

Abstract
α-tomatine is a glycoalkaloid that occurs naturally in tomatoes (Lycopersicon esculentum). In the present study, the effects of α-tomatine on human myeloid leukemia HL-60 cells were investigated. Treatment of HL-60 cells with α-tomatine resulted in growth inhibition and apoptosis in a concentration-dependent manner. Tomatidine, the aglycone of tomatine had little effect on the growth and apoptosis of HL-60 cells. Growth inhibition and apoptosis induced by α-tomatine in HL-60 cells was partially abrogated by addition of cholesterol indicating that interactions between α-tomatine and cell membrane-associated cholesterol may be important in mediating the effect of α-tomatine. Activation of nuclear factor-κB by the phorbol ester, 12-O-tetradecanoylphorbol-13-acetate failed to prevent apoptosis in HL-60 cells treated with α-tomatine. In animal experiments, it was found that treatment of mice with α-tomatine inhibited the growth of HL-60 xenografts in vivo. Results from the present study indicated that α-tomatine may have useful anti-leukemia activities.
Keywords: glycoalkaloid, α-tomatine, leukemia, apoptosis, growth inhibition, cholesterol
Introduction
The solanum-steroid-alkaloids found in plants of the Solanum species are of interest as a starting material for the synthesis of steroid hormones and exhibit notable pharmaceutical and toxicological properties (1,2). α-tomatine and tomatidine (Fig. 1), occur naturally in tomatoes (Lycopersicon esculentum), belonging to the group of solanum-steroid-alkaloids. α-tomatine is a glycoalkaloid consisting of an aglycone moiety (tomatidine) and a tetrasaccharide moiety (β-lycotetraose), which is composed of two molecules of glucose, one galactose and one xylose; the four monosaccharides form a branched structure, which is attached at the C-3 position of the aglycone (Fig. 1). Although it was known that an enzyme, termed tomatinase, catalyzes the hydrolysis of tomatine into tomatidine and β-lycotetraose (3,4), the biosynthesis and metabolism of tomatine and tomatidine remain to be elucidated. Unripe green tomatoes may contain up to 500 mg/kg tomatine fresh fruit weight. The compound is partly degraded as the tomato ripens until at maturity, levels in red tomatoes are ~5 mg/kg fresh fruit weight (5). α-tomatine has also been found at high concentrations in leaves, stems and roots, suggesting that it may be important in resistance to potential pathogens (3,4).
Figure 1.

Structures of α-tomatine and tomatidine.
α-tomatine has been reported to exert toxicity against certain types of microorganisms (6–8). Previous studies have also demonstrated that α-tomatine has cytotoxic effects on insect and rat cells (9–11). In previous years, the anticancer effect of α-tomatine has been investigated. α-tomatine inhibits the growth of human cancer cells, including the HT-29 colon cancer, HepG2 liver cancer, A549 lung cancer, PC-3 prostate cancer and MCF-7 breast cancer cell lines (12–15). α-tomatine also inhibits the growth of lymphoma and leukemia cells (16,17). Although α-tomatine has been revealed to have anticancer activities in different cancer cells, the mechanisms of action and particularly the primary target(s) remain to be elucidated. In the present study, the effects and mechanisms of α-tomatine were examined in HL-60 human myeloid leukemia cells, which are widely used as a model system to investigate the effect of different anticancer agents (18,19). The present study found that α-tomatine markedly inhibited growth and induced apoptosis in HL-60 cells. α-tomatine also inhibited the in vivo growth of HL-60 cells in a mouse xenograft model.
Materials and methods
Cells and reagents
HL-60 cells were obtained from the American Type Culture Collection (Rockville, MD, USA). Tomatidine, α-tomatine and cholesterol were purchased from Sigma-Aldrich (St. Louis, MO, USA). RPMI-1640, penicillin-streptomycin, L-glutamine and fetal bovine serum (FBS) were purchased from Gibco-BRL (Grand Island, NY, USA). HL-60 cells were maintained in RPMI-1640 culture medium containing 10% FBS that was supplemented with penicillin (100 U/ml), streptomycin (100 μg/ml) and L-glutamine (300 μg/ml) (Gibco-BRL). Cultured cells were grown at 37°C in a humidified atmosphere of 5% CO2 and were passaged twice a week.
Determination of viable cells
Cell viability was determined by the trypan blue exclusion assay, as described previously (20), which was performed by mixing 80 μl of cell suspension and 20 μl of 0.4% trypan blue solution for 2 min. Blue cells were counted as dead cells and the cells that did not absorb dye were counted as live cells.
