

Abstract
Birds are crucial to maintaining the balance of many ecosystems by providing various ecological services. The diversity of birds and their feeding guilds in different land-use types were investigated in south-central Mindanao to elucidate the effect of disturbance and habitat modification on bird communities. Point count method was employed to identify birds in three habitat types: i) agroforests; ii) ricefields; iii) roads and heavily disturbed areas. A total of 1114 bird sightings were recorded that included 33 species of 24 families; of these, 3 were Philippine endemics, and 5 were migrant species. Among all of the habitat types, the highest species diversity was found in agroforests (1/D = 16.148), and the lowest was recorded from roads and heavily disturbed habitats. The species composition of agroforests was more similar to ricefields than to areas with high levels of disturbance, such as roads. The characteristic of the vegetation and the availability of food resources may be vital to the diversity of birds in every habitat as evidenced by the high species richness of frugivores and insectivores in agroforests and ricefields, respectively, where food source is largely available. The observation of Streptopelia tranquebarica was a new record for Mindanao, and it was particularly sighted in ricefields. Therefore, this study indicates that land-use change and modification may alter bird diversity structure, and the maintenance of the vegetation in land-use types as food and resource, and as habitat is essential to the conservation of the native and ecologically-important bird species in south-central Mindanao.
Keywords: Agroforest, Avifauna, Conservation, Feeding Guild, Frugivores
Abstract
Burung-burung penting untuk mengekalkan keseimbangan ekosistem dengan membekalkan pelbagai servis ekologi. Kepelbagaian burung-burung and perseikatan pemakanan dalam pelbagai jenis guna-tanah telah dikaji di selatan-tengah Mindanao untuk menjelaskan kesan-kesan gangguan dan pengubahsuaian habitat komuniti burung. Kaedah bilang titik telah digunakan untuk mengenal pasti burung-burung dalam tiga jenis habitat: i) hutan agro; ii) sawah padi; iii) jalan raya dan kawasan yang mengalami gangguan yang teruk. Sebanyak 1114 burung telah dilihat dan direkodkan, termasuk 33 spesies daripada 24 famili; daripada ini, 3 merupakan spesies endemik Filipina, dan 5 merupakan spesies pehijrah. Daripada semua jenis habitat, diversiti spesies yang tertinggi telah dijumpai di hutan agro (1/D = 16.148), dan yang terendah direkodkan daripada jalan raya dan kawasan yang mengalami gangguan yang teruk. Komposisi spesies hutan agro lebih mirip dengan sawah padi berbanding dengan kawasan yang mengalami gangguan yang teruk, seperti jalan raya. Sifat-sifat tumbuhan dan ketersediaan sumber makanan mungkin penting untuk kepelbagaian spesies di setiap habitat seperti yang terbukti oleh kekayaan spesies frugivor dan insektivor yang tinggi di hutan agro dan sawah padi, masing-masing, yang mempunyai sumber makanan yang banyak. Pencerapan terhadap Streptopelia tranquebarica merupakan rekod baru untuk Mindanao, dan secara spesifiknya ia ditemui di sawah padi. Maka, kajian ini menunjukkan bahawa perubahan guna-tanah mungkin boleh mengubahsuai struktur kepelbagaian burung, dan pengekalan tumbuhan dalam jenis guna-tanah sebagai makanan dan sumber, dan sebagai habitat penting untuk pemuliharaan spesies-spesies burung natif dan penting secara ekologi di selatan-tengah Mindanao.
Keywords: Hutan Agro, Avifauna, Pemuliharaan, Perseikatan Pemakanan, Frugivor
INTRODUCTION
The Philippines is one of the most biodiverse countries in the world and its tropical rainforests are among the richest in Southeast Asia with exceptional species diversity and a high number of endemic species (Heaney 1993). However, the conversion of tropical rainforest into agricultural lands has resulted in the local and regional extinction of many species (Myers 1988). Lowland forest in the Philippines has decreased by about 90%, and approximately 94% of the total land area of the Philippines was once covered by forest. By the end of World War II, forest cover had been reduced to 40%, and current estimates range from 25% to less than 20% (Utzurrum 1991; Carandang 2005; Forest Management Bureau 2013).
The country’s vast archipelago harbours more than 700 known species of birds, but the number continues to increase as a result of numerous sampling and survey expeditions. The natural habitat of most of the country’s birds is tropical forest; 67% of Philippine birds spend their entire life or part of their life in forest. However, much of the tropical forest in the Philippines has been cleared for logging, mining, and agriculture as well as for industrial and development purposes (Birdlife International 2012).
Birds are present in various of habitat types and are among the important groups that play a vital role in both the structure and function of ecosystems by providing numerous ecological benefits, such as seed dispersal, the facilitation of forest restoration (et al. 2007), the pollination of many tropical plant species, and pest control services through the consumption of insects (Philpott et al. 2009; Sekercioglu 2012) and small rodents, which can devastate hectares of agricultural products. Thus, birds are an ideal study group for the valuation of ecosystem services (Wenny et al. 2011).
Although less than 1% of the world’s bird species prefer agricultural areas as their primary habitat, nearly a third of all bird species use such habitats occasionally (Sekercioglu et al. 2012). Some agroecosystems can also harbour a substantial portion of the biodiversity found under the original land cover (Vandermeer & Perfecto 1997) and can buffer and complement protected areas (Curran et al. 2004). For example, areas planted with rice (Oryza spp.) are also used by a wide variety of bird species during the non-growing season and play an important conservation role in many parts of the world (Elphick et al. 2010).
The environmental impacts on birds are typically assessed by recording changes in the population density, abundance or distribution of species in different habitat types (Temple & Wiens 1989), and the primary objective of this study is the determination of the avifaunal species assemblage and its status in different land-use types in south-central Mindanao, Philippines. This paper aims to i) identify bird species to the lowest possible level of taxonomic classification in three different land-use types (agroforests, ricefields and roads); ii) determine and compare the species diversity, conservation status and endemism of the different habitats; and iii) relate species diversity to the extent of disturbance in different habitats.
MATERIALS AND METHODS
The Study Site
This study of avifaunal diversity and the feeding guilds in different habitats was conducted on a 1024-hectare campus of the University of Southern Mindanao in south-central Mindanao (Fig. 1). The study site was chosen for its variety of habitat types, which include agroforests, orchards, ricefields and heavily disturbed habitats (roads and buildings). Additionally, the elevation is uniform among the habitat types, so the influence of elevation on bird diversity was excluded from the analysis. The elevation of the study site ranges from 33 to 60 masl with an annual mean temperature of 27.48°C and mean rainfall of 1980 to 2500 mm/year (World Climatic Database 2014). Each of the habitat types are unique in terms of floristic composition and level of disturbance, so each of the land use types were described accordingly.
