

Abstract
Inflammatory responses in the blood vessel play a pivotal role in the pathogenesis of atherosclerosis. Eupatilin, a flavone derived from Artemisia princepsPampanini, has various pharmacological activities, including antioxidant, anti-tumor, and anti-inflammatory capacities. However, there has been no research examining the function of eupatilin on vascular inflammation. Therefore, the aim of this study was to investigate the effects of eupatilin on tumor necrosis factor (TNF)-α-induced human umbilical vein endothelial cells (HUVECs) activation and the underlying molecular mechanisms. Our findings showed that eupatilin reduced U937 cells adhesion to TNF-α-stimulated HUVECs and attenuated TNF-α-induced the expression of vascular cell adhesion molecule-1 (VCAM-1) and intercellular adhesion molecule-1 (ICAM-1) in HUVECs, as well as the production of intracellular reactive oxygen species (ROS). Moreover, eupatilininhibits TNF-α-induced phosphorylation of NF-kB p65 and MAPKs in HUVECs. Taken together, the results of the present study suggest that eupatilin inhibited inflammatory reaction through suppressing the ROS/MAPK-NF-ĸB pathway in HUVECs. Thus, eupatilin is proposed as an effective new anti-inflammatory agent to suppress vascular inflammation, and further prevent atherosclerosis.
Keywords: Eupatilin, atherosclerosis, inflammation, reactive oxygen species (ROS)
Introduction
Atherosclerosis, a chronic inflammatory disease, is a major factor leading to cardiovascular complications [1]. Under inflammatory conditions, monocyte recruitment to the endothelium is enhanced on the surface of endothelial cells by adhesion molecules, including vascular cell adhesion molecule-1 (VCAM-1), intercellular adhesion molecule-1 (ICAM-1) [2]. It is also well recognized thattumor necrosis factor (TNF)-α can trigger inflammation and oxidative stress responses in endothelial cells [3-5]. Therefore, preventing TNF-α-mediated inflammation or inhibiting the expression of cell adhesion molecules in vascular endothelial cells is considered to be a promising therapeutic target for vascular inflammatory diseases.
Eupatilin[2-(3,4-dimethoxyphenyl)-5,7-dihydroxy-6-methox-ychromen-4-one] is a pharmacologically active flavones derived from Artemisia princepsPampanini, which has been widely usedas an herbal medicine in Asia.An increasing amount of evidence has demonstrated that eupatilin exhibits radical scavenging, anti-allergic, anti-tumor, and anti-inflammatory activities [6-9]. For example, Choi et al. reported that eupatilin decreased lipopolysaccharide (LPS)-induced expression of inflammatory mediators and pro-inflammatory cytokines such as cyclooxygenase-2, monocyte chemoattractant protein-1, tumor necrosis factor-α, interleukin (IL)-1β and IL-6 in macrophages [10]. In addition, eupatilin has been demonstrated to possess anti-atherogenic effect. Recently, one study reported that eupatilin significantly inhibited platelet-derived growth factor (PDGF)-BB-induced proliferation and migration of human aortic smooth muscle cells [11]. However, to our knowledge, there has been no research examining the function of eupatilin on vascular inflammation. Therefore, the aim of this study was to investigate the effects of eupatilin on TNF-α-induced human umbilical vein endothelial cells (HUVECs) activation and the underlying molecular mechanisms.
Materials and methods
Cell culture
HUVECs were from the American Type Culture Collection (ATCC, Manassas, VA, USA) and maintained in DMEM containing 1800 mg/L NaHCO3, supplemented with 10% fetal bovine serum (FBS), 100 U/mL penicillin and 100 μg/mLstreptomycin in an incubator (Life Technologies, Baltimore, MD) with a humidified atmosphere of 5% CO2 at 37°C.
U937 monocyte-like cells (ATCC) were maintained in RPMI-1640 supplemented with 10% FBS, 100 U/mLpenicillin and 100 μg/mLstreptomycinin an incubator with a humidified atmosphere of 5% CO2 at 37°C.
Cell adhesion assay
U937 cells (1×105 cells/ml) were layered over TNF-α-treated HUVEC monolayers and incubated for 2 h. Thereafter, the cells were washed with phosphate buffer saline (PBS) and fixed in 4% paraformaldehyde in PBS. The adhesion U937 cells were counted observed under a Nikon E600 fluorescent microscope and expressed as adhesion U937 cells per high-power fields.
