because they are devoid of somatic polyploidisation (Allen, 1983). Our results showed a significant 1.4-fold higher DNA content in the blood cells of marbled crayfish (Fig. 3B), which is consistent with triploidy.
Comparison of DNA methylation between marbled crayfish and Procambarus fallax
In order to test if the marbled crayfish and P. fallax clusters also differ with respect to epigenetic markers we determined global DNA methylation by mass spectrometry in identically raised and age and size-matched representatives of both crayfish. DNA methylation represents a widely conserved epigenetic mark that is often associated with polyphenism and adaptive phenotypic changes (Jaenisch and Bird, 2003; Lyko and Maleszka, 2011). Comparison of three whole juveniles and selected organs (hepatopancreas, abdominal musculature and ovary) of three adults revealed a consistently and highly significantly reduced level of DNA methylation in marbled crayfish when compared to P. fallax (Fig. 4). The ten P. fallax samples together had a DNA methylation level of 2.93±0.15% (mean±standard deviation) whereas the ten marbled crayfish samples together had a level of only 2.40±0.08%. These results suggest that marbled crayfish have a considerably different DNA methylation pattern.
Fig. 4.
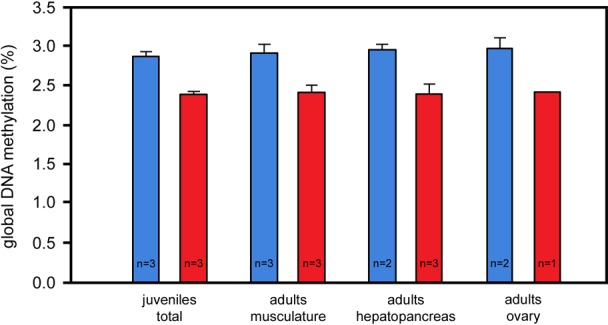
Differences in global DNA methylation between marbled crayfish (red) and P. fallax (blue). Three complete juveniles and major organs of three adult females were analysed in each crayfish. Note the consistently and significantly greater methylation levels in P. fallax (P=1.48×10−7 for the sum of all samples, Welsh two-sided t-test). Error bars represent s.d.
Comparison of morphological characters between marbled crayfish and P. fallax
Comparison of the most relevant taxonomic characters of cambarid females (Hobbs, 1972, 1981, 1989) between marbled crayfish and P. fallax corroborated the high degree of morphological similarity between the two crayfish as previously established by Kawai et al. (2009) and Martin et al. (2010). The diagnostically most meaningful trait in females of the genus Procambarus is the annulus ventralis, which is bell-shaped with a tilted S-shaped sinus in both marbled crayfish and P. fallax (Fig. 5A,B). This typical form is not found in other Procambarus species (Hobbs, 1989) as best exemplified by the differently shaped sperm receptacle of the closely related P. alleni (Fig. 5C). The areola, an unpaired structure on the dorsal midline of the carapace, is also very similar in marbled crayfish and P. fallax with respect to shape and length-to-width proportion (Fig. 5D,E). The same holds for the cheliped chelae, which closely resemble each other in both crayfish with respect to shape, dentation and setation (Fig. 5F,G), and the coloration pattern, which consists of distinct marmorated spots and dark dorsolateral stripes on the carapace and pleon (Fig. 5H,I). Size, form and coloration of the marmoration spots are highly variable not only in the sexually reproducing P. fallax but also in the genetically uniform marbled crayfish, which is a result of stochastic developmental variation (Vogt et al., 2008; Vogt, 2015b).
Fig. 5.

Comparison of morphological characters between marbled crayfish and P. fallax. (A) Annulus ventralis from exuvia of marbled crayfish. (B) Annulus ventralis of P. fallax. (C) Annulus ventralis of P. alleni. Note striking structural difference to sperm receptacles of marbled crayfish and P. fallax. (D) Areola of marbled crayfish. (E) Areola of P. fallax. (F) Left cheliped of marbled crayfish of 8.4 cm TL. (G) Left cheliped of P. fallax female of 4.7 cm TL. Form, dentation and setation of the chelae are very similar in both species. (H) Coloration of cephalothorax in marbled crayfish. (I) Coloration of cephalothorax in P. fallax male (photo: C. Lukhaup).
Comparison of life history traits between marbled crayfish and P. fallax
In contrast to the morphological characters, life history features like growth and fecundity are markedly different between marbled crayfish and P. fallax. Fig. 6 gives an example for differences in the speed of growth between identically raised laboratory populations of the same age. At day 250 after hatching, when the first females in both crayfish had reached sexual maturity, mean body weight was almost twice as large in marbled crayfish as in P. fallax females.
Fig. 6.
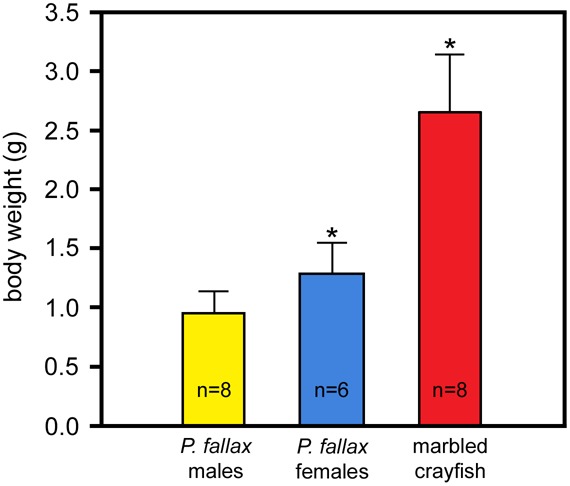
Comparison of growth between marbled crayfish and P. fallax. The three groups were reared for 250 days at 20°C under identical conditions and fed with the same food ad libitum. The differences between marbled crayfish and P. fallax females are highly significant (asterisks; P=2.06×10−5; Welsh two-sided t-test). Error bars represent s.d.
Maximum body and clutch sizes were also markedly higher in marbled crayfish. The largest specimen in our laboratory had a carapace length (CL) of 4.9 cm, a total length (TL) of 10.3 cm and a body weight of 30.1 g (Fig. 7A). In the wild, the largest of the 1084 marbled crayfish measured (Jones et al., 2009; Kawai et al., 2009; Chucholl and Pfeiffer, 2010; M. Pfeiffer and C. Chucholl, personal communication) was found in Lake Moosweiher and had a CL of 4.9 cm and a weight of 32.0 g (Chucholl and Pfeiffer, 2010). In contrast, the largest of the 4710 wild P. fallax examined (Hobbs, 1942, 1981; Hendrix et al., 2000; C. van der Heiden, PhD thesis, Florida Atlantic University, 2012) had a CL of only 3.4 cm, corresponding to a TL of 7.4 cm and a weight of approximately 11.5 g. The largest clutches of marbled crayfish in the laboratory and the wild consisted of 731 eggs (Fig. 7B) and 724 eggs (Chucholl and Pfeiffer, 2010), respectively, which is 5.6 fold higher than the largest clutch of 130 eggs reported for P. fallax in literature (Hendrix et al., 2000; P. G. VanArman, PhD thesis, Nova Southeastern University, 2003). The analysis of life history features of the slough crayfish by van der Heiden (PhD thesis, Florida Atlantic University, 2012) corroborated that P. fallax reaches only rarely a size of more than 6.5 cm TL.
Fig. 7.

Comparison of body size and fecundity between marbled crayfish and P. fallax. (A) Largest marbled crayfish from our laboratory having a total length of 10.3 cm. (B) Clutch of same specimen consisting of 731 eggs. (C) Differences in carapace length between populations of ovigerous marbled crayfish (mc) and P. fallax females (PF) from comparable climatic regions. Data for marbled crayfish (n=57) was obtained in Madagascar (Jones et al., 2009) and data for P. fallax (n=27) was obtained in Florida (Hendrix et al., 2000). Horizontal bars indicate ranges and vertical lines indicate mean values (m) and lower and upper range limits. The difference between marbled crayfish and P. fallax females is highly significant as indicated by the P-value. (D) Differences in clutch size between the same populations as in (C). The difference is highly significant as indicated by the P-value. For statistical calculations, the s.d. was taken as half the range, and a Bonferroni adjustment for multiplicity was applied.
The differences in growth and fecundity between marbled crayfish and P. fallax were also confirmed by the re-analysis of published data for egg-carrying females from comparable climatic regions. Ovigerous marbled crayfish from Madagascar had a mean CL of 3.5 cm, a mean TL of 7.4 cm and a mean clutch size of 300 eggs (Jones et al., 2009), whereas ovigerous P. fallax from the Everglades National Park in Florida had a mean CL of 1.8 cm, a mean TL of 3.8 cm and a mean clutch size of 41 eggs only (Hendrix et al., 2000), indicating that body size and fecundity is significantly increased in marbled crayfish (Fig. 7C,D). These findings identify important phenotypic differences between marbled crayfish and P. fallax that have not been recognised previously.
