

Abstract
The present study aimed to examine the protective role of Spirulina platensis (S. platensis) against arsenic-induced testicular oxidative damage in rats. Arsenic (in the form of NaAsO2 at a dose of 6.3 mg/kg body weight for 8 weeks) caused a significant accumulation of arsenic in testicular tissues as well as a decrease in the levels of testicular superoxide dismutase (SOD), catalase (CAT), reduced glutathione, and zinc. Moreover, it significantly decreased plasma testosterone, luteinizing hormone (LH), triiodothyronine (T3), and thyroxine (T4) levels and reduced sperm motility and sperm count. Arsenic (AS) led to a significant increase in testicular malondialdehyde (MDA), tumour necrosis factor alpha (TNF-α), nitric oxide (NO), and sperm abnormalities. S. platensis at a dose of 300 mg/kg was found to attenuate As-induced oxidative stress, testicular damage, and sperm abnormalities by its potent antioxidant activity. S. platensis may represent a potential therapeutic option to protect the testicular tissue from arsenic intoxication.
1. Introduction
Arsenic contamination occurs due to its industrial uses in the production of agricultural pesticides, wood preservatives, and glass production and in medicine [1, 2]. Arsenic exposure causes obvious damage in various organs, including the male reproductive function as manifested by decrease of androgenesis, suppression of spermatogenesis, and a reduction in the weight of testes and sex organs [3, 4]. However, emerging evidence supports the role of oxidative stress and inflammation with increased production of proinflammatory cytokines in the pathogenesis of arsenic-induced organ damage [5, 6]. Also, previous studies revealed that several antioxidant agents significantly protected against tissue damage due to arsenic intoxication [6, 7].
The cyanobacterium Spirulina is a filamentous blue-green alga belonging to the Oscillatoriaceae family that is generally found in tropical and subtropical regions in warm alkaline water. Spirulina is characterized by high nutritional value where it contains high protein content (60–70% by dry weight), plenty of vitamins, amino acids, gamma-linoleic acid, and minerals [8]. The consumption of Spirulina as a diet supplement has health benefits in preventing or managing hypercholesterolemia [9], hyperglycerolemia [10], obesity, inflammation [11], cancer [12], and cardiovascular disease [13]. In addition, Spirulina has antidiabetic effect [14]. Spirulina provides protection against mercuric chloride-induced oxidative stress and alteration of antioxidant defense mechanism in the liver. These activities were largely related to phycocyanin, an active protein of Spirulina [15]. Phycocyanin (Pc) is a biliprotein of the blue-green alga. This protein contains a tetrapyrrole phycocyanobilin, which is responsible for antioxidant properties of Pc [16]. It has been reported that Pc has significant antioxidant and radical scavenging properties, offering protection against oxidative stress [17]. Antioxidants can reduce arsenic toxicity through chelating it and scavenging free radicals [18]. It was reported that Pc can bind with heavy metals [19]; hence, it can chelate and remove them. In view of the above concerns, the present study was designed to evaluate the antioxidant action of S. platensis enriched with phenolic compounds in ameliorating testicular dysfunction and oxidative stress induced by arsenic.
2. Materials and Methods
2.1. Test Chemicals
Sodium arsenite was purchased from Merck, Germany, while S. platensis was obtained from Alibaba Comp., China, in the form of powder.
2.2. Animals
Four-month male Wistar albino rats, weighting 180–200 g, were got from the animal house, Faculty of Pharmacy, King Saud University. Animals were housed and fed as previously described [20]. The rats were fed a commercially available rat pellet diet ad libitum throughout the experimental period. The rats allowed to adapt to laboratory environment for seven days before the beginning of the experiment. This study was performed in the Zoology Department, Faculty of Science, King Saud University, Saudi Arabia. The care and handling of experimental animals were carried out according to the animal ethical committee of King Saud University, College of Pharmacy.
2.3. Experimental Protocol
The animals were randomly divided into four groups, consisting of eight rats in each, and they were treated for eight weeks as below:
-
Group I: normal control (rats received only water as vehicle).
-
Group II: rats received orally arsenic as sodium arsenite, 6.3 mg/kg corresponding to 15% of LD50 (41 mg/Kg) [21].
-
Group III: rats received orally 300 mg/Kg of S. platensis [22] followed by oral administration of arsenic as sodium arsenite 6.3 mg/Kg daily.
-
Group IV: rats received orally S. platensis only as in group III.
All treatments are carried out daily for eight weeks in order to evaluate their effects [23]. The rats were subjected to ether anesthesia using sliding top chamber (Kent Scientific corporation) during sample collection.