RNA extraction and quantitative reverse transcription polymerase chain reaction (RT-qPCR)
Total RNA of HUVECs was extracted using the TRIzol reagent (Invitrogen). The complementary DNA was synthesized from 5 μg of total RNA using M-MLV reverse transcriptase (Abcam, Cambridge, UK) according to the manufacturer’s instructions.The obtained complementary DNAs were then used as templates for RT-qPCR analysis.The primer sequences used for RT-PCR were as follows: VCAM-1 sense, 5’-CAAAGGTGGATCAGATTCAAG-3’ and anti-sense, 5’-GGTGAGCATTATCACCCAGAA-3’; ICAM-1 sense, 5’-CAAAGGTGGATCAGATTCAAG-3’ and anti-sense, 5’-GGTGAGCATTATCACCCAGAA-3’; β-actinsense, 5’-TCT GTG TGG ATT GGT GGC TCT A-3’ and anti-sense, 5’-CTG CTT GCT GAT CCA CAT CTG-3’. The primers were all synthesized by Sangon Biotech Co., Ltd. (Shanghai, China). The cycling conditions were as follows: 94°C for 2 min for initial denaturation; 94°C for 20 sec, 59°C for 15 sec, and 72°C for 20 sec; 2 sec for plate reading for 35 cycles; and a melt curve from 65 to 95°C. β-actin was used as an internal control. The expression levels of the relative genes were calculated using control β-actin mRNA and the 2-ΔΔCT method [12].
Western blot
Proteins were extracted from HUVECs and protein concentrations were measured by using the Bradford method. Equal amounts of protein were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred onto nitrocellulose membranes (Bio-Rad, Hercules, CA, USA). The membranes were blocked in 5% skimmed milk for 1 h at room temperature and incubated with primary antibodies overnight at 4°C. After washing with PBS containing 0.1% (v/v) Tween 20, membranes were incubated with horseradish peroxidase conjugated secondary antibodies for 1 h, followed by exposure using enhanced chemiluminescence detection reagents. The following antibodies were used: mouse anti-VCAM-1 (1:1,500), mouse anti-ICAM-1 (1:1,500), mouse anti-phospho-NF-κB p65 (1:1,000) and mouse anti-NF-κB p65 (1:1,000), mouse anti-phospho-p38 mitogen-activated protein kinase (MAPK) (1:1,000), mouse anti-p38 MAPK (1:1,000), mouse anti-phospho-ERK1/2 (1:1,000), mouse anti-ERK1/2 (1:1,000), mouseanti-phospho-JNK (1:1,000), mouse anti-JNK (1:1,000) (all purchased from Cell Signaling Technology (Danvers, MA, USA).
ROS production assay
ROS production was determined by using 5,6-chloromethyl-2’,7’-dichlorodihydrofluorescein diacetate (CMH2-DCFDA) (Sigma). HUVECs (3×106 cells/ml) were pretreated with various concentrations of eupatilin for 2 h, followed by addition of TNF-α (10 ng/ml) for 4 h. Then, 5 μMCMH2-DCFDA was added to cells and incubated at 37°C for 30 minutes in the dark. DCF fluorescence was determined using a Multi-detection microplate reader (BioTeke, Beijing, China) with an excitation wavelength of 485 nm and an emission wavelength of 530 nm.
Statistical analysis
Data were expressed as the mean ± standard deviation (SD). The statistical significance was determined by using the Student’s t-test for differences between two groups and one-way ANOVA for differences among multiple groups. P-values less than 0.05 were considered to be a statistically significant difference.
Results
Eupatilinattenuates TNF-α-activated HUVEC-monocyte interaction
Monocyte adhesion to endothelial cells is an essential event in the initiation of atherosclerosis development. To explore the effect of eupatilin on TNF-α-induced monocyte adhesion to HUVECs, we adopted a cell adhesion assay. As indicated in Figure 1, TNF-α treatment significantly increased the ability of monocytes to adhere to HUVECs, while treatment with eupatilin reduced U937 cells adhesion to TNF-α-stimulated HUVECs, exhibiting a dose-dependent manner.
Figure 1.