DISCUSSION
Our results demonstrate that marbled crayfish meets all the criteria for asexual speciation, as formalised by Barraclough et al. (2003) and Birky and Barraclough (2009): 1) The marbled crayfish is separated from its mother species, P. fallax, by reproductive isolation, significant genomic and epigenetic differences and superior life history traits; 2) Our data support a single origin of all marbled crayfish; 3) All populations known to date live outside the natural range of P. fallax, suggesting geographical isolation; and 4) All marbled crayfish populations are unified in one cluster by common phenotypic, genetic and epigenetic characteristics, despite their broad geographical distribution. These commonalities and the differences towards P. fallax make it very likely that the marbled crayfish and slough crayfish clusters will evolve differently, which is the main criterion for erecting an asexual species (Birky and Barraclough, 2009). As such, marbled crayfish should be considered as an independent species for which we propose the name Procambarus virginalis, as previously suggested by Martin et al. (2010). The formal description as a new species will be detailed in a separate publication.
Marbled crayfish appeared first in 1995 in the German aquarium trade. Thereafter, aquarists have propagated it in captivity, and since about 2003, releases have resulted in the establishment of thriving wild populations in Central Europe and Madagascar (Jones et al., 2009; Kawai et al., 2009; Chucholl and Pfeiffer, 2010; Chucholl et al., 2012; Chucholl, 2015; Faulkes, 2015a). The ‘mega-population’ (Martin, 2015) in innumerable aquarium tanks on various continents and the known wild populations are apparently all descendants of the single clone or single individual that was introduced in Germany in 1995. Our results confirm this single origin by demonstrating the identity of the mitochondrial genomes and microsatellite patterns in samples of captive and wild populations. One of the samples analysed in our study, the Heidelberg specimen, can be directly traced back to the year 1995 and to the oldest marbled crayfish for which written records exist (F. Steuerwald, personal communication).
It is unknown whether marbled crayfish emerged in the natural range of P. fallax or in captivity. Scholtz (2015), Faulkes (2015a) and Martin (2015) summarised possible scenarios for the first alternative including hybridisation with coexisting Procambarus species and geographic parthenogenesis. These authors and Chucholl (2015) also stressed that in captivity there were many more candidates for hybridisation than the naturally coexisting six Procambarus species (Hobbs, 1942, 1981) because crayfish were popular pets already in the 1990s. Faulkes (2015a) emphasised that all surveys on P. fallax in Florida and southern Georgia revealed males and females arguing against the presence of marbled crayfish populations in the natural range of P. fallax. Moreover, none of the publications on wild P. fallax (Hobbs, 1942, 1981; Hendrix and Loftus, 2000; Hendrix et al., 2000; P. G. VanArman, PhD thesis, Nova Southeastern University, 2003; C. van der Heiden, PhD thesis, Florida Atlantic University, 2012) mentioned specimens above 7.4 cm TL, which would again argue for the absence of primary populations of marbled crayfish. In sympatric populations, small and medium-sized marbled crayfish and P. fallax females would be indistinguishable by morphological criteria alone. However, marbled crayfish could now be identified by the use of genetic markers. Particularly useful is the highly specific tri-allelic microsatellite locus PclG-02, which could be assayed in large samples with reasonable expenditure. However, time for the detection of primary populations may be limited because marbled crayfish are already widespread in American aquaria (Faulkes, 2015b) and their release into the natural range of P. fallax would render the search for primary populations of marbled crayfish impossible.
Our crossbreeding experiments with marbled crayfish, P. fallax and P. alleni revealed that marbled crayfish and P. fallax still recognise each other as sexual partners but not marbled crayfish and P. alleni. Recognition of sexual partners in crayfish is mainly based on chemical signatures of the urine but may also include visual and tactile cues (Gherardi, 2002; Reynolds, 2002). Marbled crayfish and P. fallax copulate readily with each other but the progeny of such matings are pure marbled crayfish resulting from parthenogenesis. These findings demonstrate reproductive isolation and suggest that the reproductive barrier is set at the cytogenetic rather than the behavioural level. Mechanical barriers can be largely excluded because the sperm receptacles are structurally very similar in marbled crayfish and P. fallax as shown in Fig. 5 and by Martin et al. (2010). Moreover, we have repeatedly observed the insertion of the P. fallax male gonopods into the annulus ventralis of marbled crayfish. We attempted to directly prove sperm transfer by examining moulted sperm receptacles of females that had successfully produced offspring. However, we did not find any sperm remnants in these marbled crayfish or P. fallax females.
The morphological features and microsatellite patterns strongly suggest that marbled crayfish originated by autopolyploidisation and not by hybridisation with a closely related species, which is by far the most frequent cause of triploidy in animals (Leggatt and Iwama, 2003; Mallet, 2007; Abbott et al., 2013; Choleva and Janko, 2013). Fertile hybrids between two species and backcrosses of the hybrids with the parental species were occasionally observed in wild cambarid crayfish inclusive of the genus Procambarus (Cesaroni et al., 1992; Perry et al., 2001a; Zuber et al., 2012). Such hybrids are clearly recognisable because of their intermediate morphological characters and genetic signatures (Perry et al., 2001b; Zuber et al., 2012). However, marbled crayfish do not show morphological hybrid features (Kawai et al., 2009; Martin et al., 2010; this study) and there is also no evidence for hybridisation on the genetic level and no strong bias towards heterozygosity in the microsatellite pattern, which would be typical for hybrids (Soltis and Soltis, 2000; Alves et al., 2001). Of the seven microsatellite loci that were investigated in marbled crayfish so far, three were homozygous and four were heterozygous (Martin et al., 2007; Vogt et al., 2008; this study), thus largely excluding allopolyploidisation for marbled crayfish. Furthermore, Martin and colleagues have recently shown that the nuclear elongation factor 2 (EF-2) genes are identical in marbled crayfish and P. fallax but differ from other Procambarus species like P. alleni, P. clarkii, P. acutus and P. liberorum (Martin et al., 2015). These findings provide additional support for the origin of marbled crayfish by autopolyploidisation.
The combination of three different alleles per locus, as observed for PclG-02 in marbled crayfish, could be most easily explained by hybridisation. However, this pattern can also occur in autopolyploids, namely when an unreduced diploid egg with dimorphic alleles is fertilised by a sperm from the same species, or alternatively, by simultaneous fertilisation of a haploid egg by two sperms with different alleles. In shrimp, fish and bivalve aquaculture, such autopolyploid triploids are artificially produced by the prevention of polar body I extrusion in fertilised eggs either by an abrupt temperature shock or chemicals like 6-dimethylaminopurine (Sellars et al., 2006; Piferrer et al., 2009). These triploidisation experiments point to the possibility that marbled crayfish may have arisen in a captive P. fallax female by an accidental heat or cold shock during a sensitive phase of egg development.
The origin of obligatory parthenogenesis in marbled crayfish is probably a by-product of triploidisation but the causal relationship of polyploidy and parthenogenesis is not yet understood (Neiman et al., 2005; Martin, 2015). Infectious parthenogenesis by the feminising bacterium Wolbachia, which is proven for other crustaceans (Cordaux et al., 2012), was excluded by the use of molecular probes for the parasite (Vogt, 2008a). In plants, it was shown that polyploidy per se can have an immediate impact on the reproductive biology of a species (Levin, 2002). In animals, however, obligatory parthenogenesis is relatively rare. It has been described in a few vertebrate hybrids (Cuellar, 1974; Cole et al., 2010) and in several invertebrate taxa such as bdelloid rotifers, ostracods, cladocerans, mites, grass thrips, stick insects and the gastropod Potamopyrgus antipodarum (Suomalainen et al., 1987; Mark Welch and Meselson, 2000; Birky and Barraclough, 2009; Schwander and Crespi, 2009; Neiman et al., 2011; van der Kooi and Schwander, 2014). In most of these cases, the parthenogens are either diploid or allopolyploid. The combination of parthenogenesis and autopolyploidy is much less frequent. Fertile autopolyploid parthenogens are convincingly documented for the freshwater snail Potamopyrgus antipodarum (Neiman et al., 2011) and are suspected for some polyploid populations of ostracods (Little and Hebert, 1997), brine shrimp (Zhang and King, 1992) and scale insects (Gavrilov-Zimin et al., 2015). The polyploid and obligatory parthenogenetic cladoceran lineages are apparently all of hybrid origin (Dufresne and Hebert, 1995; Dufresne, 2011; Thielsch et al., 2012), which distinguishes them from marbled crayfish. Furthermore, the artificially produced autopolyploid shrimp and fish are usually sterile (Sellars et al., 2013).
