

Abstract
Purpose: The exact pathogenesis of sporadic parkinson’s disease (PD) is still unclear. Numerous evidences suggest involvement of apoptosis in the death of dopaminergic neurons. In this study we investigated the effect of sub-chronic administration of buspirone, fluoxetine and 8-hydroxy-2-[di-n-propylamino]tetralin (8-OH-DPAT) in 6-hydroxydopamine (6-OHDA)-lesioned rats and assayed striatal concentrations of apoptotic (Bax, Caspase3) and anti-apoptotic (Bcl-2) proteins.
Methods: 6-OHDA (8μg/2μl/rat) was injected unilaterally into the central region of the substantia nigra pars copmacta (SNc) of male Wistar rats and then, after 21 days lesioned rats were treated with intraperitonel (i.p) 1 mg/kg injections of buspirone, fluoxetine and 8-OH-DPAT for 10 consecutive days. Striatum of rats was removed at tenth day of drugs administration and were analyzed by western blotting method to measure Bax, caspase3 and Bcl-2 expression.
Results: The results showed that the expression of Bax and caspase3 proteins was increased three weeks after 6-OHDA injection while they were decreased significantly in parkinsonian rats which were treated by buspirone, fluoxetine and 8-OH-DPAT. Bcl-2 was decreased and increased in parkinsonian rats and parkinsonian rats treated with buspirone, fluoxetine and 8-OH-DPAT, respectively.
Conclusion: Our study indicates that sub-chronic administration of serotonergic drugs such as buspirone, fluoxetine and 8-OH-DPAT restores striatal concentration of apoptotic and anti-apoptotic factors to the basal levels of normal non-lesioned rats. We suggest that these drugs can be used as a potential adjunctive therapy in PD through attenuating neuronal apoptotic process.
Keywords: Fluoxetine, Buspirone, 8-OH-DPAT, Apoptosis, Parkinson's Disease
Introduction
The pathogenesis of sporadic Parkinson disease (PD) is still enigmatic; many factors are speculated to operate in the mechanism of cell death of the nigrostriatal dopaminergic neurons in PD. Recent studies have been focused on the factors of the pathways of programmed cell death, i.e., apoptosis, that might be involved in the neurodegeneration in PD.1 By end-stage disease in PD there is 80–95% loss of neurons in the substantia nigra.2 Increased levels of proteins that signal for apoptosis have been demonstrated not only in neurons of postmortem PD but also in experimental models of PD strongly suggested involvement of apoptosis in the death of nigrostriatal neurons.3 The major anti-apoptotic family members, Bcl-2 and Bcl-xL, which are thought to exert their effect at the mitochondrial outer membrane contribute to maintenance of membrane integrity. In contrast, one of the major pro-apoptotic family members, Bax, exerts its effects by compromising the membrane integrity leading to leakage of apoptogenic factors such as cytochrome c into the cytosol, resulting in caspase-3 activation and demise of the cell.4 Altered expressional levels of Bcl-2, Bax and increased caspase 1 and 3 activity have been reported in dopaminergic neurons of PD patients5,6 and in 6-OHDA-lesioned rats7 but the effect of serotonergic drugs on these proteins in PD have not studied. The stimulation of 5HT1A receptors induces a variable level of neuroprotection in different animal models of CNS injury.8 In vitro evidence indicates that 5HT1A agonists are able to protect neurons from apoptosis.9 Thus in this study we attempted to investigate the effect of chronic administration of serotonergic drugs (8-OH-DPAT, buspirone and fluoxetine) on 6-OHDA-induced PD in rats and possible involvement of Bax, Bcl-2 and Caspase-3 in this context.
Materials and Methods
Chemicals
All chemicals were obtained from Sigma Chemical Co. except for antibodies used for western blotting technique which were purchased from Abcam Co. Drugs and 6-OHDA were dissolved in physiological saline (0.9% NaCl) and 0.9% saline containing 0.2% (w/v) ascorbic acid respectively.
Animals and Treatment Protocol
The experiments were carried out on male Wistar rats weighing 270-300 g. The animals were given food and water ad libitum and were housed in standard polypropylene cages, four per cage at an ambient temperature of 25±2 °C under a 12-h light/12-h dark cycle. Animals were habituated to the testing conditions including being transferred to the experimental environment, handled, weighed, and restrained on the test platform for 10 min; 2 days before the investigations were conducted. The present study was carried out in accordance with the ethical guidelines for the Care and Use of Laboratory Animals of Tabriz University of Medical Sciences, Tabriz, Iran (National Institutes of Health Publication No. 85-23, revised 1985).