Figure 1:

Map of the study site in lowland south-central Mindanao, Philippines. Note: Map generated using QGIS v. 2.2.
Agroforest
The agroforest habitat (7° 6′ 36.558″ N, 124° 50′ 51.331″ E) is characterised as former secondary forest mixed with vegetation dominated by fruit-bearing trees including lanzones (Lansium domesticum), pomelo (Citrus maxima), durian (Durio sp.), rambutan (Nephelium lappaceum), chico (Achras zapota) and papaya (Carica papaya). Corn (Zea maize) and banana (Musa paradisiaca) are present at the edges of orchards, such as for chico and lanzones, and patches of cogon (Imperata cylindrica) cover some of the area, especially the edges of cornfields, while mahogany (Swietenia mahogani) covers some sections of the manmade paths.
Ricefields
The ricefield habitat (7° 7′ 10.324″ N, 124° 49′ 22.979″ E) is surrounded by oil palm (Elaeis guineensis), coconut (Coco nucifera), acacia (Samania saman) and other types of vegetation, such as shrubs, ferns and grasses. Other areas of the ricefields are dominated by corn (Z. maize) and mango trees (Mangifera indica). Three cropping seasons occur each year with growing seasons of 2.5 to 3.0 months, and the sampling period began with the start of the harvest season and ended during early field preparation, in which field flooding and soil ploughing occur. During the growing season, pesticides and herbicides were used to prevent insects and weeds from inhibiting crop growth.
Roads and Heavily Disturbed Areas
Heavily disturbed habitats (7° 6′ 59.784″ N and 124° 49′ 59.073″ E) are primarily represented by roads and areas with infrastructure, such as buildings, parks, canteens, hospitals, dorms and cottages on the campus, where human disturbance is very high and visible. Vehicles and motorcycles are also found in the study site. Trees, such as mahogany and oil palm (E. guineensis), have been planted along road edges, which are dominated by tall grasses such cogon (I. cylindrica) and other grasses and sedges.
Sampling Methods
Birds were observed through point count method as described by Sutherland et al. (2004). At least 15 points were established along existing manmade or natural trails in each habitat with approximately 200 m of linear distance between them. Every point was observed for 20 minutes beginning between 5:30 and 7:30 in the morning and between 3:30 and 5:30 in the afternoon, when the birds are usually most active. All of the birds that were perceived visually or detected through calls within 30 m of the observer were counted. The researchers walked at a slow and constant speed to ensure proper, non-biased observations, and binoculars, tablets, DSLR cameras and photographic field guides were used to confirm the observations. All observations were recorded on the field.
The observed birds were identified based on their morphology, behaviour and calls according to Kennedy (2000) and Fisher and Hicks (2006), and the ecological status, i.e., endemism, of the identified birds was determined using the same field guides. Conservation status was determined using the data provided by the International Union for the Conservation of Nature (IUCN 2014), and the published literature and field guides were used to determine the feeding guilds of the identified species.
Birds Species Diversity
The relative abundances of the bird species (%) were calculated using the equation N/n, where N is the species × the total number of individuals captured, and n is the total number of all species. Relative abundance of species and relative abundance of all species per land-use type were measured, and the avifaunal diversity in each habitat was compared using the reciprocal form of Simpson’s index (1/D) as 1/D = Σpi2 (where 1/D = reciprocal of Simpson’s D; Σpi2 = abundance of the common genera; pi = the proportion of the individual in the ith species) and calculated using Biodiversity Pro 2.0 software (McAleece et al. 1997). Furthermore, Bray-Curtis (single-link) cluster analysis was used to determine the percent similarity of the bird communities among the different land-use types in terms of diversity and foraging structure.
RESULTS
Diversity of Birds in Different Land-use Types
The point count method was employed to identify the bird species from three habitat types: i) agroforests; ii) ricefields; iii) roads and heavily disturbed areas. A total of 1114 individuals of 33 species from 24 families were recorded (Table 1). Of the 3 habitats, the highest percentage of individual sightings was in the agroforest, comprising the 44.16% of the total number of individuals, followed by ricefields (40.57%) and roads (15.26%).
Table 1:
Bird species recorded from three land-use types in lowland south-central Mindanao, Philippines.
| Family | Species name | Endemicity | Status | Feeding guild | Agroforests | Rice fields | Roads |
|---|---|---|---|---|---|---|---|
| Accipitridae | Haliastur indus | Resident | LC | Carnivore | 0 | 2 | 0 |
| Alcenidae | Halcyon chioris | Resident | LC | Carnivore | 29 | 20 | 2 |
| Apodidae | Callocalia esculenta | Resident | LC | Insectivore | 18 | 4 | 25 |
| Ardeidae | Ardeola speciosa | Migrant | LC | Carnivore | 5 | 4 | 0 |
| Bubulcus Ibis | Migrant | LC | Carnivore | 0 | 30 | 0 | |
| Egretta garzetta | Migrant | LC | Carnivore | 0 | 40 | 5 | |
| Artamidae | Artamus leucorynchus | Resident | LC | Insectivore | 2 | 14 | 3 |
| Campephagidae | Lalage nigra | Resident | LC | Insectivore | 12 | 1 | 0 |
| Columbidae | Chaicophaps indica | Resident | LC | Frugivore | 6 | 0 | 3 |
| Geopellra striata | Resident | LC | Granivore | 37 | 25 | 8 | |
| Phapitreron amethystina | Philippine Endemic | LC | Frugivore | 5 | 0 | 2 | |
| Stigmatopella chinensis | Resident | LC | Granivore | 18 | 6 | 3 | |
| Streptopella tranquebarica | Resident | LC | Granivore | 0 | 5 | 0 | |
| Corvidae | Corvus cf. macrorhynchos | Resident | LC | Carnivore | 15 | 5 | 3 |
| Cuculidae | Centropus viridis | Philippine Endemic | LC | Insectivore | 2 | 0 | 1 |
| Dicaeidae | Dicaeum australe | Philippine Endemic | LC | Nectarinivore | 8 | 0 | 0 |
| Estrildidae | Lonchura oryzivora | Resident | LC | Granivore | 21 | 29 | 2 |
| Padda oryzlvora | Introduced | VU | Granivore | 17 | 11 | 3 | |
| Hirundinidae | Hirundo tahitica | Resident | LC | Insectivore | 58 | 74 | 21 |
| Tachycineta blcolor | Resident | LC | Insectivore | 34 | 30 | 10 | |
| Laniidae | Lanius cristatus | Migrant | LC | Insectivore | 23 | 8 | 9 |
| Locustellidae | Megalurus paiustris | Resident | LC | Insectivore | 1 | 2 | 0 |