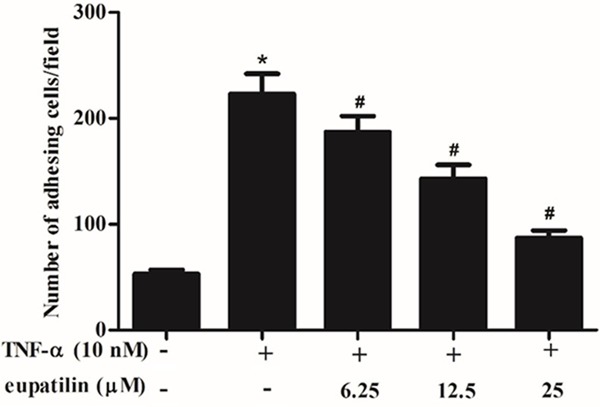
Eupatilin inhibited U937 cells adhesion to TNF-α-induced HUVEC. HUVEC were pre-incubated with various concentrations (6.25, 12.5 and 25 μM) of eupatilin for 2 h and stimulated with or without TNF-α for 4 h. U937 cells were layered over HUVEC monolayers and incubated for 2 h at 37°C. Adherent cells were photographed and analyzed by microscopy. The adherent cells were counted under a light microscope. At least three independent experiments were performed. The values shown represent the mean ± SD, the symbol * indicates differences from control group at P < 0.05; the symbol # indicates differences from TNF-α group at P < 0.05.
Eupatilin attenuated TNF-α-induced the expression of inflammatory cytokines in HUVECs
VCAM-1 and ICAM-1 are key adhesion molecules mediating leukocyte recruitment to early lesions in inflammatory vascular disease including atherosclerosis [2]. As shown in Figure 2A and 2B, TNF-α treatment resulted in a significant increase in the mRNA expression levels of VCAM-1 and ICAM-1 compared with the control group. However, eupatlin significantly inhibited TNF-α-induced the mRNA expression levels of VCAM-1 and ICAM-1 in a concentration-dependent manner. Consistent with the results of RT-PCR, Western blot analysis demonstrated that eupatilin also drastically inhibited TNF-α-induced the protein expression levels of VCAM-1 and ICAM-1 in a concentration-dependent manner (Figure 2C).
Figure 2.
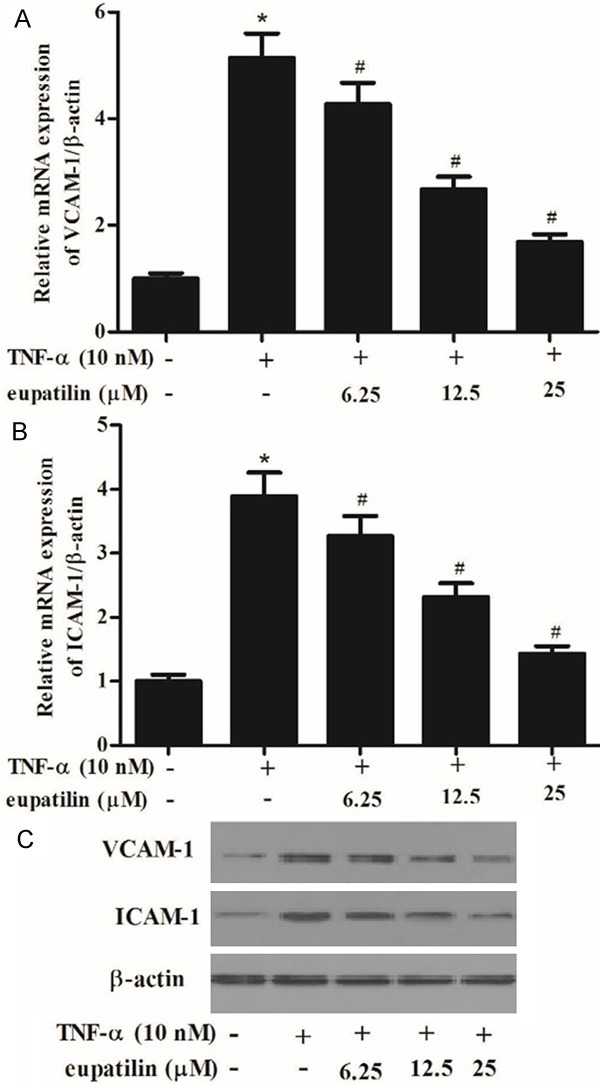
Eupatilin attenuated TNF-α-induced the expression of inflammatory cytokines in HUVECs.HUVEC were pre-incubated with various concentrations (6.25, 12.5 and 25 μM) of eupatilin for 2 h and stimulated with or without TNF-α for 4 h. A, B. VCAM-1 and ICAM-1 mRNA expression was determined by qRT-PCR. C. VCAM-1 and ICAM-1 protein expression was determined by Western blot. At least three independent experiments were performed. The values shown represent the mean ± SD, the symbol * indicates differences from control group at P < 0.05; the symbol # indicates differences from TNF-α group at P < 0.05.