Polyploids often have life history traits that are different from those of the parent species (Xiang et al., 2006; Lavania et al., 2012; Cohen et al., 2013; Krois et al., 2013). Growth, number of offspring and other quantitative traits can either be decreased or increased when compared to the diploid ancestors suggesting that polyploidy can alter life history traits per se. In marbled crayfish, growth, maximum body size and fecundity were significantly increased when compared to P. fallax, whereas the time of sexual maturity was similar (Hobbs, 1981; Jones et al., 2009; Chucholl and Pfeiffer, 2010; C. van der Heiden, PhD thesis, Florida Atlantic University, 2012; this study). Longevity may also be increased in marbled crayfish. Maximum age so far recorded is 1610 days in marbled crayfish (Vogt, 2010) and 980 days in P. fallax (Z. Faulkes, personal communication). In allopolyploids, the increase of life history traits is usually explained as the result of heterozygosity, which is well known as heterosis effect or hybrid vigor (Comai, 2005; Soltis, 2013). This explanation is not applicable for autopolyploids because autopolyploidisation enhances only the copy number of already existing genes. However, novel traits do not necessarily require new genes or new developmental pathways to come into being but can instead arise from recruitment of already existing developmental processes into new contexts (West-Eberhard, 2003; Moczek, 2009). Thus, trait alteration in marbled crayfish may have been caused by altered gene dosage, the rearrangement of gene-networks and the modulation of gene expression by changes in epigenetic regulation.
Changes in epigenetic regulation during transition from P. fallax to marbled crayfish can be deduced from the significantly reduced level of global DNA methylation in marbled crayfish. DNA methylation is an epigenetic mechanism that considerably affects plant and animal phenotypes (Jaenisch and Bird, 2003; Verhoeven et al., 2010; Lyko and Maleszka, 2011). It is responsive to environmental and genomic stresses including polyploidisation (Jaenisch and Bird, 2003) and might thus contribute to speciation in polyploids. In plants, the increase or reduction of global DNA methylation after autopolyploidisation is well known (Soltis et al., 2007; Li et al., 2011). It is also well established that DNA methylation and other epigenetic mechanisms contribute to the establishment of reproductive barriers (Durand et al., 2012; Lafon-Placette and Köhler, 2015) and the expression of hybrid vigour in allopolyploid plants (Gao et al., 2014). In marbled crayfish, epigenetic mechanisms may thus have been involved in the acquisition of novel fitness traits.
Chen et al. (2007) reported that polyploidisation is often accompanied or followed by intense rearrangements in the genome, which stabilise the new lineage. These rearrangements, which are associated with epigenetic changes, can include loss of DNA. For example, in synthetic autopolyploids of annual phlox, Phlox drummondii, an immediate loss of 17% of total DNA has been observed with a further reduction of up to 25% upon the third generation (Parisod et al., 2010). Such mechanisms may also have operated during transition from P. fallax to marbled crayfish and might explain why triploid marbled crayfish have only a 1.4-fold rather than the expected 1.5-fold increased DNA content when compared with its diploid mother species.
Marbled crayfish are successful invaders of freshwater ecosystems in Europe and Madagascar threatening native crayfish populations (Jones et al., 2009; Chucholl and Pfeiffer, 2010; Chucholl et al., 2012; Chucholl, 2015). Chucholl (2015) calculated an almost double FI-ISK (Freshwater Invertebrate Invasiveness Scoring Kit) score for marbled crayfish when compared to P. fallax, making it a high risk species for Central Europe. Moreover, Feria and Faulkes (2011) predicted with climate and habitat based Species Distribution Models that marbled crayfish could inhabit a larger geographical area than its mother species P. fallax when released in the southern states of the USA, thus illustrating the ecological superiority of marbled crayfish. This high invasive potential probably results from the enhanced fitness traits discussed above and from the saved costs for the production of males, courtship, mating and meiosis.
Evolutionary theory predicts short-term success of parthenogenetic lineages but their long-term extinction (Bell, 1982; Neiman et al., 2010; Schwander et al., 2011). The early success is usually explained by the saved costs for male production that can be used for increased growth and fecundity. The long-term extinction is explained by the accumulation of deleterious mutations (Muller's Rachet Hypothesis) and the absence of genetic variability, which impedes resistance against parasites and diseases (Red Queen Hypothesis) and adaptation to complex and changing environments (Tangled Bank Hypothesis) (Butlin, 2002; Henry et al., 2012; Martin, 2015). The short-term success for marbled crayfish is already proven by its rapid expansion (Jones et al., 2009; Chucholl et al., 2012), even though the sustainability of this expansion remains to be determined. The dead-end scenario for parthenogenetic lineages seems to apply to most animal parthenogens but not to all. Evolutionarily successful obligatory parthenogenetic examples are the bdelloid rotifers and darwinulid ostracods, often called ‘evolutionary scandals’, that propagated without sex for about 80-100 million years and generated 360 and 28 species, respectively (Butlin, 2002; Mark Welch et al., 2009; Schön et al., 2009). Furthermore, several parthenogenetic Timema stick-insect lineages have persisted without sexual recombination for more than a million generations (Schwander et al., 2011). Long-term persistence of parthenogens may be achieved by epigenetic silencing of detrimental mutations and the generation of epigenetic diversity from the same genome, allowing suppression of parasites and environmental adaptation despite genetic uniformity. This epigenetic diversity may be genetically assimilated over time finally leading to a new species (Vogt, 2015b).
In the literature, it is controversially discussed whether obligatory parthenogenetic lineages should be taxonomically considered with their parent species or be regarded as independent species (Soltis et al., 2007). According to the Biological Species Concept (Mayr, 1963) one of the most important criteria of speciation is reproductive isolation. However, this criterion is not applicable for asexuals, because the descendants of each female would have to be regarded as a new species. An asexual species should only be erected if all of the criteria specified by Birky and Barraclough (2009) for asexual speciation are met. These are reproductive and/or geographic isolation, single origin of all cluster members, phenotypic, genetic and epigenetic similarity of cluster members and separation from the sexually reproducing mother species by a broad gap in these characteristics. If regular backcrosses occur between the obligatory parthenogens and their parent-species, as is the case in waterflea Daphnia pulex (Xu et al., 2015), parthenogenetic lineages should not be considered as independent species. Sometimes, a clear taxonomic decision is avoided by summarising the sexually reproducing parent species and their asexual descendants in a ‘species complex’, particularly in hybridising and backcrossing assemblages. Examples are the Daphnia pulex complex, which includes allopolyploid obligatory parthenogenetic lineages and their diploid mother species (Dufresne, 2011), and the Carassius auratus complex, which includes diploid, triploid and polyploid fish (Bai et al., 2011). In ancient asexuals, the consideration of the various parthenogenetic lineages as species is common and cryptic species are increasingly identified (Heethoff et al., 2007; Mark Welch et al., 2009; Schön et al., 2009; Schwander et al., 2011). For example, Schön et al. (2012) successfully applied Birky and Barraclough's Evolutionary Genetic Species Concept to delineate species and to identify cryptic species among morphospecies in obligate parthenogenetic ostracods.
Polyploids and parthenogens are regarded as particularly suitable for the investigation of several puzzling questions in evolutionary biology like the sex paradox (Bell, 1982), the invasion paradox (Sax and Brown, 2000) and the role of epigenetics in speciation of clonal organisms (Vogt, 2015b). Neiman and Schwander (2011) emphasised that general insights into the advantages of sex can be generated by taking advantage of parthenogenetic taxa that differ in characteristics such as meiotic versus mitotic offspring production, single versus multiple origin, hybrid versus non-hybrid origin, and ploidy level. They mainly used the snail Potamopyrgus antipodarum as a model to investigate why sexual reproduction is extremely widespread in spite of its presumed costs and how is it maintained (Neiman et al., 2005, 2010; Neiman and Schwander, 2011). The advantage of this model is the co-occurrence of sexually reproducing diploids with parthenogenetic autotriploids of multiple origins. Similarly, the Daphnia pulex complex, which includes sexual reproducers, cyclic parthenogens and obligatory parthenogens, has proven particularly valuable to study the adaptation of sexual and asexual breeding systems to habitat and geographical latitude and to identify the genes and molecular mechanisms that are responsible for the sexual-to-asexual transition (Hebert and Finston, 2001; Eads et al., 2012; Tucker et al., 2013). In contrast to these examples, the P. fallax-marbled crayfish pair includes only sexually reproducing diploids and autotriploid parthenogens of single origin and genetic identity. This model seems to be particularly suitable to investigate chromosomal speciation, the involvement of epigenetic mechanisms in speciation, and the invasion paradox (Vogt, 2015b). The latter addresses why small genetically uniform or depauperate exotic groups can successfully conquer new environments and displace well-adapted native species (Sax and Brown, 2000).
