

Abstract
Psoriasis vulgaris is a common chronic inflammatory skin disease characterized by the hyperproliferation and abnormal differentiation of keratinocytes. CCATT/enhancer binding protein α (C/EBPα) is abundant in the epidermis and is associated with the proliferation of keratinocytes. However, the role of C/EBPα in the proliferation of keratinocytes and the pathogenesis of psoriasis vulgaris are yet to be elucidated. In the present study, using two-step immunohistochemistry, the expression levels of C/EBPα and Ki-67 were examined in skin biopsies harvested from 30 patients with psoriasis vulgaris and 30 healthy control subjects. The proliferation index (PI) was calculated and the correlation between C/EBPα expression levels and the PI was assessed using Pearson's correlation coefficient. In addition, the effect on HaCaT immortalized human keratinocytic cells of treatment with various concentrations of interleukin (IL)-17 was investigated. Subsequently, cell proliferation rates were examined using a Cell Counting kit-8 assay and the mRNA and protein expression levels of C/EBPα were analyzed using semiquantitative reverse transcription-polymerase chain reaction and western blotting, respectively, in order to analyze the effects of IL-17 stimulation on C/EBPα expression levels. C/EBPα expression was predominantly detected in the cytoplasm of the keratinocytes and C/EBPα expression levels were significantly lower in the psoriatic lesions (P<0.05), as compared with the control group. An inverse correlation was detected between the expression levels of C/EBPα and the PI in the psoriatic lesions. Furthermore, a significant increase in the cell proliferation rate and significant reductions in the mRNA and protein expression levels of C/EBPα were detected in HaCaT cells following treatment with IL-17. These results demonstrated that C/EBPα may act as a downstream target of IL-17 and may be associated with the pathogenesis of psoriasis.
Keywords: psoriasis vulgaris, keratinocyte, proliferation, CCATT/enhancer binding protein α, interleukin-17
Introduction
Psoriasis vulgaris is a chronic inflammatory disorder characterized by the hyperproliferation and abnormal differentiation of keratinocytes and the infiltration of inflammatory cells into the dermis and epidermis. Although its exact pathogenesis is yet to be elucidated, psoriasis vulgaris is associated with various genetic, immunological, psychological and environmental factors (1). T-cell-mediated specific and nonspecific immune disorders associated with the genetic background of the patient have been demonstrated to have a central role in the pathogenesis of psoriasis (2). T-helper (Th)17 cells, which are a newly discovered subset of Th cells, are capable of secreting an important pro-inflammatory cytokine, interleukin (IL)-17, and the Th-17/IL-17 axis has a key role in the immunopathogenesis of psoriasis vulgaris (3,4).
The CCAAT/enhancer binding protein (C/EBP) family, which consists of a group of basic transcription factors containing a leucine zipper structure, is critical to the proliferation and differentiation of various cell types (5). C/EBPα, which was the first cloned member of the C/EBP family with 358 amino acid residues, contains a C-terminal leucine-zipper protein interaction domain, a basic DNA binding region and an N-terminal transcriptional activation domain (6). C/EBPα is expressed in various highly differentiated cells and tissues, including peripheral blood mononuclear cells, and adipose, liver and lung tissues (7). Previous studies have demonstrated that C/EBPα is expressed in every layer of the epidermis; therefore, C/EBPα may regulate the proliferation and differentiation of keratinocytes (8,9).
In the present study, the expression levels of C/EBPα and Ki-67 were investigated in psoriasis vulgaris lesions using immunohistochemistry, and the proliferation index (PI) was subsequently calculated to explore the correlation between C/EBPα expression levels and PI. In addition, the mRNA and protein expression levels of C/EBPα and the cell proliferation rate were examined in HaCaT cells, an immortalized human keratinocytic cell line, following treatment with various concentrations of IL-17 in vitro.
Materials and methods
Patients and skin biopsies
Skin biopsies were obtained from 30 patients with active stage psoriasis vulgaris, including 14 males and 16 females aged 13–45 years (mean, 27.77±8.64 years), who presented at the Department of Dermatology at Qilu Hospital (Jinan, China). None of the participants had previously been diagnosed with another systemic disease and no local or systematic therapy was administered for ≥2 months prior to the biopsy. Control skin samples were obtained from 30 gender- and age-matched healthy participants (14 males and 16 females) aged 9–51 years old (mean, 31.80±14.86 years). Written informed consent was provided by all participants prior to the skin biopsies and the present study was approved by the Qilu Hospital Ethics Committee of Shandong University.
