

Abstract
In vivo protection by antimicrobial neutralizing Abs can require the contribution of effector functions mediated by Fc-Fcγ receptor (Fc-FcγR) interactions for optimal efficacy. In influenza, broadly neutralizing anti-hemagglutinin (anti-HA) stalk mAbs require Fc-FcγR interactions to mediate in vivo protection, but strain-specific anti-HA head mAbs do not. Whether this rule applies only to anti-stalk Abs or is applicable to any broadly neutralizing Ab (bNAb) against influenza is unknown. Here, we characterized the contribution of Fc-FcγR interactions during in vivo protection for a panel of 13 anti-HA mAbs, including bNAbs and non-neutralizing Abs, against both the stalk and head domains. All classes of broadly binding anti-HA mAbs required Fc-FcγR interactions to provide protection in vivo, including those mAbs that bind the HA head and those that do not neutralize virus in vitro. Further, a broadly neutralizing anti-neuraminidase (anti-NA) mAb also required FcγRs to provide protection in vivo, but a strain-specific anti-NA mAb did not. Thus, these findings suggest that the breadth of reactivity of anti-influenza Abs, regardless of their epitope, necessitates interactions with FcγRs on effector cell populations to mediate in vivo protection. These findings will guide the design of antiviral Ab therapeutics and inform vaccine design to elicit Abs with optimal binding properties and effector functions.
Introduction
Influenza virus epidemics contribute to 250,000 to 500,000 deaths per year worldwide (1). Vaccination to elicit anti-influenza Abs and passive administration of Abs represent approaches for providing protection in humans (2, 3). Hemagglutinin (HA), the glycoprotein to which the majority of the influenza immune response is directed, is the main target of the post-vaccination immune response. Influenza HA is composed of two domains: the immunodominant globular head (HA1), which varies from strain to strain, and a stalk (HA2) that is relatively conserved between strains and subtypes (4). Thus, most Abs against the head react with a single or highly similar influenza strain, while Abs against the stalk domain often bind a breadth of strains and subtypes.
The mechanisms underlying Ab-mediated antiviral neutralization and protection are distinct. Ab-mediated neutralization is quantified during in vitro assays and results from steric blockade of epitopes required for viral entry (anti-HA head mAbs), fusion (anti-HA stalk mAbs), or budding and egress (anti-HA and anti-neuraminidase [anti-NA] mAbs) (5). By contrast, Ab-mediated protection is measured in vivo and results from a complex interplay between immune effector cells and effector mechanisms mediated by these cells, in addition to the above-mentioned in vitro–defined neutralization mechanisms (6). Thus, recent studies have identified that interactions between the Ab’s constant Fc domain and members of the Fcγ receptor (FcγR) family are required for anti-influenza mAb–mediated protection in vivo (7, 8). FcγRs are an important family of cell-surface receptors expressed by immune cells that couple innate and adaptive immunity after Ab engagement. Both activating FcγRs (murine FcγRI, FcγRIII, and FcγRIV) and an inhibitory FcγR (FcγRIIb) are coexpressed by most immune cells, and the overall balance of signaling between these two classes determines the biological effect of circulating immune complexes or Abs bound to pathogens or cells (9).
Previous studies demonstrated that broadly neutralizing anti-HA stalk mAbs required Fc-FcγR interactions to mediate protection from lethal influenza infection in vivo, while strain-specific anti-HA head mAbs did not (7). Mechanistically, it was demonstrated that anti-HA stalk mAbs efficiently engage FcγRs to trigger Ab-dependent cellular cytotoxicity (ADCC), while anti-head mAbs do not. Whether this general rule applies to all anti-HA head and stalk mAbs is unknown. Here, we analyze a panel of 13 anti-HA mAbs, including broadly neutralizing Abs (bNAbs) and non-neutralizing Abs against both the stalk and head domains. We demonstrate that each of the broadly binding and neutralizing mAbs requires Fc-FcγR interactions to mediate protection in vivo, suggesting that the breadth of reactivity of anti-influenza Abs, regardless of their epitope, necessitates interactions with FcγRs on effector cell populations for in vivo protection.