| Meropidae | Merops philippinus | Resident | LC | Insectivore | 12 | 41 | 0 |
| Muscicapidae | Rhipidura Javanica | Resident | LC | Insectivore | 22 | 8 | 8 |
| Muscicapa sp. | Resident | LC | Insectivore | 2 | 0 | 0 | |
| Nectariniidae | Nactarinia jugularis | Resident | LC | Nectarinivore | 25 | 0 | 9 |
| Oriolidae | Oriolus chinensis | Resident | LC | Frugivore | 12 | 0 | 14 |
| Passeridae | Passer montanus | Resident | LC | Granivore | 18 | 30 | 9 |
| Pycnonotidae | Pycnonotous goiavier | Resident | LC | Frugiore | 64 | 11 | 3 |
| Scolopacidae | Tringa stagnatilis | Migrant | LC | Carnivore | 0 | 9 | 0 |
| Sturnidae | Aplonis panayensis | Resident | LC | Frugiore | 21 | 27 | 25 |
| Turdidae | Copsychus saularls | Resident | LC | Insectivore | 5 | 0 | 2 |
| Cisticolidae | Cisticola juncidls | Resident | LC | Insectivore | 0 | 16 | 0 |
| Abundance | 1114 | 492 | 452 | 170 | |||
| Species richness | 33 | 27 | 25 | 22 |
Note LC’ stands for least concern’: VU’ stands for ‘vulnerable’
In the agroforests, a total of 497 individuals belonging to 27 species were recorded; Pycnonotous goiavier was the most frequently recorded species, representing 13% of the total sightings, followed by Hirundo tahitica (11.79%) and Geopelia striata (7.52%). Some species, such as Diceaum australe and Muscicapa sp., were only recorded in the agroforest habitat. In the ricefields, a total of 452 individuals were recorded representing 25 bird species. Ardeidae and Columbidae were the most well-represented families with 3 species each (11%) followed by Estrildidae and Hirundidae with 2 species (7%), and the rest of the families were each represented by a single species (3%). Of all of the species recorded in the area, H. tahitica (16.37%) was the most abundant of the recorded species. The area with the fewest individuals and species was the heavily disturbed habitat represented by roads; a total of 170 individuals and 22 species were recorded in this area. Columbidae was the most species-rich family with four species, namely, Chalcophaps indica, G. striata, Phapitreron amethystinus and Spilopelia chinensis. Two species, Collocalia esculenta and Aplonis payanensis, were the most often sighted species and represented 14.70% of the total number of individuals. Bird species diversity was determined by species dominance; the reciprocal form of the Simpson’s diversity index (1/D) was employed to measure the diversity of birds recorded from the three land-use types. The most diverse habitat was the agroforest (1/D = 16.146) followed by the ricefields (1/D = 13.934) and finally the roads and heavily disturbed habitat (1/D = 12.579), and this result was further supported by the degree of community similarity among the habitats. Using Bray-Curtis cluster analysis (Fig. 2), greater similarity was found between agroforests and ricefields in terms of species composition compared to areas with high disturbance and human encroachment, such as roads. The result of this investigation also demonstrated that as the structure of the vegetation changed and the amount of disturbance increased, the abundance of bird sightings also decreased. Fifteen species (45.46%) were common in all three habitat types, namely, A. payanensis, Artamus leucorynchus, C. esculenta, Corvus cf. macrorhynchos, G. striata, Halcyon chloris, H. tahitica, Lanius cristatus, Lonchura atricapilla, Padda oryzivora, Passer montanus, P. goiavier, Rhipidura javanica, Stigmatopelia chinensis and Tachycineta bicolor.
Figure 2:

Similarity of species composition among three habitat types in lowland south-central Mindanao, Philippines.
Of the 33 species recorded from the different habitat types, 3 species were Philippine endemics, namely, P. amethystina, Centropus viridis and Dicaeum australe. C. viridis is an endemic Philippine coucal, and it was only recorded in two habitat types, roads and agroforest. This species can be found alone or in pairs in a wide variety of habitats from grassland, mixed cultivation, secondary growth and primary forest up to 2000 masl. Coucals are shy and hard to see except when they are perching in the open or on the tops of grasses or small trees, and they have an undulating flight pattern and skulk through dense vegetation (Kennedy 2000). The columbid P. amethystina is an endemic dove species; it can be found in lowlands but prefers middle and higher elevations from 500 to 2000 masl in primary and secondary forest (Kennedy 2000). In the study site, it was recorded in flocks or in pairs in trees along roads and deep inside the agroforest. The endemic Philippine flowerpecker, D. australe, was only recorded foraging in the agroforest habitat, and according to Kennedy (2000), this species dwells in the canopies of forests, forest edges, secondary growth, and shrubs and fruiting tress in open country. It occurs singly or in groups and mixed flocks, usually below 1000 masl. Based on the data collected in this study, no endemic species were recorded in the ricefields, but wetland and migrant species such as Ardeola speciosa, Bubulcus ibis, Egretta garzetta and Tringa stagnitilis were found foraging in the ricefields. The occurrence of T. stagnitilis, a rare migrant in the Philippines, is noteworthy; this species was observed in a flock feeding on insects in the paddy of a ricefield. Another noteworthy finding is the new Mindanao record of Streptopelia tranquebarica, which had previously only been known to occur in Bataan, Calayan, Lubang, Mindoro and Luzon (Kennedy 2000) but was recorded foraging in the ricefields in the study site.
The conservation status of the bird species was assessed using the latest information available from the IUCN Red List. Of all of the recorded species, only one was determined to be threatened; P. oryzivora is considered to be Vulnerable by the IUCN Red List (IUCN 2014). According to the IUCN, this species is endemic to the islands of Java, Bali and probably Madura, Indonesia, but it has been widely introduced. Its population had been increasing, but extensive trapping for the domestic and international caged bird trade has probably been underway for centuries, reaching a peak in the 1960s and 1970s, and is the main cause of the population decline. The tendency of the species to flock, particularly at roost sites, renders it especially susceptible to mass trapping, and even feral populations, which were originally introduced through trade, have been decimated. In this study, this species was recorded in all of the habitat types.
