

Researchers uncover a large variation in the lifetime migratory decisions of young white storks.
Keywords: Animal movement, migration costs, energy expenditure, White storks, high-resolution GPS, acceleration, lifetime tracking, population comparison
Abstract
Annual migratory movements can range from a few tens to thousands of kilometers, creating unique energetic requirements for each specific species and journey. Even within the same species, migration costs can vary largely because of flexible, opportunistic life history strategies. We uncover the large extent of variation in the lifetime migratory decisions of young white storks originating from eight populations. Not only did juvenile storks differ in their geographically distinct wintering locations, their diverse migration patterns also affected the amount of energy individuals invested for locomotion during the first months of their life. Overwintering in areas with higher human population reduced the stork’s overall energy expenditure because of shorter daily foraging trips, closer wintering grounds, or a complete suppression of migration. Because migrants can change ecological processes in several distinct communities simultaneously, understanding their life history decisions helps not only to protect migratory species but also to conserve stable ecosystems.
INTRODUCTION
The seasonal long-distance movements of birds have fascinated humans for hundreds of years. Empirical and theoretical research on these often intercontinental journeys has increased our understanding of behavioral, ecological, and evolutionary aspects of migration (1). The recent advances and miniaturization of tracking technology provide new possibilities for following a migrant throughout its annual cycle or even throughout its life with extreme detail. High-resolution GPS (Global Positioning System) together with three-axial acceleration allows us now to monitor the behavior of free-flying birds almost continuously (2). Beyond a description of flight behavior, the accelerometry technique has been used for estimating energy expenditure in a variety of free-living animals (3–5), offering new opportunities to explore the physiological basis of life histories. One of the main goals of this mechanistic approach to ecology is to understand how animals interact with their physical environment. It has become apparent that movement decisions depend not only on the internal state and capacities of an organism but also on biotic and abiotic environmental factors (6). Therefore, deciphering mechanisms that govern animal movement—especially those that provide the link between energy expenditure and survival—is of fundamental relevance to animal ecology.
One frequently studied migrant is the white stork, Ciconia ciconia. Numerous studies provide detailed knowledge on their population dynamics, as well as their breeding and migratory behavior (7, 8). However, even in such a popular model, questions concerning the energetics and costs of migration are largely unanswered. Here, we used biologging technology to get a deep insight into the energetics and timing of stork migration. White storks breed from Europe to Northwest Africa and Western Asia. Because migration varies greatly between the different populations, storks provide a suitable system for exploring the costs and implications of migratory decisions. Similar to most raptors, white storks are soaring migrants that rarely use flapping flight (9). They gain altitude using thermal uplift (that is, columns of rising air created by solar radiation) and then move forward through gliding. Soaring flight is energetically cheaper than flapping, allowing birds to travel hundreds of kilometers with minimal energy expenditure (10, 11). Soaring species actively choose their migration routes and times to experience stronger updrafts (12) and minimize their cost of transport (13).
Here, using high-resolution GPS devices with three-dimensional (3D) acceleration sensors, we compared the migratory behavior of juvenile storks from eight populations to contrast the use of preferential environmental conditions by birds from different origins. Very little is known about the subpopulations at the edge of the species’ distribution range (for example, Armenia, Uzbekistan, and Tunisia), because most of the tracking effort on white storks has focused on adults of the central European population. Lifetime trajectories allowed us to record and evaluate the movement costs of a white stork’s life in light of its decision-making from the earliest stages of its independent life (that is, as soon as it leaves the nest). We continuously monitor the youngster’s movements throughout the first 5 months of its migratory journey using GPS and accelerometers. Because many juveniles do not survive this trip (14, 15), those first 5 months can be seen as a representation of a juvenile stork’s lifetime movement costs. Time and energy are key currencies in every migrant’s life (10, 16); therefore, choosing an optimal migration strategy seems of utmost importance. On the basis of these assumptions, we predicted that migratory decisions such as timing, routes, and destinations are major determinants of the lifetime movement costs.
RESULTS
The migratory behavior (timing, routes, lengths, and destinations) varied drastically among the stork populations (Table 1 and Fig. 1A). Migration initiation date (of those storks that left their natal grounds) differed strongly between the different populations [F6,50 = 16.83, n = 57, P < 0.001, adjusted R2 = 0.63, linear model (LM); Fig. 1, B and C; see table S1 and Materials and Methods for details]. It spanned, on average, 36 days (±4 days SD) between July 27th (average for Spanish storks) and September 1st (average for Armenian storks) and was unrelated to the maximum distance reached (here, we examined only birds that survived throughout the first 5 months after departing; F1,27 = 0.11, n = 29, P = 0.75, adjusted R2 = −0.03, LM; Fig. 1C). The seven populations that migrated could be distinguished by their maximum distance reached (F6,22 = 7.02, n = 29, P < 0.001, adjusted R2 = 0.56, LM; see table S2): storks that migrated along the well-described Eastern flyway (that is, Russian, Polish, and Greek storks) traveled as far as South Africa, whereas birds from Spain, Tunisia, and southwest Germany did not move south of the Sahel zone. Armenian storks traveled, on average, only 982.2 km (±195 km SD) toward the Persian Gulf, and Uzbek birds turned out to be residents, wintering within a radius of about 150 km from their natal sites around Tashkent (see Fig. 1, A and C). The sedentary behavior of the latter population was confirmed by 11 additional juveniles tagged the following season (2014).