Results and Discussion
Anti-HA Ab panel.
We have collected a panel of 13 anti-HA mAbs, including bNAbs and non-neutralizing Abs against both the stalk and head domains (Table 1 and Supplemental Tables 1–4 display virus strains recognized and neutralized by each clone; supplemental material available online with this article; doi:10.1172/JCI84428DS1). Five mAbs — FI6, 6F12, 2B06, 2G02, and 1F02 — are broadly neutralizing and bind to the HA stalk. 7B2, PY102, and 4C04 are strain-specific mAbs that engage the HA head. Finally, 4G05 and 1F05 are broadly neutralizing anti-HA head mAbs, while 1A01, 1A05, and 4G01 are pan-H1, but non-neutralizing, anti-HA head mAbs. All mAbs bound cell-surface HA on A549 cells infected with the 2009 pandemic H1N1 strain A/Netherlands/602/2009 (Neth09) (Figure 1A and Supplemental Figure 1B). Each mAb also bound to purified 2009 pandemic A/California/04/2009 (Cal09) virus, Cal09 HA protein, and the 2014–2015 trivalent influenza vaccine (Supplemental Figure 1, A, C, and D). All anti-head mAbs bound to recombinant Cal09 HA1 (head) domain proteins (with the exception of the HAI+ mAb IF05, which only recognizes HA on whole virus or infected cells), while all anti-stalk bNAbs bound to a chimeric HA protein containing H5 head and H1 stalk domains (Supplemental Figure 1, E and F). As previously described (7), strain-specific anti-HA head mAbs showed the most potent in vitro neutralization during in vitro plaque reduction neutralization assays, while broadly neutralizing anti-HA stalk mAbs showed weaker neutralization capabilities in vitro (Figure 1B). Broadly neutralizing anti-HA head mAbs showed intermediate neutralization Abs, while the pan-H1, but non-neutralizing, mAbs showed no activity. Within each subset of Abs, each clone was from a different patient and used distinct VH genes (except for two anti-stalk bNAbs from different individuals that utilized the same VH). Heavy-chain variable, joining, and diversity (VDJ) junction analysis demonstrated that each clone was unique in CDR3 length and amino acid sequence. Thus, each subset of anti-HA Abs was composed of independent Ab clones derived from unique individuals and unique selection events.