Speciation by autopolyploidisation as observed in the marbled crayfish is a special case of chromosomal speciation that is well-known in plants (Soltis et al., 2007) but rare in animals. Chromosomal speciation is a complementary concept to the better known speciation by changes in allele frequency distribution and can result in the almost instantaneous production of new species and phenotypic novelty within one generation (King, 1993; Faria and Navarro, 2010; de Storme and Mason, 2014). This ‘saltational speciation or ‘saltational evolution’ (Theißen, 2009; Rubinoff and Le Roux, 2008; Minelli, 2015) has largely been ignored by gradualism-based Modern Synthesis, which may be due to its rarity in animals, the lack of mechanistic understanding and the dearth of suitable models. Marbled crayfish represents a contemporary animal example of autopolyploid speciation, which likely started about 20-30 generations ago. Comparative whole-genome and epigenome sequencing approaches will be required to disentangle the contribution of genetic and epigenetic changes in the speciation of marbled crayfish and to monitor future genetic and epigenetic diversification.
CONCLUSION
Marbled crayfish can be regarded as a new species that originated from P. fallax by triploidisation and concomitant epigenetic alterations, as shown by our combined morphological, behavioural, genetic and epigenetic study. Marbled crayfish is morphologically very similar to its mother species but has superior fitness traits. Genetic data suggest an instantaneous speciation by autopolyploidisation and parallel change of the mode of reproduction from gonochorism to parthenogenesis. The young evolutionary age of marbled crayfish, which is possibly less than three decades, may offer the possibility to identify key events for this type of speciation. The combination of autopolyploidy and obligate parthenogenesis is common in plants but very rare in animals. Thus, the P. fallax-marbled crayfish pair provides an interesting new model system to study asexual speciation and saltational evolution in animals and to determine how much genetic and epigenetic change is necessary to create a new species.
MATERIALS AND METHODS
Animals
The following animals were used: (1) marbled crayfish Procambarus fallax (Hagen, 1870) f. virginalis from our laboratory lineages named ‘Heidelberg’ and ‘Petshop’ and from two wild populations in Germany and Madagascar, (2) Procambarus fallax (Hagen, 1870) from our laboratory population and the aquarium trade, (3) Procambarus alleni (Faxon, 1884) from the aquarium trade, and (4) Procambarus clarkii (Girard, 1852) from an invasive Swiss population. The Heidelberg lineage was founded by G.V. in February 2003 from a single female, which originated from the oldest documented marbled crayfish aquarium population founded in 1995 by F. Steuerwald. The Petshop lineage was established by G.V. in February 2004 from a single female purchased in a pet shop. The wild marbled crayfish were from Lake Moosweiher, Germany (provided by M. Pfeiffer), and a market in Antananarivo, Madagascar (provided by F. Glaw). Our P. fallax laboratory population was founded in February 2014 by a single pair obtained from the aquarium trade. All laboratory crayfish were raised under the same conditions. Animals were kept either individually or communally in plastic containers of 30×25×20 cm equipped with gravel and shelters. Tap water was used as the water source and replaced once a week. Water temperature was maintained at 20°C. All animals and life stages were fed with TetraWafer Mix pellets. All crayfish experiments were performed by approval of the institutional animal welfare committee, in compliance with local standards and guidelines.
Cross-breeding experiments
For the 38 crossbreeding experiments we used three P. fallax males with total lengths (TL=tip of rostrum to end of telson) of 3.1-5.2 cm, five P. fallax females with TLs of 3.5-4.2 cm, 14 marbled crayfish females with TLs of 4.0-6.3 cm and two P. alleni males with TLs of 5.1-5.3 cm. All males were in the reproductively competent Form I as indicated by the presence of hooks on the ischia of the 3rd and 4th peraeopods. Eight of the 14 marbled crayfish females and 4 of the 5 P. fallax females had well-developed glair glands on the underside of the pleon indicating ovarian maturity and receptiveness. The behavioural experiments were performed in aquaria with an area of 26×16 cm without shelter. Pairs were observed for 2 h and copulation was regarded as successful when the partners remained in typical copulation position for more than 10 min. Parentage of the offspring was determined by microsatellite analysis.
Microsatellite analysis
For microsatellite analysis, walking legs of specimens were fixed in 80% ethanol prior to extraction of nuclear DNA with the Blood & Cell Culture DNA Kit (Genomic Tips) from Qiagen (Hilden, Germany). A total of five microsatellite primer pairs were tested. Four of them were originally designed for P. clarkii (PclG-02, PclG-04, PclG-08, PclG-48) (Belfiore and May, 2000) and one pair (PclG-26) was designed for marbled crayfish based on the P. clarkii sequences (Vogt et al., 2008). The same microsatellite loci were additionally investigated in P. alleni and P. clarkii. PCR was carried out using a Primus 96 Cycler (Peqlab Biotechnologie, Erlangen, Germany). Fragment analysis was performed on a Beckman Coulter CEQ 8000 eight capillary sequencer (Beckman Coulter, Krefeld, Germany) using the Beckman Coulter DNA Size Standard Kit 400 bp. Loci were scored with GeneMarker, v.2.6 (SoftGenetics, State College, PA, USA).
Sequencing, assembly and comparison of mitochondrial genomes
For comparison of complete mitochondrial genomes we used two cultured marbled crayfish from the Heidelberg and Petshop lineages, two wild marbled crayfish from Lake Moosweiher and Madagascar, one P. fallax female and one P. alleni female. DNA was isolated from hepatopancreases and abdominal musculature as described above and sequenced on an Illumina HiSeq platform. Read pairs were quality trimmed (quality value ≥30, minimum length ≥30) and the mitochondrial genome of the Heidelberg animal was assembled by Velvet 2.0 (Zerbino and Birney, 2008). The sequences of the other specimens were established by mapping against the Heidelberg sequence using Bowtie 2 (Langmead and Salzberg, 2012). For the identification of single nucleotide polymorphisms (SNPs) between the marbled crayfish populations, we used mpileup and bcftools from SAMtools (Li et al., 2009), requiring a quality value >30 for SNP calling. Mitochondrial genome sequences of P. fallax and P. alleni were generated by MITObim 1.6 (Hahn et al., 2013) using published mitochondrial DNA fragments from P. fallax (FJ619800) and P. alleni (HQ171462, FJ619802, HQ171451) as seed sequences. Mismatches in comparison to marbled crayfish sequences were identified by blastn alignments.
Measurement of DNA content by flow cytometry
Flow cytometry was used to determine the DNA content in haemocytes of P. fallax and marbled crayfish. Haemolymph was withdrawn through the articulating membrane between coxa and basis of the chelipeds, mixed 1:1 with crayfish anticoagulant buffer solution (100 mM glucose, 34 mM trisodium citrate, 26 mM citric acid, 15.8 mM EDTA, pH 4.6) and centrifuged for 5 min at 1400 rpm. The pellet was washed and re-suspended with 100 µl PBS. Samples were either stored in 10% DMSO at −80°C or immediately used for analysis of the DNA content. For flow cytometry, 4 µl RNase A (Sigma-Aldrich, Munich, Germany) stock solution (50 mg/ml) was added to the samples and incubated for 5 min at room temperature followed by an incubation for 60 min with 5 µl propidium iodide (Life Technologies, Darmstadt, Germany) stock solution (1 mg/ml). The samples were then mixed 1:1 with PBS and the DNA-related fluorescence intensities of single cells were measured on a BD Accuri C6 Cytometer (BD Sciences, Heidelberg, Germany) with blue laser 488 nm and detection filter FL2 585/40 nm.
Measurement of global DNA methylation by mass spectrometry
Global DNA methylation was determined in three whole juveniles and selected tissues (hepatopancreas, abdominal musculature and ovary) of three adults of marbled crayfish and P. fallax. Sample preparation and LC-MS/MS analyses were conducted as previously described (Kellner et al., 2014) and were performed on an Agilent 1260 LC system connected to an Agilent 6460 TripleQuad mass spectrometer (Agilent, Böblingen, Germany). Briefly, after enzymatic hydrolysis to nucleosides, the samples were spiked with 250 fmol [D3]-5-methylcytosine as internal standard. The mass transitions resulting from the loss of desoxyribose (5-methylcytidine: 242 Th→126 Th, [D3]-5-methylcytidine: 245 Th→129 Th) by collision induced dissociation (CID) were analysed in dynamic multiple reaction monitoring mode (DMRM). Calibration curves using a stable isotope labelled internal standard were established for quantification of 5-methylcytidine. The linear regressions resulting from the double logarithmic plots were used to correlate the respective signals from LC-MS/MS analysis to known amounts of substance. The yield of detected modification was normalised to guanosine content (as equivalent to cytidine content) because of better signal quality. To assess the amount of guanosine, the areas of the DAD results, gained during the LC analysis, were correlated to their respective amounts of substance in the same way as above.