Table 1. Locations and sample sizes of the different study populations.
Central GPS location of study sites; number of birds tagged (in parenthesis: number of birds with high-resolution data, that is, GPS and acceleration data taken every 5 min); number of birds still alive after 5 and 12 months (in parenthesis: percentage of total number tagged).
| Population |
Central GPS coordinates of study site |
No. of birds | |||
| Longitude | Latitude | Tagged (high resolution) | Alive after 5 months | Alive after 12 months | |
| Armenia | 44°31′1.2″ | 40°12′0″ | 8 (1) | 2 (25%) | 2 (25%) |
| Greece | 26°10′4.8″ | 40°53′27.6″ | 10 (1) | 5 (50%) | 2 (20%) |
| Poland | 15°30′7.2″ | 51°56′13.2″ | 4 (2) | 2 (50%) | 2 (50%) |
| Russia | 37°23′31.2″ | 55°38′49.2″ | 10 (4) | 7 (70%) | 4 (40%) |
| Spain | −6°1′33.6″ | 37°21′25.2″ | 11 (1) | 3 (27%) | 1 (9%) |
| Southwest Germany | 8°56′2.4″ | 47° 45′ 10.8″ | 12 (5) | 6 (46%) | 5 (38%) |
| Tunisia | 10°15′39.6″ | 36° 48′ 32.4″ | 9 (2) | 4 (44%) | 2 (22%) |
| Uzbekistan | 69°2′16.8″ | 40° 59′ 45.6″ | 6 (3) | 4 (67%) | 3 (50%) |
| TOTAL | — | — | 70 (19) | 33 (47%) | 21 (30%) |
Fig. 1. Migratory behavior of juveniles from eight different populations.

(A) Migration paths of 62 individuals tracked with GPS/GSM (Global System for Mobile Communications) (eight individuals died before migrating). Maps depicted are OpenStreetMap images (www.openstreetmap.org/copyright; https://creativecommons.org/licenses/by-sa/2.0/) accessed through the R package OpenStreetMap. (B) Departure date of the studied populations. Color scale indicates departure dates (white indicating no departure). (C) Departure date as a function of maximum distance reached (each color represents one population). Dots in the light gray–shaded area represent individuals that left their natal grounds but survived for less than 150 days. SW, southwest.
Hereafter, to explore their movement activity in detail, we focused on birds that yielded high-resolution data (n = 19; Table 1 and Materials and Methods). With the aim of assessing energetic costs for movement, we obtained a quantitative measure of animal activity from three-axial acceleration data—overall dynamic body acceleration (ODBA). This measure has been used as a proxy for energy expenditure in a variety of studies (3–5). The overall costs of the individuals’ first 5 months of life following departure (total ODBA) were calculated as the sum of daily activity values (ODBA). On the basis of total distance and total ODBA, we detected three different migratory patterns. First, birds that migrated but did not cross 33°N (that is, they overwintered north of the Sahara; n = 11) flew a total distance of 4867 ± 230 km (mean ± SD) with a total ODBA of 352.6 ± 18.8g (mean ± SD). This pattern was displayed by some individuals born in Germany and all Armenian storks. A second pattern was followed by storks that overwinter south of the Sahara (for example, the “typical” eastern migrants born in Poland, Russia, and Greece; Tunisian storks and some western migrants from southwest Germany; N = 5). They traveled 16,550 ± 3716 km (mean ± SD; including Sahara crossings) and showed a total ODBA of 473 ± 24.3g (mean ± SD). Despite the total distance being 3.4 times larger in the latter group compared to that of storks wintering north of the Sahara desert, the total ODBA was only 1.3 times larger, revealing a nonlinear relationship between movement activity (Y) and distance traveled (X) (Y = 387.2 + 1.344 × 10−11 X3; F1,14 = 23.7, n = 16, P < 0.001, LM, adjusted R2 = 0.60; Fig. 2A). The third strategy was displayed by storks from Uzbekistan (n = 3). Although data from these birds were partly interpolated (see Materials and Methods) and thus not included in the previous analyses, they revealed a unique pattern that separates Uzbek storks from the other populations. The maximum distance from the natal site was, on average, only 107.6 km (±32.5 km SD), but the total distance flown added up to 5486.5 ± 993 km (mean ± SD), similar to migratory storks wintering north of the Sahara. However, the total ODBA of Uzbek storks was 444.0 ± 37.5g (mean ± SD) and thus more in agreement with the birds crossing the Sahara and traveling to Africa along the Eastern route (Fig. 2A).