Morphological assessment of apoptotic cells
Apoptosis was determined by morphological assessment in cells stained with propidium iodide (21). Briefly, cytospin slides were prepared following each experiment and cells were fixed with acetone/methanol (1:1) for 10 min at room temperature, followed by 10 min with propidium iodide staining (1 μg/ml in phosphate-buffered saline; Gibco-BRL) and analyzed using a fluorescence microscope (Nikon Eclipse TE200; Nikon, Tokyo, Japan). Apoptotic cells were identified by classical morphological features, including nuclear condensation, cell shrinkage and formation of apoptotic bodies (21).
Nuclear extract preparation and electrophoretic mobility shift assay (EMSA)
Mini-extracts prepared from cells (1×107 cells/ml) were collected by centrifugation (13000 × g for 10 min), resuspended in hypotonic buffer and incubated on ice to obtain the nuclear pellet as described in our previous study (22). Oligonucleotides were synthesized by the DNA Synthesis and Sequencing Laboratory at the Cancer Institute of New Jersey (New Brunswick, NJ, USA). Double-stranded oligonucleotides were labeled by incubation with the Klenow enzyme fragment of DNA polymerase in the presence of 32P-dCTP, 32P-dGTP, dATP and dTTP deoxynucleoside triphosphates. Radiolabeled oligonucleotides (at least 1×108 cpm/μg) were incubated with 8 μg of nuclear protein and 3 μg of poly(dI-dC) in a total volume of 16 μl. DNA-protein complexes were analyzed by electrophoresis on 6% acrylamide gels run in 1X Tris-borate buffer (Sigma-Aldrich) (22).
HL-60 xengrafts in immunodeficient mice
Female severe combined immunodeficient (SCID) mice (6–7 weeks old) were obtained from Taconic Farms Inc. (Germantown, NY, USA). The animals were housed in sterile filter-capped microisolator cages and provided with sterilized food and water. HL-60 cells (1.0×106 cells/0.1 ml/mouse) suspended in 50% Matrigel (Collaborative Research, Bedford, MA, USA) in RPMI-1640 medium were injected subcutaneously into the right flank of the mice. After 3 weeks, mice with subcutaneous tumors were divided into two groups. One group of animals received intraperitoneal (IP) injection of vehicle, which consisted of propylene glycol, polysorbate 80, benzyl alcohol, ethanol and water (40: 0.5: 1: 10: 48.5). The other group of animals received IP injection of α-tomatine (5 mg/kg; 5 μl vehicle/g). The mice received treatment three times a week for 3 weeks. Tumor size (length × width) and body weight were measured. The animal experiment was performed under an Institutional Animal Care and Use Committee-approved protocol (#02-001; Rutgers University, Piscataway, NJ, USA).
Statistical analysis
The analysis of variance method with the Tukey-Kramer test was used for the comparison of growth inhibition and apoptosis. Student’s t-test was used to assess the differences of tumor size and body weight between the control group and the treatment group.
Results
Effects of α-tomatine on the growth of HL-60 cells
HL-60 cells were treated with different concentrations of α-tomatine for 72 h. The number of viable cells was determined at 24, 48 and 72 h using the trypan blue exclusion assay (Fig. 2). Treatment of HL-60 cells with α-tomatine resulted in a time- and concentration-dependent growth inhibition. α-tomatine at 2 and 5 μM markedly inhibited the growth of HL-60 cells. Treatment with α-tomatine (5 μM) resulted in ~98% decrease in the number of viable cells (Fig. 2A). Cytosine arabinoside (Ara-C), a commonly used chemotherapeutic drug for the treatment of myeloid leukemia, was included in the experiment. Treatment of HL-60 cells with Arc-C also resulted in growth inhibition in a time- and concentration-dependent manner (Fig. 2B). α-tomatine at 5 μM exhibited a more significant growth inhibitory effect than Ara-C. By contrast to the marked growth inhibitory effect of α-tomatine, tomatidine (the aglycone of tomatine, see Fig. 1) had little or no effect on the growth of HL-60 cells (Fig. 2C).
Figure 2.

Effects of α-tomatine, tomatidine and Ara-C on the growth of HL-60 cells. HL-60 cells were seeded at a density of 1×105 cells/ml in 35 mm tissue culture dishes and treated with various concentrations of (A) α-tomatine, (B) Ara-C and (C) tomatidine for 24, 48 and 72 h. The number of viable cells was determined using the trypan blue exclusion assay. Each value represents the mean ± standard error of the mean from three separate experiments. Ara-c, cytosine arabinoside.