Investigation of morphological characters and life history traits
For qualitative comparison of morphological characters between marbled crayfish and P. fallax we used 18 marbled crayfish with TLs of 4.0-8.4 cm and body weights of 1.4-15.2 g and 12 P. fallax females with TLs of 3.6-5.7 cm and weights of 1.1-4.5 g. We focussed on comparison of annulus ventralis (sperm receptacle), areola of the carapace, cheliped chelae and coloration, the taxonomically most relevant characters in female Cambaridae (Hobbs, 1972, 1981, 1989). For quantitative comparison of life history traits we analysed growth and time of sexual maturity in eight marbled crayfish, six P. fallax females and eight P. fallax males from our laboratory. Each group consisted of batch mates and was communally raised for 250 days under identical conditions. Growth was determined by measurement of carapace length (CL), total length (TL) and body weight. Sexual maturity was deduced from the presence of glair glands. Additionally, we re-analysed published body sizes and clutch sizes of wild marbled crayfish (n=57) and P. fallax females (n=27) from comparable climatic regions (Hendrix et al., 2000; Jones et al., 2009). The maximum values of these samples were compared with the maximum values of our laboratory population. All quantitative data were statistically analysed using the Welsh two-sided t-test.
Data accessibility
The mitochondrial DNA sequences have been deposited in GenBank under the accession numbers KT074363, KT074364 and KT074365.
Acknowledgements
We thank Michael Pfeiffer (Gobio, March-Hugstetten, Germany) and Christoph Chucholl (Fisheries Research Station Baden-Württemberg, Langenargen, Germany) for providing marbled crayfish from Lake Moosweiher and for information on the biology of marbled crayfish in this lake, Frank Glaw (Bavarian State Collection of Zoology, Munich, Germany) and Miguel Vences (Braunschweig University of Technology, Braunschweig, Germany) for the Madagascar sample, the Swiss Federal Office for the Environment (BAFU, Bern, Switzerland) for the Procambarus clarkii samples, Frank Steuerwald (KABS, Waldsee, Germany) for information on the oldest known marbled crayfish, Chris Lukhaup (Hinterweidenthal, Germany) for Fig. 5i, Thomas Carell (Ludwig-Maximilians-University, Munich, Germany) for providing [D3]-dm5C internal standard for mass spectrometry, Günter Raddatz and Carine Legrand (DKFZ) for statistical help, the DKFZ Flow Cytometry and DKFZ Genomics and Proteomics core facilities for flow cytometry and DNA sequencing services, and Gerhard Scholtz (Humboldt University, Berlin, Germany), Bronwyn W. Williams (North Carolina Museum of Natural Sciences, Raleigh, USA), Zen Faulkes (University of Texas-Pan American, Edinburg, USA) and two anonymous referees for valuable comments.
Footnotes
Competing interests
The authors declare no competing or financial interests.
Author contributions
G.V. conceived the study, participated in the design of the study, sampled the tissues, performed the cross-breeding experiments and analysed the morphological and life history data; C.F. carried out the assembly and analysis of mitochondrial genome sequences and the determination of DNA contents by flow cytometry; K.H. maintained laboratory crayfish cultures and prepared DNA samples; A.S., J.P. and R.S. performed the analysis of the microsatellite markers; K.S and M.H. carried out the mass spectrometric measurement of DNA methylation; F.L. participated in the design of the study and coordinated the study. G.V. and F.L. wrote the manuscript. All authors revised the manuscript and gave final approval for publication.
Funding
This research received no specific grant from any funding agency in the public, commercial or not-for-profit sectors.
References
- Abbott R., Albach D., Ansell S., Arntzen J. W., Baird S. J. E., Bierne N., Boughman J., Brelsford A., Buerkle C. A., Buggs R. et al. (2013). Hybridization and speciation. J. Evol. Biol. 26, 229-246. 10.1111/j.1420-9101.2012.02599.x [DOI] [PubMed] [Google Scholar]
- Allen S. K., Jr (1983). Flow cytometry: assaying experimental polyploid fish and shellfish. Aquaculture 33, 317-328. 10.1016/0044-8486(83)90412-X [DOI] [Google Scholar]
- Alves M. J., Coelho M. M. and Collares-Pereira M. J. (2001). Evolution in action through hybridisation and polyploidy in an Iberian freshwater fish: a genetic review. Genetica 111, 375-385. 10.1023/A:1013783029921 [DOI] [PubMed] [Google Scholar]
- Alwes F. and Scholtz G. (2006). Stages and other aspects of the embryology of the parthenogenetic Marmorkrebs (Decapoda, Reptantia, Astacida). Dev. Genes Evol. 216, 169-184. 10.1007/s00427-005-0041-8 [DOI] [PubMed] [Google Scholar]
- Bai Z., Liu F., Li J. and Yue G. H. (2011). Identification of triploid individuals and clonal lines in Carassius auratus complex using microsatellites. Int. J. Biol. Sci. 7, 279-285. 10.7150/ijbs.7.279 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barraclough T. G., Birky C. W. Jr and Burt A. (2003). Diversification in sexual and asexual organisms. Evolution 57, 2166-2172. 10.1111/j.0014-3820.2003.tb00394.x [DOI] [PubMed] [Google Scholar]
- Belfiore N. M. and May B. (2000). Variable microsatellite loci in red swamp crayfish, Procambarus clarkii, and their characterization in other crayfish taxa. Mol. Ecol. 9, 2230-2234. 10.1046/j.1365-294X.2000.105339.x [DOI] [PubMed] [Google Scholar]
- Bell G. (1982). The Masterpiece of Nature: The Evolution and Genetics of Sexuality. Berkley: University of California Press. [Google Scholar]
- Birky C. W. Jr and Barraclough T. G. (2009). Asexual speciation. In Lost Sex: The Evolutionary Biology of Parthenogenesis (ed. Schön I., Martens K. and van Dijk P.), pp. 201-216. Dordrecht: Springer. [Google Scholar]
- Birky C. W. Jr, Adams J., Gemmel M. and Perry J. (2010). Using population genetic theory and DNA sequences for species detection and identification in asexual organisms. PLoS ONE 5, e10609 10.1371/journal.pone.0010609 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Butlin R. (2002). The costs and benefits of sex: new insights from old asexual lineages. Nat. Rev. Genet. 3, 311-317. 10.1038/nrg749 [DOI] [PubMed] [Google Scholar]
- Cesaroni D., Allegrucci G. and Sbordoni V. (1992). A narrow hybrid zone between two crayfish species from a Mexican cave. J. Evol. Biol. 5, 643-659. 10.1046/j.1420-9101.1992.5040643.x [DOI] [Google Scholar]
- Chen Z. J., Ha M. and Soltis D. (2007). Polyploidy: genome obesity and its consequences. New Phytol. 174, 717-720. 10.1111/j.1469-8137.2007.02084.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choleva L. and Janko K. (2013). Rise and persistence of animal polyploidy: evolutionary constraints and potential. Cytogenet. Genome Res. 140, 151-170. 10.1159/000353464 [DOI] [PubMed] [Google Scholar]
- Chucholl C. (2014). Predicting the risk of introduction and establishment of an exotic aquarium animal in Europe: insights from one decade of Marmorkrebs (Crustacea, Astacida, Cambaridae) releases. Manag. Biol. Invasions 5, 309-318. 10.3391/mbi.2014.5.4.01 [DOI] [Google Scholar]
- Chucholl C. (2015). Marbled crayfish gaining ground in Europe: the role of the pet trade as invasion pathway. In Freshwater Crayfish: A Global Overview (ed. Kawai T., Faulkes Z. and Scholtz G.), pp. 83-114. Boca Raton: CRC Press. [Google Scholar]
- Chucholl C. and Pfeiffer M. (2010). First evidence for an established Marmorkrebs (Decapoda, Astacida, Cambaridae) population in Southwestern Germany, in syntopic occurrence with Orconectes limosus (Rafinesque, 1817). Aquat. Invasions 5, 405-412. 10.3391/ai.2010.5.4.10 [DOI] [Google Scholar]
- Chucholl C., Morawetz K. and Groß H. (2012). The clones are coming – strong increase in Marmorkrebs [Procambarus fallax (Hagen, 1870) f. virginalis] records from Europe. Aquat. Invasions 7, 511-519. 10.3391/ai.2012.7.4.008 [DOI] [Google Scholar]