Table 1. mAb characteristics.
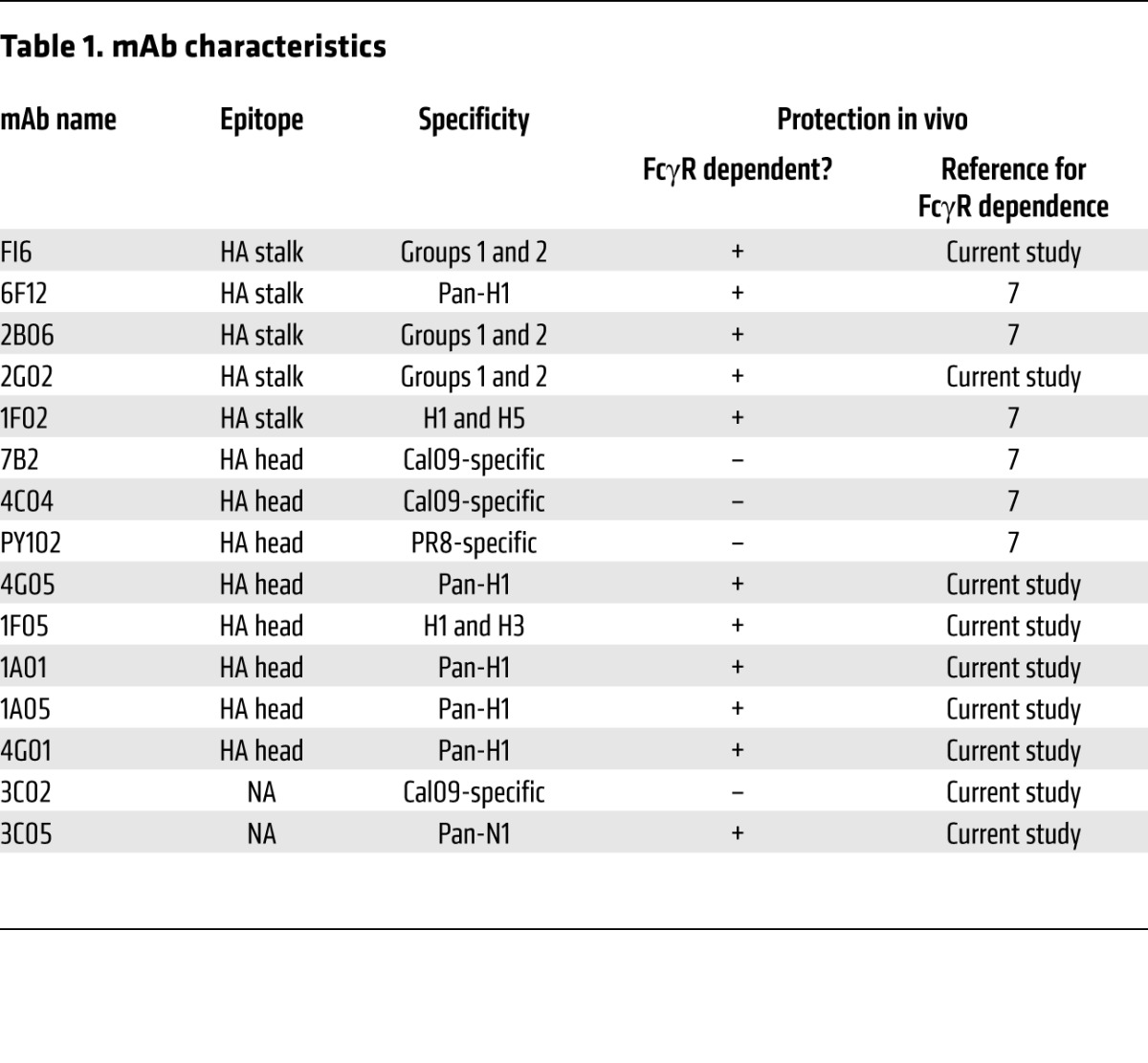
Figure 1. Broadly binding and neutralizing anti-HA stalk mAbs require Fc-FcγR interactions for protection in vivo.

(A) Broadly neutralizing anti-stalk (red lines), strain-specific anti-head (blue lines), broadly neutralizing anti-head (green lines), and non-neutralizing anti-head (purple lines) mAb binding to Neth09-infected cells by flow cytometric analysis and (B) neutralization of Neth09 virus (mean ± SEM) in duplicate samples. (C and D) WT mice were given the indicated doses of IgG2a (red circles) or DA265-mutant (blue squares) FI6 mAb (C), 2G02 mAb (D), or PBS (black triangles) before Neth09 virus infection. Values represent the mean ± SEM percentage of weight change compared with day-0 values (left panels) and percentage of survival (right panels). n = 4–5 mice per group. Significant differences between the IgG2a and DA265 samples are shown. *P < 0.05 and **P < 0.01, by 2-tailed Student’s t test.
Broadly neutralizing anti-HA stalk mAbs require Fc-FcγR interactions to mediate protection in vivo.
To determine the contributions of Fc-FcγR interactions during influenza virus neutralization in vivo, we generated mAb constructs with different Fc domains that either preferentially engaged activating FcγRs (murine IgG2a) or were null for FcγR binding (DA265-mutant Fc) (10). We compared IgG2a and DA265 constructs for H1 protein binding (Supplemental Figure 2) and detected no differences. Modification of the Fc domain by introducing point mutations or switching isotypes does not affect Fab-mediated activities, including binding or neutralization (7).