Fig. 2. Activity measures of juvenile white storks.
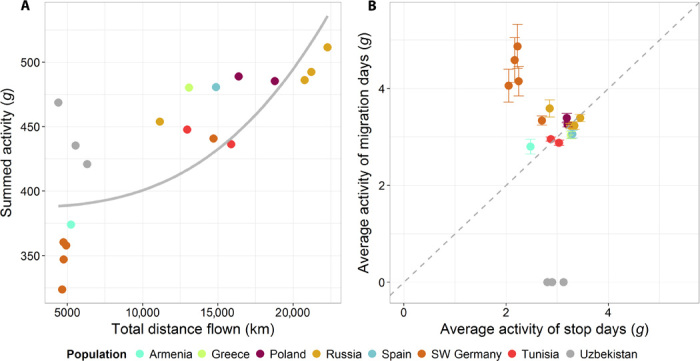
(A) Summed activity (ODBA) of the first 5 months of a juvenile’s life as a function of total distance flown during the same time. The solid gray line represents the third-order polynomial regression. The best-fitting regression is provided by the equation Y = 387.2 + 1.344 × 10−11 X3. (B) Average activity (ODBA) of a migration day in relation to average activity of a stopover day. Color represents the different populations. Gray dashed line is a reference line.
Next, to examine the three distinct migration patterns in more detail, we looked at the movement activity (ODBA) during flight and stopover days separately (Fig. 2B). A general linear mixed model (GLMM; gamma distribution with individual as random factor; n = 16; R2 = 0.71) revealed that ODBA on flight days was influenced significantly by the bird’s location (that is, a categorical variable that indicates whether it was north or south of 33°N; Wald χ2 = 68.3, P < 0.001) and an interaction between this location and thermal uplift (12) (Wald χ2 = 26.4, P < 0.001; Fig. 3; see table S3 and Materials and Methods for details). Thus, the more favorable flight conditions in Africa allowed birds to reach foraging locations further south while investing a minimum amount of energy for flight. In contrast, the storks that remained within Europe, the Middle East, and Northern Africa displayed fewer flight days and experienced more strenuous flight conditions, which led to a disproportional increase in their daily ODBA values.
Fig. 3. Activity and thermal uplift.
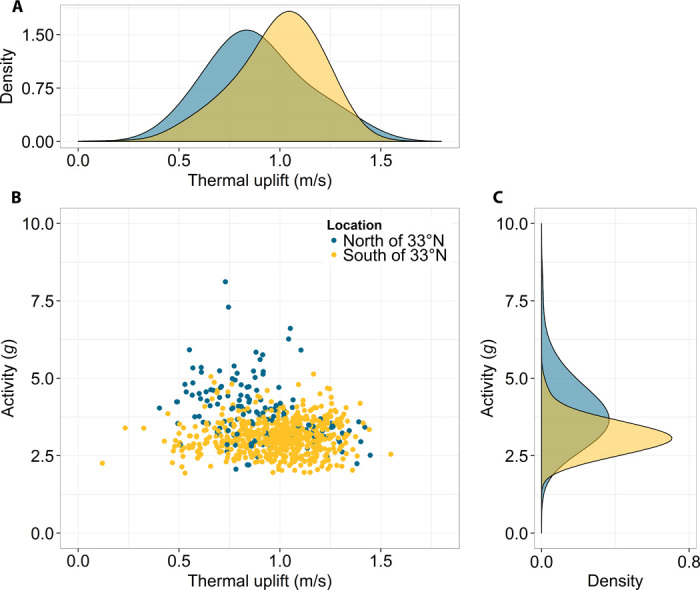
Activity (sum of daily ODBA values) as a function of thermal uplift (mean of daily thermal uplift), during flight days, depending on the location of the bird (B). Blue and light brown correspond to Europe/Middle East (north of 33°N) and Africa (south of 33°N), respectively. Plots on the top (A) and the side (C) are the density histograms of thermal uplift (top) and ODBA (right).