2.4. Sample Preparation and Biochemical Analysis
At the end of the experimentation period, blood and organs were collected as previously described [20]. Plasma testosterone, luteinizing hormone (LH), triiodothyronine (T3), and thyroxine (T4) concentrations were assayed by enzyme immunoassay using commercial kits from Diagnostic products Co., Los Angeles, CA, USA. Testes, vas deferens, epididymis, prostate gland, and seminal vesicle were isolated from surrounding tissues and placed into tubes. The organs were dried between two sheets of filter paper and their wet weight was determined. The organ weight/body weight ratio × 100 was calculated and expressed as relative organ weight beside absolute weight. Epididymis and testes were processed as previously described in order to perform histological, biochemical, and sperm analysis [20].
The supernatant of testicular homogenates was used for determination of malondialdehyde, reduced glutathione, catalase, and superoxide dismutase levels using colorimetric assay kits according to the recommendations of the manufacturer (BioDiagnostic, Egypt). The testicular level of nitric oxide was assayed using colorimetric assay kit following the manufacturer's instructions (Cayman Chemical Company, USA). Also, the level of tumour necrosis factor-α in testicular homogenates was determined by enzyme-linked immunosorbent assay (ELISA) using rat TNF-α immunoassay kit according to the guidance of the manufacturer (R&D Systems, USA). In addition, arsenic and zinc levels in testes were estimated by atomic absorption (Perkin-Elmer, UK).
2.5. Sperm Analysis
Sperm motility, count, and abnormalities were evaluated as previously described [20, 24].
2.6. Statistical Analysis
All values were expressed as mean ± SE. Statistical analysis of data was performed using two-way ANOVA followed by least significant difference (LSD) for comparison of various treatments using the spss 13.0.
3. Results
3.1. Biochemical Analysis
The results demonstrated that supplementation of Spirulina to arsenic exposed rats reduced the arsenic content remarkably in the testis (Figure 1). On the other hand, testicular zinc concentration of arsenic treated groups (Figure 2) decreased significantly as compared with control. Testicular zinc concentration in S. platensis + arsenic group is significantly higher than those treated with arsenic only. Arsenic treatment without S. platensis significantly enhanced the levels of testicular MDA, TNF-α, and nitric oxide concentrations (P ≤ 0.1), while SOD, catalase, and GSH levels decreased significantly as compared with control (Table 1). The administration of S. platensis followed by arsenic intoxication attenuated these effects.
Figure 1.
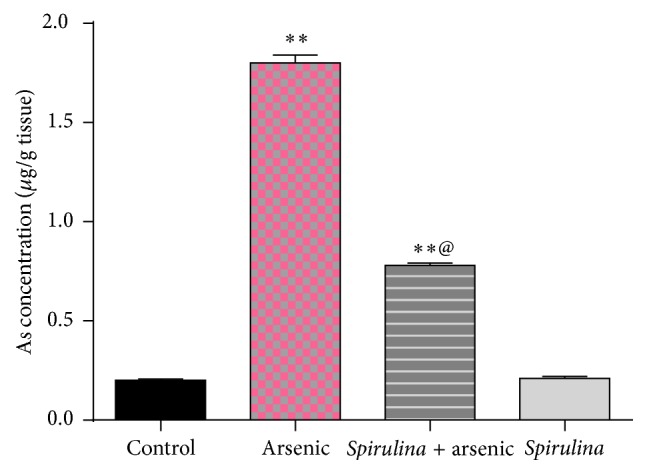
Effect of Spirulina platensis on testicular arsenic concentration (μg/g tissue) in arsenic intoxicated rats. All numbers are mean + standard error, n = 8. AS: arsenic. ∗∗Significantly different from control value, ∗∗ P < 0.01. @Significantly different from arsenic group value, @ P < 0.01.
Figure 2.
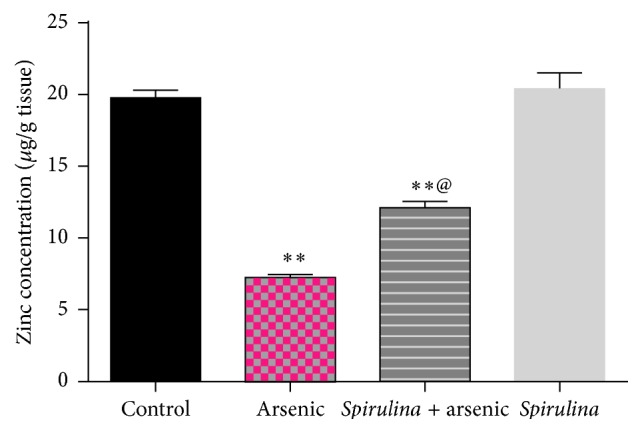
Effect of Spirulina platensis on testicular zinc concentration (µg/g tissue) in arsenic intoxicated rats. All numbers are mean + standard error, n = 8. ∗∗Significantly different from control value, ∗∗ P < 0.01. @Significantly different from arsenic group value, @ P < 0.01.