To test whether Fc-FcγR interactions contributed to bNAb-mediated protection in vivo, we treated WT mice that had received 6 mg/kg body weight FI6 or 2G02 anti-stalk bNAbs with five 50% mouse lethal doses (mLD50) of Neth09 and monitored weight loss and survival. Mice that received the IgG2a FI6 and 2G02 bNAbs showed minimal weight loss compared with that seen in PBS-treated mice (P < 0.0001) on day 7, whereas DA265-treated mice showed weight loss curves similar to those of the PBS-treated animals (Figure 1, C and D). FI6 and 2G02 IgG2a bNAb–treated mice showed 100% survival, whereas no animals in the DA265- or PBS-treated groups survived (P = 0.005 and P = 0.002, respectively). Similarly, Fcer1g–/– mice (11), which lack the FcR-γ chain and do not express activating FcγRs (FcγRI, FcγRIII, or FcγRIV), were not protected by administration of IgG2a isotype 2G02 mAb (Supplemental Figure 3). Lung viral titers correlate with weight loss and survival in animals treated with Abs that mediate protection in an Fc-dependent manner (7). We used PBS as a control treatment in all experiments, because no differences in weight or survival curves are observed in mice receiving PBS or isotype control mAb (7). Ab isotype–specific differences in in vivo half-lives are similar between IgG2a and DA265 mutant Abs (7, 12) and therefore do not influence these results. Thus, Fc-FcγR interactions are required for broadly neutralizing anti-HA stalk mAb–mediated protection during in vivo viral challenge.
Broadly neutralizing anti-HA head mAbs require Fc-FcγR interactions to mediate protection in vivo.
All strain-specific anti-HA head mAbs that have been tested thus far do not require Fc-FcγR interactions for protection in vivo (Table 1 and ref. 7). However, whether broadly neutralizing anti-head mAbs mediate protection in vivo through Fc-mediated effector mechanisms remains unknown. Therefore, we tested whether Fc-FcγR interactions mediate protection by two broadly neutralizing anti-head mAbs. Mice receiving IgG2a 4G05 and 1F05 anti-head bNAbs showed minimal weight loss after lethal Neth09 challenge compared with mice receiving the DA265 bNAbs (P < 0.0012 on day 7; Figure 2, A and B). 4G05 and 1F05 IgG2a bNAb–treated mice had 100% survival, whereas no animals in the DA265- or PBS-treated groups survived (P ≤ 0.002). In addition, FcRα-null mice, which lack all activating and inhibitory FcγRs (FcγRI, FcγRIIb, FcγRIII, or FcγRIV) (13), were not protected by administration of IgG2a isotype 4G05 or 1F05 mAb (Supplemental Figure 4, A and B). Thus, while strain-specific anti-HA head mAbs do not require Fc-FcγR interactions to mediate protection in vivo, broadly neutralizing anti-HA head mAbs are protective in vivo in an FcγR-dependent manner.
Figure 2. Broadly neutralizing anti-HA head mAbs and pan-H1, non-neutralizing anti-HA head mAbs require Fc-FcγR interactions for protection in vivo.

WT mice were given the indicated doses of IgG2a (red circles) or DA265-mutant (blue squares) 4G05 (A), 1F05 (B), 1A01 (C), 1A05 (D), or 4G01 (E) mAb or PBS (black triangles) before Neth09 viral infection. Values represent the mean ± SEM percentage of weight change compared with day-0 values (left panels) and percentage of survival (right panels). n = 5–7 mice per group. Significant differences between the IgG2a and DA265 samples are shown. *P < 0.05 and **P < 0.01, by 2-tailed Student’s t test.
Pan-H1, non-neutralizing anti-HA head mAbs require Fc-FcγR interactions to mediate protection in vivo.