The parkinsonian rats were randomly allocated to equal groups (six rats per group) and were treated with 8-OH-DPAT (1mg/kg, i.p), fluoxetine (1mg/kg, i.p) and buspirone (1 mg/kg, i.p) for ten days. According to our previous study10-12 1mg/kg dose of these drugs had more profound anti-cataleptic effect in 6-OHDA-induced hemiparkinsonian rats. Then their striatal apoptotic proteins (Bcl-2, Bax and Caspase 3) were assessed by western blotting technique.
6-OHDA-Induced SNc Lesion
Animals were anesthetized with an intraperitoneal (i.p) injection of ketamine (50 mg/kg) and xylazine (5 mg/kg) and were mounted in a stereotaxic frame in the flat skull position. A central incision made to expose the skull. 6-OHDA was injected thorough a guide cannula (23 gauge stainless steel) implanted in the SNc. The coordinates for this site were based on the rat brain atlas13 anteroposterior (AP): –5.0 mm from the bregma; mediolateral (ML): –2.1 mm from the midline and dorsoventral (DV): –7.7 from the skull. Desipramine (25 mg/kg, i.p) was injected 30 min before the intra-SNc injection of 6-OHDA to avoid the destruction of noradrenergic neurons. Then, 6-OHDA (8 μg/per rat in 2 μl saline with 0.2% ascorbic acid) was infused with an infusion pump at a constant flow rate of 0.2 μl/min into the left SNc.
Western Blot Analysis
Animals were sacrificed after ten days administration of drugs and their striatum was dissected and homogenized in lysis buffer (Cocktail Protease Inhibitor 10 µl/ml and Phenyl Methane Sulfonyl Fluoride 10 µl/ml). Lysates (30 µg protein) were resolved by SDS-PAGE for Bax, Caspase3 and Bcl-2 assay; then were transferred to polyvinylidene difluoride filters (PVDF) which were blocked with 5% milk and were incubated overnight at 4°C with the following primary antibodies: Bax, Bcl-2 and GAPDH, 1:1000 , Caspase-3, 1:500. Subsequently, PVDF membranes were incubated for 1 hr at room temperature with an HRP-conjugated secondary antibody (1:3000); incubated with the chemiluminescent substrate for 90 s; and exposed for 30 s to 5 min, to Hyperfilm. Finally the developed films were scanned, and density of immonoreactive bands was measured using “Image J” software and was normalized to the internal control bands (GAPDH was the loading control).
Statistical Analysis
Statistical analysis of each data set was performed by use of SPSS software (version 16.0). Data were expressed as the mean±SEM, and were analyzed by one-way ANOVA in behavioral and biochemical experiments. In the case of significant variation (p<0.05), the values were compared by Tukey test.
Results
The Effects of Buspirone, Fluoxetine and 8-OH-DPAT on Bax Expression
The Expression of striatal Bax protein in 6-OHDA-lesioned rats increased significantly (p<0.001) when compared to non-lesioned (control) rats. The amount of this protein in buspirone, fluoxetine and 8-OH-DPAT-treated parkinsonian rats was almost same as the control level. On the other hand, these drugs decreased expression of Bax protein in 6-OHDA-lesioned rats (Figure 1).
Figure 1.