Table 1.
Effect of S. platensis on testicular oxidative stress parameters in arsenic treated rats.
| Parameter | Treatment | |||
|---|---|---|---|---|
| Control | Arsenic | S. platensis + arsenic | S. platensis | |
| MDA (nmol/mg protein) | 10.48 ± 0.36 | 22.83 ± 0.89∗∗ | 14.95 ± 0.65∗∗@ | 8.76 ± 0.67∗ |
| SOD (unit/mg protein) | 24.27 ± 0.65 | 9.11 ± 0.35∗∗ | 16.81 ± 0.44∗∗@ | 26.89 ± 0.21∗∗ |
| Catalase (μmol/min/mg protein) | 30.28 ± 1.06 | 15.86 ± 0.35∗∗ | 22.75 ± 0.74∗∗@ | 32.11 ± 2.10 |
| GSH (nmol/mg protein) | 27.25 ± 1.47 | 14.13 ± 0.89∗∗ | 21.25 ± 1.14∗∗@ | 39.87 ± 1.86∗∗ |
| TNF-α (pg/100 mg tissue) | 11.48 ± 0.21 | 108.12 ± 3.15∗∗ | 46.92 ± 2.71∗∗@ | 10.65 ± 0.47 |
| Nitric oxide (nmol/100 mg tissue) | 85.20 ± 4.14 | 216.92 ± 5.78∗∗ | 130.41 ± 6.37∗∗@ | 87.14 ± 3.60 |
All numbers are mean + standard error, n = 8.
∗Significantly different from control value, ∗ P < 0.05, ∗∗ P < 0.01.
@Significantly different from arsenic group value, @ P < 0.01.
MDA: malondialdehyde; SOD: superoxide dismutase; GSH: reduced glutathione; TNF-α: tumor necrosis factor-alpha.
3.2. Reproductive Organ Weights
Sodium arsenite intoxication significantly decreased the testis, vas deferens, epididymis, prostate, and seminal vesicle weights. Treatment with Spirulina prior to arsenic administration, however, kept the weight of reproductive organs close to normal (Tables 2(a) and 2(b)).
Table 2.
(a) Absolute reproductive organ weights (g) of arsenic intoxicated rats treated with S. platensis. (b) Effect of S. platensis on reproductive organ weights (g) relative to body weight in arsenic intoxified rats.
(a).
| Parameter | Treatment | |||
|---|---|---|---|---|
| Control | Arsenic | S. platensis + arsenic | S. platensis | |
| Left testis | 1.76 ± 0.05 | 1.40 ± 0.07∗∗ | 1.67 ± 0.05@ | 1.79 ± 0.08 |
| Vas deferens | 0.21 ± 0.01 | 0.13 ± 0.01∗∗ | 0.17 ± 0.007∗∗ | 0.19 ± 0.006 |
| Epididymis | 0.86 ± 0.03 | 0.57 ± 0.02∗∗ | 0.73 ± 0.03∗ | 0.81 ± 0.04 |
| Prostate | 0.80 ± 0.02 | 0.37 ± 0.03∗∗ | 0.65 ± 0.05∗∗@ | 0.77 ± 0.05 |
| Seminal vesicle | 1.51 ± 0.08 | 0.86 ± 0.06∗∗ | 1.18 ± 0.06∗∗@ | 1.46 ± 0.07 |
(b).
| Treatment | ||||
|---|---|---|---|---|
| Parameter | Control | Arsenic | S. platensis + arsenic | S. platensis |
| Left testis | 0.62 ± 0.02 | 0.51 ± 0.02∗∗ | 0.59 ± 0.01@ | 0.60 ± 0.02 |
| Vas deferens | 0.07 ± 0.003 | 0.05 ± 0.002∗∗ | 0.05 ± 0.002∗∗ | 0.06 ± 0.005 |
| Epididymis | 0.25 ± 0.002 | 0.21 ± 0.006∗∗ | 0.23 ± 0.007∗ | 0.24 ± 0.008 |
| Prostate | 0.31 ± 0.02 | 0.13 ± 0.01∗∗ | 0.20 ± 0.009∗∗@ | 0.29 ± 0.01 |
| Seminal vesicle | 0.52 ± 0.01 | 0.31 ± 0.01∗∗ | 0.47 ± 0.02∗@ | 0.50 ± 0.04 |
All numbers are mean + standard error, n = 8.