Eupatilin attenuated TNF-α-induced intracellular ROS production in HUVECs
It has been reported that TNF-αinduces oxidative stress in cells and increase intracellular reactive oxygen species (ROS) generation.Thus, we investigated the effect of eupatilin on the production of TNF-α-induced ROS in HUVECs. As shown in Figure 3, TNF-α obviously increased the level of intracellular ROS, as compared with untreated cells. However, pretreatment with eupatilin significantly decreased TNF-α-induced ROS levels in a concentration dependent manner.
Figure 3.
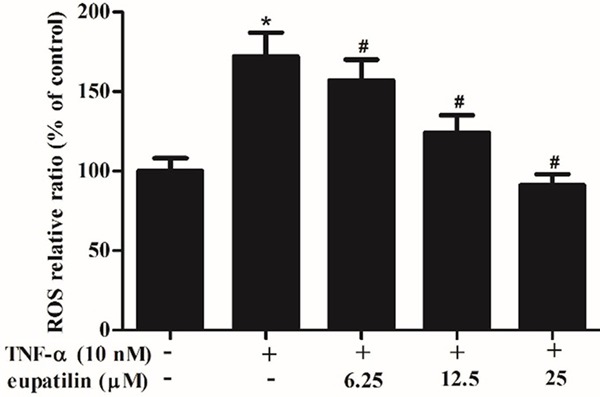
Eupatilin attenuated TNF-α-induced intracellular ROS production in HUVECs. HUVEC were pre-incubated with various concentrations (6.25, 12.5 and 25 μM) of eupatilin for 2 h and stimulated with or without TNF-α for 4 h. ROS level was analyzed by fluorescence intensity values. At least three independent experiments were performed. The values shown represent the mean ± SD, the symbol *indicates differences from control group at P < 0.05; the symbol #indicates differences from TNF-α group at P < 0.05.
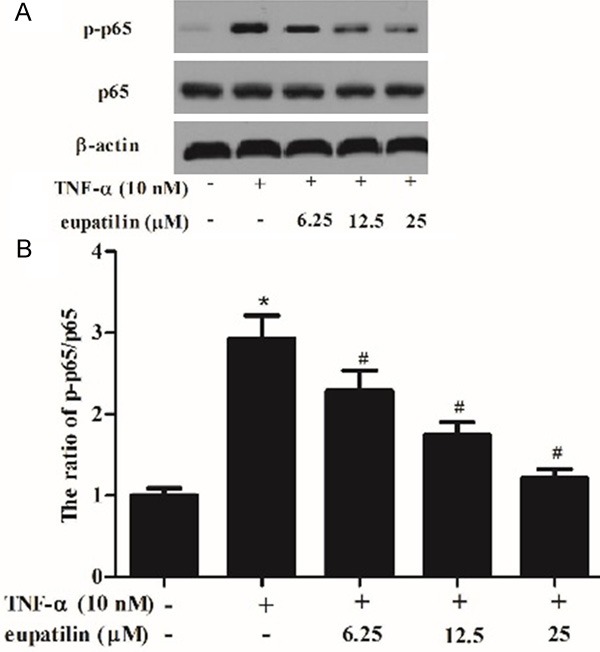
Eupatilin suppressed the TNF-α-activated NF-κB p65 phosphorylation in HUVECs
Activation of the NF-κBis involved in the expression of inflammatory mediators. Therefore, we investigated the effect of eupatilin on TNF-α-induced phosphorylation of NF-κB p65. As indicated in Figure 4, TNF-α significantly induced phosphorylation of p65, whereas, eupatilin prevented the TNF-α-induced phosphorylation of p65 in HUVECs. These results suggest that eupatilin inhibits TNF-α-induced NF-κB activation.