Avifaunal Foraging Structure
A feeding guild is a group of species that exploits similar food resources in a habitat, and its characterisation is usually based on the type of food being consumed, which in turn determines the feeding behaviour of the different bird species and the availability of food resources. Foraging guilds can be a useful way to compare changes between species-rich communities because their functional organisation can be investigated even if no species are shared (Terborgh & Robinson 1986). Five types of feeding guild were identified in the study area: carnivore, frugivore, insectivore, granivore and nectarivore. The entire community and all 3 of the habitats were dominated by insectivores with 36% in agroforests, 30% in ricefields and 24% in heavily disturbed habitats (Fig. 3). A. leucorynchus, C. esculenta, G. striata, H. tahitica, L. cristatus, R. javanica and T. bicolor were the most common insectivore species occurring in the study site.
Figure 3:
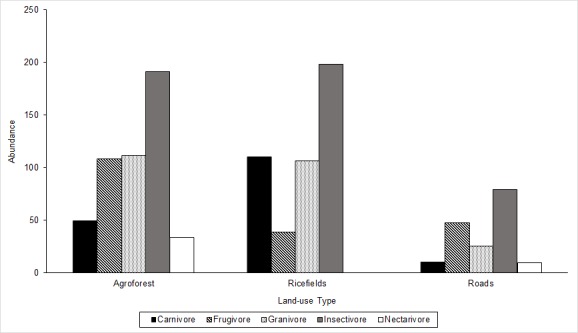
Bird abundance in terms of feeding guild in different land-use types in lowland south-central Mindanao, Philippines.
The similarity in feeding guild structure was greater between agroforests and ricefields with 79.98% similarity compared to roads (51.76%) (Fig. 4). Frugivorous bird species were most abundant in agroforests and roads with five species (15%) in each habitat, and the most abundant frugivores in the agroforest habitat were P. goiavier and A. payanensis; these species were observed foraging in fruiting trees with small and ripe fruits. Along roads, A. payanensis was the most abundant frugivore, constituting 14.75% of the total number of observations. This species was observed perching on wooden posts besides roads and other types of infrastructure in urban sites. The fewest number of frugivore species, two (6%), was recorded in the ricefields, but carnivorous species were primarily observed in the ricefields. The most abundant carnivorous species were E. garzetta (8.85%) followed by B. ibis (6.64%) and H. chloris (4.42%), and the large carnivores Haliastur indus and C. macrorhynchos were observed hovering and foraging in rice fields on small vertebrates, such as rodents. Two species of carnivores were recorded in both the agroforests and roads. H. chloris (5.9%) and E. garzetta (3%) were the most abundant carnivores in the agroforest habitat and roads, respectively, but the E. garzetta observation may be accidental as there was only a single sighting throughout the duration of sampling. Species from the Estrildidae family dominated most of the granivore species; P. montanus was the most abundant and dominant species in the rice fields and was followed by L. atricapilla and P. oryzivora, all of which were present in all three habitats. Three additional granivores were the dove species, G. striata, Stegmatopelia chinensis and S. tranquebarica. Moreover, only two species of nectarivorous species were recorded; D. australe was only recorded in the agroforest habitat, and Nectarinia jugularis was recorded in both the agroforest and along roads. Both species were absent from the rice fields.
Figure 4:

Similarity of the feeding guilds in different land-use types in lowland south-central Mindanao, Philippines.
DISCUSSION
The results of this study suggest that higher bird diversity is found in agroforests than in ricefields and roads, which conform to the conclusions of Azman et al. (2011) concerning the structure of bird communities in different habitat types; bird diversity is higher in areas with high vegetation diversity, such as primary and secondary forests. The agroforest examined in this study is described as a mixture of tall, large fruit-bearing trees with a wide canopy cover, and the greater diversity of birds in this habitat may be influenced by the presence of important resources that allow birds to tolerate disturbance due to land use change. Most of the species identified in this habitat were residents, and few were forest-dependent. However, although it contains some endemic species, the diversity of the habitat is still very low compared to the forest habitats in the region (see Achondo et al. 2011, 2014). Of all of the bird species recorded, Pycnonotus goiavier was the most abundant, and this species belongs to the Pycnonotidae family, a group important to forest restoration due to their efficient seed dispersal activity in lowland forests (Sritongchuay et al. 2014). The overall result of this study agrees with the findings of Harvey and Villalobos (2007) that agroforests contain bird assemblages that are as abundant, species-rich and diverse as natural forest ecosystems. However, the structure of the community was highly altered with fewer forest-dependent species and different dominant species, so bird diversity is more closely related to the structural and floristic characteristics of the different land use types. One of the important benefits of agroforests to biodiversity is the provision of habitat for tolerant species and corridors that connect different habitats (Jose 2009). Birds plays a vital role in seed dispersal and forest recovery in agroforest ecosystems in that they increase seed deposition, which can result in high seedling regeneration of species that have not been locally present in previous years (Lozada et al. 2007). According to Van Bael et al. (2007), the presence of large tress in an agroforest habitat provides important breeding niches and food resources for many bird species, and this may influence the high diversity of birds in the habitat. Almazán-Núñez et al. (2015) found that bird abundance was higher in natural forests in the dry season, and the high species richness of dispersers and seed predators was positively correlated with vegetation complexity. Waltert et al. (2005) also found that young secondary forests and agroforests sustain high numbers of bird species that are similar to adjacent, near-primary forest. Greater conservation value has been found in agroforest habitats than in crop monocultures, and traditional agroforestry systems are often complex and more supportive of diversity than monocultures. However high levels of wild biodiversity may often depend on proximity to natural habitat, which is still most favourable to many wildlife species, such as birds (Mcneely & Schroth 2006; Harvey & Villalobos 2007). Forests and agroforests that have been replaced with simplified agricultural systems will shift towards less specialised bird communities with altered proportions of functional groups (Sekercioglu 2012). Furthermore, the proximity of agroecosystems to natural forest is vital to the maintenance of bird diversity and ecological function (Tscharntke et al. 2008).