∗Significantly different from control value, ∗ P < 0.05, ∗∗ P < 0.01.
@Significantly different from arsenic group value, @ P < 0.01.
3.3. Plasma Hormones Level
We observed that sodium arsenite intoxication decreased the levels of testosterone, LH, T3, and T4 significantly (P ≤ 0.01) compared to control values (Figures 3 and 4). Treatment with S. platensis was found to be effective in alleviation of alteration in hormone levels induced by the arsenic.
Figure 3.
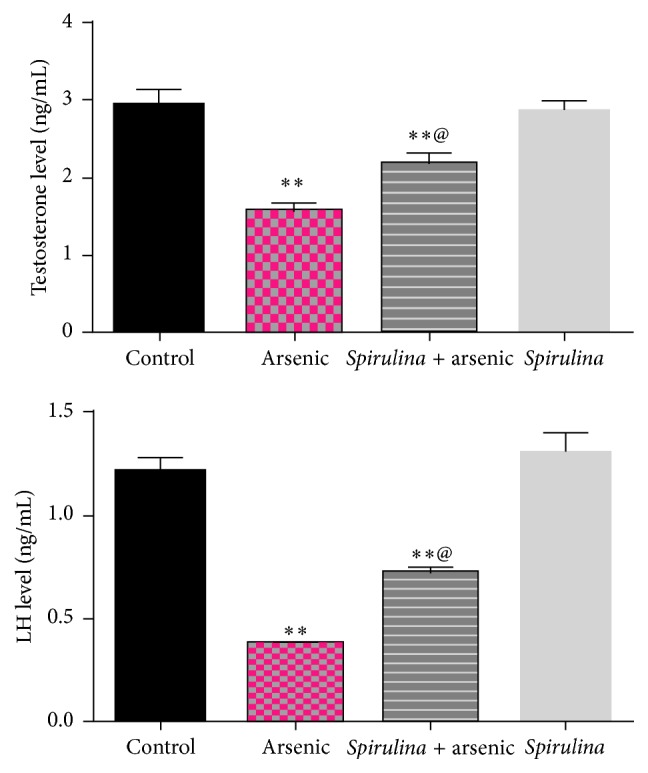
Effect of S. platensis on plasma testosterone and luteinizing hormone (LH) levels in arsenic intoxicated rats. All numbers are mean + standard error, n = 8. ∗∗Significantly different from control value, ∗∗ P < 0.01. @Significantly different from arsenic group value, @ P < 0.01.
Figure 4.
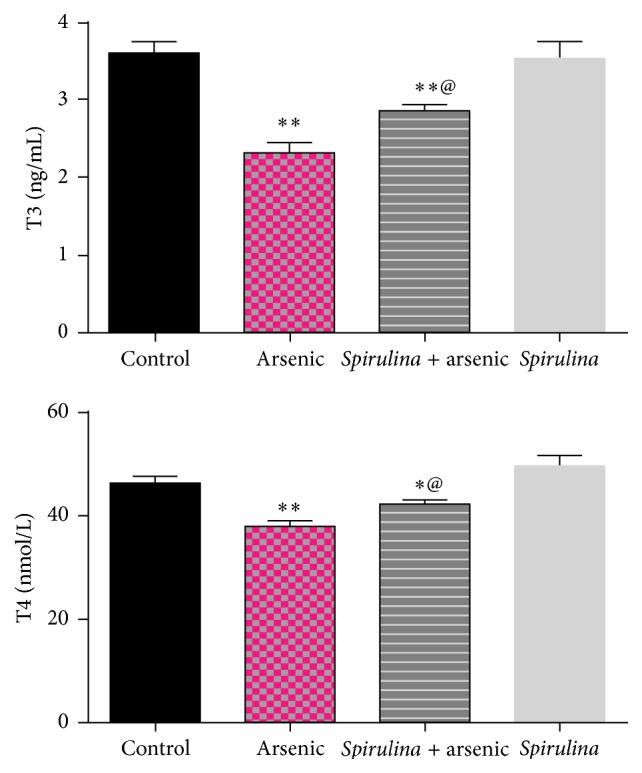
Effect of S. platensis on T3 and T4 concentrations in arsenic intoxicated rats. All numbers are mean + standard error, n = 8. ∗Significantly different from control value, ∗ P < 0.05, ∗∗ P < 0.01. @Significantly different from arsenic group value, @ P < 0.01. T3: triiodothyronine; T4: thyroxine.