Figure 4.
Eupatilin suppressed the TNF-α-activated NF-κB p65 phosphorylation in HUVECs. HUVEC were pre-incubated with various concentrations (6.25, 12.5 and 25 μM) of eupatilin for 2 h and stimulated with or without TNF-α for 30 min. A. Protein levels of p65 were detected by Western blotting. B and C. Protein expression was analyzed using BandScan 5.0 software and normalized to β-actin. At least three independent experiments were performed. The values shown represent the mean ± SD, the symbol *indicates differences from control group at P < 0.05; the symbol #indicates differences from TNF-α group at P < 0.05.
Eupatilin suppressed the TNF-α-activated MAPK phosphorylation in HUVECs
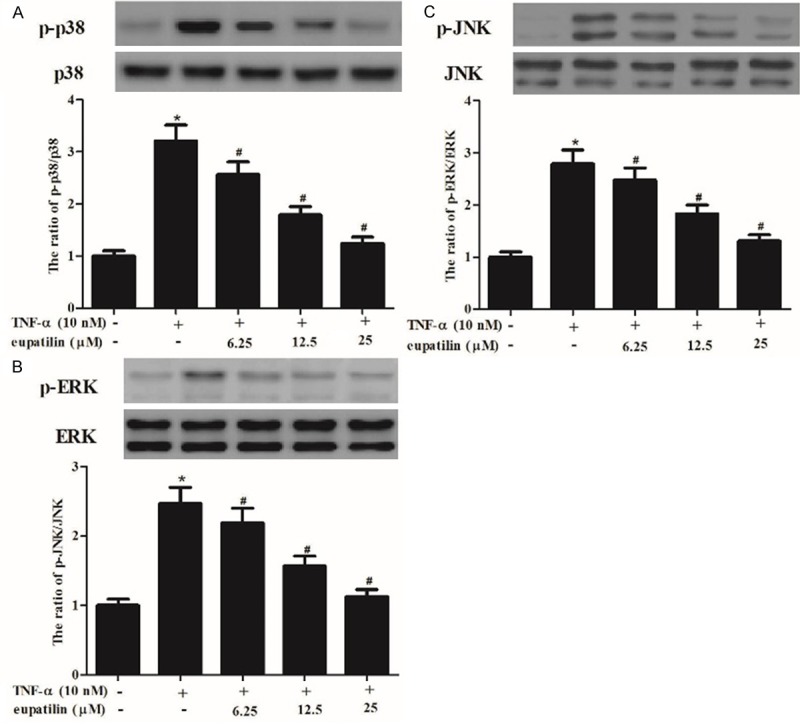
The MAP kinase (MAPK) pathways are involved in controlling the expression of the pro-inflammatory mediators in endothelial cells. Therefore, we investigated the effect of eupatilin on TNF-α-induced phosphorylation of MAPKs in HUVECs. As indicated in Figure 5, when HUVECs were treated with TNF-α alone at 30 min, the levels of phosphorylation of p38 MAPK, ERK1/2 and JNK wereincreased compared with untreated control. However, pretreatment with eupatilin inhibited the TNF-α induced phosphorylation of MAPKs in HUVECs.
Figure 5.
Eupatilin suppressed the TNF-α-activated phosphorylation of MAPKs in HUVECs. HUVEC were pre-incubated with various concentrations (6.25, 12.5 and 25 μM) of eupatilin for 2 h and stimulated with or without TNF-α for 30 min. A-C. Protein expression of p-p38, p38, p-ERK, ERK, p-JNK and JNK was analyzed by western blot analysis, and protein expression was analyzed using BandScan 5.0 software and normalized to β-actin.At least three independent experiments were performed. The values shown represent the mean ± SD, the symbol *indicates differences from control group at P < 0.05; the symbol #indicates differences from TNF-α group at P < 0.05.
Discussion
Atherosclerosis is a cellular response to a continuous insult of vascular inflammation resulting in successive intimal thickening. In the present study, our findings showed that eupatilin reduced U937 cells adhesion to TNF-α-stimulated HUVECs and attenuated TNF-α-induced the expression of inflammatory cytokines in HUVECs, as well as the production of intracellular ROS. Moreover, eupatilininhibits TNF-α-induced phosphorylation of MAPKs and NF-κB p65 in HUVECs.