We next tested whether pan-H1, but non-neutralizing, anti-HA head mAbs are protective in vivo and whether this protection is Fc dependent. Although IgG2a isotype 1A01, 1A05, and 4G01 mAbs were unable to neutralize Neth09 virus during in vitro assays (Figure 1B), all three mAbs protected mice from lethal Neth09 challenge in vivo (Figure 2, C–E) at doses ranging from 4 mg/kg to 6 mg/kg. Surprisingly, these doses are similar to those necessary for in vivo protection by broadly neutralizing anti-HA stalk mAbs (Figure 1, C and D, ref. 7), as well as by the broadly neutralizing anti-HA head mAb 1F05 (Figure 2B). By contrast, similar doses of DA265-mutant versions of 1A01, 1A05, and 4G01 mAbs were unable to prevent weight loss after lethal challenge (P ≤ 0.0001 at day 7). Accordingly, IgG2a non-neutralizing mAb–treated mice had 100% survival, whereas no animals in the DA265- or PBS-treated groups survived (P = 0.0009, P = 0.0034, and P = 0.0009, respectively). Fcer1g–/– or FcRα-null mice were not protected by IgG2a isotype 1A01, 1A05, or 4G01 mAbs (Supplemental Figure 5, A–C). Thus, while strain-specific anti-HA head mAbs do not require Fc-FcγR interactions to mediate protection in vivo (7), broad-binding mAbs reactive with the HA head protect in vivo in an FcγR-dependent manner, regardless of whether the mAbs neutralize virus in vitro. Collectively, these results (Figures 1 and 2) demonstrate that anti-HA head or stalk Abs that recognize a breadth of influenza HA subtypes require interactions with FcγRs to mediate protection in vivo. These findings suggest that it is the breadth of neutralizing Abs against HA that necessitates Fc-FcγR interactions to mediate protection during influenza infection in vivo.
Broadly neutralizing anti-NA mAb requires Fc-FcγR interactions to mediate protection, but strain-specific anti-NA mAb does not.
Although we have demonstrated that the breadth of neutralizing Abs against HA necessitates FcγR-mediated effector functions for protection in vivo, it is unknown whether this is a generalizable property of broadly reactive Abs. Therefore, we tested two anti-NA mAbs during in vitro neutralization assays and in vivo protection experiments. 3C02 is a strain-specific anti-NA mAb that recognizes N1 from the 2009 pandemic H1N1 strain (Table 1). By contrast, anti-NA mAb 3C05 is a broadly neutralizing anti-NA mAb that reacts with multiple N1 strains (Table 1). Both Abs neutralized Neth09 virus in vitro, but strain-specific 3C02 did so with greater than 2 logs more potency than did broadly neutralizing 3C05 (Figure 3A). In vivo, both IgG2a and DA265-mutant 3C02 mAbs provided similar protection at the suboptimal dose of 0.5 mg/kg after Neth09 challenge, with no statistically significant differences detected in weight loss on day 7 or in overall survival (Figure 3B). We noted no differences in weight loss or survival between mice receiving IgG2a or DA265 3C02 mAbs at any dose between 0.25 mg/kg and 6 mg/kg (Supplemental Figure 6). By contrast, an IgG2a isotype version of the broadly neutralizing anti-NA mAb 3C05 provided full weight loss protection at day 7 (P = 0.0005) and 100% survival, while no mice receiving DA265-mutant 3C05 mAb survived the challenge (P = 0.004). Thus, as seen with anti-HA mAbs, an anti-NA mAb that recognizes a breadth of influenza viruses requires Fc-FcγR interactions to mediate protection from lethal infection in vivo, while a strain-specific anti-NA mAb does not.
Figure 3. Broadly neutralizing anti-NA mAb requires Fc-FcγR interactions to mediate protection in vivo, but strain-specific anti-NA mAb does not.

(A) Broadly neutralizing anti-HA stalk mAb FI6 (red circles), strain-specific anti-HA head mAb 4C04 (blue squares), strain-specific anti-NA mAb 3C02 (green circles), and broadly neutralizing anti-NA mAb 3C05 (green squares) neutralization of Neth09 virus. Values represent the mean ±SEM of duplicate samples. (B and C) Mice were given the indicated doses of IgG2a (red circles) or DA265-mutant (blue squares) 3C02 (B) or 3C05 (C) mAb or PBS (black triangles) before Neth09 viral infection. Values represent the mean ± SEM percentage of weight change compared with day-0 values (left panels) and percentage of survival (right panels). n = 4–5 mice per group. Significant differences between the IgG2a sample and DA265 sample are shown. **P < 0.01 by Student’s t test.