Immunohistochemistry
Following excision, the skin samples were immediately fixed in 10% neutral formaldehyde, embedded in paraffin and serially cut into 3-µm-thick sections. Subsequently, the sections were deparaffinized with xylene and rehydrated using a graded ethanol series. Antigen retrieval was conducted by incubation with 1 mM ethylenediaminetetraacetic acid (EDTA) buffer (pH 8.0) at 120°C under high pressure for 5 min. Endogenous peroxidase activity was terminated using 3% hydrogen peroxide solution and the sections were subsequently blocked with 5% goat serum (C-0005; BIOSS; Beijing, China) to prevent nonspecific background staining. Following this, the sections were incubated at 37°C for 1 h with rabbit anti-human C/EBPα polyclonal antibody (1:100; bs-16030R) and mouse anti-human Ki-67 ready-to-use monoclonal antibody (1:100; bs-2130R; both BIOSS). After washing three times with phosphate-buffered saline, sections were incubated using a PV-6000 Polymer Detection system (ZSGB-Bio, Beijing, China), according to the manufacturers protocol, stained with 3,3-diaminobenzidine substrate (ZSGB-Bio) for color development and subsequently counterstained with Harris hematoxylin (Beyotime Institute of Biotechnology, Jiangsu, China) for examination under a light microscope (DM1000 LED; Leica Microsystems, Inc., Buffalo Grove, IL, USA). Keratinocytes were deemed positive for C/EBPα or Ki-67 expression if yellow to brown granules were detected in the cytoplasm or nucleus, respectively. Three fields were randomly selected under high magnification (×400) and the mean gray value of the positive area was analyzed using Image Pro Plus (IPP) software, version 6.0 (Media Cybernetics, Inc., Rockville, MD, USA), where a high grey value indicated lower expression levels. Five representative fields with the highest number of positive cells were counted in each section (magnification, ×400) and the PI was calculated as the percentage of total epidermal cells that were Ki-67-positive.
Cell culture
HaCaT immortalized human keratinocyte cell line was purchased from the American Type Culture Collection (Manassas, VA, USA). HaCaT cells were cultured in Dulbeccos modified Eagles medium (DMEM) (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented with 10% fetal bovine serum (vol/vol), 100 U/ml penicillin and 100 µg/ml streptomycin (C0222; Beyotime Institute of Biotechnology) in a humidified incubator containing 5% CO2 at 37°C. Cells were digested with 0.25% trypsin/0.02% EDTA (1:1) solution (C0203; Beyotime Institute of Biotechnology) and passaged (1:2) every 2–3 days. Subsequently, the HaCaT cells were cultured in 12-well plates (1×105 cells/ml) and grown to 60–70% confluence, prior to stimulation with 50, 70 and 90 ng/ml IL-17 for 24 h. Following treatment, cells were harvested for RNA isolation and protein extraction, respectively. DMEM was simultaneously used without IL-17 to treat the control group. Experiments were performed in triplicate.
Cell Counting kit-8 Assay
A total of 5×103 HaCaT cells/ml in the logarithmic growth phase were cultured in 96-well plates. At 60–70% confluence, cells were treated with 50, 70 or 90 ng/ml IL-17 for 24 h. Subsequently, cells were incubated with CCK-8 reagent at 37°C for 2 h, according to the manufacturer's protocol (Dojindo Molecular Technologies, Inc., Kumamoto, Japan). A Varioskan™ Flash microplate reader (Thermo Fisher Scientific, Inc.) was used to measure the optical density (OD) at 450 nm. Cell viability (% of control) was calculated as follows: (ODtest - ODblank)/(ODcontrol - ODblank). Experiments were performed in sextuplicate.
Semi-quantitative reverse transcription-polymerase chain reaction (RT-PCR)
Following treatment with IL-17 for 24 h, total RNA was isolated from the HaCaT cells using an RNAiso Plus kit (Takara Biotechnology Co., Ltd., Dalian, China), according to the manufacturer's protocol, and detected using a Nanodrop™ 2000 spectrophotometer at 260 nm (Nanodrop; Thermo Fisher Scientific, Inc.). A total of 1 µg RNA was reverse transcribed into cDNA using oligodeoxythymine nucleotide primers and subsequently amplified using a Power RT kit (BioTeke Corporation, Beijing, China). Specific primer pairs were synthesized by Sangon Biotech Co., Ltd. (Shanghai, China) and their sequences are listed in Table I. PCR was performed using a 50-µl system containing: 25 µl 2X Super Taq PCR MasterMix (PR1732; BioTeke Corporation), 2.5 µl forward and reverse primers, 10 µl template and 10 µl triple-distilled water. Reaction conditions were as follows: 95°C for 5 min, followed by 35 cycles of 95°C for 1 min, 60°C for 45 sec, 72°C for 45 sec, and 72°C for 10 min. The amplified PCR products were separated by 2% agarose gel electrophoresis. The sizes of the C/EBPα and β-actin fragments were 375 and 495 bp, respectively. Ethidium bromide-stained PCR products were visualized using an ultraviolet transilluminator (Tanon™ 2500R; Tanon Science and Technology Co., Ltd., Shanghai, China) and gray values of the DNA bands were simultaneously measured using the automatic image analyzer software. Relative gray values were measured as the ratio of gray value between C/EBPα and β-actin.
