

Abstract
Inflammation participates in the pathogenesis of many liver diseases, including liver cirrhosis. Certain inflammatory citokines, such as interleukin (IL)-1β and IL-18, are produced after the activation of a multiprotein complex known as the inflammasome. Activation of the inflammasome has been documented in several liver diseases, but its role in the development and progression of liver cirrhosis or the complications associated with this disease is still largely unknown. We have recently studied the impact of the inflammasome in the sterile inflammatory response that takes place in the ascitic fluid of patients with decompensated cirrhosis, providing evidence that activation of the absent in melanoma 2 (AIM2) inflammasome is an important response in these patients. Ascitic fluid-derived macrophages were able to mount a very robust AIM2-mediated response even in the absence of a priming signal, which is usually required for the full activation of all the inflammasomes. In addition, high level of inflammasome activation in these patients was associated with a higher degree of liver disease and an increased incidence of spontaneous bacterial peritonitis. These results may help explain the exacerbated inflammatory response that usually occurs in patients with decompensated cirrhosis in the absence of detectable infections. Thus, inflammasomes should be considered as possible therapeutic targets in sterile inflammatory complications in patients with cirrhosis.
Keywords: Cirrhosis, Ascites, Inflammasome, Absent in melanoma 2, Interleukin-1β
Core tip: In this Editorial I discuss the involvement of the inflammasome in the inflammatory reactions that occur in patients with liver cirrhosis and ascites. I focus on a recent work in which we observed that the absent in melanoma 2 inflammasome is highly activated in the ascitic fluid of patients with advanced cirrhosis and that its activation is linked to the severity of liver disease. These findings are important for the understanding of the sterile inflammatory reactions in these patients, and could have important therapeutic implications.
TEXT
Liver cirrhosis is the result of a long pathologic process initiated by chronic infection with hepatitis B virus or hepatitis C virus (HCV), excessive alcohol consumption, accumulation of fat in liver cells, and other metabolic alterations. The most important complications of liver cirrhosis include intestinal bleeding, encephalopathy, and ascites, and the development of any of these complications is clinically known as decompensated cirrhosis. Ascites is the most common cause of hepatic decompensation, and usually precedes the others. Decompensation of cirrhosis is usually associated with a systemic inflammatory response characterized by activation of innate immune cells and elevated expression of pro-inflammatory cytokines [tumor necrosis factor α, interleukin (IL)-1β, IL-6] in the ascitic fluid. This inflammatory response is usually the result of bacterial translocation from the intestinal lumen to extra-intestinal sites, such as mesenteric lymph nodes and ascitic fluid. Bacterial translocation does not necessarily mean bacterial infection, since the translocating bacteria is often killed by the innate immune system. However, the solely presence of molecules of microbial origin [such as lipopolysaccharide (LPS) or bacterial DNA] is sufficient to mount a sterile inflammatory response in the ascitic fluid in the absence of active infection[1-3].
Since its first description in 2002 by Martinon et al[4], the inflammasome has been a key subject of research in multiple inflammatory diseases. Very comprehensive reviews of the expression, activation, and function of the inflammasomes have been published elsewhere[5-7], and therefore these topics are discussed here only briefly. The inflammasome is a cytosolic multiprotein complex that controls the activation of the enzyme caspase-1[4,6]. Once activated, caspase-1 mediates the maturation and release of pro-inflammatory cytokines such as IL-1β and IL-18. Caspase-1 activity can also result in a highly inflammatory form of cell death called “pyroptosis” in some cells[8], which occurs most frequently upon infection with intracellular pathogens[9]. Inflammasomes are assembled upon recognition of pathogen-associated molecular patterns (PAMPs), as well as host-derived signals known as damage-associated molecular patterns (DAMPs) that are released as a result of tissue damage or cellular stress. Several members of the NLR family (nucleotide-binding and oligomerization domain and leucine-rich-repeat-containing proteins) have been reported to exhibit inflammasome activity, including NLRP1, NLRP3, NLRP6 or NLRC4. In addition to NLRs, the HIN-200 domain-containing protein absent in melanoma 2 (AIM2) has also the ability to induce inflammasome activation. Full activation of the inflammasome requires two different signals. The first signal is provided by the activation of pattern recognition receptors, such as Toll-like receptors, resulting in the accumulation of inactive pro-IL-1β and pro-IL-18 inside the cell. The second signal is then provided by the activation of NLRPs or AIM2 by different danger signals[5-7]. For example, NLRP3 is activated by a wide range of PAMPs and DAMPs (e.g., toxins, uric acid, ATP), whereas AIM2 is activated only by double-stranded DNA (dsDNA) of any origin[10,11] (Figure 1).