Nevertheless, these birds compensated for the short and expensive journey by spending very little energy for locomotion during nonmigration days. Those birds that migrated south of 33°N moved 43.7 ± 27.4 km (mean ± SD) each day. In contrast, birds remaining in Europe, the Middle East, or Northern Africa reduced their daily distance to 19.6 km (±18.8 km SD). Hence, on stopover days, storks varied in their daily movement distances, depending on whether they overwintered in Uzbekistan, south or north of the Sahara (Wald χ2 = 63.2, df = 2, n = 19, P < 0.001, R2 = 0.61, GLMM; see table S4 and Materials and Methods). Although Uzbek birds did not migrate, they moved more (locally) than did conspecifics remaining within Europe and Northern Africa (Tukey post hoc test, z = −7.9, P < 0.001). Linked to this result, the most striking difference between the groups was the choice of habitat during nonmigration days. Using a GLMM (gamma distribution with individual as random factor; R2 = 0.71), we found that movement activity exhibited during nonmigration days was significantly affected by human population density (Wald χ2 = 118.44; n = 19; P < 0.001) and vegetation cover (Wald χ2 = 153.43; n = 19; P < 0.001). The storks wintering north of the Sahara mainly dwelled in areas with higher human population density and low levels of vegetation cover (for example, crops, grass, and shrubs), whereas long-distance flyers encountered regions with very low human density and higher values of vegetation cover (Fig. 4). This indicates that white storks spend more energy overwintering in African rural areas compared to densely populated regions of Europe.
Fig. 4. Activity in relation to human population density and vegetation cover.

Daily activity (sum of hourly ODBA averages) as a function of human population density (mean of the day) (B) and vegetation cover of shrubs and low-lying bushes [“low vegetation cover,” European Centre for Medium-Range Weather Forecasts (ECMWF), mean of the day] (D). Blue and light brown correspond to birds that overwintered north and south of 33°N, respectively. Plots on the top and the side are the density histograms of population density (A), vegetation cover (C), and ODBA (E).
DISCUSSION
Our results highlight the large extent of variation in migration within a single, widely distributed, soaring food opportunist. The eight stork populations differed in their geographically distinct overwintering grounds, migration routes, and wintering locations, with significant consequences on the amount of energy invested during the first months of life. Traveling from their respective natal grounds through Europe was more demanding compared to flying on the African continent. The activity measure ODBA indicates a reduction of 15% of energy used for locomotion when flying over Africa. Although flying seems less costly, this might not equate with energy savings of 15%, because costs for thermo- and hydroregulation might be much higher in hot climates. Although it was previously known that the speed of soaring migrants can differ between geographical regions along the migration path (17–20), our data provide quantitative evidence for regional differences in movement costs. In addition to favorable weather effects, the decreased flight costs in Africa could be potentially affected by other factors, such as improved flight performance through learning (14) or, alternatively, increased flock sizes (21), which provide more social cues (22) to find and use updrafts more efficiently. Although a high juvenile mortality (7) (Table 1) hinders longitudinal assessments of migratory behavior throughout an individual’s life, a long-term study on black kites recently described the ontogeny of migratory performance, revealing within-individual improvements and selective mortality (14). The continuation of lifetime tracking, together with detailed analyses on social interactions and individual improvements, will eventually allow us to understand the role of social and asocial components in the development of flight abilities, unraveling additional links between movement strategies and life history decisions.
The decision to migrate, distance moved, and final destination were revealed here as main factors shaping the lifetime movement costs in juvenile migrants. The lack of relationship between departure date and final distance reached indicates that departure time may not be determined by the destination but by the geographical constraints encountered en route (for example, high mountains or water bodies). Finding the most suitable meteorological conditions to cross these barriers may strongly influence the timing of departure (23). Further, our results showed that many storks deviate from the well-described eastern and western routes. Birds from Russia, Poland, and Greece displayed the most traditional routes along the eastern flyway, through the Sahel region of East Africa all the way down to South Africa. Although it is documented that an increasing number of white storks overwinter on the Iberian Peninsula (24), extensively feeding on landfill sites, all of the Spanish juveniles migrated across the Sahara desert to the western Sahel. In contrast, storks from southwest Germany in this study were clearly affected by these human-induced changes, and four of the six birds that survived for at least 5 months overwintered on garbage dumps in Northern Morocco instead of migrating to the Sahel zone. Our results showed that birds that stayed in Morocco moved less during stopover days and had lower ODBA values compared to birds wintering south of the Sahara. Hence, feeding on anthropogenic food sources such as landfills seems to be beneficial because birds can shorten their migration distance and decrease their daily energy expenditure. These changes may result in higher survival and fitness (25), potentially leading to rapid microevolutionary changes in migratory patterns (26).