Table I.
Primer sequences.
| Gene | Primer | Sequence (5′-3′) |
|---|---|---|
| C/EBPα | Upstream | CACCGCTCCAATGCCTAC |
| Downstream | CCCATCGCAGTGAGTTCCG | |
| β-actin | Upstream | AAGTACTCCGTGTGGATCGG |
| Downstream | ATGCTATCACCTCCCCTGTG |
C/EBPα, CCATT/enhancer binding protein α.
Western blotting
HaCaT cells were dissolved in lysis buffer containing 1 mM phenylmethanesulfonyl fluoride (Cell Lysis Buffer for Western and IP; Beyotime Institute of Biotechnology) on ice. Samples were centrifuged at 12,000 × g for 5 min and the total protein content in the supernatant was analyzed using an Enhanced BCA Protein Assay kit (Beyotime Institute of Biotechnology). A total of 30 µg protein from the cell lysates was separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to a polyvinylidene difluoride (PVDF) membrane. The PVDF membrane was subsequently blocked with 5% skimmed milk at room temperature for 1 h and incubated at 4℃ overnight with rabbit anti-human C/EBPα polyclonal antibody (1:400; bs-16030R; BIOSS), or mouse anti-human anti-β-actin antibody (1:1,000; sc-47778; Santa Cruz Biotechnology, Inc., Dallas, TX, USA). After washing with 0.01 M Tris-buffered saline containing 0.1% Tween-20, the membrane was incubated with horseradish peroxidase-labeled goat-anti-rabbit or goat-anti-mouse immunoglobulin G antibody (1:1,000; E030120-01; EarthOx Life Sciences, Millbrae, CA, USA) at room temperature for 1 h. A standard enhanced chemiluminescence reaction (Sangon Biotech Co., Ltd., Shanghai, China) was performed, according to the manufacturer's protocol, and densitometric analysis of the band intensity was performed using a Tanon™ 5000 chemiluminescence imaging analysis system (Tanon Science and Technology Co., Ltd.). Relative protein expression levels were calculated as gray value ratios as follows: C/EBPα integrated option density (IOD)/β-actin IOD.
Statistical analysis
SPSS software, version 19.0 (SPSS, Inc., Chicago, IL, USA) was used for all statistical analyses. Data were expressed as the mean ± standard deviation. The intensity of immunohistochemical staining of C/EBPα between the psoriasis and control groups was analyzed using Students t-test. Analyses of the correlation between C/EBPα expression levels and PI in the psoriatic lesions were performed using Pearson's correlation coefficient. mRNA and protein expression levels of C/EBPα were analyzed between the psoriasis and control groups following the administration of various concentrations of IL-17 using one-way analysis of variance with Student-Newman-Keuls post-hoc analysis. P<0.05 was considered to indicate a statistically significant difference.
Results
Immunohistochemical staining of C/EBPα in psoriatic lesions
C/EBPα was expressed in the cytoplasm of the majority of keratinocytes and in the whole epidermis of the healthy control skin samples, with sporadic expression detected in the basal layer and enhanced expression levels detected in the stratum corneum. Staining of C/EBPα was weaker in the psoriasis vulgaris group, as compared with the control group. Immunohistochemical staining demonstrated the grey value of C/EBPα was 51.94±7.60 in the control group, as compared with 85.44±13.88 in the psoriasis vulgaris group. Therefore, significantly reduced C/EBPα expression levels were detected in the psoriatic lesions (t=7.819, P<0.01), as compared with the control skin samples (Fig. 1).
Figure 1.