Following agroforests, the ricefield habitat was the second most diverse habitat in terms of birds. The similarity index between agroforest and ricefields was 79.91%, and the birds recorded in rice fields included wetland and migrant species such as A. speciosa, B. ibis, E. garzetta, L. cristatus and T. stagnatilis. Ricefields function as temporary wetlands with conditions that change drastically over the course of the year. This habitat type is essential for many bird species; it provides valuable and suitable habitat for the foraging and breeding activities of various kinds of bird species including migrants (Tourenq et al. 2001). Seasonal change is one of the temporal effects on the habitats used by bird species (Tourenq et al. 2001) such as T. stagnatilis, which utilises ricefields for foraging and breeding during the post-harvest season and field preparation. This species inhabits warm inland wetlands from open steppes to boreal forests including shallow freshwater and grassy or marshy lake edges (Kennedy 2000; Birdlife International 2012). In this study, flocks of T. stagnatilis were found feeding on many serious ricefield paddy pests, which further suggests that the post-harvest season and field preparation can result in important surges in prey availability for birds. According to Comin et al. (2001), wetland birds prefer habitats with intermediate plant cover for resting and sleeping rather than ricefields or very open or very dense wetlands with tall vegetation. However, ricefields play an important role in mitigating the impact of wetland modification on many bird species, but no legal protections for this habitat type have been proposed (Sanchez-Guzman et al. 2007). Additionally, ricefields serve as refuges for wetland birds during years when marshes and other wetlands are dry (Tourenq et al. 2001). Farming strategies and ricefield cultivation may also affect the presence and diversity of wetland bird species (Tourenq et al. 2001). Wood et al. (2010) stated that traditional practices with one crop per year and long periods of fallow flooding may be beneficial to many important species. In contrast, modern intensive farming with multiple cropping seasons and high chemical usage may be implicated in the decline of biodiversity. Moreover, flooded fields apparently provide foraging habitat that is equivalent to semi-natural wetlands with reduced predation, making it a safer habitat for many wetland birds. Therefore, if ricefields are managed properly, they can be a valuable agricultural habitat for many wetland birds (Elphick 2000), and for migrant species, such as E. garzetta, A. bacchus, and B. ibis, ricefields are important feeding, stopover and wintering sites (Acosta et al. 2010).
To further determine the structure of bird communities related to the modification of habitat types, roads and heavily disturbed habitats were assessed. The lowest species diversity was found in this habitat type with 51% similarity to the agroforests and ricefields. The increasing prevalence of roads is one potential contributor to the worldwide decline in bird populations (Kociolek et al. 2011). The study by Palomino and Carrascal (2007) revealed that proximity to urban sites does not affect the total bird species richness in natural habitats, which conforms to the results of this study. However, bird abundance increases with a reduction in the distance to the nearest urban development. Studies of bird diversity in cities and urbanised landscapes have revealed significant impacts on bird species richness and relative abundance (Gatesire et al. 2014), and increasing diversity from more to less urbanised areas has also been observed in previous studies (Clergeau et al. 1998). Road density has also been found to influence bird communities (Minor & Urban 2009), and the availability of food resources along roads and in urbanised habitats is an important factor that can explain the low diversity in our study. Further, to mitigate the negative effects of urbanisation, the use of native vegetation is recommended in urban sites, such as roads and buildings (Mills et al. 1989), and edge effects might be partially mitigated through vegetation management and restoration (Kociolek et al. 2011).
As a whole, the most commonly encountered species among the habitats were as follows: C. esculenta, G. striata, H. tahitica, L. malacca, N. jugularis, P. montanus, P. goiavier, R. javanica and T. bicolor. These birds are mostly associated with human communities and some are generalist feeders. Their characteristics allow them to easily adapt to their environments, so they are of lower risk of extinction (Blair 1996). Very low endemicity was noted in the agroforest with only three Philippine endemic species (9%) recorded, and only two endemic species were recorded along roads with no endemic records from the ricefields. Both P. amethystina and D. australe are usually found in primary and secondary forest as well as fruiting and flowering trees from 2000 masl in elevation and below while C. viridis usually prefers grasslands, mixed cultivated areas, forests and secondary growth (Kennedy 2000). However, the majority of the species were residents (64%), such as A. payanensis, H. tahitica, G. striata, T. bicolor and L. atricapilla. According to Paz et al. (2013), more endemic species are present in advanced secondary and old growth forests, and island-endemic birds are thought to seldom use disturbed habitats, such as plantations and agroforests, due to the evolution of specialised morphological and behavioural characteristics, which results in a preference for specific natural forest resources. However, the presence of tropical trees in land use types is important for the maintenance of many resident bird species (Waltert et al. 2005), and agroforests can still support a high number of endemic species. A study in central Sulawesi (Abrahamczyk et al. 2008) revealed that agroecosystems, particularly cacao plantations, may maintain highly diverse forest tree cover, which when properly managed can harbour up to 60% of the forest specialists and endemic birds species. Given the importance of habitat structure, vegetation and levels of human disturbance may be important determinants of endemic bird diversity, species richness and abundance.
Feeding Guilds
The foraging habits of the identified birds were assessed to explore the variation in avifaunal composition among habitat types; according to Pearman (2002), variation in vegetation structure affects the distribution of bird foraging guilds. Five feeding guilds were assessed by the study and insectivores were the most dominant group. The insectivorous feeding guild was mostly composed of species from families Apodidae and Hirundidae, such as C. esculenta and H. tahitica, respectively. In the ricefields and agroforests, the insectivore H. tahitica was the most abundant species, and insectivorous species diversity was found to be greater in agroforests compared to monoculture crops and heavily disturbed habitats, such as ricefields and roads, respectively. According to Blake and Loiselle (2001), insectivores are often found to be the most species rich and abundant in tropical forests, which supports the findings of Rajashekara and Venkatesha (2014), who found greater numbers of insectivores in agroforests. The availability of a variety of food sources for both adults and young and safe habitats for nesting and roosting in and around agroecosystems are important for the occurrence and abundance of insectivorous species. On the one hand, insectivores in agroforests provide benefits to farms by reducing the populations of insect pests in agricultural habitats, which further enhances the conservation value of farms for birds and other wildlife species (Johnson et al. 2010). Laurance et al. (2004) further stated that, even without the noise and traffic, roads have greatly affected populations of insectivorous bird species, which is consistent with the low insectivore abundance and richness found along roads in the study site.
Next to insectivores, carnivorous birds were the second most abundant group, particularly in ricefields. The most abundant species were E. garzetta and B. ibis, which conforms to the findings of Munira et al. (2014) that carnivorous species are abundant before the harvest season in northern Peninsular Malaysia. The presence of a high number of carnivorous species in ricefields was primarily influenced by the abundance of food sources, such as small frogs, fish, molluscs and small vertebrate species. Stafford et al. (2010) stated that the abundance of carnivorous bird species in ricefields may be due to the large quantity of food resources, such as polychaetes, crustaceans and mollusks, in the rice plots during the migration season. Furthermore, King et al. (2010) noted that the rice fields in many countries support large numbers of migratory waterbirds and may be important for many species.