During the past decades, it has been documented that numerous migratory species have shortened their migration distance or turned into residents in response to anthropogenic environmental change (27–29). Although previous data are lacking, we hypothesize that human-induced supplementary feeding [that is, feeding on fish farms (I. Pokrovsky, personal observation)] could have driven the suppression of migratory behavior of Uzbek storks, a subspecies (C. ciconia asiatica) traditionally thought to overwinter in Afghanistan and Pakistan (30). Migratory animals can have fundamental effects on ecosystems by altering ecological networks, influencing pest control and pollination, or affecting infectious disease dynamics (31, 32). Understanding how human actions alter migratory patterns may be the key not only to protecting migratory species but also to maintaining diverse and stable ecosystems.
MATERIALS AND METHODS
Data set
To track an individual’s movements throughout its entire lifetime, we equipped juvenile white storks 1 week before fledging with high-resolution, solar GSM-GPS-ACC loggers (e-obs GmbH). We focused our tracking efforts on eight different white stork populations, thereby covering almost the complete breeding range of the species (for details on locations and sample size of the populations, see Table 1). The data used in this study are available on Movebank [movebank.org, study name “MPIO white stork lifetime tracking data (2013-2014)”] and are published in the Movebank Data Repository with doi:10.5441/001/1.78152p3q (33).
The transmitters (weight, 54 g) were attached using a Teflon-nylon harness (weight, ~12 g). The total weight of the transmitters and the harness was 66 g, corresponding to approximately 2% of the mean body mass of white storks (34). We recorded GPS locations and 3D body acceleration for 18 hours/day (between 4:00 and 22:00 local time at the natal grounds). GPS positions had a positional accuracy of ±3.6 m (that is, when stationary, 50% of fixes remain within a radius of 3.6 m within 24 hours) and were recorded every 5 min. Missing fixes due to low battery power or bad satellite coverage were interpolated using linear approximation (3% of all data points). Three-axial body acceleration was measured every 5 min for 3.8 s at 10.54 Hz (that is, 40 data points per axis and 120 data points per ACC burst). Data were stored onboard the device until they were downloaded via an ultrahigh-frequency radio link from a distance of approximately 300 m (35). In addition, all loggers had a GSM unit that sent two short text messages [SMS (Short Message Service)] per day (limited to areas of cellular coverage), each containing five GPS locations recorded at 1-hour intervals, providing low-resolution positional data. Raw acceleration data were converted from millivolts into gravitational acceleration (g) (rate of change of velocity; 1g = 9.81 ms−2) using tag-specific calibration values. The three signals were individually smoothed using running means over 3.8 s. Next, for each axis, the smoothed data were subtracted from the corresponding unsmoothed data; the sum of all three axes provided a value of ODBA experienced by the animal (5, 36).
Movement parameters from GSM data
We used low-resolution GSM data to identify individual departure dates from the natal grounds. Because storks from all populations traveled southward, we defined departure date as the first day a bird reduced its latitude by more than 0.5° (that is, approximately 60 km southward). For birds that survived the following 5 months, we also calculated the maximum distance reached from the nest site.
Movement parameters from high-resolution data
To compare the energy expenditure of different migration strategies, we analyzed the high-resolution data (GPS + acceleration) of storks that survived for at least 150 days after departure. Total migration costs were measured using the sum of all daily ODBA values (that is, 150 observations per bird), each representing the sum of hourly averages. Because most juveniles die within these 150 days, these total costs can be seen as a representation of the costs of a juvenile’s stork life. Total migration distance was defined as the sum of all distances between consecutive GPS fixes. To compare migration and stopover days, we defined the latter as days when storks traveled less than 100 km/day. Because Uzbek storks remained at their natal grounds, the colder, less sunny weather caused low battery power and intermittent interruptions in the data recordings in three birds (47, 36, and 19 days). To compare these birds to all other individuals, we obtained estimates of energy expenditure, distance traveled, and days through interpolation using randomly generated ODBA and distance values in the range of recordings from previous days.
Environmental data annotation
We used the Environmental Data Automated Track Annotation (Env-DATA) system (37) to annotate the tracking data with ambient atmospheric observations and underlying landscape information. Env-DATA is a service on Movebank (38), an open, online system for management, archiving, analysis, and sharing of animal movement data. The human population density was provided by the Socioeconomic Data and Applications Center at Columbia University (http://sedac.ciesin.columbia.edu/data/collection/gpw-v3). The ECMWF global reanalysis data set was accessed to annotate the tracks with low vegetation cover (for example, crops, grass, tundra, marshes, and shrubs) and thermal uplift velocity. The estimates of thermal uplift velocity provided by Env-DATA are calculated using estimates of temperature, relative humidity, surface pressure, boundary layer height, and instantaneous moisture and surface heat fluxes using ECMWF data [for details on the equations used, see Bohrer et al. (12)]. To annotate the movement tracks, Movebank uses the data values at locations and times provided by the environmental data set to calculate an estimated value of the variable for the location and time of each animal location (interpolation). Records from all the environmental data sets were annotated to the tracks using inverse weighted distance interpolation in space and time [for further details, see Dodge et al. (37)].