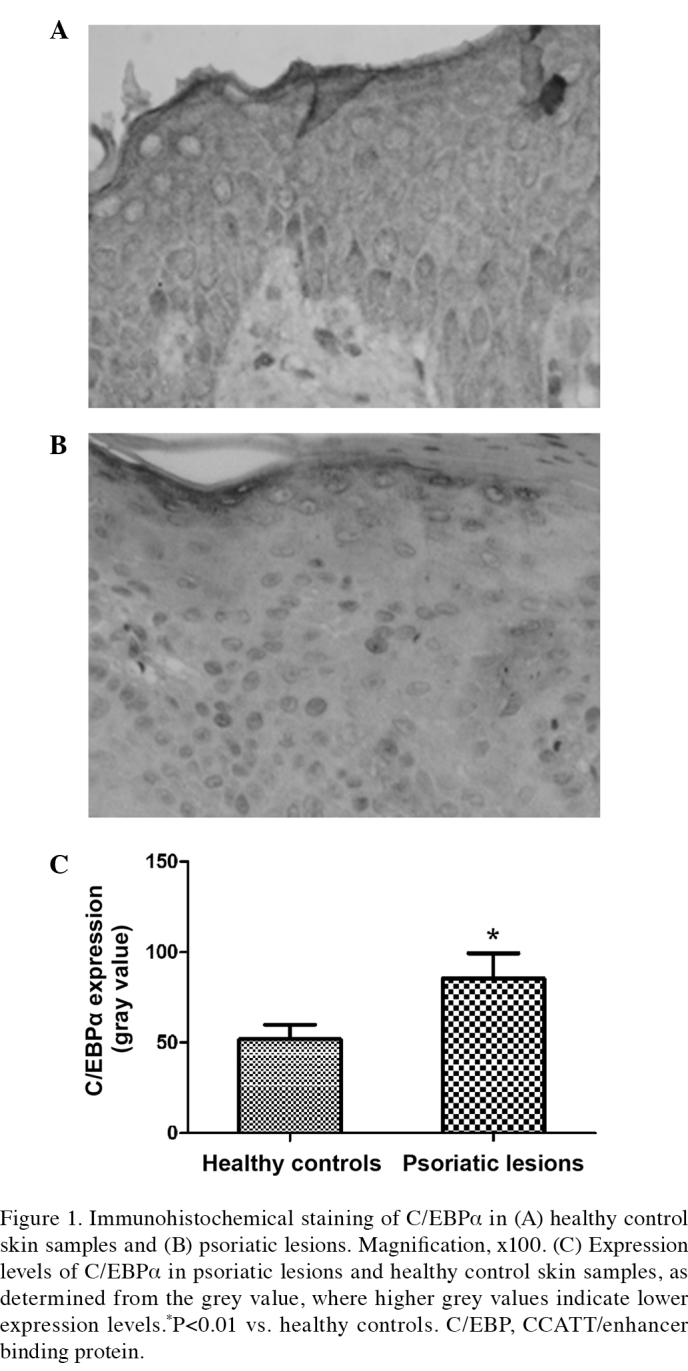
Immunohistochemical staining of C/EBPα in (A) healthy control skin samples and (B) psoriatic lesions. Magnification, ×100. (C) Expression levels of C/EBPα in psoriatic lesions and healthy control skin samples, as determined from the grey value, where higher grey values indicate lower expression levels.*P<0.01 vs. healthy controls. C/EBP, CCATT/enhancer binding protein.
Correlation between C/EBPα expression levels and PI
Immunohistochemical staining demonstrated that Ki-67 was predominantly expressed in the cytoplasm of keratinocytes and was sporadically expressed in the basal layer of the control skin samples. Ki-67-positive cells were detected in the basal layer and, notably, in the middle and lower parts of the stratum spinosum of psoriasis vulgaris lesions. The PI was significantly increased in the psoriasis vulgaris group (0.20±0.09), as compared with 0.66±0.03 in the control group (t=4.535; P<0.01; Fig. 2A). Pearson correlation analysis demonstrated that there was a significant positive correlation between the grey values of C/EBPα and the PI (r=0.654, P<0.01; Fig. 2B), which implied that the expression levels of C/EBPα in the keratinocytes of psoriasis vulgaris were inversely correlated with the PI.
Figure 2.
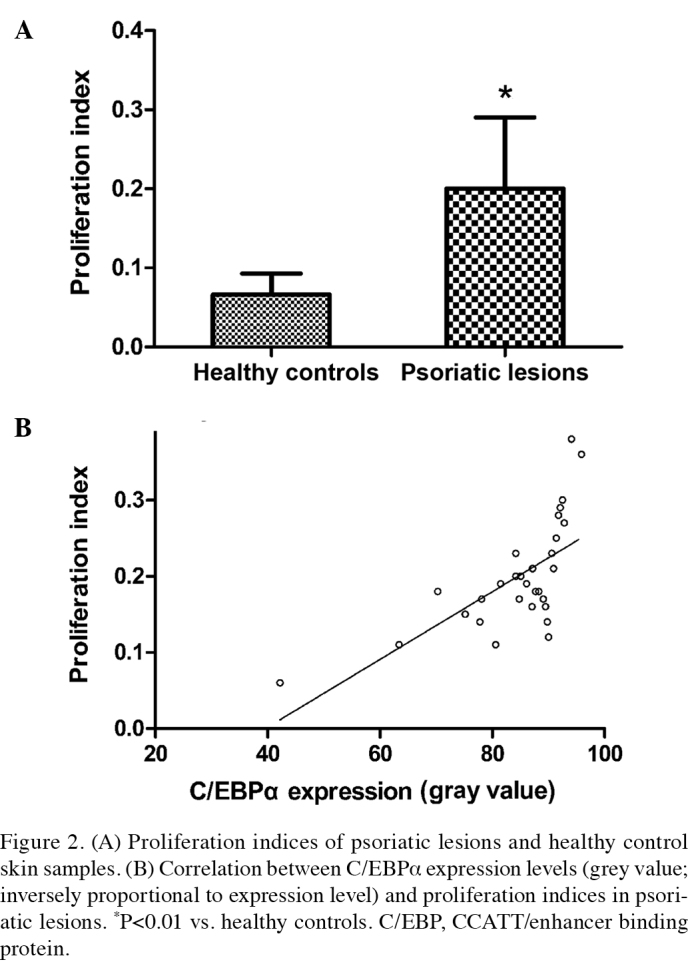
(A) Proliferation indices of psoriatic lesions and healthy control skin samples. (B) Correlation between C/EBPα expression levels (grey value; inversely proportional to expression level) and proliferation indices in psoriatic lesions. *P<0.01 vs. healthy controls. C/EBP, CCATT/enhancer binding protein.