3.4. Sperm Motility, Count, and Abnormalities
Arsenic intoxication decreased the sperm motility and count compared to the normal control (Table 3). In addition, a significant increase of sperm abnormalities was found in rats treated with arsenic. S. platensis administration reduced the toxic effects of arsenic on sperms.
Table 3.
Effect of S. platensis on sperm morphological parameters in experimental arsenic exposed rats.
| Parameter | Treatment | |||
|---|---|---|---|---|
| Control | Arsenic | S. platensis + arsenic | S. platensis | |
| Sperm motility (%) | 83.56 ± 1.18 | 72.29 ± 2.00∗∗ | 84.56 ± 0.67@ | 90.42 ± 2.10∗ |
| Sperm count per epididymis (million/epididymis) | 17.47 ± 1.06 | 7.09 ± 0.41∗∗ | 12.68 ± 0.85∗∗@ | 25.5 ± 1.15∗∗ |
| Abnormal sperm rate (%) | ||||
| Head | 2.16 ± 0.13 | 8.92 ± 0.56∗∗ | 4.94 ± 0.19∗∗@ | 1.90 ± 0.07 |
| Tail | 1.83 ± 0.11 | 2.88 ± 0.14∗∗ | 2.14 ± 0.18@ | 2.11 ± 0.12 |
| Total | 3.99 ± 0.14 | 11.34 ± 0.51∗∗ | 6.92 ± 0.32∗∗@ | 4.00 ± 0.16 |
All numbers are mean + standard error, n = 8.
∗Significantly different from control value, ∗ P < 0.05, ∗∗ P < 0.01.
@Significantly different from arsenic group value, @ P < 0.01.
3.5. Histopathological Observation
Histological observation of the testes of control animals showed normal spermatogenic cells with normal arrangement (Figure 5(a)). The section of testis of arsenic treated rat showed (Figure 5(b)) thickening of tubules basement membrane, vascular degeneration, marked decrease in spermatogenic cells population, hemorrhage in interstitial tissues, and deformation of Leydig cells. Moreover, the sperm bundles were absent in some tubules. Pretreatment with S. platensis could, however, prevent the As-toxicity and maintain the normalcy of the testicular architecture (Figure 5(c)).
Figure 5.

(a) Photomicrograph of the testis of control rat showing normal structure of seminiferous tubules containing different types of spermatogenic cells (H&E, ×400). (b) Photomicrograph of the testis of rat that received arsenic showing deformed Leydig cells (white arrow), vacuolated spermatogenic cells (VD), thickened basement membrane (dotted arrow), and congestion of blood vessel (black arrow) (H&E, ×400). (c) Photomicrograph of the testis of rat treated with S. platensis + As showing normal spermatogenesis and cell arrangement (H&E, ×400). (d) Photomicrograph of the testis of rat treated with S. platensis showing normal structure (H&E, ×400).
4. Discussion
The response of male rats to the protective effects of S. platensis against arsenic-induced oxidative stress and reprotoxicity was examined in this study. Our results proposed that the increase of testicular MDA level may result from arsenic accumulation in the testis suggesting oxidative stress following free radical generation. Enhancement of lipid peroxidation and inhibition of the antioxidant enzymes in the testes are important mechanisms for arsenic pathogenesis [25]. The testicular tissue is provided with an antioxidant defense system including several enzymes functioning in a collective manner for the removing free radicals generated within the cell. SOD and catalase are major enzymes that get rid of reactive oxygen species (ROS) [26]. In the present study, the animals treated with arsenic showed decreased activities of testicular antioxidant enzymes, SOD, and CAT that may indicate the antioxidant imbalance induced by arsenic. A decrease in the activity of SOD can be referred to as an enhanced superoxide production during arsenic metabolism. SOD catalyzes the dismutation of superoxide anions and prevents the subsequent formation of hydroxyl radicals [27]. The observed decreased testicular SOD might be responsible for increased lipid peroxidation following arsenic treatment [28]. The superoxide radical also reduced the activity of catalase [29]. Moreover, exposure to arsenic reduces the testicular GSH content of the present rats as previously found [30, 31].
The improved antioxidant status of testicular tissues by S. platensis can be deduced from elevated levels of testicular SOD, CAT, zinc, and GSH and a decrease of MDA and arsenic concentrations of S. platensis + arsenic group as compared to arsenic group. The antioxidant properties of S. platensis may be attributed to the presence of potent antioxidant components as β-carotene, vitamin C, vitamin E, selenium, and manganese [32–37]. Moreover, phycocyanin of S. platensis significantly inhibited peroxyl radical induced lipid peroxidation [16] and it may chelate arsenic as it binds with heavy metals [38].