Statistical analysis
We examined whether departure date and maximum distance reached differed between the populations (categorical variable) using LMs. Also, of those birds that yielded high-resolution data and left their natal grounds (n = 16), we related movement activity to total distance traveled using an LM.
When analyzing the effect of thermal uplift, population density, and vegetation cover on daily activity (ODBA), we used GLMMs. GLMMs allow cases where the response variable is determined by both random and fixed effects. We fitted bird identity as a random effect because of repeated sampling of the same individual. ODBA was not normally distributed, necessitating the use of a gamma distribution. The significance of the explanatory terms, the fixed effects, was assessed by their Wald statistics (distributed as χ2) for each term when fitted last in the model. The effect size was indicated by marginal R2, as suggested by Nakagawa and Schielzeth (39). All statistical analyses were performed using R and the R package lme4 (40, 41). Post hoc tests with Tukey style contrasts were used for pairwise comparisons of treatment means, using the glht function in the multcomp package.
Supplementary Material
Acknowledgments
We thank all the people who helped during fieldwork, especially W. Schäfle, F. Torres, S. Palacios, S. Cabezas, J. De la Puente, C. Shamina, M. Shamin, Y. Galchyonkov, A. Stupnikov, V. Sudarev, T. Qhobela, H. Dlensi, S. Ben Mahrez, W. Bashimam, J. Tahri, E. Stets, A. Athanasiadis, V. Sirkelidis, S. Feldman, and H. Eggers. Thanks to the Municipality of Alexandroupolis, Greece, and the Hellenic Public Power Corporation. Special thanks to B. Keeves for help with retrieving GPS devices. We thank W. Heidrich and F. Kümmeth (e-obs) for help with logger programming and S. Davidson for setting up the movebank.org data repository. Funding: A.F. was supported by the German Aerospace Center (DLR); J.B. was supported by projects i-link0564, CGL2012-32544, and 511/2012 from the Spanish Consejo Superior de Investigaciones Científicas, National Parks, National Ministries, and FEDER (European Regional Development Fund) funds. We acknowledge the support of Deutsch-Israelische Projektkooperation grants (Deutsche Forschungsgemeinschaft) NA 846/1-1 and WI 3576/1-1. Author contributions: A.F., W.F., M.W., and R.N. conceived the idea; A.F., W.F., and M.W. wrote the manuscript; A.F., J.B., W.F., and M.W. edited the manuscript; A.F., W.F., J.B., I.P., M.K., M.M., K.A., I.F., E.M., L.J., H.A., C.F.-A., S.R., T.M.M., and M.W. collected the data; and A.F. analyzed the data. Competing interests: The authors declare that they have no competing interests. Data and materials availability: All data needed to evaluate the conclusions in the paper are present in the paper and/or the Supplementary Materials. Additional data related to this paper may be requested from the authors. The data used in this study are available on Movebank [movebank.org, study name “MPIO white stork lifetime tracking data (2013-2014)”] and are published in the Movebank Data Repository, with doi:10.5441/001/1.78152p3q (33).
SUPPLEMENTARY MATERIALS
Supplementary material for this article is available at http://advances.sciencemag.org/cgi/content/full/2/1/e1500931/DC1
Table S1. Results of linear regression models concerning the effects of population on departure date.
Table S2. Results of linear regression models concerning the effects of population on maximum distance reached.
Table S3. GLMM model results for animal activity (ODBA) showing the effects of the main predictors. North of 33°N was incorporated in the intercept.
Table S4. GLMM model results concerning the effect of daily distance moved on the location of the wintering grounds.
Table S5. GLMM model results for animal activity (ODBA) showing the effects of human population density (log) and vegetation cover (centered).