Effect of IL-17 treatment on the proliferation of HaCaT cells and C/EBPα expression levels
In the in vitro experiment, immortalized human keratinocytic HaCaT cells were treated with 50, 70 and 90 ng/ml IL-17 for 24 h. As compared with the control group, IL-17 treatment significantly increased the proliferation of HaCaT cells in a concentration-dependent manner (Fig.3). Following culture with 50, 70 and 90 ng/ml IL-17, semiquantitative RT-PCR and western blot analyses were performed using a β-actin housekeeping control in order to examine the mRNA and protein expression levels of C/EBPα. The results demonstrated that, as compared with the control group, 50, 70 and 90 ng/ml IL-17 significantly reduced the mRNA (q=4.11, P<0.05; q=9.12, P<0.001; q=12.44, P<0.001, respectively) and protein (q=5.293, P<0.01; q=10.17, P<0.001; q=16.82, P<0.001, respectively) expression levels of C/EBPα in a concentration-dependent manner (Fig. 4).
Figure 3.

HaCaT cell proliferation rates following treatment with 50, 70 or 90 ng/ml IL-17 for 24 h, as determined by a Cell Counting kit-8 assay. IL, interleukin.
Figure 4.
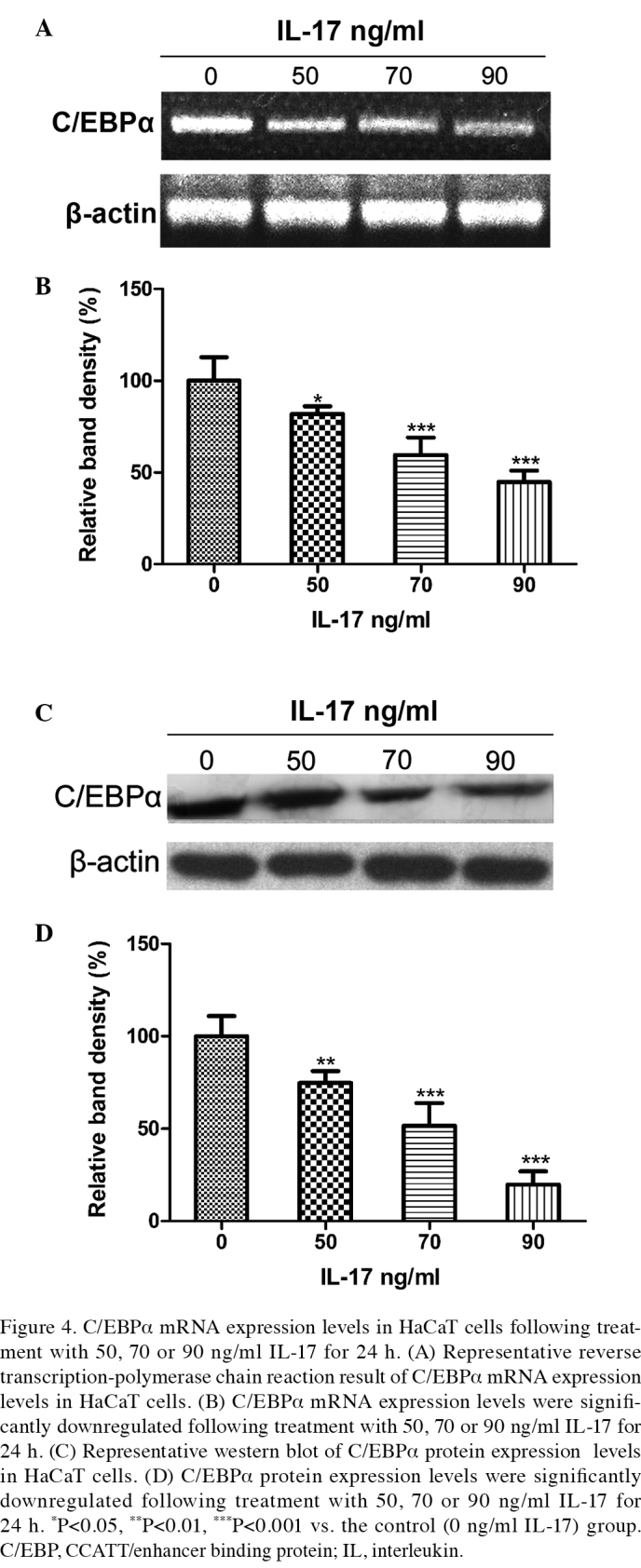
C/EBPα mRNA expression levels in HaCaT cells following treatment with 50, 70 or 90 ng/ml IL-17 for 24 h. (A) Representative reverse transcription-polymerase chain reaction result of C/EBPα mRNA expression levels in HaCaT cells. (B) C/EBPα mRNA expression levels were significantly downregulated following treatment with 50, 70 or 90 ng/ml IL-17 for 24 h. (C) Representative western blot of C/EBPα protein expression levels in HaCaT cells. (D) C/EBPα protein expression levels were significantly downregulated following treatment with 50, 70 or 90 ng/ml IL-17 for 24 h. *P<0.05, **P<0.01, ***P<0.001 vs. the control (0 ng/ml IL-17) group. C/EBP, CCATT/enhancer binding protein; IL, interleukin.
Discussion
Psoriasis vulgaris is a chronic inflammatory disease characterized by a thickened epidermis, hyperproliferation and parakeratosis of keratinocytes and marked inflammatory cell infiltration. It is hypothesized that the abnormal proliferation and differentiation of keratinocytes in psoriatic lesions is associated with the activation of T cells by external stimuli, which subsequently recognize a self-antigen and lead to an immune response, in which the Th-17/IL-17 axis has an important role (10). Th17 cells, which are named according to their secretion of IL-17, are a newly reported subtype of CD4+ T cells that gather in psoriatic lesions (11). Significantly increased expression levels of IL-17 have been detected in psoriatic