Effect of α-tomatine on the apoptosis of HL-60 cells
The effects of α-tomatine and tomatidine on stimulation of apoptosis in HL-60 cells were determined using morphological assessment of apoptotic cells. Apoptotic cells were identified by classical morphological features, including nuclear condensation, cell shrinkage and formation of apoptotic bodies. Morphologically distinct apoptotic cells from representative samples are shown in Fig. 3B. Treatment of HL-60 cells with α-tomatine stimulated apoptosis in a concentration-dependent manner (Fig. 3C). A low concentration of α-tomatine (1 μM) had little stimulatory effect on HL-60 cell apoptosis. Treatment with 2 μM α-tomatine resulted in ~30% apoptotic cells and treatment with a higher concentration of α-tomatine (5 μM) resulted in ~60% apoptotic cells. By contrast, tomatidine had little or no stimulatory effect on the apoptosis of HL-60 cells (Fig. 3C).
Figure 3.

Effect of α-tomatine and tomatidine on the apoptosis of HL-60 cells. HL-60 cells were seeded at a density of 1×105 cells/ml in 35 mm tissue culture dishes and treated with various concentrations of α-tomatine and tomatidine for 48 h. Apoptosis was determined by propidium iodide staining and morphological assessment. Representative micrographs of propidium iodide-stained cells from the (A) control group and cells treated with (B) 2 μM α-tomatine (arrows indicate apoptotic cells). (C) Percentage of apoptotic cells as determined by morphological assessment in HL-60 cells treated with different concentrations of α-tomatine (grey bar) and tomatidine (open bar). Each value represents the mean ± standard error of the mean from three separate experiments.
Effects of cholesterol on α-tomatine-induced growth inhibition and apoptosis
It has been established that α-tomatine and tomatidine can form complexes with cholesterol (23,24). It was further investigated whether binding of α-tomatine with cell membrane cholesterol is required for the effect of this compound on growth inhibition and apoptosis in HL-60 cells. In these experiments, HL-60 cells were treated with α-tomatine in the presence or absence of cholesterol and cell growth and apoptosis were determined. The present study found that the addition of cholesterol significantly abrogated α-tomatine-induced growth inhibition and apoptosis in HL-60 cells. As shown in Fig. 4A, treatment with 2 μM α-tomatine decreased the number of viable cells by ~50%. The addition of an equal molar concentration of cholesterol abrogated the effect of α-tomatine. The addition of an equal molar concentration of cholesterol also abrogated the effect of tomatine on the stimulation of apoptosis in HL-60 cells (Fig. 4B).
Figure 4.
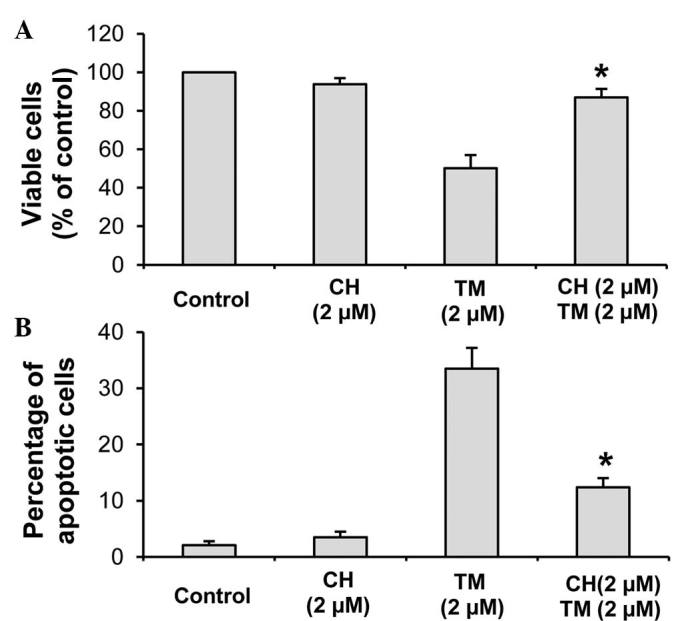
Effects of CH on TM-induced growth inhibition and apoptosis. HL-60 cells were seeded at a density of 1×105 cells/ml in 35 mm tissue culture dishes and treated with TM in the presence or absence of CH for 48 h. (A) The number of viable cells was determined by the trypan blue exclusion assay. (B) Apoptosis was determined by propidium iodide staining and morphological assessment. Each value represents the mean ± standard error of the mean from three separate experiments. Differences in the number of viable and apoptotic cells between the TM-treated group and TM+CH-treated group were analyzed by analysis of variance with the Tukey-Kramer multiple comparison test. *P<0.01 vs. TM-treated group. CH, cholesterol; TM, α-tomatine.