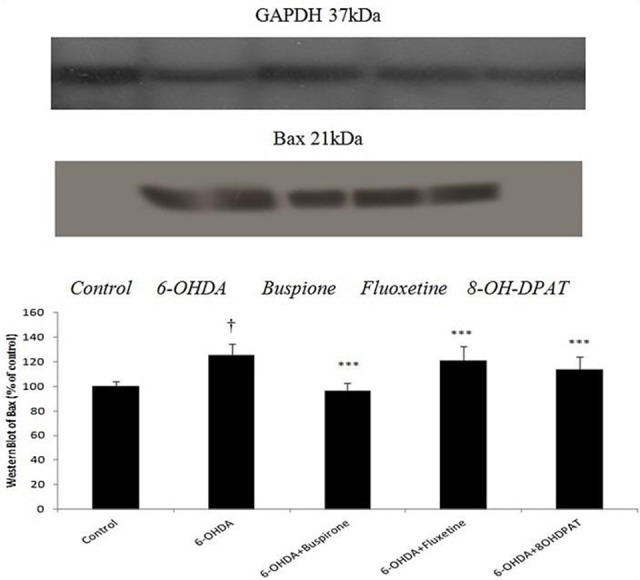
Effect of sub-chronically(10 days) administered buspirone,fluoxetine and 8-OH-DPAT (1 mg/kg, i.p) on SNcbax expression analyzed by Western Blot. Data represent the mean percentage of controls (±SEM) for 6 animals in each group. Data were analyzed by ANOVA followed by tukey’s-test. † p<0.001 when compared with normal group (control), ***p<0.001 when compared with 6-OHDA-lesioned group .GAPDH was loading control.
The Effects of Buspirone, Fluoxetine and 8-OH-DPAT on Caspase-3 Expression
The striatum of aforementioned groups was investigated for evaluation of pro-apoptotic Caspase-3 protein activity. Results showed that in 6-OHDA-lesioned rats expression of this protein was more than control group (p<0.001), while in 6-OHDA-lesioned rats which were treated with buspirone, fluoxetine and 8-OH-DPAT expression of Caspase-3 protein was reduced to control level (Figure 2).
Figure 2.
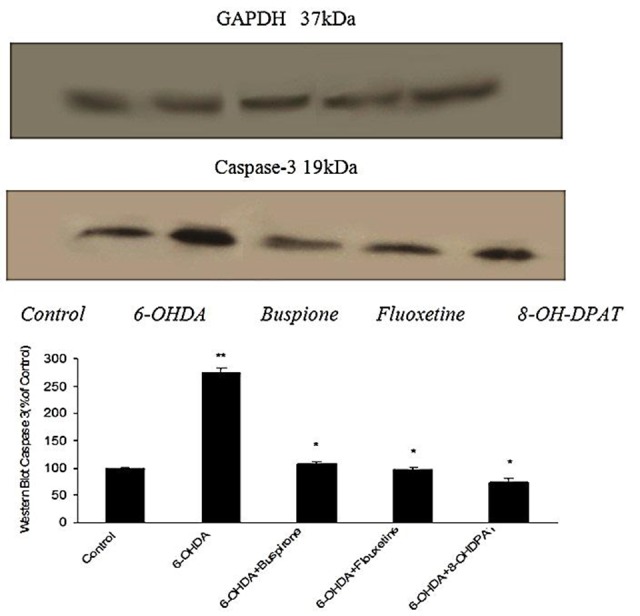
Effect of chronically (10 days) administered buspirone,fluoxetine and 8-OH-DPAT (1 mg/kg, i.p) on SNc Caspase-3 activity analyzed byWestern Blot. Data represent the mean percentage of controls (±SEM) for 6 animals in each group. Data were analyzed by ANOVA followed by tukey’s-test. **p<0.001 when compared with normal group (control), *p<0.001 when compared with 6-OHDA-lesioned group .GAPDH was loading control.
The Effects of Buspirone, Fluoxetine and 8-OH-DPAT on Bcl-2 Expression
As it has been shown in Figure 3, the expression of anti-apoptotic Bcl-2 protein in the striatum of 6-OHDA-lesioned rats was less than (p<0.001) control group. In 6-OHDA-lesioned rats which were treated with buspirone, fluoxetine and 8-OH-DPAT, expression of Bcl-2 protein was greater (p<0.001) than 6-OHDA-lesioned rats.
Figure 3.