Figure 1.
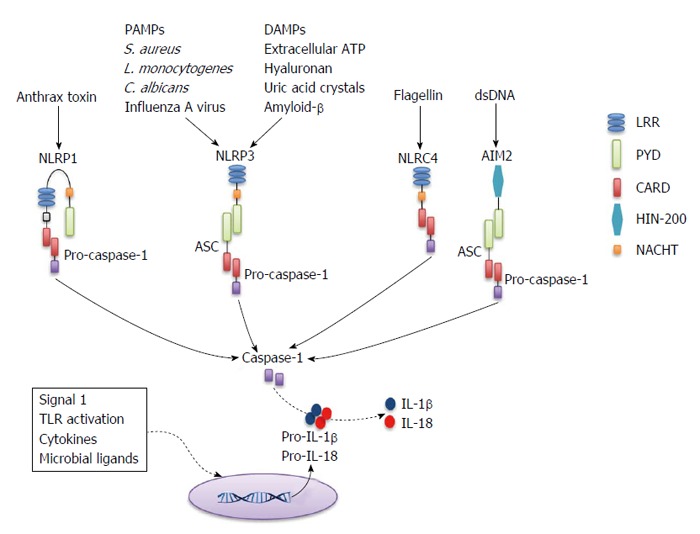
Basic representation of inflammasome activation. Inflammasomes are formed after NLR or PYHIN family members recognize signals associated with tissue damage or infection. Receptors that have a CARD domain can recruit pro-caspase-1 directly (e.g., NLRC4), whereas those that contain a PYD domain (e.g., NLRP3 and AIM2) recruit pro-caspase-1 through the accessory protein ASC (which contains a PYD and a CARD). NLRP1 contains a CARD and can bypass the requirement for ASC, but also contains a PYD and its interaction with ASC enhances the activity of the NLRP1 inflammasome. CARD: Caspase-1 recruitment domain; PYD: Pyrin domain; ASC: Apoptosis-associated speck-like protein containing a CARD; LRR: Leucine rich repeat; HIN-200: Hematopoietic interferon-inducible nuclear antigen with 200 amino-acid repeat; PAMPs: Pathogen-associated molecular patterns; DAMPs: Damage-associated molecular patterns; AIM2: Absent in melanoma 2; dsDNA: Double-stranded DNA; IL: Interleukin; S. aureus: Staphylococcus aureus; L. monocytogenes: Listeria monocytogenes; C. albicans: Candida albicans; TLR: Toll-like receptor.
Recent studies have suggested that the inflammasome also plays an important role in chronic liver disease[12]. For example, the inflammasome is activated in response to HCV infection[13], in drug-induced liver injury[14], or in the pathogenesis of non-alcoholic steatohepatitis[15,16]. However, the inflammasome-mediated response in decompensated cirrhosis was unexplored until publication of our recent study by Lozano-Ruiz et al[17]. In this study we show that activation of the inflammasome is an important response in the ascitic fluid of cirrhotic patients. Macrophages from ascitic fluid showed high levels of pro-IL-1β and pro-IL-18 mRNA, constitutive activation of caspase-1 and enhanced expression of AIM2 protein and mRNA when compared to blood-derived macrophages from the same patients. Moreover, contrary to blood macrophages, activation of the AIM2 inflammasome did not require a priming signal in these cells, demonstrating the pre-activated state of the inflammasome in the ascitic fluid. This pre-activated state of the AIM2 inflammasome was associated with the presence of bacterial DNA fragments in the ascitic fluid of these patients, suggesting that translocation of bacteria and their products could be responsible for this priming[17].