Effects of TPA on α-tomatine-induced growth inhibition and apoptosis
The phorbol ester TPA was demonstrated to activate the nuclear transcription factor, nuclear factor (NF)-κB in cancer cells, including leukemia cells (22,25,26). Multiple studies have indicated that NF-κB has an anti-apoptotic effect and activation of NF-κB protects cancer cells, including leukemia cells from apoptosis (27–29). It is therefore important to investigate whether pretreatment with TPA may protect HL-60 cells from α-tomatine-induced apoptosis. In initial experiments, HL-60 cells were treated with TPA and its effect on activation of NF-κB was determined using the EMSA. As shown in Fig. 5A, treatment of HL-60 cells with TPA for 1 h caused a marked activation of NF-κB. In subsequent experiments, HL-60 cells were pretreated with TPA for 1 h and then treated with tomatine. However, pretreatment with TPA failed to protect the cells from α-tomatine-induced growth inhibition (Fig. 5B) and apoptosis (Fig. 5C).
Figure 5.
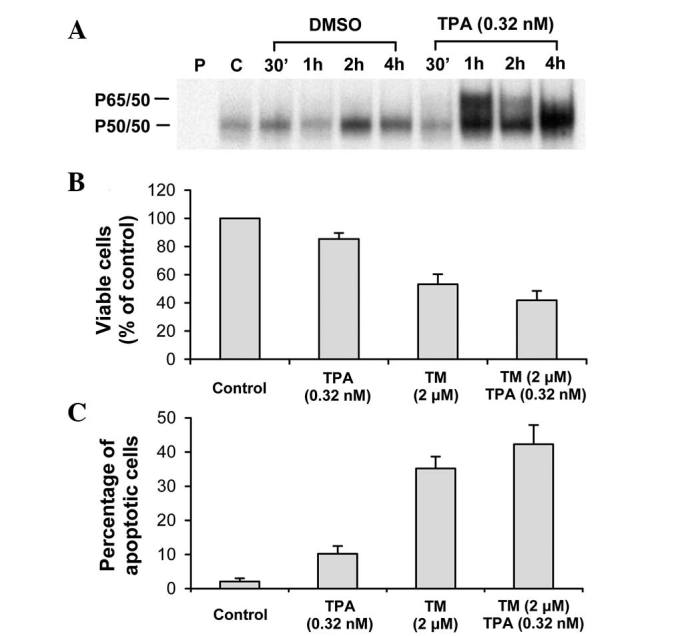
Effects of pre-treatment with TPA on TM-induced apoptosis in HL-60 cells. (A) HL-60 cells were seeded in 100 mm culture dishes and treated with 0.32 nM TPA for different time intervals. Activation of NF-κB in HL-60 cells was determined by electrophoretic mobility shift assay. P=probe only. C=control. Treatment of HL-60 cells with TPA for 1 h resulted in a marked activation of NF-κB. (B and C) HL-60 cells were seeded at a density of 1×105 cells/ml in 35 mm culture dishes. The cells were treated with TM for 48 h with or without pre-treatment with 0.32 nM TPA for 1 h. (B) The number of viable cells was determined by the trypan blue exclusion assay and expressed as a percentage of the control. (C) Apoptosis was determined by propidium iodide staining and morphological assessment. TPA, 12-O-tetradecanoylphorbol-13-acetate; TM, α-tomatine; CH, cholesterol; DMSO, dimethyl sulfoxide.
Effect of α-tomatine on the growth of HL-60 xenograft tumors
Female SCID mice with subcutaneous HL-60 xenograft tumors were injected IP with vehicle (5 μl/g body weight) or α-tomatine (5 mg/kg; 5 μl vehicle/g) three times a week for 3 weeks. As shown in Fig. 6, treatment with α-tomatine significantly inhibited the growth of HL-60 tumors. The mean ± standard error of the mean (SEM) for the percentage of the initial tumor size after 3 weeks of treatment was 475.2±62.2 for the control group and 263.9±25.7 for the α-tomatine-treated group. Statistical analysis using Student’s t-test revealed that the difference in the percentage of the initial tumor size between the control group and the α-tomatine-treated group were statistically significant (P<0.01). The mean ± SEM for the body weight (g) was 22.7±0.9 for the vehicle-treated control group and 21.8±1.0 for the α-tomatine-treated group. Statistical analysis using Student’s t-test revealed that the difference in the body weight between the control group and the treatment group was not statistically significant (P>0.05).
Figure 6.