Monocyte-HUVECs adhesion is an early step in inflammatory disorders such as atherosclerosis [13]. Thus, reducing monocyte adhesion to HUVECs is a promising pharmacological target for the prevention of atherosclerosis. In this study, we found that eupatlinsuppressed the ability of monocytes to adhere to HUVECs induced by TNF-α. In corroboration of these results, eupatilin significantly inhibited induction of expression of adhesion molecules in a concentration-dependent manner after stimulation with the inflammatory cytokine, TNF-α, at the level of protein and mRNA. These results suggested that eupatilin suppressed TNF-α-induced monocyte/endothelial cell interactions by downregulating the expression of the adhesion molecules ICAM-1 and VCAM-1.
Several reports have shown that the oxidative stress was correlated with pathophysiological damage in endothelial dysfunction [14-16]. Moreover, previous studies have shown that cytokines such as TNF-α could induce the production of intercellular ROS in endothelial cells [17,18]. Consistent with the above-mentioned studies, in this study, we observed that eupatilin inhibited TNF-α-induced ROS generation in HUVECs. These results suggested that free radical scavenging effects of eupatlin may contribute to its anti-inflammatory effects.
Nuclear factor-kappa B (NF-κB) is a transcription factor which has been considered to be the critical regulator of the inflammatory process [19,20]. It is well known that NF-κB is involved in the regulation of VCAM-1, ICAM-1 and E-selectin expression and cytokine production [21,22]. Moreover, it has been reported that TNF-α induces oxidative stress in endothelial cells and increase intracellular ROS generation, leads to the activation of NF-κB [23]. In line with these results, in this study, we found thateupatilin inhibited the TNF-α induced phosphorylation of NF-ĸB p65 in HUVECs. These results suggest that eupatilin attenuated the TNF-α-induced inflammatory mediator expression through inhibition of NF-ĸB activation in HUVECs.
MAP kinases are a family of serine-threonine kinases, participating in the regulation of cell adhesion molecules expressed on cells in response to external stimuli including TNF-α [24,25]. Furthermore, there is growing evidence shows that MAPK signaling pathways are involved in the regulation of NF-ĸB activation in TNF-α-induced endothelial cells [26-28]. In this study, we found that eupatilin inhibited the TNF-α induced phosphorylation of MAPKs in HUVECs. These results suggest that eupatilin inhibited TNF-α-induced NF-ĸB activation and adhesion molecules expression involved of inhibiting MAPK signaling pathway.
In summary, the results of the present study suggest that eupatilininhibited inflammatory reaction through suppressing the ROS/MAPK-NF-ĸB pathway in HUVECs. Thus, eupatilin is proposed as an effective new anti-inflammatory agent to suppress vascular inflammation, and further prevent atherosclerosis.
Disclosure of conflict of interest
None.
References
- 1.Glass CK, Witztum JL. Atherosclerosis: the road ahead. Cell. 2001;104:503–516. doi: 10.1016/s0092-8674(01)00238-0. [DOI] [PubMed] [Google Scholar]
- 2.Blankenberg S, Barbaux S, Tiret L. Adhesion molecules and atherosclerosis. Atherosclerosis. 2003;170:191–203. doi: 10.1016/s0021-9150(03)00097-2. [DOI] [PubMed] [Google Scholar]