It was previously shown that bacterial translocation and inflammation increases depending on the degree of liver damage and the clinical stage of the disease[18]. Thus, it was conceivable that the severity of liver disease could affect the extent of inflammasome activation in cirrhosis. Indeed, activation of caspase-1 and AIM2-mediated production of IL-1β and IL-18 were increased in patients with Child-Pugh C score, compared to those with Child-Pugh B. Additionally, high level of IL-18 in ascitic fluid showed a significant association with the occurrence of spontaneous bacterial peritonitis (SBP) in these patients independently of the Child-Pugh score, suggesting that increased inflammasome activation might be a marker of increased risk of SBP.
In summary, these findings are important for the understanding of the sterile inflammatory reactions in patients with advanced cirrhosis. In these patients, complications associated with high mortality are normally accompanied by excessive inflammation, and therefore our results could have important translational implications. We propose that a two-hit process could explain the exacerbated inflammasome activation in advanced cirrhosis (Figure 2). First, bacterial translocation would lead to an abnormal influx of exogenous PAMPs (e.g., LPS or bacterial DNA) that induce a pre-activation state of the inflammasome in ascitic fluid cells. Second, endogenous DAMPs released from damaged liver cells (e.g., host dsDNA) would provide the second signal for the activation of the AIM2 inflammasome and the promotion of inflammation in the absence of active infection[17]. However, some questions remain that need to be further clarified. For example, it is not clear whether the inflammasome contributes to, or is a consequence of, cirrhosis progression. In addition, the use of the inflammasome as a therapeutic target in cirrhosis needs to be carefully addressed. Several IL-1β blocking agents are currently approved and used in patients suffering from different inflammatory diseases[19], but the increased risk of infections would argue against using these immunosuppressive drugs in certain situations, such as in SBP. Therefore, more studies are needed to determine the exact role of the inflammasome in the pathogenesis of advanced cirrhosis and its potential use as a therapeutic target for the treatment or prevention of inflammatory complications.
Figure 2.
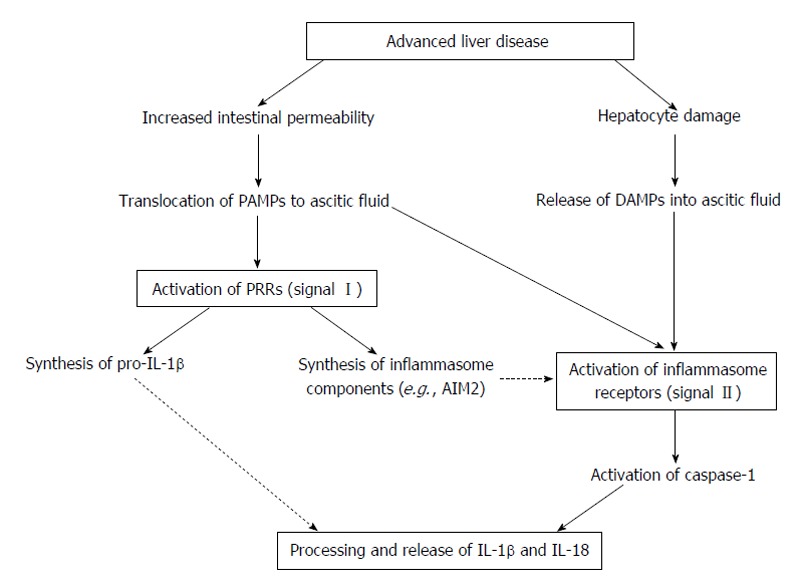
Theoretical mechanism of inflammasome activation in ascitic fluid. Advanced cirrhosis is typically associated with overgrowth of intestinal bacteria and increased intestinal permeability, which results in the translocation of bacterial products (e.g., DNA or LPS) to the ascitic fluid. The presence of these PAMPs activates PRRs in innate immune cells of the ascitic fluid, inducing the synthesis of IL-1β and IL-18 precursors and inflammasome components (signal I). At the same time, continuous liver damage (e.g., by virus or alcohol) would result in hepatocyte death and release of DAMPs (e.g., host dsDNA). These DAMPs (and probably new translocation events of PAMPs from the intestinal lumen) would activate inflammasome-forming receptors such as AIM2 (providing signal II), which in turn results in the activation of caspase-1 and the maturation and release of IL-1β and IL-18 into the ascitic fluid. IL: Interleukin; PAMPs: Pathogen-associated molecular patterns; DAMPs: Damage-associated molecular patterns; AIM2: Absent in melanoma 2; dsDNA: Double-stranded DNA; PRRs: Pattern recognition receptors; LPS: Lipopolysaccharide.