During the rice-growing cycle, fields have been found to be most important to carnivorous birds during the sowing stage, whereas post-harvest flooded fields are most valuable for granivorous species (Acosta et al. 2010). This is supported by the results of this study in which greater granivore species richness was found in ricefields compared to other habitat types, and the most abundant granivore species include G. striata, L. atricapilla, P. oryzivora, P. montanus and S. chinensis. Columbids, such as G. striata and S. chinensis, were commonly observed on the ground and in grassy habitats were they would pick grains from plant sources such as rice and grasses. P. montanus is a species that is known to occur throughout the year in ricefields and habitats inhabited by humans, and L. atricapilla was once considered to be the national bird of the Philippines and is usually found in flocks consuming grains and seeds in ricefields, grasslands and open country. Both species (P. montanus and L. atricapilla) are considered to be pests by local rice farmers in the Philippines (Kennedy 2000). Moorcroft et al. (2002) concluded that fields left fallow after harvest (i.e., stubble fields) support high wintering densities of many species of granivorous birds, and they emphasised that variation in the abundance and availability of weeds affects the diversity of granivorous species. Furthermore, the presence of a high diversity of granivores in a habitat indicates habitat disturbance (Gray et al. 2007).
More frugivorous birds were found in agroforests compared to ricefields, and the abundance of the available food resources in the agroforest is one reason for the high species richness in the habitat. The abundance and richness of fruiting plants is important and associated with the diversity of frugivorous bird species and foraging behaviours in certain habitat types (Moegenburg & Levey 2003). The related findings of Blake and Loiselle (1991) that the capture rate of frugivores in primary forest is generally greater during periods when fruit is abundant, and the presence of species belonging to the family Sturnidae, Pycnonotidae, Columbidae and Oriolidae is a good indicator of forest regeneration in semi-degraded/disturbed habitats such as agroforest. The tolerance of frugivorous species to degraded landscapes is important during initial forest succession and restoration in tropical regions (Herrera 1984; Corlett 1998). Finally, nectarivores were the least abundant in all of the habitats and were absent in ricefields. In the agroforest habitat, two species were recorded including the endemic flowerpecker D. australe, and the presence of a nectarivore in an agroforest may be explained by the availability of flowering resources and the seasonality of flowering. Fleming (1992) emphasised that the structure and composition of avian communities changes in space and time with the availability of food resources, and variation tends to be most visible among bird species that feed on patchy and temporary food resources, such as nectar and fruit. Waltert et al. (2005) found fewer species of nectarivores in agroecosystems as the intensity of land modification increased. Cotton (2006) added that the abundance and diversity of nectarivores are correlated with an increase in nectar availability. In this study, the nectarivores were the least abundant and most poorly documented guild; Loiselle (1988) noted that nectarivores are difficult to observe because of their small size and are likely underestimated relative to other guilds. Furthermore, according to Li et al. (2013), higher bird species richness was found in native forest than in rubber plantations, and the bird communities varied significantly among the two habitat types. No strict frugivores were found in rubber agroforests, and no granivores were found in semi-deciduous monsoon forest; the species richness of insectivores and nectarivores was lower in rubber agroforest than in native forest.
This study provided information about the diversity and structure of bird communities in different land-uses in lowland south-central Mindanao, and the results suggest that the availability of diverse food sources and foraging and roosting sites are essential factors for the diversity and abundance of bird species in certain habitat types. In a similar case in Malaysia, Azman et al. (2011) concluded that the conversion of forest to agriculture, such as to oil palm plantations or paddy fields, causes changes in bird diversity and the distribution of avian guilds. Based on their study of rubber plantations and natural forests, Li et al. (2013) noted that native forests are still important habitats for bird species as richness was higher in the native versus the agroforest, which supports the results of this study. Furthermore, this suggests that to conserve native, endemic and migrant species in all land-use systems, important food sources and habitat structure for a variety of bird species must be maintained. Moreover, if deforestation in Southeast Asia continues, it will increase the probability of extinction of many bird species in the region (Brooks et al. 1997).
CONCLUSION
The factors influencing avifaunal diversity and the structure of foraging guilds were elucidated in this study, and the following conclusions were drawn: i) among all land-use types, the highest avifaunal diversity was recorded in the agroforest; ii) higher species similarity was found between the agroforest and the ricefields than the roads and urban sites; iii) a low number of endemic species was found in all of the land-use types; iv) insectivorous and frugivorous species were more abundant in agroforests and ricefields; and v) vegetation structure and the availability of food resources and foraging/nesting sites are important for bird species in different land use types.
Studies of the effects of land-use on birds in the Philippines may be scarce, so further study of additional factors affecting bird diversity in different degraded habitats is recommended, such as considering spatio-temporal factors and the availability of specific food resources.
Acknowledgments
The researchers are very grateful to the experts and critics who provided constructive comments and suggestions, which greatly improved this paper. Furthermore, we gratefully acknowledge a small grant from the project funded by the Rufford Foundation that also contributed to the success of this paper.