REFERENCES AND NOTES
- 1.I. Newton, The Migration Ecology of Birds (Academic Press, London, 2008). [Google Scholar]
- 2.Kays R., Crofoot M. C., Jetz W., Wikelski M., Terrestrial animal tracking as an eye on life and planet. Science 348, aaa2478 (2015). [DOI] [PubMed] [Google Scholar]
- 3.Halsey L. G., Shepard E. L. C., Wilson R. P., Assessing the development and application of the accelerometry technique for estimating energy expenditure. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 158, 305–314 (2011). [DOI] [PubMed] [Google Scholar]
- 4.Williams T. M., Wolfe L., Davis T., Kendall T., Richter B., Wang Y., Bryce C., Elkaim G. H., Wilmers C. C., Instantaneous energetics of puma kills reveal advantage of felid sneak attacks. Science 346, 81–85 (2014). [DOI] [PubMed] [Google Scholar]
- 5.Wilson R. P., White C. R., Quintana F., Halsey L. G., Liebsch N., Martin G. R., Butler P. J., Moving towards acceleration for estimates of activity-specific metabolic rate in free-living animals: The case of the cormorant. J. Anim. Ecol. 75, 1081–1090 (2006). [DOI] [PubMed] [Google Scholar]
- 6.Nathan R., Getz W. M., Revilla E., Holyoak M., Kadmon R., Saltz D., Smouse P. E., A movement ecology paradigm for unifying organismal movement research. Proc. Natl. Acad. Sci. U.S.A. 105, 19052–19059 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Schaub M., Kania W., Köppen U., Variation of primary production during winter induces synchrony in survival rates in migratory white storks Ciconia ciconia. J. Anim. Ecol. 74, 656–666 (2005). [Google Scholar]
- 8.P. Tryjanowski, T. Sparks, L. Jerzak, The White Stork in Poland: Studies in Biology, Ecology and Conservation (Bogucki Wydawnictwo Naukowe, Poznań, Poland, 2006). [Google Scholar]
- 9.Leshem Y., Yom-Tov Y., The use of thermals by soaring migrants. Ibis 138, 667–674 (1996). [Google Scholar]
- 10.Hedenstrom A., Migration by soaring or flapping flight in birds: The relative importance of energy cost and speed. Philos. Trans. R. Soc. London Ser. B 342, 353–361 (1993). [Google Scholar]
- 11.C. J. Pennycuick, Modelling the Flying Bird (Elsevier, Burlington, MA, 2008). [Google Scholar]
- 12.Bohrer G., Brandes D., Mandel J. T., Bildstein K. L., Miller T. A., Lanzone M., Katzner T., Maisonneuve C., Tremblay J. A., Estimating updraft velocity components over large spatial scales: Contrasting migration strategies of golden eagles and turkey vultures. Ecol. Lett. 15, 96–103 (2012). [DOI] [PubMed] [Google Scholar]
- 13.Sapir N., Wikelski M., McCue M. D., Pinshow B., Nathan R., Flight modes in migrating European bee-eaters: Heart rate may indicate low metabolic rate during soaring and gliding. PLOS One 5, e13956 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Sergio F., Tanferna A., De Stephanis R., Jiménez L. López, Blas J., Tavecchia G., Preatoni D., Hiraldo F., Individual improvements and selective mortality shape lifelong migratory performance. Nature 515, 410–413 (2014). [DOI] [PubMed] [Google Scholar]
- 15.Menu S., Gauthier G., Reed A., Survival of young Greater Snow Geese (Chen caerulescens atlantica) during fall migration. The Auk 122, 479–496 (2005). [Google Scholar]
- 16.T. Alerstam, Å. Lindström, in Bird Migration, P. D. E. Gwinner, Ed. (Springer, Berlin, Germany, 1990), pp. 331–351. [Google Scholar]
- 17.Alerstam T., Hake M., Kjellén N., Temporal and spatial patterns of repeated migratory journeys by ospreys. Anim. Behav. 71, 555–566 (2006). [Google Scholar]
- 18.Fuller M. R., Seegar W. S., Schueck L. S., Routes and travel rates of migrating Peregrine Falcons Falco peregrinus and Swainson’s Hawks Buteo swainsoni in the Western Hemisphere. J. Avian Biol. 29, 433–440 (1998). [Google Scholar]
- 19.Klaassen R. H. G., Strandberg R., Hake M., Alerstam T., Flexibility in daily travel routines causes regional variation in bird migration speed. Behav. Ecol. Sociobiol. 62, 1427–1432 (2008). [Google Scholar]
- 20.Shamoun-Baranes J., Baharad A., Alpert P., Berthold P., Yom-Tov Y., Dvir Y., Leshem Y., The effect of wind, season and latitude on the migration speed of white storks Ciconia ciconia, along the Eastern migration route. J. Avian Biol. 34, 97–104 (2003). [Google Scholar]
- 21.Leshem Y., Yom-Tov Y., The magnitude and timing of migration by soaring raptors, pelicans and storks over Israel. Ibis 138, 188–203 (1996). [Google Scholar]
- 22.van Loon E. E., Shamoun-Baranes J., Bouten W., Davis S. L., Understanding soaring bird migration through interactions and decisions at the individual level. J. Theor. Biol. 270, 112–126 (2011). [DOI] [PubMed] [Google Scholar]