- Cohen H., Fait A. and Tel-Zur N. (2013). Morphological, cytological and metabolic consequences of autopolyploidization in Hylocereus (Cactaceae) species. BMC Plant Biol. 13, 173 10.1186/1471-2229-13-173 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cole C. J., Hardy L. M., Dessauer H. C., Taylor H. L. and Townsend C. R. (2010). Laboratory hybridization among North American whiptail lizards, including Aspidoscelis inornata arizonae × A. tigris marmorata (Squamata: Teiidae), ancestors of unisexual clones in nature. Am. Mus. Novitates 3698, 1-43. 10.1206/3698.2 [DOI] [Google Scholar]
- Comai L. (2005). The advantages and disadvantages of being polyploid. Nat. Rev. Genet. 6, 836-846. 10.1038/nrg1711 [DOI] [PubMed] [Google Scholar]
- Cordaux R., Pichon S., Ben Afia Hatira H., Doublet V., Grève P., Marcadé I., Braquart-Varnier C., Souty-Grosset C., Charfi-Cheikhrouha F. and Bouchon D. (2012). Widespread Wolbachia infection in terrestrial isopods and other crustaceans. ZooKeys 176, 123-131. 10.3897/zookeys.176.2284 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coyne J. A. and Orr H. A. (2004). Speciation. Sunderland: Sinauer Associates. [Google Scholar]
- Cuellar O. (1974). On the origin of parthenogenesis in vertebrates: the cytogenetic factors. Am. Nat. 108, 625-648. 10.1086/282940 [DOI] [Google Scholar]
- De Storme N. and Mason A. (2014). Plant speciation through chromosome instability and ploidy change: cellular mechanisms, molecular factors and evolutionary relevance. Curr. Plant Biol. 1, 10-33. 10.1016/j.cpb.2014.09.002 [DOI] [Google Scholar]
- Dufresne F. (2011). The history of the Daphnia pulex complex: asexuality, hybridization, and polyploidy. In Phylogeography and Population Genetics in Crustacea (ed. Held C., Koenemann S. and Schubart C. D.), Crustacean Issues 19, pp. 217-232. Boca Raton: CRC Press. [Google Scholar]
- Dufresne F. and Hebert P. D. N. (1995). Polyploidy and clonal diversity in an arctic cladoceran. Heredity 75, 45-53. 10.1038/hdy.1995.102 [DOI] [Google Scholar]
- Durand S., Bouché N., Perez Strand E., Loudet O. and Camilleri C. (2012). Rapid establishment of genetic incompatibility through natural epigenetic variation. Curr. Biol. 22, 326-331. 10.1016/j.cub.2011.12.054 [DOI] [PubMed] [Google Scholar]
- Eads B. D., Tsuchiya D., Andrews J., Lynch M. and Zolan M. E. (2012). The spread of a transposon insertion in Rec8 is associated with obligate asexuality in Daphnia. Proc. Natl. Acad. Sci. USA 109, 858-863. 10.1073/pnas.1119667109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Faria R. and Navarro A. (2010). Chromosomal speciation revisited: rearranging theory with pieces of evidence. Trends Ecol. Evol. 25, 660-669. 10.1016/j.tree.2010.07.008 [DOI] [PubMed] [Google Scholar]
- Faulkes Z. (2015a). Marble crayfish as a new model organism and a new threat to native crayfish conservation. In Freshwater Crayfish: A Global Overview (ed. Kawai T., Faulkes Z. and Scholtz G.), pp. 31-53. Boca Raton: CRC Press. [Google Scholar]
- Faulkes Z. (2015b). Marmorkrebs (Procambarus fallax f. virginalis) are the most popular crayfish in the North American pet trade. Knowl. Manag. Aquat. Ecosyst. 416, 20 10.1051/kmae/2015016 [DOI] [Google Scholar]
- Feria T. P. and Faulkes Z. (2011). Forecasting the distribution of Marmorkrebs, a parthenogenetic crayfish with high invasive potential, in Madagascar, Europe, and North America. Aquat. Invasions 6, 55-67. 10.3391/ai.2011.6.1.07 [DOI] [Google Scholar]
- Gao M., Huang Q., Chu Y., Ding C., Zhang B. and Su X. (2014). Analysis of the leaf methylomes of parents and their hybrids provides new insight into hybrid vigor in Populus deltoides. BMC Genet. 15 Suppl. 1, S8 10.1186/1471-2156-15-S1-S8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gavrilov-Zimin I. A., Stekolshchikov A. and Gautam D. C. (2015). General trends of chromosomal evolution in Aphidococca (Insecta, Homoptera, Aphidinea+ Coccinea). Comp. Cytogen. 9, 335-422. 10.3897/CompCytogen.v9i3.4930 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gherardi F. (2002). Behaviour. In Biology of Freshwater Crayfish (ed. Holdich D. M.), pp. 258-290. Oxford: Blackwell Science. [Google Scholar]
- Hahn C., Bachmann L. and Chevreux B. (2013). Reconstructing mitochondrial genomes directly from genomic next-generation sequencing reads – a baiting and iterative mapping approach. Nucleic Acids Res. 41, e129 10.1093/nar/gkt371 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hebert P. D. N. and Finston T. L. (2001). Macrogeographic patterns of breeding system diversity in the Daphnia pulex group from the United States and Mexico. Heredity 87, 153-161. 10.1046/j.1365-2540.2001.00885.x [DOI] [PubMed] [Google Scholar]
- Heethoff M., Domes K., Laumann M., Maraun M., Norton R. A. and Scheu S. (2007). High genetic divergences indicate ancient separation of parthenogenetic lineages of the oribatid mite Platynothrus peltifer (Acari, Oribatida). J. Evol. Biol. 20, 392-402. 10.1111/j.1420-9101.2006.01183.x [DOI] [PubMed] [Google Scholar]
- Hendrix A. N. and Loftus W. F. (2000). Distribution and relative abundance of the crayfishes Procambarus alleni (Faxon) and Procambarus fallax (Hagen) in Southern Florida. Wetlands 20, 194-199. 10.1672/0277-5212(2000)020[0194:DARAOT]2.0.CO;2 [DOI] [Google Scholar]
- Hendrix A. N., Armstrong D. and Grue C. (2000). Everglades Crayfish Final Report South Florida Ecosystem Restoration Program, RES97–9. Washington: U.S. Department of the Interior, National Park Service. [Google Scholar]
- Henry L., Schwander T. and Crespi B. J. (2012). Deleterious mutation accumulation in asexual Timema stick insects. Mol. Biol. Evol. 29, 401-408. 10.1093/molbev/msr237 [DOI] [PubMed] [Google Scholar]
- Hobbs H. H., Jr (1942). The crayfishes of Florida. Univ. Florida Publ. Biol. Sci. Ser. 3(2), 1-179. [Google Scholar]
- Hobbs H. H., Jr (1972). Crayfishes (Astacidae) of North and Middle America. Biota of Freshwater Ecosystems. Identification Manual 9. Washington, DC: Environmental Protection Agency. [Google Scholar]
- Hobbs H. H., Jr (1981). The crayfishes of Georgia. Smithson. Contrib. Zool. 318, 1-549. 10.5479/si.00810282.318 [DOI] [Google Scholar]
- Hobbs H. H., Jr (1989). An illustrated checklist of the American crayfishes (Decapoda: Astacidae, Cambaridae, and Parastacidae). Smithson. Contrib. Zool. 480, 1-236. 10.5479/si.00810282.480 [DOI] [Google Scholar]
- Jaenisch R. and Bird A. (2003). Epigenetic regulation of gene expression: how the genome integrates intrinsic and environmental signals. Nat. Genet. 33 Suppl., 245-254. 10.1038/ng1089 [DOI] [PubMed] [Google Scholar]
- Jones J. P. G., Rasamy J. R., Harvey A., Toon A., Oidtmann B., Randrianarison M. H., Raminosoa N. and Ravoahangimalala O. R. (2009). The perfect invader: a parthenogenic crayfish poses a new threat to Madagascar's freshwater biodiversity. Biol. Invasions 11, 1475-1482. 10.1007/s10530-008-9334-y [DOI] [Google Scholar]
- Kawai T., Scholtz G., Morioka S., Ramanamandimby F., Lukhaup C. and Hanamura Y. (2009). Parthenogenetic alien crayfish (Decapoda: Cambaridae) spreading in Madagascar. J. Crust. Biol. 29, 562-567. 10.1651/08-3125.1 [DOI] [Google Scholar]
- Keller N. S., Pfeiffer M., Roessink I., Schulz R. and Schrimpf A. (2014). First evidence of crayfish plague agent in populations of the marbled crayfish (Procambarus fallax forma virginalis). Knowl. Manag. Aquat. Ecosyst. 414, 15 10.1051/kmae/2014032 [DOI] [Google Scholar]
- Kellner S., Ochel A., Thüring K., Spenkuch F., Neumann J., Sharma S., Entian K.-D., Schneider D. and Helm M. (2014). Absolute and relative quantification of RNA modifications via biosynthetic isotopomers. Nucleic Acids Res. 42, e142 10.1093/nar/gku733 [DOI] [PMC free article] [PubMed] [Google Scholar]
- King M. (1993). Species Evolution: The Role of Chromosome Change. Cambridge: Cambridge University Press. [Google Scholar]