- 3.Basuroy S, Bhattacharya S, Leffler CW, Parfenova H. Nox4 NADPH oxidase mediates oxidative stress and apoptosis caused by TNF-α in cerebral vascular endothelial cells. Am J Physiol Cell Physiol. 2009;296:C422–C432. doi: 10.1152/ajpcell.00381.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Basuroy S, Bhattacharya S, Tcheranova D, Qu Y, Regan RF, Leffler CW, Parfenova H. HO-2 provides endogenous protection against oxidative stress and apoptosis caused by TNF-α in cerebral vascular endothelial cells. Am J Physiol Cell Physiol. 2006;291:C897–C908. doi: 10.1152/ajpcell.00032.2006. [DOI] [PubMed] [Google Scholar]
- 5.Zahler S, Kupatt C, Becker BF. Endothelial preconditioning by transient oxidative stress reduces inflammatory responses of cultured endothelial cells to TNF-α. FASEB J. 2000;14:555–564. doi: 10.1096/fasebj.14.3.555. [DOI] [PubMed] [Google Scholar]
- 6.Jeong JH, Moon SJ, Jhun JY, Yang EJ, Cho ML, Min JK. Eupatilin Exerts Antinociceptive and Chondroprotective Properties in a Rat Model of Osteoarthritis by Downregulating Oxidative Damage and Catabolic Activity in Chondrocytes. PLoS One. 2015;10:e0130882. doi: 10.1371/journal.pone.0130882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Jung J, Ko SH, Yoo DY, Lee JY, Kim YJ, Choi SM, Kang KK, Yoon HJ, Kim H, Youn J. 5, 7-Dihydroxy-3, 4, 6-trimethoxyflavone inhibits intercellular adhesion molecule 1 and vascular cell adhesion molecule 1 via the Akt and nuclear factor-κB-dependent pathway, leading to suppression of adhesion of monocytes and eosinophils to bronchial epithelial cells. Immunology. 2012;137:98–113. doi: 10.1111/j.1365-2567.2012.03618.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Park BB, sun Yoon J, shil Kim E, Choi J, woong Won Y, hye Choi J, Lee YY. Inhibitory effects of eupatilin on tumor invasion of human gastric cancer MKN-1 cells. Tumour Biol. 2013;34:875–885. doi: 10.1007/s13277-012-0621-y. [DOI] [PubMed] [Google Scholar]
- 9.Min SW, Kim NJ, Baek NI, Kim DH. Inhibitory effect of eupatilin and jaceosidin isolated from Artemisia princeps on carrageenan-induced inflammation in mice. J Ethnopharmacol. 2009;125:497–500. doi: 10.1016/j.jep.2009.06.001. [DOI] [PubMed] [Google Scholar]
- 10.Choi EJ, Lee S, Chae JR, Lee HS, Jun CD, Kim SH. Eupatilin inhibits lipopolysaccharideinduced expression of inflammatory mediators in macrophages. Life Sci. 2011;88:1121–1126. doi: 10.1016/j.lfs.2011.04.011. [DOI] [PubMed] [Google Scholar]
- 11.Son JE, Lee E, Seo SG, Lee J, Kim JE, Kim J, Lee KW, Lee HJ. Eupatilin, a major flavonoid of Artemisia, attenuates aortic smooth muscle cell proliferation and migration by inhibiting PI3K, MKK3/6, and MKK4 activities. Planta Med. 2013;79:1009–1016. doi: 10.1055/s-0033-1350621. [DOI] [PubMed] [Google Scholar]
- 12.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2- ΔΔCT method. Methods. 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 13.Moon MK, Lee YJ, Kim JS, Kang DG, Lee HS. Effect of caffeic acid on tumor necrosis factor-alpha-induced vascular inflammation in human umbilical vein endothelial cells. Biol Pharm Bull. 2009;32:1371–1377. doi: 10.1248/bpb.32.1371. [DOI] [PubMed] [Google Scholar]
- 14.Yachie A, Niida Y, Wada T, Igarashi N, Kaneda H, Toma T, Ohta K, Kasahara Y, Koizumi S. Oxidative stress causes enhanced endothelial cell injury in human heme oxygenase-1 deficiency. J Clin Invest. 1999;103:129–135. doi: 10.1172/JCI4165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Cai H, Harrison DG. Endothelial dysfunction in cardiovascular diseases: the role of oxidant stress. Circ Res. 2000;87:840–844. doi: 10.1161/01.res.87.10.840. [DOI] [PubMed] [Google Scholar]
- 16.Schulz E, Jansen T, Wenzel P, Daiber A, Münzel T. Nitric oxide, tetrahydrobiopterin, oxidative stress, and endothelial dysfunction in hypertension. Antioxid Redox Signal. 2008;10:1115–1126. doi: 10.1089/ars.2007.1989. [DOI] [PubMed] [Google Scholar]