- 23.Shamoun-Baranes J., Van Loon E., Alon D., Alpert P., Yom-Tov Y., Leshem Y., Is there a connection between weather at departure sites, onset of migration and timing of soaring-bird autumn migration in Israel? Glob. Ecol. Biogeogr. 15, 541–552 (2006). [Google Scholar]
- 24.Massemin-Challet S., Gendner J.-P., Samtmann S., Pichegru L., Wulgué A., Le Maho Y., The effect of migration strategy and food availability on White Stork Ciconia ciconia breeding success. Ibis 148, 503–508 (2006). [Google Scholar]
- 25.Tortosa F. S., Caballero J. M., Reyes-López J., Effect of rubbish dumps on breeding success in the White Stork in Southern Spain. Waterbirds Int. J. Waterbird Biol. 25, 39–43 (2002). [Google Scholar]
- 26.Pulido F., Berthold P., Current selection for lower migratory activity will drive the evolution of residency in a migratory bird population. Proc. Natl. Acad. Sci. U.S.A. 107, 7341–7346 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.W. Fiedler, F. Bairlein, U. Köppen, Birds and Climate Change, in Advances in Ecological Research, W. Fiedler, P. Berthold, and A. Møller, Eds. (Academic Press, London, 2004), vol. 35, pp. 49–67. [Google Scholar]
- 28.Visser M. E., Perdeck A. C., Van Balen J. H., Both C., Climate change leads to decreasing bird migration distances. Glob. Chang. Biol. 15, 1859–1865 (2009). [Google Scholar]
- 29.Wilcove D. S., Wikelski M., Going, going, gone: Is animal migration disappearing. PLOS Biol. 6, e188 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.V. A. Andronov, V. A. Adamackaya, T. B. Adamackaya, Artukhin, Birds of Russia and Adjacent Regions: Pelecaniformes, Ciconiiformes, Phoenicopteriformes (KMK Sci Press, Moscow, Russia, 2011). [Google Scholar]
- 31.Altizer S., Bartel R., Han B. A., Animal migration and infectious disease risk. Science 331, 296–302 (2011). [DOI] [PubMed] [Google Scholar]
- 32.Bauer S., Hoye B. J., Migratory animals couple biodiversity and ecosystem functioning worldwide. Science 344, 1242552 (2014). [DOI] [PubMed] [Google Scholar]
- 33.A. Flack, W. Fiedler, J. Blas, I. Pokrovski, M. Kaatz, M. Mitropolsky, K. Aghababyan, Y. Fakriadis, E. Makrigianni, L. Jerzak, C. Shamina, H. Azafzaf, S. Rotics, R. Nathan, M. Wikelski, Data from: Migration costs of eight Eurasian white stork populations (2015); available at dx.doi.org/10.5441/001/1.78152p3q.
- 34.G. Creutz, Der Weisstorch: Ciconia ciconia (Ziemsen, Lutherstadt Wittenberg, Germany, 1985). [Google Scholar]
- 35.Holland R. A., Wikelski M., Kümmeth F., Bosque C., The secret life of oilbirds: New insights into the movement ecology of a unique avian frugivore. PLOS One 4, e8264 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Gleiss A. C., Wilson R. P., Shepard E. L. C., Making overall dynamic body acceleration work: On the theory of acceleration as a proxy for energy expenditure. Ecol. Evol. 2, 23–33 (2011). [Google Scholar]
- 37.Dodge S., Bohrer G., Weinzierl R., Davidson S. C., Kays R., Douglas D., Cruz S., Han J., Brandes D., Wikelski M., The environmental-data automated track annotation (Env-DATA) system: Linking animal tracks with environmental data. Mov. Ecol. 1, 3 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Kranstauber B., Cameron A., Weinzerl R., Fountain T., Tilak S., Wikelski M., Kays R., The movebank data model for animal tracking. Environ. Model. Software 26, 834–835 (2011). [Google Scholar]
- 39.Nakagawa S., Schielzeth H., A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 4, 133–142 (2013). [Google Scholar]
- 40.R Development Core Team, R: A Language and Environment for Statistical Computing (R Foundation for Statistical Computing, Vienna, Austria, 2009).
- 41.D. Bates, M. Maechler, Package “lme4” (version 0.999375-32): linear mixed-effects models using S4 classes (2009); available (April 2011) at http://cran.r-project.org/web/packages/lme4/lme4.pdf.
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary material for this article is available at http://advances.sciencemag.org/cgi/content/full/2/1/e1500931/DC1
Table S1. Results of linear regression models concerning the effects of population on departure date.
Table S2. Results of linear regression models concerning the effects of population on maximum distance reached.
Table S3. GLMM model results for animal activity (ODBA) showing the effects of the main predictors. North of 33°N was incorporated in the intercept.
Table S4. GLMM model results concerning the effect of daily distance moved on the location of the wintering grounds.
Table S5. GLMM model results for animal activity (ODBA) showing the effects of human population density (log) and vegetation cover (centered).