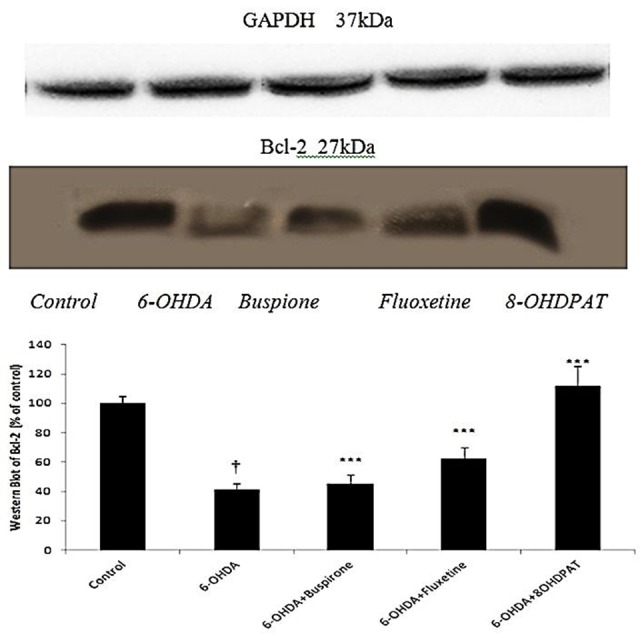
Effect of chronically (10 days) administered buspirone,fluoxetine and 8-OH-DPAT (1 mg/kg, i.p) on SNc Bcl-2 expression analyzed byWestern Blot. Data represent the mean percentage of controls (±SEM) for 6 animals in each group. Data were analyzed by ANOVA followed by tukey’s-test. †p<0.001 when compared with normal group (control); ***p <0.001 when compared with 6-OHDA-lesioned group .GAPDH was loading control.
Discussion
In our previous studies we have shown that 6-OHDA (8 μg/2 μl/rat) could induce significant catalepsy in bar test when compared with control (normal) and sham-operated animals. 10-12, 14-17 Furthermore, 8-OH-DPAT (1 mg/kg, i.p),10 fluoxetine (1 mg/kg, i.p)11 and buspirone (1 mg/kg, i.p)12 attenuated 6-OHDA-induced catalepsy in hemiparkinsonian rats. Thus, in this study we attempted to assess the effect of these drugs on expression of pro-apoptotic proteins, Bax and Caspase-3 as well as anti-apoptotic Bcl-2 protein in 6-OHDA-lesioned rats. Our results showed that expression of Bax protein was increased significantly in 6-OHDA-lesioned rats. This is in agreement with previous studies showing an over expression of Bax protein in parkinsonian patients18 and 6-OHDA-treated rats.19-20 In those groups of parkinsonian rats which were treated sub-chronically by buspirone, fluoxetine and 8-OH-DPAT, striatal expression of Bax protein was dropped markedly to the level of control group. The attenuating effect of buspirone and 8-OH-DPAT on expression of Bax Protein was greater than fluoxetine.
Caspase family has 14 members that many of these proteases have been implicated in neuronal apoptosis. The first evidence that caspase activity is required for neuronal apoptosis came from experiments using pharmacological inhibitors of caspases. Moreover caspases may play pivotal roles in a number of acute and chronic neurologic disorders including stroke, Amyotrophic lateral sclerosis (ALS), PD and Huntington’s disease.21 For example, mice transgenic for a dominant negative mutant of caspase-1 exhibit reduced sensitivity to ischemia as well as trophic factor deprivation. Accordingly, inhibition of caspase-3 expression plays a crucial neuroprotective role in a number of animal and human models of neurodegenerative disease such as Alzheimer disease (AD) and PD.22 Our data demonstrate that in 6-OHDA-lesioned rats there was a significant increase in striatal Caspase-3 level when compared to control group. This is in accordance with previous studies reporting an increase of in 6-OHDA, Methyl-4-phenyl-1,2,3,6-tetrahydropyridine ( MPTP) and lipopolysaccharide (LPS)-treated rats.3,23 Additionally we assessed Caspase-3 activity in striatum of buspirone, fluoxetine and 8-OH-DPAT-treated parkinsonian rats. In parkinsonian rats which were treated with these drugs Caspase-3 expression diminished to the level of non-lesioned (control) animals. The effect of 8-OH-DPAT on Caspase-3 expression was more profound than buspirone and fluoxetine. In a previous study it has been shown that 8-OH-DPAT can reduce morphine-induced apoptosis in hippocampus of rats.24 Since 8-OH-DPAT is a pure agonist of 5-HT1A receptors, therefore we may suggest possible involvement of these receptors in apoptosis process of PD.