lesions, as compared with samples from control skin and skin surrounding psoriatic lesions (11). Furthermore, the administration of IL-17 monoclonal antibody to patients with psoriasis is capable of quickly and effectively alleviating the clinical symptoms, improving pathological characteristics and attenuating keratinocyte proliferation and inflammatory cell infiltration into the lesions (12,13). In the form of homogenous or heterogeneous dimers, IL-17 binds to its IL-17 receptor (IL-17R) and subsequently activates the signaling pathways of phosphoinositide 3-kinase (PI3K), nuclear factor (NF)-κB and mitogen-activated protein kinase in order to regulate cell proliferation (14,15). A previous study, based on the upregulation of IL-17R in the keratinocytes of psoriatic lesions, has demonstrated that the administration of IL-17R receptor antagonists may rapidly control the symptoms of psoriasis, and reduce the psoriasis area and severity index (16). This suggests that IL-17 may mediate this immune disorder, resulting in the excessive proliferation and differentiation of keratinocytes.
C/EBPα, which is selectively expressed in a variety of tissue cells, interacts with various tissue-specific transcription factors in order to regulate proliferation and differentiation (17). Furthermore, C/EBPα may be an important negative regulator of cell proliferation. Reduced C/EBPα expression levels have been detected in various types of cancer, including lung, breast, liver and other epithelial tumors; therefore, C/EBPα may be associated with the pathogenesis of cancer (18–20). C/EBPα is abundantly expressed in normal skin tissue and reduced C/EBPα expression levels in some skin tumors have been correlated with uncontrolled cell proliferation and tumor development, in which the re-expression and enhanced expression of C/EBPα was demonstrated to inhibit hyperproliferation (21). Previous studies have demonstrated that C/EBPα protein dimers, rather than the leucine zipper domain, directly interact with various cell cycle-related proteins in order to control the cell cycle process (22,23). C/EBPα is capable of regulating, stabilizing and activating the cell-cycle inhibitor, p21, and is also capable of inhibiting the activities of cyclin-dependent kinase (CDK)2 and CDK4, and E2F-mediated transcriptional processes, to block cell cycle progression (23–25). These mechanisms induce cell cycle arrest at the G1/S phase and inhibit cell proliferation. However, the activation of the PI3K/Akt signaling pathway has a negative regulatory role on the expression of C/EBPα (26).