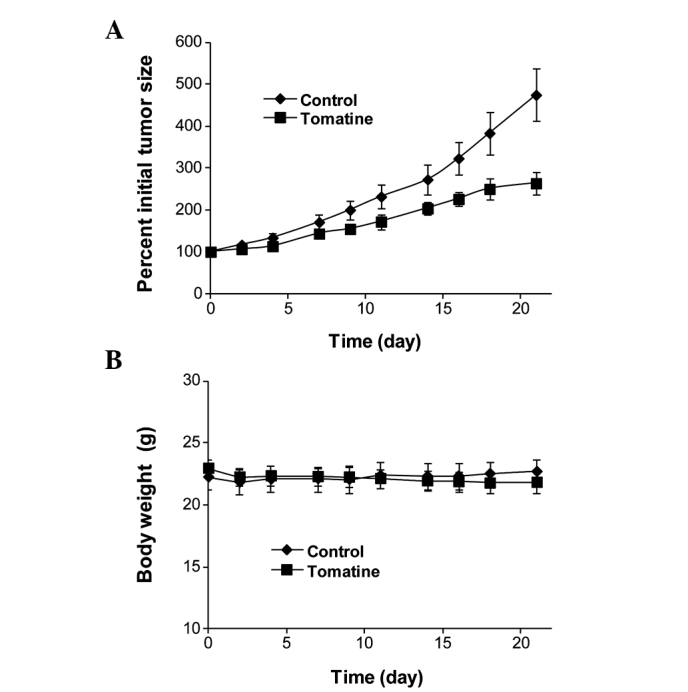
Effects of α-tomatine on (A) the growth of HL-60 xenografts and (B) body weight of SCID mice. Female SCID mice were injected subcutaneously with HL-60 cells in 50% Matrigel (1.0×106 cells/0.1 ml) suspended in RPMI medium. After 3 weeks, mice with HL-60 xenograft tumors (0.6–1.0 cm wide and 0.6–1.0 cm long) were intraperitoneally injected with vehicle (5 μl/g body weight; n=12) or with α-tomatine (5 mg/kg body weight; n=12) three times a week for 3 weeks. (A) Tumor size (length x width; cm2) was measured and expressed as a percentage of initial tumor size. (B) Body weight (g). SCID, severe combined immunodeficient.
Discussion
Glycoalkaloids are nitrogen-containing secondary plant metabolites found in numerous plants, including potatoes and tomatoes (30). The glycoalkaloid α-tomatine has been hypothesized to be involved in host-plant resistance against phytopathogens and has a variety of pharmacological and toxicological properties in animals and humans (5). α-tomatine has previously been found to exert anticancer activities (13–17,31). In the present study, the effect of α-tomatine and its aglycone, tomatidine was determined in myeloid leukemia HL-60 cells. Although it has previously been reported that α-tomatine inhibited growth and induced apoptosis in leukemia cells, the effect of tomatidine on leukemia cells has not been investigated. In the present study it was demonstrated for the first time, to the best of our knowledge, that α-tomatine, but not tomatidine inhibited growth and induced apoptosis in HL-60 cells indicating the importance of the glycone compartment of this compound. It was also found that α-tomatine was as potent as the widely used anti-leukemia drug Ara-C for inhibiting the growth and stimulating the apoptosis of HL-60 cells.
The mechanisms by which α-tomatine induces growth inhibition and apoptosis in myeloid leukemia cells remain to be elucidated. A previous study revealed that treatment with α-tomatine resulted in activation of Bak and Mcl-1s, and caused release of apoptosis inducing factor and suppression of survivin (16). Other studies demonstrated that α-tomatine inhibited the NF-κB and phosphatidyl-inositol-3-kinase/Akt signaling pathways activation in lung and prostate cancer cells (13,15). However, the primary cellular target(s) for α-tomatine and its mechanisms for modulating apoptosis-associated pathways remain to be elucidated. Previous in vitro studies revealed that α-tomatine forms a robust complex with cholesterol in aqueous media (23,24) and α-tomatine was used as a cholesterol probe (32). Keukens et al (33) demonstrated that α-tomatine and other glycoalkaloids interacted with membrane-associated cholesterol. In the present study, it was investigated whether the interaction of α-tomatine with cell membrane-associated cholesterol is essential for its effects on leukemia cells. It was found that inhibiting the interaction of α-tomatine with cell membrane-associated cholesterol by addition of equal molar concentrations of free cholesterol in the medium abrogated the effect of α-tomatine. This is the first study, to the best of our knowledge, indicating that the interaction of tomatine with cholesterol is important for mediating its effect on growth and apoptosis in leukemia cells. The interaction of α-tomatine with cholesterol and the disruptive effect on the cell membrane may be one of the mechanisms by which α-tomatine induces apoptosis in HL-60 cells. It is also possible that the formation of complexes of α-tomatine and cholesterol may modulate the responsiveness of cell membrane receptors to growth stimuli and thus decrease the growth of HL-60 cells.