In these studies, all neutralizing and non-neutralizing anti-HA and anti-NA Abs that recognize a breadth of influenza strains and/or subtypes required FcγRs for protection in vivo, while strain-specific mAbs did not. While previous studies assessing strain-specific anti-HA head mAbs showed that this class of mAb did not require FcγRs for protection in vivo (7), it is now clear that at least some broad-binding Abs against the head domain require Fc-FcγR interactions to mediate protection in vivo. Additional studies will be required to generalize this requirement. Further, we demonstrate that a broadly neutralizing anti-NA mAb required Fc-FcγR interactions for protection, while a strain-specific anti-NA mAb did not. Therefore, the current studies strongly suggest that the breadth of reactivity of anti-influenza mAbs necessitates FcγR-mediated effector functions for protection in vivo.
Surprisingly, both broadly neutralizing anti-HA stalk and non-neutralizing Abs mediated protection in vivo in an FcγR-dependent manner at similar doses. These findings suggest that the in vitro neutralization mechanisms of broadly neutralizing mAbs, such as inhibition of viral fusion or egress, do not dominate in vivo at the doses tested. Therefore, our findings suggest that non-neutralizing Abs targeting other influenza-derived antigens expressed on the surface of virally infected cells may mediate protection in vivo through FcγRs and warrant further investigation. The mechanism of protection by all classes of broadly neutralizing anti-influenza Abs is likely cytotoxicity and phagocytosis of infected cells, since FcγRs contribute to protection after viral entry into target cells (7) and clodronate liposome–sensitive cell populations mediate protection by anti-influenza Abs (14).
Importantly, for Abs with in vitro–defined neutralizing capabilities (such as inhibition of virus binding, viral fusion, or egress), dramatically increasing the dose of mAb (8- to 10-fold) bypasses the FcγR requirement through effective utilization of the in vitro–defined neutralization mechanisms (7). In vivo, bNAbs are generated at much lower frequencies in the Ab repertoire compared with strain-specific mAbs (15). Thus, engagement of the FcγR system may represent an important mechanism to compensate for low concentrations of these mAbs in the serum. It is tempting to speculate that some selection event occurs during the Ab response to select for mAbs with appropriate effector functions; further studies will be required to address this. In addition, we demonstrate that non-neutralizing Abs likely play a crucial role during in vivo antiviral Ab responses by triggering FcγR-mediated effector mechanisms to destroy infected cells expressing viral antigens. Thus, the “polyclonality” of the Ab response, including neutralizing and non-neutralizing Abs with various degrees of effector functions, serves to provide optimal antiviral immunity in vivo.
Our findings demonstrate a general FcγR-mediated mechanism underlying the in vivo protective activities of anti-influenza Abs with a breadth of reactivity. Thus, ideal antiviral immunotherapies, such as passively administered bNAbs and vaccine-elicited bNAbs, must be optimized for FcγR-mediated effector functions.
Methods
Detailed methods are described in the Supplemental Methods.
Statistics.
Statistical differences between survival were analyzed by comparing Kaplan-Meier curves using the log-rank test and GraphPad Prism software, version 5.0 (GraphPad Software). All other statistical differences were compared using a 2-tailed Student’s t test. A P value of less than 0.05 was considered statistically significant.
Study approval.
All animal studies were approved by the IACUC of The Rockefeller University.
Author contributions
DJD, PCW, and JVR designed the experiments. DJD and PCW generated reagents. PP provided reagents and methodological advice. DJD performed the experiments. DJD and JVR analyzed the data. DJD and JVR wrote the manuscript.
Supplementary Material
Acknowledgments
We thank Florian Krammer (Icahn School of Medicine, New York, New York, USA) for providing HA protein and purified virus. This work was supported by the National Institute of Allergy and Infectious Diseases (NIAID), NIH (5U19AI109946, to J.V. Ravetch). The content is solely the responsibility of the authors and does not necessarily represent the official views of the NIH. D.J. DiLillo received a postdoctoral fellowship from the Leukemia and Lymphoma Society of America.