The present study demonstrated that the proliferation of HaCaT cells was significantly increased and C/EBPα mRNA and protein expression levels were significantly decreased following treatment with 50, 70 and 90 ng/ml IL-17 for 24 h. Furthermore, downregulated expression of C/EBPα was detected in the psoriatic lesions, which was negatively correlated with PI. Therefore, the results of the present study suggest that IL-17 is capable of activating the PI3K pathway via IL-17R, and activation of the PI3K/Akt pathway may reduce the expression of C/EBPα. Hence, IL-17 administration may inhibit keratinocyte proliferation by decreasing the expression levels of C/EBPα, and PI3K may have a critical role in this mechanism.
In conclusion, IL-17 may result in the downregulation of C/EBPα expression in keratinocytes and the IL-17/C/EBPα axis may be associated with hyperproliferation of keratinocytes in psoriasis vulgaris lesions.
Glossary
Abbreviations
- IL-17
interleukin-17
- C/EBPα
CCAAT/enhancer binding protein α
References
- 1.Bowcock AM, Kreuger JG. Getting under the skin: The immunogenetics of psoriasis. Nat Rev Immunol. 2005;5:699–711. doi: 10.1038/nri1689. [DOI] [PubMed] [Google Scholar]
- 2.Lynde CW, Poulin Y, Vender R, Bourcier M, Khalil S. Interleukin 17A: Toward a new understanding of psoriasis pathogenesis. J Am Acad Dermatol. 2014;71:141–150. doi: 10.1016/j.jaad.2013.12.036. [DOI] [PubMed] [Google Scholar]
- 3.Raychaudhuri SP. Role of IL-17 in psoriasis and psoriatic arthritis. Clin Rev Allergy Immunol. 2013;44:183–193. doi: 10.1007/s12016-012-8307-1. [DOI] [PubMed] [Google Scholar]
- 4.Nakajima K. Critical role of the interleukin-23/T-helper 17 cell axis in the pathogenesis of psoriasis. J Dermatol. 2012;39:219–224. doi: 10.1111/j.1346-8138.2011.01458.x. [DOI] [PubMed] [Google Scholar]
- 5.Nerlov C. The C/EBP family of transcription factors: A paradigm for interaction between gene expression and proliferation control. Trends Cell Biol. 2007;17:318–324. doi: 10.1016/j.tcb.2007.07.004. [DOI] [PubMed] [Google Scholar]
- 6.Roe JS, Vakoc CR. C/EBPα: Critical at the origin of leukemic transformation. J Exp Med. 2014;211:1–4. doi: 10.1084/jem.20132530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Zaragoza K, Bégay V, Schuetz A, Heinemann U, Leutz A. Repression of transcriptional activity of C/EBPalpha by E2F-dimerization partner complexes. Mol Cell Biol. 2010;30:2293–2304. doi: 10.1128/MCB.01619-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Johnson PF. Molecular stop signs: R egulation of cell-cycle arrest by C/EBP transcription factors. J Cell Sci. 2005;118:2545–2555. doi: 10.1242/jcs.02459. [DOI] [PubMed] [Google Scholar]
- 9.Oh WJ, Rishi V, Orosz A, Gerdes MJ, Vinson C. Inhibition of CCAAT/enhancer binding protein family DNA binding in mouse epidermis prevents and regresses papillomas. Cancer Res. 2007;67:1867–1876. doi: 10.1158/0008-5472.CAN-06-2746. [DOI] [PubMed] [Google Scholar]
- 10.Leonardi C, Matheson R, Zachariae C, Cameron G, Li L, Edson-Heredia E, Braun D, Banerjee S. Anti-interleukin-17 monoclonal antibody ixekizumab in chronic plaque psoriasis. N Engl J Med. 2012;366:1190–1199. doi: 10.1056/NEJMoa1109997. [DOI] [PubMed] [Google Scholar]
- 11.Kagami S, Rizzo HL, Lee JJ, Koguchi Y, Blauvelt A. Circulating Th17, Th22, and Th1 cells are increased in psoriasis. J Invest Dermatol. 2010;130:1373–1383. doi: 10.1038/jid.2009.399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Reich K. Anti-interleukin-17 monoclonal antibody ixekizumab in psoriasis. N Engl J Med. 2012;367:274–275. doi: 10.1056/NEJMc1205835. [DOI] [PubMed] [Google Scholar]
- 13.Papp KA, Leonardi C, Menter A, Ortonne JP, Krueger JG, Kricorian G, Aras G, Li J, Russell CB, Thompson EH, Baumgartner S. Brodalumab, an anti-interleukin-17-receptor antibody for psoriasis. N Engl J Med. 2012;366:1181–1189. doi: 10.1056/NEJMoa1109017. [DOI] [PubMed] [Google Scholar]