Bcl-2 is specifically considered as an important anti-apoptotic protein and is thus classified as an oncogene. Many members of the Bcl-2 family have been implicated in the regulation of neuronal cell death. The exact mechanism by which the Bcl-2 proteins control apoptosis is still not entirely clear but studies have shown that it’s over expression in cell culture and in transgenic mice increases resistance of neurons to death during development and protects from apoptosis induced by excitotoxic, metabolic and oxidative insults relevant to AD, stroke and other disorder.22,25 In this study, Bcl-2 level was decreased in 6-OHDA-lesioned rats.26 Following treatment with buspirone, fluoxetine and 8-OH-DPAT, striatal level of this protein was elevated significantly in comparison to 6-OHDA-lesioned rats. The effect of 8-OH-DPAT and buspirone (as a partial agonist of 5-HT1A receptors) was greater than fluoxetine.
Previous studies suggested that 5-HT1A receptors have neuroprotective effect.27-30 We have shown that these drugs via stimulation of 5-HT1A receptors, can improve 6-OHDA-induced catalepsy in rats.10-12 The stimulation of 5HT1A receptors induces a variable level of neuroprotection in different animal models of CNS injury such as ischemia, N-methyl-d-aspartate (NMDA) excitotoxicity, acute subdural hematoma, and traumatic brain injury. Furthermore, in vitro evidence indicates that 5HT1A agonists are able to protect neurons from apoptosis.24 There are different hypotheses on the mechanisms involved in 5HT1A-mediated neuroprotection, including neuronal membrane hyperpolarization that reduces excitability, reduced glutamate release, and blockade of voltage-sensitive Na+ channels.24 Other neuroprotective mechanisms have also been proposed for 5HT1A agonists such as stimulation of the BCL-2 expression through the MAPK/ERK signaling pathway and suppression of the pro-apoptotic protein caspase-3 in a MAPK- and protein kinase C-α dependent manner.31,32 Thus, we may suggest that the observed anti-apoptotic effects of serotonergic drugs i.e. buspirone, fluoxetine and 8-OH-DPAT is exerted through 5-HT1A receptors.
According to the obtained results buspirone, fluoxetine and 8-OH-DPAT diminished striatal expression of Bax and Caspase-3 proteines in parkinsonian rats, whereas they increased Bcl-2 level. On the other hand, these drugs could attenuate apoptotic process in 6-OHDA-induced experimental model of PD. This correlates well with the results of our previous behavioral studies10-12 reporting an anti-cataleptic effect for buspirone, fluoxetine and 8-OH-DPAT in 6-OHDA-lesioned rats.
Conclusion
In conclusion, we suggest that serotonergic agents may have beneficial effect in preventing the progression of PD through attenuating apoptotic events. However, further clinical investigations should be carried out to prove this.
Ethical Issues
Not applicable.
Conflict of Interest
The authors report no conflicts of interest.
References
- 1.Nagatsu T. Parkinson's disease: Changes in apoptosis-related factors suggesting possible gene therapy. J Neural Transm. 2002;109(5-6):731–45. doi: 10.1007/s007020200061. [DOI] [PubMed] [Google Scholar]
- 2.Double KL, Reyes S, Werry EL, Halliday GM. Selective cell death in neurodegeneration: Why are some neurons spared in vulnerable regions? Prog Neurobiol. 2010;92(3):316–29. doi: 10.1016/j.pneurobio.2010.06.001. [DOI] [PubMed] [Google Scholar]
- 3.Singh S, Dikshit M. Apoptotic neuronal death in parkinson's disease: Involvement of nitric oxide. Brain Res Rev. 2007;54(2):233–50. doi: 10.1016/j.brainresrev.2007.02.001. [DOI] [PubMed] [Google Scholar]