Free radicals are able to induce cytokine production from various cell types [39]. The decreased antioxidant enzyme activities with elevated lipid peroxidation, TNF-α, and NO levels indicated impaired antioxidative defense mechanisms with an oxidative injury in the testes of arsenic group. It was reported that there was a link between TNF-α or NO and oxidative stress. Both TNF-α and NO can increase the production of reactive oxygen species and oxidative stress [40, 41]. It was found that both NO and TNF-α inhibited testosterone synthesis pathways [42, 43]. The significant decrease in the plasma level of testosterone in the present rats treated with arsenic may be due to its direct effect on the testis or suppression of luteinizing hormone secretion. S. platensis represses proinflammatory cytokine expression and secretion through suppression of nuclear factor kappa (NF-κB). Activation of NF-κB pathway is a major pathway for the development of inflammatory diseases [44]. The antioxidants found in S. platensis maintain the endogenous antioxidants and inhibit elevation of testicular NO and TNF-α, thus reducing oxidative stress and relieving the pathological changes induced by arsenic in testis which may lead to improvement of testosterone level.
A significant decrease in the weights of testis and accessory sex organs was observed in arsenic exposed rats, which may be due to the inhibition of spermatogenesis and decreased steroidogenesis. It is well known that the testosterone stimulates normal growth and function of male reproductive system [45]. The weight of the testis is also largely dependent on the mass of the differentiated spermatogenic cells and reduction in the testicular weight indicates germ cell loss [3]. Our results showed that S. platensis alleviated the reduction in T3 and T4 levels induced by arsenic. It is well known that thyroid hormones affect spermatogenesis [46]. In addition, the number of sperm production by testes was decreased significantly in hypothyroid rats and increased in hyperthyroid rats in comparison with the control group rats. It was shown that thyroid hormone receptor expresses in the germ cells from spermatogonia to primary spermatocytes [47].
A higher ROS production or a decreased antioxidant capacity is responsible for stimulation of lipid peroxidation production which affects sperm motility [48]. The observed decrease in the number of sperm count and motility and increase of sperm morphological abnormalities may result from less production of androgen in arsenic exposed rats or from increased level of testicular lipid peroxidation. Spermatozoa are particularly liable to ROS-induced damage because their plasma membranes have large quantities of polyunsaturated fatty acids and their cytoplasm comprises low concentration of the scavenging enzymes [49]. It is documented that ROS generation can induce abnormal sperm morphology [50]. It appeared that S. platensis, containing potent antioxidants, significantly reversed the deleterious effects of arsenic on sperms. Thus, the antioxidative properties of S. platensis may play a positive role in the defense against oxidative stress induced by arsenic. Our previous findings clearly highlight the role of S. platensis as a protective modulator of mercuric chloride-induced testicular injuries and oxidative stress [20]. Here, S. platensis significantly lessen the increase in arsenic concentration, and the reduction in zinc concentration of testicular tissue resulted from sodium arsenite administration. Zinc acts as a cofactor for superoxide dismutase, preserves the reduced glutathione, and induces metallothionein which has antioxidant and metal-chelating properties [51]. Zinc acts as an effective anti-inflammatory and antioxidant agent [52]. It can be speculated that S. platensis through its antioxidant activity decreased the arsenic burden in testicular tissue and restored the depleted zinc which results in an additional protective effect against arsenic-mediated testicular toxicity.
The present investigation showed that the treatment of the rats with S. platensis improves sperm characteristics as manifested by increase of sperm motility and count. The improvement of sperm parameters may be due to antioxidant components of S. platensis [53, 54].
In conclusion, the protective actions of S. platensis against arsenic are believed to originate from its free radical scavenging, antioxidant activities, maintenance of antioxidant enzymes, and a decrease in the production of inflammatory mediators that are implicated in the pathogenesis of arsenic-induced testicular injury. Therefore, S. platensis represents a potential agent to prevent testicular injury and dysfunction induced by arsenic exposure.
Acknowledgments
The authors would like to thank Dr. Ibrahim EL-Hazza Scientific Group, for their guidance and helping during the experiment period. The authors would also like to extend their sincere appreciation to the Deanship of Scientific Research at King Saud University for funding this Research Group no. RGP-1435-093.
Conflict of Interests
There is no conflict of interests regarding the publication of this paper.