- 14.Kim HR, Cho ML, Kim KW, Juhn JY, Hwang SY, Yoon CH, Park SH, Lee SH, Kim HY. Up-regulation of IL-23p19 expression in rheumatoid arthritis synovial fibroblasts by IL-17 through PI3-kinase-, NF-kappaB-and p38 MAPK-dependent signalling pathways. Rheumatology (Oxford) 2007;46:57–64. doi: 10.1093/rheumatology/kel159. [DOI] [PubMed] [Google Scholar]
- 15.Gu C, Wu L, Li X. IL-17 family, Cytokines, receptors and signaling. Cytokine. 2013;64:477–485. doi: 10.1016/j.cyto.2013.07.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Papp KA, Reid C, Foley P, Sinclair R, Salinger DH, Williams G, Dong H, Krueger JG, Russell CB, Martin DA. Anti-IL-17 receptor antibody AMG 827 leads to rapid clinical response in subjects with moderate to severe psoriasis, Results from a phase I, randomized, placebo-controlled trial. J Invest Dermatol. 2012;132:2466–2469. doi: 10.1038/jid.2012.163. [DOI] [PubMed] [Google Scholar]
- 17.Ramji D, Foka P. CCAAT/enhancer-binding proteins: Structure function and regulation. Biochem J. 2002;365:561–575. doi: 10.1042/bj20020508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Tada Y, Brena RM, Hackanson B, Morrison C, Otterson GA, Plass C. Epigenetic modulation of tumor suppressor CCAAT/enhancer binding protein alpha activity in lung cancer. J Natl Cancer Inst. 2006;98:396–406. doi: 10.1093/jnci/djj093. [DOI] [PubMed] [Google Scholar]
- 19.Seifeddine R, Dreiem A, Blanc E, Fulchignoni-Lataud MC, Le Frère Belda MA, Lecuru F, Mayi TH, Mazure N, Favaudon V, Massaad C, et al. Hypoxia down-regulates CCAAT/enhancer binding protein-alpha expression in breast cancer cells. Cancer Res. 2008;68:2158–2165. doi: 10.1158/0008-5472.CAN-07-1190. [DOI] [PubMed] [Google Scholar]
- 20.Tseng HH, Hwang YH, Yeh KT, Chang JG, Chen YL, Yu HS. Reduced expression of C/EBP alpha protein in hepatocellular carcinoma is associated with advanced tumor stage and shortened patient survival. J Cancer Res Clin Oncol. 2009;135:241–247. doi: 10.1007/s00432-008-0448-5. [DOI] [PubMed] [Google Scholar]
- 21.Shim M, Powers KL, Ewing SJ, Zhu S, Smart RC. Diminished expression of C/EBPalpha in skin carcinomas is linked to oncogenic Ras and reexpression of C/EBPalpha in carcinoma cells inhibits proliferation. Cancer Res. 2005;65:861–867. [PubMed] [Google Scholar]
- 22.Reckzeh K, Cammenga J. Molecular mechanisms underlying deregulation of C/EBPalpha in acute myeloid leukemia. Int J Hematol. 2010;91:557–568. doi: 10.1007/s12185-010-0573-1. [DOI] [PubMed] [Google Scholar]
- 23.Harris TE, Albrecht JH, Nakanishi M, Darlington GJ. CCAAT/enhancer-binding protein-alpha cooperates with p21 to inhibit cyclin-dependent kinase-2 activity and induces growth arrest independent of DNA binding. J Biol Chem. 2001;276:29200–29209. doi: 10.1074/jbc.M011587200. [DOI] [PubMed] [Google Scholar]
- 24.Wang GL, Shi X, Salisbury E, Sun Y, Albrecht JH, Smith RG. andT imchenko NA: Cyclin D3 maintains growth-inhibitory activity of C/EBPalpha by stabilizing C/EBPalpha-cdk2 and C/EBPalpha-Brm complexes. Mol Cell Biol. 2006;26:2570–2582. doi: 10.1128/MCB.26.7.2570-2582.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Rishi L, Hannon M, Salomè M, Hasemann M, Frank AK, Campos J, Timoney J, O'Connor C, Cahill MR, Porse B, Keeshan K. Regulation of Trib2 by an E2F1-C/EBPα feedback loop in AML cell proliferation. Blood. 2014;123:2389–2400. doi: 10.1182/blood-2013-07-511683. [DOI] [PubMed] [Google Scholar]
- 26.Datta J, Majumder S, Kutay H, Motiwala T, Frankel W, Costa R, Cha HC, MacDougald OA, Jacob ST, Ghoshal K. Metallothionein expression is suppressed in primary human hepatocellular carcinomas and is mediated through inactivation of CCAAT/enhancer binding protein alpha by phosphatidylinositol 3-kinase signaling cascade. Cancer Res. 2007;67:2736–2746. doi: 10.1158/0008-5472.CAN-06-4433. [DOI] [PMC free article] [PubMed] [Google Scholar]