- Krois N. R., Cherukuri A., Puttagunta N. and Neiman M. (2013). Higher rate of tissue regeneration in polyploid asexual versus diploid sexual freshwater snails. Biol. Lett. 9, 20130422 10.1098/rsbl.2013.0422 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lafon-Placette C. and Köhler C. (2015). Epigenetic mechanisms of postzygotic reproductive isolation in plants. Curr. Opin. Plant Biol. 23, 39-44. 10.1016/j.pbi.2014.10.006 [DOI] [PubMed] [Google Scholar]
- Langmead B. and Salzberg S. L. (2012). Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357-359. 10.1038/nmeth.1923 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lavania U. C., Srivastava S., Lavania S., Basu S., Misra N. K. and Mukai Y. (2012). Autopolyploidy differentially influences body size in plants, but facilitates enhanced accumulation of secondary metabolites, causing increased cytosine methylation. Plant J. 71, 539-549. 10.1111/j.1365-313X.2012.05006.x [DOI] [PubMed] [Google Scholar]
- Leggatt R. A. and Iwama G. K. (2003). Occurrence of polyploidy in the fishes. Rev. Fish Biol. Fisheries 13, 237-246. 10.1023/B:RFBF.0000033049.00668.fe [DOI] [Google Scholar]
- Levin D. A. (2002). The Role of Chromosomal Change in Plant Evolution. New York: Oxford University Press. [Google Scholar]
- Li H., Handsaker B., Wysoker A., Fennell T., Ruan J., Homer N., Marth G., Abecasis G., Durbin R.. and 1000 Genome Project DataProcessing Subgroup. (2009). The Sequence Alignment/Map format and SAMtools. Bioinformatics 25, 2078-2079. 10.1093/bioinformatics/btp352 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li A., Hu B.-Q., Xue Z.-Y., Chen L., Wang W.-X., Song W.-Q., Chen C.-B. and Wang C.-G. (2011). DNA methylation in genomes of several annual herbaceous and woody perennial plants of varying ploidy as detected by MSAP. Plant. Mol. Biol. Rep. 29, 784-793. 10.1007/s11105-010-0280-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Little T. J. and Hebert P. D. N. (1997). Clonal diversity in high arctic ostracodes. J. Evol. Biol. 10, 233-252. 10.1007/s000360050020 [DOI] [Google Scholar]
- Lyko F. and Maleszka R. (2011). Insects as innovative models for functional studies of DNA methylation. Trends Genet. 27, 127-131. 10.1016/j.tig.2011.01.003 [DOI] [PubMed] [Google Scholar]
- Mallet J. (2007). Hybrid speciation. Nature 446, 279-283. 10.1038/nature05706 [DOI] [PubMed] [Google Scholar]
- Mark Welch D. B. and Meselson M. (2000). Evidence for the evolution of bdelloid rotifers without sexual reproduction or genetic exchange. Science 288, 1211-1215. 10.1126/science.288.5469.1211 [DOI] [PubMed] [Google Scholar]
- Mark Welch D. B., Ricci C. and Meselson M. (2009). Bdelloid rotifers: progress in understanding the success of an evolutionary scandal. In Lost Sex: The Evolutionary Biology of Parthenogenesis (ed. Schön I., Martens K. and van Dijk P.), pp. 259-280. Dordrecht: Springer. [Google Scholar]
- Martin P. (2015). Parthenogenesis: mechanisms, evolution, and its relevance to the role of marbled crayfish as model organism and potential invader. In Freshwater Crayfish: A Global Overview (ed. Kawai T., Faulkes Z. and Scholtz G.), pp. 63-82. Boca Raton: CRC Press. [Google Scholar]
- Martin P., Kohlmann K. and Scholtz G. (2007). The parthenogenetic Marmorkrebs (marbled crayfish) produces genetically uniform offspring. Naturwissenschaften 94, 843-846. 10.1007/s00114-007-0260-0 [DOI] [PubMed] [Google Scholar]
- Martin P., Dorn N. J., Kawai T., van der Heiden C. and Scholtz G. (2010). The enigmatic Marmorkrebs (marbled crayfish) is the parthenogenetic form of Procambarus fallax (Hagen, 1870). Contrib. Zool. 79, 107-118. [Google Scholar]
- Martin P., Thonagel S. and Scholtz G. (2015). The parthenogenetic Marmorkrebs (Malacostraca: Decapoda: Cambaridae) is a triploid organism. J. Zool. Syst. Evol. Res. (in press). [Google Scholar]
- Mayr E. (1963). Animal Species and Evolution. Cambridge: Harvard University Press. [Google Scholar]
- Mayr E. (1996). What is a species, and what is not? Philos. Sci. 63, 262-277. 10.1086/289912 [DOI] [Google Scholar]
- Minelli A. (2015). Grand challenges in evolutionary developmental biology. Front. Ecol. Evol. 2, 85 10.3389/fevo.2014.00085 [DOI] [Google Scholar]
- Moczek A. P. (2009). The origin and diversification of complex traits through micro- and macroevolution of development: insights from horned beetles. Curr. Top. Dev. Biol. 86, 135-162. 10.1016/S0070-2153(09)01006-0 [DOI] [PubMed] [Google Scholar]
- Neiman M., Jokela J. and Lively C. M. (2005). Variation in asexual lineage age in Potamopyrgus antipodarum, a New Zealand snail. Evolution 59, 1945-1952. 10.1111/j.0014-3820.2005.tb01064.x [DOI] [PubMed] [Google Scholar]
- Neiman M., Hehman G., Miller J. T., Logsdon J. M. Jr and Taylor D. R. (2010). Accelerated mutation accumulation in asexual lineages of a freshwater snail. Mol. Biol. Evol. 27, 954-963. 10.1093/molbev/msp300 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neiman M., Paczesniak D., Soper D. M., Baldwin A. T. and Hehman G. (2011). Wide variation in ploidy level and genome size in a New Zealand freshwater snail with coexisting sexual and asexual lineages. Evolution 65, 3202-3216. 10.1111/j.1558-5646.2011.01360.x [DOI] [PubMed] [Google Scholar]
- Neiman M. and Schwander T. (2011). Using parthenogenetic lineages to identify advantages of sex. Evol. Biol. 38, 115-123. 10.1007/s11692-011-9113-z [DOI] [Google Scholar]
- Parisod C., Holderegger R. and Brochmann C. (2010). Evolutionary consequences of autopolyploidy. New Phytol. 186, 5-17. 10.1111/j.1469-8137.2009.03142.x [DOI] [PubMed] [Google Scholar]
- Perry W. L., Feder J. L., Dwyer G. and Lodge D. M. (2001a). Hybrid zone dynamics and species replacement between Orconectes crayfishes in a northern Wisconsin lake. Evolution 55, 1153-1166. 10.1111/j.0014-3820.2001.tb00635.x [DOI] [PubMed] [Google Scholar]
- Perry W. L., Feder J. L. and Lodge D. M. (2001b). Implications of hybridization between introduced and resident Orconectes crayfishes. Conserv. Biol. 15, 1656-1666. 10.1046/j.1523-1739.2001.00019.x [DOI] [Google Scholar]
- Piferrer F., Beaumont A., Falguière J.-C., Flajšhans M., Haffray P. and Colombo L. (2009). Polyploid fish and shellfish: production, biology and applications to aquaculture for performance improvement and genetic containment. Aquaculture 293, 125-156. 10.1016/j.aquaculture.2009.04.036 [DOI] [Google Scholar]
- Reynolds J. D. (2002). Growth and reproduction. In Biology of Freshwater Crayfish (ed. Holdich D. M.), pp. 152-191. Oxford: Blackwell Science. [Google Scholar]
- Rubinoff D. and Le Roux J. J. (2008). Evidence of repeated and independent saltational evolution in a peculiar genus of sphinx moths (Proserpinus: Sphingidae). PLoS ONE 3, e4035 10.1371/journal.pone.0004035 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sax D. F. and Brown J. H. (2000). The paradox of invasion. Global Ecol. Biogeogr. 9, 363-371. 10.1046/j.1365-2699.2000.00217.x [DOI] [Google Scholar]
- Scholtz G. (2015). Happy birthday! The first decade of Marmorkrebs research – results and perspectives. In Freshwater Crayfish: A Global Overview (ed. Kawai T., Faulkes Z. and Scholtz G.), pp. 3-12. Boca Raton: CRC Press. [Google Scholar]
- Scholtz G., Braband A., Tolley L., Reimann A., Mittmann B., Lukhaup C., Steuerwald F. and Vogt G. (2003). Ecology: Parthenogenesis in an outsider crayfish. Nature 421, 806 10.1038/421806a [DOI] [PubMed] [Google Scholar]
- Schön I., Rossetti G. and Martens K. (2009). Darwinulid ostracods: ancient asexual scandals or scandalous gossip? In Lost Sex: The Evolutionary Biology of Parthenogenesis (ed. Schön I., Martens K. and van Dijk P.), pp. 217-240. Dordrecht: Springer. [Google Scholar]