REFERENCES
- Abrahamczyk S, Kessler M, Dwi Putra D, Waltert M, Tscharntke T. The value of differently managed cacao plantations for forest bird conservation in Sulawesi, Indonesia. Bird Conservation International. 2008;18(4):349–362. [Google Scholar]
- Achondo MJMM, Fernandez-Casim L, Suetos KJE, Agduma AR, Bretana BLP, Supremo JP, Salem JGC, Mancao LS, Bello VP. Do conservation areas support avifaunal diversity? Alliance Research Publication. 2014;1(1):19–31. [Google Scholar]
- Achondo MJMM, Casim LF, Bello VP, Tanalgo KC, Agduma AR, Bretaña BLP, Salem JGC, Mancao LS, Supremo JP. Rapid assessment and feeding guilds of birds in selected rubber and oil palm plantations in North Cotabato. Asian Journal of Biodiversity. 2011;2(1):103–120. [Google Scholar]
- Acosta M, Mugica L, Blanco D, López-Lanús B, Dias RA, Doodnath LW, Hurtado J. Birds of rice fields in the Americas. Waterbirds. 2010;33(1):105–122. [Google Scholar]
- Almazán-Núñez RC, Arizmendi MDC, Eguiarte LE, Corcuera P. Distribution of the community of frugivorous birds along a successional gradient in a tropical dry forest in south-western Mexico. Journal of Tropical Ecology. 2015;31(01):57–68. [Google Scholar]
- Azman NM, Latip NSA, Sah SAM, Akil MAMM, Shafie NJ, Khairuddin NL. Avian diversity and feeding guilds in a secondary forest, an oil palm plantation and a paddy field in riparian areas of the Kerian River Basin, Perak, Malaysia. Tropical Life Sciences Research. 2011;22(2):45–64. [PMC free article] [PubMed] [Google Scholar]
- Birdlife International . Birdlife International country profile: Philippines. 2012. http://www.birdlife.org/datazone/country/philippines (accessed on 5 November 2014) [Google Scholar]
- Blair RB. Land use and avian species diversity along an urban gradient. Ecological Applications. 1996;6(2):506–519. [Google Scholar]
- Blake JG, Loiselle BA. Bird assemblages in second-growth and old-growth forests, Costa Rica: Perspectives from mist nets and point counts. The Auk. 2001;118(2):304–326. [Google Scholar]
- Blake JG, Loiselle BA. Variation in resource abundance affects capture rates of birds in three lowland habitats in Costa Rica. The Auk. 1991;180(1):114–130. [Google Scholar]
- Brooks TM, Pimm SL, Collar NJ. Deforestation predicts the number of threatened birds in insular Southeast Asia. Conservation Biology. 1997;11(2):382–394. [Google Scholar]
- Carandang AP. Forest resource assessment—National forest assessment: Forestry policy analysis: Philippine. Philippines: Food and Agriculture Organization (FAO); 2005. [Google Scholar]
- Clergeau P, Savard JPL, Mennechez G, Falardeau G. Bird abundance and diversity along an urban-rural gradient: A comparative study between two cities on different continents. Condor. 1998;100(3):413–425. [Google Scholar]
- Comín FA, Romero JA, Hernández O, Menéndez M. Restoration of wetlands from abandoned rice fields for nutrient removal, and biological community and landscape diversity. Restoration Ecology. 2001;9(2):201–208. [Google Scholar]
- Corlett RT. Frugivory and seed dispersal by vertebrates in the oriental (Indomalayan) region. Biological Reviews of the Cambridge Philosophical Society. 1998;73:413–448. doi: 10.1017/s0006323198005234. [DOI] [PubMed] [Google Scholar]
- Cotton PA. Seasonal resource tracking by Amazonian hummingbirds. Ibis. 2006;149:135–142. [Google Scholar]
- Curran LM, Trigg SN, McDonald AK, Astiani D, Hardiono YM, Siregar P, Kasischke E. Lowland forest loss in protected areas of Indonesian Borneo. Science. 2004;303(5660):1000–1003. doi: 10.1126/science.1091714. [DOI] [PubMed] [Google Scholar]
- Elphick CS. Functional equivalency between rice fields and seminatural wetland habitats. Conservation Biology. 2000;14(1):181–191. [Google Scholar]
- Elphick CS, Taft O, Lourenço PM. Management of rice fields for birds during the non-growing season. Waterbirds. 2010;33(sp.1):181–192. [Google Scholar]
- Fisher T, Hicks N. A photographic guide to birds of the Philippines. London: New Holland Publishers; 2006. [Google Scholar]
- Fleming TH. How do fruit- and nectar-feeding birds and mammals track their food resources? In: Hunter MD, Ohgushi T, Price PW, editors. Effects of resource distribution on animal-plant interactions. San Diego: Academic; 1992. pp. 355–391. [Google Scholar]
- Forest Management Bureau . The Philippine forestry statistics 2013. Philippines: Department of Environment and Natural Resources; 2013. http://forestry.denr.gov.ph/statbook.htm (accessed on 12 December 2014). [Google Scholar]
- Gatesire T, Nsabimana D, Nyiramana A, Seburanga JL, Mirville MO. Bird diversity and distribution in relation to urban landscape types in Northern Rwanda. The Scientific World Journal. 20142014 doi: 10.1155/2014/157824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gray MA, Baldauf SL, Mayhew PJ, Hill JK. The response of avian feeding guilds to tropical forest disturbance. Conservation Biology. 2007;21(1):133–141. doi: 10.1111/j.1523-1739.2006.00557.x. [DOI] [PubMed] [Google Scholar]
- Harvey CA, Villalobos JAG. Agroforestry systems conserve species-rich but modified assemblages of tropical birds and bats. Biodiversity and Conservation. 2007;16(8):2257–2292. [Google Scholar]
- Heaney LR. Biodiversity patterns and the conservation of mammals in the Philippines. Asia Life Sciences. 1993;2:261–274. [Google Scholar]
- Herrera CM. A study of avian frugivores, bird-dispersed plants, and their interaction in Mediterranean scrublands. Ecological Monographs. 1984;54:1–23. [Google Scholar]
- International Union for the Conservation of Nature (IUCN) The IUCN Red List of threatened species. 2014 http://www.iucnredlist.org/ (accessed on 7 November 2014). [Google Scholar]
- Johnson MD, Kellermann JL, Stercho AM. Pest reduction services by birds in shade and sun coffee in Jamaica. Animal Conservation. 2010;13(2):140–147. [Google Scholar]
- Jose S. Agroforestry for ecosystem services and environmental benefits: An overview. Agroforestry Systems. 2009;76(1):1–10. [Google Scholar]
- Kennedy R, editor. A guide to the birds of the Philippines. Oxford: Oxford University Press; 2000. [Google Scholar]
- King S, Elphick CS, Guadagnin D, Taft O, Amano T. Effects of landscape features on waterbird use of rice fields. Waterbirds. 2010;33:151–159. [Google Scholar]
- Kociolek AV, Clevenger AP, St Clair CC, Proppe DS. Effects of road networks on bird populations. Conservation Biology. 2011;25(2):241–249. doi: 10.1111/j.1523-1739.2010.01635.x. [DOI] [PubMed] [Google Scholar]