Footnotes
Supported by Grant PI13/00315 from the Instituto de Salud Carlos III (co-financed by FEDER funds); and grants UGP-14-123 and UGP-14-248 from FISABIO Research Foundation.
Conflict-of-interest statement: The author declares no conflict of interest.
Open-Access: This article is an open-access article which was selected by an in-house editor and fully peer-reviewed by external reviewers. It is distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Peer-review started: January 20, 2015
First decision: March 6, 2015
Article in press: January 19, 2016
P- Reviewer: Liu ZH, Tsuchiya A, Uchiyama H S- Editor: Gong XM L- Editor: A E- Editor: Liu SQ
References
- 1.Francés R, Zapater P, González-Navajas JM, Muñoz C, Caño R, Moreu R, Pascual S, Bellot P, Pérez-Mateo M, Such J. Bacterial DNA in patients with cirrhosis and noninfected ascites mimics the soluble immune response established in patients with spontaneous bacterial peritonitis. Hepatology. 2008;47:978–985. doi: 10.1002/hep.22083. [DOI] [PubMed] [Google Scholar]
- 2.González-Navajas JM, Bellot P, Francés R, Zapater P, Muñoz C, García-Pagán JC, Pascual S, Pérez-Mateo M, Bosch J, Such J. Presence of bacterial-DNA in cirrhosis identifies a subgroup of patients with marked inflammatory response not related to endotoxin. J Hepatol. 2008;48:61–67. doi: 10.1016/j.jhep.2007.08.012. [DOI] [PubMed] [Google Scholar]
- 3.Zapater P, Francés R, González-Navajas JM, de la Hoz MA, Moreu R, Pascual S, Monfort D, Montoliu S, Vila C, Escudero A, et al. Serum and ascitic fluid bacterial DNA: a new independent prognostic factor in noninfected patients with cirrhosis. Hepatology. 2008;48:1924–1931. doi: 10.1002/hep.22564. [DOI] [PubMed] [Google Scholar]
- 4.Martinon F, Burns K, Tschopp J. The inflammasome: a molecular platform triggering activation of inflammatory caspases and processing of proIL-beta. Mol Cell. 2002;10:417–426. doi: 10.1016/s1097-2765(02)00599-3. [DOI] [PubMed] [Google Scholar]
- 5.Schroder K, Tschopp J. The inflammasomes. Cell. 2010;140:821–832. doi: 10.1016/j.cell.2010.01.040. [DOI] [PubMed] [Google Scholar]
- 6.Rathinam VA, Vanaja SK, Fitzgerald KA. Regulation of inflammasome signaling. Nat Immunol. 2012;13:333–342. doi: 10.1038/ni.2237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Strowig T, Henao-Mejia J, Elinav E, Flavell R. Inflammasomes in health and disease. Nature. 2012;481:278–286. doi: 10.1038/nature10759. [DOI] [PubMed] [Google Scholar]