- 4.Zaldana M, Bonastre M, Aguilar E, Marin C. Differential nigral expression of bcl-2 protein family in chronically haloperidol and clozapine-treated rats: Role in neurotoxicity and stereotyped behavior. Exp Neurol. 2007;203(2):302–8. doi: 10.1016/j.expneurol.2006.08.009. [DOI] [PubMed] [Google Scholar]
- 5.Hartmann A, Troadec JD, Hunot S, Kikly K, Faucheux BA, Mouatt-Prigent A. et al. Caspase-8 is an effector in apoptotic death of dopaminergic neurons in parkinson's disease, but pathway inhibition results in neuronal necrosis. J Neurosci. 2001;21(7):2247–55. doi: 10.1523/JNEUROSCI.21-07-02247.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hartmann A, Mouatt-Prigent A, Vila M, Abbas N, Perier C, Faucheux BA. et al. Increased expression and redistribution of the antiapoptotic molecule bcl-xl in parkinson's disease. Neurobiol Dis. 2002;10(1):28–32. doi: 10.1006/nbdi.2002.0494. [DOI] [PubMed] [Google Scholar]
- 7.Kramer BC, Mytilineou C. Alterations in the cellular distribution of bcl-2, bcl-x and bax in the adult rat substantia nigra following striatal 6-hydroxydopamine lesions. J Neurocytol. 2004;33(2):213–23. doi: 10.1023/b:neur.0000030696.62829.ec. [DOI] [PubMed] [Google Scholar]
- 8.Oosterink BJ, Harkany T, Luiten PG. Post-lesion administration of 5-ht1a receptor agonist 8-oh-dpat protects cholinergic nucleus basalis neurons against nmda excitotoxicity. Neuroreport. 2003;14(1):57–60. doi: 10.1097/01.wnr.0000051543.96524.3c. [DOI] [PubMed] [Google Scholar]
- 9.Ahlemeyer B, Glaser A, Schaper C, Semkova I, Krieglstein J. The 5-ht1a receptor agonist bay x 3702 inhibits apoptosis induced by serum deprivation in cultured neurons. Eur J Pharmacol. 1999;370(2):211–6. doi: 10.1016/s0014-2999(99)00136-3. [DOI] [PubMed] [Google Scholar]
- 10.Sharifi H, Nayebi AM, Farajnia S. Chronic administration of 8-OH-DPAT attenuates catlepsy in 6-hydroxydopamine-lesioned rats through modulation of inflammatory cytokines. Iran J Basic Med Sci. 2013;16(12):1270–75. [PMC free article] [PubMed] [Google Scholar]
- 11.Sharifi H, Mohajjel Nayebia A, Farajnia S. Dose-dependent effect of flouxetine on 6-ohda-induced catalepsy in male rats: A possible involvement of 5-ht1a receptors. Advanced pharmaceutical bulletin. 2013;3(1):203–6. doi: 10.5681/apb.2013.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Sharifi H, Mohajjel Nayebia A, Farajnia S. The effect of chronic administration of buspirone on 6-hydroxydopamine-induced catalepsy in rats. Adv Pharm Bull. 2012;2(1):127–31. doi: 10.5681/apb.2012.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Paxinos G, Watson C. The rat brain in stereotaxic coordinates. 6th ed. Sydney: Academic Press; 1982. [Google Scholar]
- 14.Mahmoudi J, Mohajjel Nayebi A, Reyhani-Rad S, Samini M. Fluoxetine improves the effect of levodopa on 6-hydroxy dopamine-induced motor impairments in rats. Adv Pharm Bull. 2012;2(2):149–55. doi: 10.5681/apb.2012.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Javad Mahmoudi, Alireza Mohajjel Nayebi, Morteza Samini, Siamak Reyhani-Rad, Vahab Babapour. Buspirone improves the anti-cataleptic effect of levodopa in 6-hydroxydopamine-lesioned rats. Pharmacolo Rep. 2011;63(4):908–914. doi: 10.1016/s1734-1140(11)70606-5. [DOI] [PubMed] [Google Scholar]
- 16.Mahmoudi J, Mohajjel Nayebi A, Samini M, Reyhani-Rad S, Babapour V. 5-ht(1a) receptor activation improves anti-cataleptic effects of levodopa in 6-hydroxydopamine-lesioned rats. Daru. 2011;19(5):338–43. [PMC free article] [PubMed] [Google Scholar]
- 17.Nayebi AM. Hypothesis: A promising effect of 5-HT1A receptor agonists in alleviating motor symptoms of Parkinsons disease. Afr J Pharm Pharmaco. 2010;4(6):289–90. [Google Scholar]