Footnotes
Conflict of interest: The authors have declared that no conflict of interest exists.
Reference information:J Clin Invest. 2016;126(2):605–610. doi:10.1172/JCI84428.
References
- 1.WHO Influenza (Seasonal) Fact sheet 211. [March 1, 2014]; [November 30, 2015];WHO Web site. http://www.who.int/mediacentre/factsheets/fs211/en/
- 2.Pica N, Palese P. Toward a universal influenza virus vaccine: prospects and challenges. Annu Rev Med. 2013;64:189–202. doi: 10.1146/annurev-med-120611-145115. [DOI] [PubMed] [Google Scholar]
- 3.Wohlbold TJ, Krammer F. In the shadow of hemagglutinin: a growing interest in influenza viral neuraminidase and its role as a vaccine antigen. Viruses. 2014;6(6):2465–2494. doi: 10.3390/v6062465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Wang TT, Palese P. Biochemistry. Catching a moving target. Science. 2011;333(6044):834–835. doi: 10.1126/science.1210724. [DOI] [PubMed] [Google Scholar]
- 5.Krammer F, Palese P. Advances in the development of influenza virus vaccines. Nat Rev Drug Discov. 2015;14(3):167–182. doi: 10.1038/nrd4529. [DOI] [PubMed] [Google Scholar]
- 6.Bournazos S, DiLillo DJ, Ravetch JV. The role of Fc-FcγR interactions in IgG-mediated microbial neutralization. J Exp Med. 2015;212(9):1361–1369. doi: 10.1084/jem.20151267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.DiLillo DJ, Tan GS, Palese P, Ravetch JV. Broadly neutralizing hemagglutinin stalk-specific antibodies require FcgammaR interactions for protection against influenza virus in vivo. Nat Med. 2014;20(2):143–151. doi: 10.1038/nm.3443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Corti D, et al. A neutralizing antibody selected from plasma cells that binds to group 1 and group 2 influenza A hemagglutinins. Science. 2011;333(6044):850–856. doi: 10.1126/science.1205669. [DOI] [PubMed] [Google Scholar]
- 9.Pincetic A, et al. Type I and type II Fc receptors regulate innate and adaptive immunity. Nat Immunol. 2014;15(8):707–716. doi: 10.1038/ni.2939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Nimmerjahn F, Ravetch JV. Divergent immunoglobulin g subclass activity through selective Fc receptor binding. Science. 2005;310(5753):1510–1512. doi: 10.1126/science.1118948. [DOI] [PubMed] [Google Scholar]
- 11.Takai T, Li M, Sylvestre D, Clynes R, Ravetch JV. FcR gamma chain deletion results in pleiotrophic effector cell defects. Cell. 1994;76(3):519–529. doi: 10.1016/0092-8674(94)90115-5. [DOI] [PubMed] [Google Scholar]
- 12.et al. FcγRs modulate the anti-tumor activity of antibodies targeting the PD-1/PD-L1 axis. Cancer Cell. 2015;28(3):285–295. doi: 10.1016/j.ccell.2015.08.004. [DOI] [PubMed] [Google Scholar]
- 13.Smith P, DiLillo DJ, Bournazos S, Li F, Ravetch JV. Mouse model recapitulating human Fcgamma receptor structural and functional diversity. Proc Natl Acad Sci U S A. 2012;109(16):6181–6186. doi: 10.1073/pnas.1203954109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.El Bakkouri K, et al. Universal vaccine based on ectodomain of matrix protein 2 of influenza A: Fc receptors and alveolar macrophages mediate protection. J Immunol. 2011;186(2):1022–1031. doi: 10.4049/jimmunol.0902147. [DOI] [PubMed] [Google Scholar]
- 15.Corti D, et al. Heterosubtypic neutralizing antibodies are produced by individuals immunized with a seasonal influenza vaccine. J Clin Invest. 2010;120(5):1663–1673. doi: 10.1172/JCI41902. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.