- 8.Bergsbaken T, Fink SL, Cookson BT. Pyroptosis: host cell death and inflammation. Nat Rev Microbiol. 2009;7:99–109. doi: 10.1038/nrmicro2070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Case CL, Shin S, Roy CR. Asc and Ipaf Inflammasomes direct distinct pathways for caspase-1 activation in response to Legionella pneumophila. Infect Immun. 2009;77:1981–1991. doi: 10.1128/IAI.01382-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Fernandes-Alnemri T, Yu JW, Datta P, Wu J, Alnemri ES. AIM2 activates the inflammasome and cell death in response to cytoplasmic DNA. Nature. 2009;458:509–513. doi: 10.1038/nature07710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hornung V, Ablasser A, Charrel-Dennis M, Bauernfeind F, Horvath G, Caffrey DR, Latz E, Fitzgerald KA. AIM2 recognizes cytosolic dsDNA and forms a caspase-1-activating inflammasome with ASC. Nature. 2009;458:514–518. doi: 10.1038/nature07725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Szabo G, Csak T. Inflammasomes in liver diseases. J Hepatol. 2012;57:642–654. doi: 10.1016/j.jhep.2012.03.035. [DOI] [PubMed] [Google Scholar]
- 13.Burdette D, Haskett A, Presser L, McRae S, Iqbal J, Waris G. Hepatitis C virus activates interleukin-1β via caspase-1-inflammasome complex. J Gen Virol. 2012;93:235–246. doi: 10.1099/vir.0.034033-0. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 14.Imaeda AB, Watanabe A, Sohail MA, Mahmood S, Mohamadnejad M, Sutterwala FS, Flavell RA, Mehal WZ. Acetaminophen-induced hepatotoxicity in mice is dependent on Tlr9 and the Nalp3 inflammasome. J Clin Invest. 2009;119:305–314. doi: 10.1172/JCI35958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kamari Y, Shaish A, Vax E, Shemesh S, Kandel-Kfir M, Arbel Y, Olteanu S, Barshack I, Dotan S, Voronov E, et al. Lack of interleukin-1α or interleukin-1β inhibits transformation of steatosis to steatohepatitis and liver fibrosis in hypercholesterolemic mice. J Hepatol. 2011;55:1086–1094. doi: 10.1016/j.jhep.2011.01.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Henao-Mejia J, Elinav E, Jin C, Hao L, Mehal WZ, Strowig T, Thaiss CA, Kau AL, Eisenbarth SC, Jurczak MJ, et al. Inflammasome-mediated dysbiosis regulates progression of NAFLD and obesity. Nature. 2012;482:179–185. doi: 10.1038/nature10809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Lozano-Ruiz B, Bachiller V, García-Martínez I, Zapater P, Gómez-Hurtado I, Moratalla A, Giménez P, Bellot P, Francés R, Such J, et al. Absent in melanoma 2 triggers a heightened inflammasome response in ascitic fluid macrophages of patients with cirrhosis. J Hepatol. 2015;62:64–71. doi: 10.1016/j.jhep.2014.08.027. [DOI] [PubMed] [Google Scholar]
- 18.Cirera I, Bauer TM, Navasa M, Vila J, Grande L, Taurá P, Fuster J, García-Valdecasas JC, Lacy A, Suárez MJ, et al. Bacterial translocation of enteric organisms in patients with cirrhosis. J Hepatol. 2001;34:32–37. doi: 10.1016/s0168-8278(00)00013-1. [DOI] [PubMed] [Google Scholar]
- 19.Hoffman HM, Wanderer AA. Inflammasome and IL-1beta-mediated disorders. Curr Allergy Asthma Rep. 2010;10:229–235. doi: 10.1007/s11882-010-0109-z. [DOI] [PMC free article] [PubMed] [Google Scholar]