To further investigate whether the glycone group is required for α-tomatine to inhibit growth and induce apoptosis, the effects of α-tomatine with its aglycone tomatidine on HL-60 cells were investigated. Although previous studies have revealed that tomatidine was able to interact and form a complex with cell membrane-associated cholesterol (23,24), it was found that tomatidine had little effect on growth inhibition and apoptosis at the doses assessed. Other studies have demonstrated that glycoalkaloids caused membrane disruption leading to the release of peroxidase previously enclosed in lipid vesicles (33,34). It was observed that α-tomatine increased the fluorescence-measured membrane permeability of frog embryos while tomatidine had a weak effect (5). α-tomatine also decreased active transport via the activity of the sodium-potassium pump in frog skin, however, tomatidine had no effect on frog skin (5). These results, together with the present findings suggest that the glycone component is important for mediating the effects of α-tomatine on growth inhibition and apoptosis. In additional studies, it was investigated whether pre-treatment with the phorbol ester TPA may protect HL-60 cells from apoptosis induced by α-tomatine. Although the present study demonstrated that treatment with TPA resulted in a marked activation of NF-κB, pre-treatment with TPA did not protect HL-60 cells from α-tomatine-induced apoptosis. This result suggests that α-tomatine may also trigger pathways that are not associated with NF-κB to induce apoptosis in HL-60 cells.
The present study also demonstrated an in vivo effect of α-tomatine. Treatment of SCID mice with IP injection of α-tomatine three times a week significantly inhibited the growth of HL-60 cells in vivo. At the dose of 5 mg/kg body weight used in the present study, α-tomatine appeared to be non toxic as no body weight loss and no abnormalities in the major organs were observed in the animals. Previous studies revealed that α-tomatine was not toxic when consumed orally by rats (35–37). When administered intravenously, it had a median lethal dose value equal to 18 mg/kg body weight (35,36). Nishie et al (38) revealed that IP injection of α-tomatine (30–100 mg/kg) in rabbits produced neither fatalities nor abnormal ECG signals. Although the plasma concentrations of α-tomatine in these studies were not known, it is reasonable to assume that blood concentration of α-tomatine at micromolar level may be achievable without severe toxicity. Further studies are required to establish the plasma levels of α-tomatine in association with its inhibitory effect on leukemia and other types of cancer in suitable animal models.
In conclusion, the present study demonstrated that α-tomatine had a marked inhibitory effect on growth and a marked stimulatory effect on apoptosis in human myeloid leukemia HL-60 cells. α-tomatine also significantly inhibited the in vivo growth of HL-60 cells in the SCID mouse xenograft model. The present study suggests that cell membrane-associated cholesterol serves as a primary target for mediating the effect of α-tomatine in leukemia cells. The present study also indicates that the glycone component is critical for α-tomatine to convey signals for growth inhibition and apoptosis. The results from the present study indicate that α-tomatine may be a candidate for the development of novel anti-leukemia agents.
Acknowledgments
The present study was supported by the Guangdong Province Leadership Grant, China National Science Foundation (grant nos. 81272452, 21102020 and 21272043) and the Rutgers Cancer Institute of New Jersey (grant no. CCSG P30-CA072720 RSD).
References
- 1.Willker W, Leibfritz D. Complete assignment and conformational studies of tomatine and tomatidine. Magn Reson Chem. 1992;30:645–650. doi: 10.1002/mrc.1260300713. [DOI] [Google Scholar]
- 2.Friedman M. Tomato glycoalkaloids: role in the plant and in the diet. J Agric Food Chem. 2002;50:5751–5780. doi: 10.1021/jf020560c. [DOI] [PubMed] [Google Scholar]
- 3.Roddick JG. The steroidal glycoalkaloid α-tomatine. Phytochemistry. 1974;13:9–25. doi: 10.1016/S0031-9422(00)91261-5. [DOI] [Google Scholar]