References
- 1.Wang J. P., Qi L., Moore M. R., Ng J. C. A review of animal models for the study of arsenic carcinogenesis. Toxicology Letters. 2002;133(1):17–31. doi: 10.1016/s0378-4274(02)00086-3. [DOI] [PubMed] [Google Scholar]
- 2.Lengfelder E., Hofmann W.-K., Nowak D. Impact of arsenic trioxide in the treatment of acute promyelocytic leukemia. Leukemia. 2012;26(3):433–442. doi: 10.1038/leu.2011.245. [DOI] [PubMed] [Google Scholar]
- 3.Sarkar M., Chaudhuri G. R., Chattopadhyay A., Biswas N. M. Effect of sodium arsenite on spermatogenesis, plasma gonadotrophins and testosterone in rats. Asian Journal of Andrology. 2003;5(1):27–31. [PubMed] [Google Scholar]
- 4.Pant N., Murthy R. C., Srivastava S. P. Male reproductive toxicity of sodium arsenite in mice. Human and Experimental Toxicology. 2004;23(8):399–403. doi: 10.1191/0960327104ht467oa. [DOI] [PubMed] [Google Scholar]
- 5.Chang S. I., Jin B., Youn P., Park C., Park J.-D., Ryu D.-Y. Arsenic-induced toxicity and the protective role of ascorbic acid in mouse testis. Toxicology and Applied Pharmacology. 2007;218(2):196–203. doi: 10.1016/j.taap.2006.11.009. [DOI] [PubMed] [Google Scholar]
- 6.Das J., Ghosh J., Manna P., Sinha M., Sil P. C. Taurine protects rat testes against NaAsO2-induced oxidative stress and apoptosis via mitochondrial dependent and independent pathways. Toxicology Letters. 2009;187(3):201–210. doi: 10.1016/j.toxlet.2009.03.001. [DOI] [PubMed] [Google Scholar]
- 7.Ramanathan K., Anusuyadevi M., Shila S., Panneerselvam C. Ascorbic acid and α-tocopherol as potent modulators of apoptosis on arsenic induced toxicity in rats. Toxicology Letters. 2005;156(2):297–306. doi: 10.1016/j.toxlet.2004.12.003. [DOI] [PubMed] [Google Scholar]
- 8.Hoseini S. M., Khosravi-Darani K., Mozafari M. R. Nutritional and medical applications of spirulina microalgae. Mini-Reviews in Medicinal Chemistry. 2013;13(8):1231–1237. doi: 10.2174/1389557511313080009. [DOI] [PubMed] [Google Scholar]
- 9.Ferreira-Hermosillo A., Torres-Duran P. V., Juarez-Oropeza M. A. Hepatoprotective effects of Spirulina maxima in patients with non-alcoholic fatty liver disease: a case series. Journal of Medical Case Reports. 2010;4(103):1–5. doi: 10.1186/1752-1947-4-103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Deng R., Chow T.-J. Hypolipidemic, antioxidant, and antiinflammatory activities of microalgae spirulina. Cardiovascular Therapeutics. 2010;28(4):e33–e45. doi: 10.1111/j.1755-5922.2010.00200.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Coskun Z. K., Kerem M., Gurbuz N., et al. The study of biochemical and histopathological effects of spirulina in rats with TNBS-induced colitis. Bratislava Medical Journal. 2011;112(5):235–243. [PubMed] [Google Scholar]
- 12.Ismail M. F., Ali D. A., Fernando A., et al. Chemoprevention of rat liver toxicity and carcinogenesis by Spirulina. International Journal of Biological Sciences. 2009;5(4):377–387. doi: 10.7150/ijbs.5.377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Khan M., Shobha J. C., Mohan I. K., et al. Protective effect of Spirulina against doxorubicin-induced cardiotoxicity. Phytotherapy Research. 2005;19(12):1030–1037. doi: 10.1002/ptr.1783. [DOI] [PubMed] [Google Scholar]
- 14.Karkos P. D., Leong S. C., Karkos C. D., Sivaji N., Assimakopoulos D. A. Spirulina in clinical practice: evidence-based human applications. Evidence-Based Complementary and Alternative Medicine. 2011;2011:4. doi: 10.1093/ecam/nen058.531053 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Romay C., Armesto J., Remirez D., González R., Ledon N., García I. Antioxidant and anti-inflammatory properties of C-phycocyanin from blue-green algae. Inflammation Research. 1998;47(1):36–41. doi: 10.1007/s000110050256. [DOI] [PubMed] [Google Scholar]