- 18.Levy OA, Malagelada C, Greene LA. Cell death pathways in parkinson's disease: Proximal triggers, distal effectors, and final steps. Apoptosis. 2009;14(4):478–500. doi: 10.1007/s10495-008-0309-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Ayllon V, Cayla X, Garcia A, Roncal F, Fernandez R, Albar JP. et al. Bcl-2 targets protein phosphatase 1 alpha to bad. J Immunol. 2001;166(12):7345–52. doi: 10.4049/jimmunol.166.12.7345. [DOI] [PubMed] [Google Scholar]
- 20.Djordjevic A, Djordjevic J, Elakovic I, Adzic M, Matic G, Radojcic MB. Fluoxetine affects hippocampal plasticity, apoptosis and depressive-like behavior of chronically isolated rats. Prog Neuropsychopharmacol Biol Psychiatry. 2012;36(1):92–100. doi: 10.1016/j.pnpbp.2011.10.006. [DOI] [PubMed] [Google Scholar]
- 21.Yuan J, Yankner BA. Apoptosis in the nervous system. Nature. 2000;407(6805):802–9. doi: 10.1038/35037739. [DOI] [PubMed] [Google Scholar]
- 22.Becker EB, Bonni A. Cell cycle regulation of neuronal apoptosis in development and disease. Prog Neurobiol. 2004;72(1):1–25. doi: 10.1016/j.pneurobio.2003.12.005. [DOI] [PubMed] [Google Scholar]
- 23.Han BS, Hong HS, Choi WS, Markelonis GJ, Oh TH, Oh YJ. Caspase-dependent and -independent cell death pathways in primary cultures of mesencephalic dopaminergic neurons after neurotoxin treatment. J Neurosci. 2003;23(12):5069–78. doi: 10.1523/JNEUROSCI.23-12-05069.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Charkhpour M, Nayebi AR, Doustar Y, Hassanzadeh K. 8-oh-dpat prevents morphine-induced apoptosis in rat dorsal raphe nucleus: A possible mechanism for attenuating morphine tolerance. Anesth Analg. 2010;111(5):1316–21. doi: 10.1213/ANE.0b013e3181f1bb9a. [DOI] [PubMed] [Google Scholar]
- 25.Mattson MP. Apoptosis in neurodegenerative disorders. Nat Rev Mol Cell Biol. 2000;1(2):120–9. doi: 10.1038/35040009. [DOI] [PubMed] [Google Scholar]
- 26.Guo DJ, Li F, Yu PH, Chan SW. Neuroprotective effects of luteolin against apoptosis induced by 6-hydroxydopamine on rat pheochromocytoma pc12 cells. Pharm Biol. 2013;51(2):190–6. doi: 10.3109/13880209.2012.716852. [DOI] [PubMed] [Google Scholar]
- 27.Berends AC, Luiten PG, Nyakas C. A review of the neuroprotective properties of the 5-ht1a receptor agonist repinotan hcl (bayx3702) in ischemic stroke. CNS drug reviews. 2005;11(4):379–402. doi: 10.1111/j.1527-3458.2005.tb00055.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ohman J, Braakman R, Legout V. Repinotan (bay x 3702): A 5ht1a agonist in traumatically brain injured patients. J Neurotrauma. 2001;18(12):1313–21. doi: 10.1089/08977150152725614. [DOI] [PubMed] [Google Scholar]
- 29.Meller E. 5-ht1a receptor-mediated apoptosis: Death by jnk? Biochim Biophys Acta. 2007;1773(6):691–3. doi: 10.1016/j.bbamcr.2007.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Cowen DS, Johnson-Farley NN, Travkina T. 5-ht receptors couple to activation of akt, but not extracellular-regulated kinase (erk), in cultured hippocampal neurons. J Neurochem. 2005;93(4):910–7. doi: 10.1111/j.1471-4159.2005.03107.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kukley M, Schaper C, Becker A, Rose K, Krieglstein J. Effect of 5- hydroxytryptamine 1A receptor agonist BAY X 3702 on BCL-2 and BAX proteins level in the ipsilateral cerebral cortex of rats after transient focal ischemia. Neuroscience. 2001;107(3):405–13. doi: 10.1016/s0306-4522(01)00369-4. [DOI] [PubMed] [Google Scholar]
- 32.Adayev T, Ray I, Sondhi R, Sobocki T, Banerjee P. The g protein-coupled 5-ht1a receptor causes suppression of caspase-3 through mapk and protein kinase calpha. Biochim Biophys Acta. 2003;1640(1):85–96. doi: 10.1016/s0167-4889(03)00023-5. [DOI] [PubMed] [Google Scholar]