- 4.Lairini K, Perez-Espinosa A, Pineda M, Ruiz-Rubio M. Purification and characterization of tomatinase from Fusarium oxysporum f. sp lycopersici. Appl Environ Microbiol. 1996;62:1604–1609. doi: 10.1128/aem.62.5.1604-1609.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Blankemeyer JT, White JB, Stringer BK, Friedman M. Effect of α-tomatine and tomatidine on membrane potential of frog embryos and active transport of ions in frog skin. Food Chem Toxicol. 1997;35:639–646. doi: 10.1016/S0278-6915(97)00038-0. [DOI] [PubMed] [Google Scholar]
- 6.Jiratko J. Comparison of antifungal activity of tomatine and tomato extract. Ochrani Rostlenia. 1993;29:93–98. [Google Scholar]
- 7.Chan HT, Tam SYT. Toxicity of α-tomatine to larvae of the Mediterranean fruit fly (Diptera: Tephrididae) J Economic Entomology. 1985;78:305–307. doi: 10.1093/jee/78.2.305. [DOI] [Google Scholar]
- 8.Chu YI, Lu FM. The ovicidal effect of tomatine against deposited eggs of the diamondback moth, Plutella xylostella L. Chinese Journal of Entomology. 1992;12:213–216. [Google Scholar]
- 9.Weissenberg M, Levy A, Svoboda JA, Ishaaya I. The effect of some Solanum steroidal alkaloids and glycoalkaloids on larvae of the red flour beetle, Tribolium castaneum, and the tobacco hornworm, Manduca sexta. Phytochemistry. 1998;47:203–209. doi: 10.1016/S0031-9422(97)00565-7. [DOI] [PubMed] [Google Scholar]
- 10.Chataing B, Concepcion JL, Lobaton R, Usubillaga A. Inhibition of Trypanosoma cruzi growth in vitro by Solanum alkaloids: a comparison with ketoconazole. Planta Med. 1998;64:31–36. doi: 10.1055/s-2006-957361. [DOI] [PubMed] [Google Scholar]
- 11.Bergers Lee WW, Alink GM. Toxic effect of the glycoalkaloids solanine and tomatine on cultured neonatal rat heart cells. Toxicol Lett. 1980;6:29–32. doi: 10.1016/0378-4274(80)90098-3. [DOI] [PubMed] [Google Scholar]
- 12.Lee KR, Kozukue N, Han JS, et al. Glycoalkaloids and metabolites inhibit the growth of human colon (HT29) and liver (HepG2) cancer cells. J Agric Food Chem. 2004;52:2832–2839. doi: 10.1021/jf030526d. [DOI] [PubMed] [Google Scholar]
- 13.Shieh JM, Cheng TH, Shi MD, et al. α-Tomatine suppresses invasion and migration of human non-small cell lung cancer NCI-H460 cells through inactivating FAK/PI3K/Akt signaling pathway and reducing binding activity of NF-κB. Cell Biochem Biophys. 2011;60:297–310. doi: 10.1007/s12013-011-9152-1. [DOI] [PubMed] [Google Scholar]
- 14.Shih YW, Shieh JM, Wu PF, et al. α-tomatine inactivates PI3K/Akt and ERK signaling pathways in human lung adenocarcinoma A549 cells: effect on metastasis. Food Chem Toxicol. 2009;47:1985–1995. doi: 10.1016/j.fct.2009.05.011. [DOI] [PubMed] [Google Scholar]
- 15.Lee ST, Wong PF, Cheah SC, Mustafa MR. α-tomatine induces apoptosis and inhibits nuclear factor-κB activation on human prostatic adenocarcinoma PC-3 cells. PLoS One. 2011;6:e18915. doi: 10.1371/journal.pone.0018915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Chao MW, Chen CH, Chang YL, et al. α-Tomatine-mediated anti-cancer activity in vitro and in vivo through cell cycle- and caspase-independent pathways. PLoS One. 2012;7:e44039. doi: 10.1371/journal.pone.0044093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Yang YW, Wu CA, Morrow WJ. The apoptotic and necrotic effects of tomatine adjuvant. Vaccine. 2004;22:2316–2327. doi: 10.1016/j.vaccine.2003.10.036. [DOI] [PubMed] [Google Scholar]
- 18.Collins SJ. The HL-60 promyelocytic leukemia cell line: proliferation, differentiation, and cellular oncogene expression. Blood. 1987;70:1233–1244. [PubMed] [Google Scholar]
- 19.Semizarov D, Glesne D, Laouar A, et al. A lineage-specific protein kinase crucial for myeloid maturation. Proc Natl Acad Sci USA. 1998;95:15412–15417. doi: 10.1073/pnas.95.26.15412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Zheng X, Chang RL, Cui XX, et al. Synergistic effects of clinically achievable concentrations of 12-O-tetradecanoylphorbol-13-acetate in combination with all-trans retinoic acid, 1α,25 dihydroxyvitamin D3, and sodium butyrate on differentiation in HL-60 cells. Oncol Res. 2000;12:419–427. [PubMed] [Google Scholar]