- 17.Chen JW, Chen YH, Lin FY, Chen YL, Lin SJ. Ginkgo biloba extract inhibits tumor necrosis factor-α-induced reactive oxygen species generation, transcription factor activation, and cell adhesion molecule expression in human aortic endothelial cells. Arterioscler Thromb Vasc Biol. 2003;23:1559–1566. doi: 10.1161/01.ATV.0000089012.73180.63. [DOI] [PubMed] [Google Scholar]
- 18.Corda S, Laplace C, Vicaut E, Duranteau J. Rapid reactive oxygen species production by mitochondria in endothelial cells exposed to tumor necrosis factor-α is mediated by ceramide. Am J Respir Cell Mol Biol. 2001;24:762–768. doi: 10.1165/ajrcmb.24.6.4228. [DOI] [PubMed] [Google Scholar]
- 19.Javaid K, Rahman A, Anwar KN, Frey RS, Minshall RD, Malik AB. Tumor necrosis factor-α induces early-onset endothelial adhesivity by protein kinase Cζ–dependent activation of intercellular adhesion molecule-1. Circ Res. 2003;92:1089–1097. doi: 10.1161/01.RES.0000072971.88704.CB. [DOI] [PubMed] [Google Scholar]
- 20.Lockyer JM, Colladay JS, Alperin-Lea WL, Hammond T, Buda AJ. Inhibition of nuclear factor-κB-mediated adhesion molecule expression in human endothelial cells. Circ Res. 1998;82:314–320. doi: 10.1161/01.res.82.3.314. [DOI] [PubMed] [Google Scholar]
- 21.Wan M, Liu J, Ouyang X. Nucleotide‐binding oligomerization domain 1 regulates Porphyromonas gingivalis‐induced vascular cell adhesion molecule 1 and intercellular adhesion molecule 1 expression in endothelial cells through NF‐κB pathway. J Periodontal Res. 2015;50:189–196. doi: 10.1111/jre.12192. [DOI] [PubMed] [Google Scholar]
- 22.Wang L, Xu Y, Yu Q, Sun Q, Xu Y, Gu Q, Xu X. H-RN, a novel antiangiogenic peptide derived from hepatocyte growth factor inhibits inflammation in vitro and in vivo through PI3K/AKT/IKK/NF-κB signal pathway. Biochem Pharmacol. 2014;89:255–265. doi: 10.1016/j.bcp.2014.02.026. [DOI] [PubMed] [Google Scholar]
- 23.Liu X, Pan L, Wang X, Gong Q, Zhu YZ. Leonurine protects against tumor necrosis factor-α-mediated inflammation in human umbilical vein endothelial cells. Atherosclerosis. 2012;222:34–42. doi: 10.1016/j.atherosclerosis.2011.04.027. [DOI] [PubMed] [Google Scholar]
- 24.Ju JW, Kim SJ, Jun CD, Chun JS. p38 Kinase and c‐Jun N‐Terminal Kinase Oppositely Regulates Tumor Necrosis Factor α‐Induced Vascular Cell Adhesion Molecule‐1 Expression and Cell Adhesion in Chondrosarcoma Cells. IUBMB Life. 2002;54:293–299. doi: 10.1080/15216540215674. [DOI] [PubMed] [Google Scholar]
- 25.Ho AW, Wong CK, Lam CW. Tumor necrosis factor-α up-regulates the expression of CCL2 and adhesion molecules of human proximal tubular epithelial cells through MAPK signaling pathways. Immunobiology. 2008;213:533–544. doi: 10.1016/j.imbio.2008.01.003. [DOI] [PubMed] [Google Scholar]
- 26.Kanaji N, Nelson A, Wang X, Sato T, Nakanishi M, Gunji Y, Basma H, Michalski J, Farid M, Rennard SI. Differential roles of JNK, ERK1/2, and p38 mitogen-activated protein kinases on endothelial cell tissue repair functions in response to tumor necrosis factor-α. J Vasc Res. 2013;50:145–156. doi: 10.1159/000345525. [DOI] [PubMed] [Google Scholar]
- 27.Hu W, Zhang Q, Yang X, Wang Y, Sun L. Puerarin Inhibits Adhesion Molecule Expression in TNF-α-Stimulated Human Endothelial Cells via Modulation of the Nuclear Factor ĸB Pathway. Pharmacology. 2010;85:27–35. doi: 10.1159/000264938. [DOI] [PubMed] [Google Scholar]
- 28.Zhang L, Zhang S. Modulating Bcl‐2 Family Proteins and Caspase‐3 in Induction of Apoptosis by Paeoniflorin in Human Cervical Cancer Cells. Phytother Res. 2011;25:1551–1557. doi: 10.1002/ptr.3534. [DOI] [PubMed] [Google Scholar]