- Schön I., Pinto R. L., Halse S., Smith A. J., Martens K. and Birky C. W. Jr (2012). Cryptic species in putative ancient asexual darwinulids (Crustacea, Ostracoda) . PLoS ONE 7, e39844 10.1371/journal.pone.0039844 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwander T. and Crespi B. J. (2009). Multiple direct transitions from sexual reproduction to apomictic parthenogenesis in Timema stick insects. Evolution 63, 84-103. 10.1111/j.1558-5646.2008.00524.x [DOI] [PubMed] [Google Scholar]
- Schwander T., Henry L. and Crespi B. J. (2011). Molecular evidence for ancient asexuality in Timema stick insects. Curr. Biol. 21, 1129-1134. 10.1016/j.cub.2011.05.026 [DOI] [PubMed] [Google Scholar]
- Seitz R., Vilpoux K., Hopp U., Harzsch S. and Maier G. (2005). Ontogeny of the Marmorkrebs (marbled crayfish): a parthenogenetic crayfish with unknown origin and phylogenetic position. J. Exp. Zool. 303A, 393-405. 10.1002/jez.a.143 [DOI] [PubMed] [Google Scholar]
- Sellars M. J., Degnan B. M. and Preston N. P. (2006). Production of triploid Kuruma shrimp, Marsupenaeus (Penaeus) japonicus (Bate) nauplii through inhibition of polar body I, or polar body I and II extrusion using 6-dimethylaminopurine. Aquaculture 256, 337-345. 10.1016/j.aquaculture.2006.02.052 [DOI] [Google Scholar]
- Sellars M., Wood A., Murphy B., Coman G., Arnold S., McCulloch R. and Preston N. (2013). Reproductive performance and mature gonad morphology of triploid and diploid Black Tiger shrimp (Penaeus monodon) siblings. Aquacult. Res. 44, 1493-1501. 10.1111/j.1365-2109.2012.03156.x [DOI] [Google Scholar]
- Shen H., Braband A. and Scholtz G. (2013). Mitogenomic analysis of decapod crustacean phylogeny corroborates traditional views on their relationships. Mol. Phylogenet. Evol. 66, 776-789. 10.1016/j.ympev.2012.11.002 [DOI] [PubMed] [Google Scholar]
- Soltis P. S. (2013). Hybridization, speciation and novelty. J. Evol. Biol. 26, 291-293. 10.1111/jeb.12095 [DOI] [PubMed] [Google Scholar]
- Soltis P. S. and Soltis D. E. (2000). The role of genetic and genomic attributes in the success of polyploids. Proc. Natl. Acad. Sci. USA 97, 7051-7057. 10.1073/pnas.97.13.7051 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soltis D. E., Soltis P. S., Schemske D. W., Hancock J. F., Thompson J. N., Husband B. C. and Judd W. S. (2007). Autopolyploidy in angiosperms: have we grossly underestimated the number of species? Taxon 56, 13-30. [Google Scholar]
- Suomalainen E., Saura A. and Lokki J. (1987). Cytology and Evolution in Parthenogenesis. Boca Raton: CRC Press. [Google Scholar]
- Tang C. Q., Obertegger U., Fontaneto D. and Barraclough T. G. (2014). Sexual species are separated by larger genetic gaps than asexual species in rotifers. Evolution 68, 2901-2916. 10.1111/evo.12483 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Theißen G. (2009). Saltational evolution: hopeful monsters are here to stay. Theory Biosci. 128, 43-51. 10.1007/s12064-009-0058-z [DOI] [PubMed] [Google Scholar]
- Thielsch A., Völker E., Kraus R. H. S. and Schwenk K. (2012). Discrimination of hybrid classes using cross-species amplification of microsatellite loci: methodological challenges and solutions in Daphnia. Mol. Ecol. Res. 12, 697-705. 10.1111/j.1755-0998.2012.03142.x [DOI] [PubMed] [Google Scholar]
- Tucker A. E., Ackerman M. S., Eads B. D., Xu S. and Lynch M. (2013). Population-genomic insights into the evolutionary origin and fate of obligately asexual Daphnia pulex. Proc. Natl. Acad. Sci. USA 110, 15740-15745. 10.1073/pnas.1313388110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- van der Kooi C. J. and Schwander T. (2014). Evolution of asexuality via different mechanisms in grass thrips (Thysanoptera: Aptinothrips). Evolution 68, 1883-1893. 10.1111/evo.12402 [DOI] [PubMed] [Google Scholar]
- Verhoeven K. J. F., van Dijk P. J. and Biere A. (2010). Changes in genomic methylation patterns during the formation of triploid asexual dandelion lineages. Mol. Ecol. 19, 315-324. 10.1111/j.1365-294X.2009.04460.x [DOI] [PubMed] [Google Scholar]
- Vogt G. (2008a). The marbled crayfish: a new model organism for research on development, epigenetics and evolutionary biology. J. Zool. 276, 1-13. 10.1111/j.1469-7998.2008.00473.x [DOI] [Google Scholar]
- Vogt G. (2008b). Investigation of hatching and early post-embryonic life of freshwater crayfish by in vitro culture, behavioral analysis, and light and electron microscopy. J. Morphol. 269, 790-811. 10.1002/jmor.10622 [DOI] [PubMed] [Google Scholar]
- Vogt G. (2010). Suitability of the clonal marbled crayfish for biogerontological research: a review and perspective, with remarks on some further crustaceans. Biogerontology 11, 643-669. 10.1007/s10522-010-9291-6 [DOI] [PubMed] [Google Scholar]
- Vogt G. (2011). Marmorkrebs: natural crayfish clone as emerging model for various biological disciplines. J. Biosci. 36, 377-382. 10.1007/s12038-011-9070-9 [DOI] [PubMed] [Google Scholar]
- Vogt G. (2015a). Research on stem cells, aging, cancer resistance, and epigenetics in marbled crayfish and relatives: potential benefits for human biology and medicine. In Freshwater Crayfish: A Global Overview (ed. Kawai T., Faulkes Z. and Scholtz G.), pp. 115-157. Boca Raton: CRC Press. [Google Scholar]
- Vogt G. (2015b). Stochastic developmental variation, an epigenetic source of phenotypic diversity with far-reaching biological consequences. J. Biosci. 40, 159-204. 10.1007/s12038-015-9506-8 [DOI] [PubMed] [Google Scholar]
- Vogt G., Tolley L. and Scholtz G. (2004). Life stages and reproductive components of the Marmorkrebs (marbled crayfish), the first parthenogenetic decapod crustacean. J. Morphol. 261, 286-311. 10.1002/jmor.10250 [DOI] [PubMed] [Google Scholar]
- Vogt G., Huber M., Thiemann M., van den Boogaart G., Schmitz O. J. and Schubart C. D. (2008). Production of different phenotypes from the same genotype in the same environment by developmental variation. J. Exp. Biol. 211, 510-523. 10.1242/jeb.008755 [DOI] [PubMed] [Google Scholar]
- Walker D., Porter B. A. and Avise J. C. (2002). Genetic parentage assessment in the crayfish Orconectes placidus, a high-fecundity invertebrate with extended maternal brood care. Mol. Ecol. 11, 2115-2122. 10.1046/j.1365-294X.2002.01609.x [DOI] [PubMed] [Google Scholar]
- West-Eberhard M. J. (2003). Developmental Plasticity and Evolution. New York: Oxford University Press. [Google Scholar]
- Williams B. W., Davis C. S. and Coltman D. W. (2010). Isolation and characterization of nine polymorphic microsatellite loci in the northern crayfish (Orconectes virilis). Conserv. Genet. Resour. 2, 235-237. 10.1007/s12686-010-9247-9 [DOI] [Google Scholar]
- Xiang J., Li F., Zhang C., Zhang X., Yu K., Zhou L. and Wu C. (2006). Evaluation of induced triploid shrimp Penaeus (Fenneropenaeus) chinensis cultured under laboratory conditions. Aquaculture 259, 108-115. 10.1016/j.aquaculture.2006.05.033 [DOI] [Google Scholar]
- Xu S., Spitze K., Ackerman M. S., Ye Z., Bright L., Keith N., Jackson C. E., Shaw J. R. and Lynch M. (2015). Hybridization and the origin of contagious asexuality in Daphnia pulex. Mol. Biol. Evol., Epub ahead of print 10.1093/molbev/msv190 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zerbino D. R. and Birney E. (2008). Velvet: algorithms for de novo short read assembly using de Bruijn graphs. Genome Res. 18, 821-829. 10.1101/gr.074492.107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang L. and King C. E. (1992). Genetic variation in sympatric populations of diploid and polyploid brine shrimp (Artemia parthenogenetica). Genetica 85, 211-221. 10.1007/BF00132273 [DOI] [PubMed] [Google Scholar]
- Zuber S. T., Muller K., Laushman R. H. and Roles A. J. (2012). Hybridization between an invasive and a native species of the crayfish genus Orconectes in North-Central Ohio. J. Crust. Biol. 32, 962-971. 10.1163/1937240X-00002091 [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The mitochondrial DNA sequences have been deposited in GenBank under the accession numbers KT074363, KT074364 and KT074365.