- 21.Zheng X, Cui XX, Khor TO, et al. Inhibitory effect of a γ-tocopherol-rich mixture of tocopherols on the formation and growth of LNCaP prostate tumors in immunodeficient mice. Cancers (Basel) 2011;3:3762–3772. doi: 10.3390/cancers3043762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Hansson A, Marín YE, Suh J, et al. Enhancement of TPA-induced growth inhibition and apoptosis in myeloid leukemia cells by BAY 11–7082, an NF-κB inhibitor. Int J Oncol. 2005;27:941–948. [PubMed] [Google Scholar]
- 23.Micich TJ. Behavior of polymer-supported tomatine toward cholesterol in the presence and absence of butter oil. J Agricu Food Chem. 1993;39:1610–1613. doi: 10.1021/jf00009a014. [DOI] [Google Scholar]
- 24.Roddick JG. Complex formation between solanaceous steroidal glycoalkaloids and free sterols in vitro. Phytochemistry. 1979;18:1467–1470. doi: 10.1016/S0031-9422(00)98476-0. [DOI] [Google Scholar]
- 25.Maiti A, Cuendet M, Kondratyuk T, et al. Synthesis and cancer chemopreventive activity of zapotin, a natural product from Casimiroa edulis. J Med Chem. 2007;50:350–355. doi: 10.1021/jm060915+. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kamiya T, Makino J, Hara H, et al. Extracellular-superoxide dismutase expression during monocytic differentiation of U937 cells. J Cell Biochem. 2011;112:244–255. doi: 10.1002/jcb.22917. [DOI] [PubMed] [Google Scholar]
- 27.Bortul R, Tazzari PL, Cappellini A, et al. Constitutively active Akt1 protects HL60 leukemia cells from TRAIL-induced apoptosis through a mechanism involving NF-κB activation and cFLIP(L) up-regulation. Leukemia. 2003;17:379–389. doi: 10.1038/sj.leu.2402793. [DOI] [PubMed] [Google Scholar]
- 28.Baldwin AS. Control of oncogenesis and cancer therapy resistance by the transcription factor NF-κB. J Clin Invest. 2001;107:241–246. doi: 10.1172/JCI11991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Cuní S, Pérez-Aciego P, Pérez-Chacón G, et al. A sustained activation of PI3K/NF-κB pathway is critical for the survival of chronic lymphocytic leukemia B cells. Leukemia. 2004;18:1391–1400. doi: 10.1038/sj.leu.2403398. [DOI] [PubMed] [Google Scholar]
- 30.Friedman M, McDonald GM. Potato glycoalkaloids: chemistry, analysis, safety, and plant physiology. Crit Rev Plant Sci. 1997;16:55–132. doi: 10.1080/07352689709701946. [DOI] [Google Scholar]
- 31.Friedman M. Anticarcinogenic, cardioprotective, and other health benefits of tomato compounds lycopene, α-tomatine, and tomatidine in pure form and in fresh and processed tomatoes. J Agric Food Chem. 2013;61:9534–9550. doi: 10.1021/jf402654e. [DOI] [PubMed] [Google Scholar]
- 32.Severs NJ, Simms HL. Failure of filipin to detect cholesterol-rich domains in smooth muscle plasma membane. Nature. 1983;303:637–638. doi: 10.1038/303637a0. [DOI] [PubMed] [Google Scholar]
- 33.Keukens EAJ, de Vrije T, van den Boom C, et al. Molecular basis of glycoalkaloid induced membrane disruption. Biochim Biophys Acta. 1995;1240:216–228. doi: 10.1016/0005-2736(95)00186-7. [DOI] [PubMed] [Google Scholar]
- 34.Roddick JG, Rijnenberg AL, Osman SF. Synergistic interaction between potato glycoalkaloids α-solanine and α-chaconine in relation to destabilization of cell membranes: Ecological implications. J Chem Ecol. 1988;14:889–902. doi: 10.1007/BF01018781. [DOI] [PubMed] [Google Scholar]
- 35.Cayen MN. Effect of dietary tomatine on cholesterol metabolism in the rat. J Lipid Res. 1971;12:482–490. [PubMed] [Google Scholar]
- 36.Friedman M, Henika PR, Mackey BE. Feeding of potato, tomato and eggplant alkaloids affects food consumption and body and liver weights in mice. J Nutr. 1996;126:989–999. doi: 10.1093/jn/126.4.989. [DOI] [PubMed] [Google Scholar]
- 37.Wilson RH, Poley GW, De Eds F. Some pharmacologic and toxicologic properties of tomatine and its derivatives. Toxicol Appl Pharmacol. 1961;3:39–48. doi: 10.1016/0041-008X(61)90006-0. [DOI] [PubMed] [Google Scholar]
- 38.Nishie K, Norred WP, Swain AP. Pharmacology and toxicology of chaconine and tomatine. Res Comm Chem Pathol Pharm. 1975;12:657–668. [PubMed] [Google Scholar]