- Laurance SG, Stouffer PC, Laurance WF. Effects of road clearings on movement patterns of understory rainforest birds in central Amazonia. Conservation Biology. 2004;18(4):1099–1109. [Google Scholar]
- Li S, Zou F, Zhang Q, Sheldon FH. Species richness and guild composition in rubber plantations compared to secondary forest on Hainan Island, China. Agroforestry Systems. 2013;87(5):1117–1128. [Google Scholar]
- Loiselle BA. Bird abundance and seasonality in a Costa Rican lowland forest canopy. Condor. 1988;90:761–772. [Google Scholar]
- Lozada T, De Koning GHJ, Marché R, Klein AM, Tscharntke T. Tree recovery and seed dispersal by birds: comparing forest, agroforestry and abandoned agroforestry in coastal Ecuador. Perspectives in Plant Ecology, Evolution and Systematics. 2007;8(3):131–140. [Google Scholar]
- McAleece N, Gage JDG, Lambshead PJD, Paterson GLJ. BioDiversity professional statistics analysis software. UK: Scottish Association for Marine Science and the London Natural History Museum; 1997. [Google Scholar]
- McNeely JA, Schroth G. Agroforestry and biodiversity conservation–traditional practices, present dynamics, and lessons for the future. Biodiversity & Conservation. 2006;15(2):549–554. [Google Scholar]
- Mills GS, Dunning JB, Jr, Bates JM. Effects of urbanization on breeding bird community structure in southwestern desert habitats. Condor. 1989;91:416–428. [Google Scholar]
- Minor E, Urban D. Forest bird communities across a gradient of urban development. Urban Ecosystems. 2010;13(1):51–71. [Google Scholar]
- Moegenburg SM, Levey DJ. Do frugivores respond to fruit harvest? An experimental study of short-term responses. Ecology. 2003;84(10):2600–2612. [Google Scholar]
- Moorcroft D, Whittingham MJ, Bradbury RB, Wilson JD. The selection of stubble fields by wintering granivorous birds reflects vegetation cover and food abundance. Journal of Applied Ecology. 2002;39(3):535–547. [Google Scholar]
- Munira A, Salmi AN, Anuar MS, Muin MMA, Abdul M, Amirrudin A, Juliani SN. Diversity and temporal distribution of birds in rice-growing landscape, Northern Peninsular Malaysia. Sains Malaysiana. 2014;43(4):513–520. [Google Scholar]
- Myers N. Threatened biotas: “Hot spots” in tropical forests. Environmentalist. 1988;8(3):187–208. doi: 10.1007/BF02240252. [DOI] [PubMed] [Google Scholar]
- Paz SL, Ngoprasert D, Nuneza OM, Mallari NAD, Gale GA. Philippine-endemic and Mindanao-endemic bird communities on Canticol and Mt. Hilonghilong, Philippines. Asian Journal of Biodiversity. 2013;4(1):135–168. [Google Scholar]
- Palomino D, Carrascal LM. Threshold distances to nearby cities and roads influence the bird community of a mosaic landscape. Biological Conservation. 2007;140(1):100–109. [Google Scholar]
- Pearman PB. The scale of community structure: habitat variation and avian guilds in tropical forest understory. Ecological Monographs. 2002;72(1):19–39. [Google Scholar]
- Philpott SM, Soong O, Lowenstein JH, Pulido AL, Lopez DT, Flynn DF, DeClerck F. Functional richness and ecosystem services: Bird predation on arthropods in tropical agroecosystems. Ecological Applications. 2009;19(7):1858–1867. doi: 10.1890/08-1928.1. [DOI] [PubMed] [Google Scholar]
- Rajashekara S, Venkatesha MG. Insectivorous bird communities of diverse agro-ecosystems in the Bengaluru region, India. Journal of Entomology and Zoology Studies. 2014;2(5):142–155. [Google Scholar]
- Sánchez-Guzmán JM, Morán R, Masero JA, Corbacho C, Costillo E, Villegas A, Santiago-Quesada F. Identifying new buffer areas for conserving waterbirds in the Mediterranean basin: The importance of the rice fields in Extremadura, Spain. Biodiversity and Conservation. 2007;16(12):3333–3344. [Google Scholar]
- Sekercioglu CH. Bird functional diversity and ecosystem services in tropical forests, agroforests and agricultural areas. Journal of Ornithology. 2012;153(1):153–161. [Google Scholar]
- Sritongchuay T, Gale GA, Stewart A, Kerdkaew T, Bumrungsri S. Seed rain in abandoned clearings in a lowland evergreen rain forest in Southern Thailand. Tropical Conservation Science. 2014;7(3):572–585. [Google Scholar]
- Stafford JD, Kaminski RM, Reinecke KJ. Avian foods, foraging and habitat conservation in world rice fields. Waterbirds. 2010;33(sp1):133–150. [Google Scholar]
- Sutherland WJ, Newton I, Green R. Bird ecology and conservation: A handbook of techniques: A handbook of techniques. UK: Oxford University Press; 2004. [Google Scholar]
- Temple SA, Wiens JA. Bird populations and environmental changes: Can birds be bio-indicators? American Birds. 1989;43(2):260–270. [Google Scholar]
- Terborgh J, Robinson S. Guilds and their utility in ecology. In: Kikkawa J, Anderson DJ, editors. Community ecology: Pattern and process. London: Blackwell Scientific Publications; 1986. [Google Scholar]
- Tourenq C, Bennetts RE, Kowalski H, Vialet E, Lucchesi JL, Kayser Y, Isenmann P. Are ricefields a good alternative to natural marshes for waterbird communities in the Camargue, southern France? Biological Conservation. 2001;100(3):335–343. [Google Scholar]
- Tscharntke T, Sekercioglu CH, Dietsch TV, Sodhi NS, Hoehn P, Tylianakis JM. Landscape constraints on functional diversity of birds and insects in tropical agroecosystems. Ecology. 2008;89(4):944–951. doi: 10.1890/07-0455.1. [DOI] [PubMed] [Google Scholar]
- Utzurrum RCB. Philippine island biogeographic patterns: practical considerations for resource conservation and management. Association of Systematic Biology of the Philippines Communications. 1991;3:19–32. [Google Scholar]
- Van Bael SA, Bichier P, Ochoa I, Greenberg R. Bird diversity in cacao farms and forest fragments of western Panama. Biodiversity and Conservation. 2007;16(8):2245–2256. [Google Scholar]
- Vandermeer J, Perfecto I. The agroecosystem: A need for the conservation biologist’s lens. Conservation Biology. 1997;11(3):591–592. [Google Scholar]
- Waltert M, Bobo KS, Sainge NM, Fermon H, Mühlenberg M. From forest to farmland: Habitat effects on afrotropical forest bird diversity. Ecological Applications. 2005;15(4):1351–1366. [Google Scholar]
- Wenny DG, Devault TL, Johnson MD, Kelly D, Sekercioglu CH, Tomback DF, Whelan CJ. The need to quantify ecosystem services provided by birds. The Auk. 2011;128(1):1–14. [Google Scholar]
- Wood C, Qiao Y, Li P, Ding P, Lu B, Xi Y. Implications of rice agriculture for wild birds in China. Waterbirds. 2010;33(sp1):30–43. [Google Scholar]
- World Climatic Database WorldClim – Global climate data. 2014 http://www.worldclim.org/ (accessed on 9 November 2014) [Google Scholar]