- 16.Bhat V. B., Madyastha K. M. Scavenging of peroxynitrite by phycocyanin and phycocyanobilin from Spirulina platensis: protection against oxidative damage to DNA. Biochemical and Biophysical Research Communications. 2001;285(2):262–266. doi: 10.1006/bbrc.2001.5195. [DOI] [PubMed] [Google Scholar]
- 17.Lissi E. A., Pizarro M., Aspee A., Romay C. Kinetics of phycocyanine bilin groups destruction by peroxyl radicals. Free Radical Biology and Medicine. 2000;28(7):1051–1055. doi: 10.1016/s0891-5849(00)00193-3. [DOI] [PubMed] [Google Scholar]
- 18.Rana S. V. S. Protective effect of ascorbic acid against oxidative stress induced by inorganic arsenic in liver and kidney of rat. Indian Journal of Experimental Biology. 2007;45(4):371–375. [PubMed] [Google Scholar]
- 19.Gelagutashvili E. S., Belokobyl'skii A. I., Rcheulishvili A. N., Mosulishvili L. M. Interaction of Pb(II) ions with C-phycocyanin from Spirulina platensis: effect of ionic strength. Biofizika. 2003;48(4):589–594. [PubMed] [Google Scholar]
- 20.El-Desoky G. E., Bashandy S. A., Alhazza I. M., Al-Othman Z. A., Aboul-Soud M. A. M., Yusuf K. Improvement of mercuric chloride-induced testis injuries and sperm quality deteriorations by Spirulina platensis in rats. PLoS ONE. 2013;8(3) doi: 10.1371/journal.pone.0059177.e59177 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Lewis R., Saxs J. Dangerous Properties of Industrial Materials. 9th. New York, NY, USA: V. N. Reinhold; 1996. [Google Scholar]
- 22.Simsek N., Karadeniz A., Kalkan Y., Keles O. N., Unal B. Spirulina platensis feeding inhibited the anemia- and leucopenia-induced lead and cadmium in rats. Journal of Hazardous Materials. 2009;164(2-3):1304–1309. doi: 10.1016/j.jhazmat.2008.09.041. [DOI] [PubMed] [Google Scholar]
- 23.Sengupta P. A scientific review of age determination for a laboratory rat: how old is it in comparison with human age? Biomedicine International. 2011;2:81–89. [Google Scholar]
- 24.Narayana K., Prashanthi N., Nayanatara A., Kumar H. H. C., Abhilash K., Bairy K. L. Effects of methyl parathion (o,o-dimethyl o-4-nitrophenyl phosphorothioate) on rat sperm morphology and sperm count, but not fertility, are associated with decreased ascorbic acid level in the testis. Mutation Research. 2005;588(1):28–34. doi: 10.1016/j.mrgentox.2005.08.012. [DOI] [PubMed] [Google Scholar]
- 25.Chen Y.-C., Lin-Shiau S.-Y. N., Lin J.-K. Involvement of reactive oxygen species and caspase 3 activation in arsenite-induced apoptosis. Journal of Cellular Physiology. 1998;177(2):324–333. doi: 10.1002/(sici)1097-4652(199811)177:2<324::aid-jcp14>3.0.co;2-9. [DOI] [PubMed] [Google Scholar]
- 26.Rasyidah T. I., Suhana S., Nur-Hidayah H., Kaswandi M. A., Noah R. M. Evaluation of antioxidant activity of Zingiber officinale (ginger) on formalin—induced testicular toxicity in rats. Journal of Medical and Bioengineering. 2014;3(3):149–153. doi: 10.12720/jomb.3.3.149-153. [DOI] [Google Scholar]
- 27.Imlay J. A., Linn S. DNA damage and oxygen radical toxicity. Science. 1988;240(4857):1302–1309. doi: 10.1126/science.3287616. [DOI] [PubMed] [Google Scholar]
- 28.Maiti S., Chatterjee A. K. Differential response of cellular antioxidant mechanism of liver and kidney to arsenic exposure and its relation to dietary protein deficiency. Environmental Toxicology and Pharmacology. 2000;8(4):227–235. doi: 10.1016/S1382-6689(00)00046-6. [DOI] [PubMed] [Google Scholar]
- 29.Kono Y., Fridovich I. Superoxide radical inhibits catalase. Journal of Biological Chemistry. 1982;257(10):5751–5754. [PubMed] [Google Scholar]
- 30.Zakharyan R. A., Aposhian H. V. Enzymatic reduction of arsenic compounds in mammalian systems: the rate-limiting enzyme of rabbit liver arsenic biotransformation is MMA(V) reductase. Chemical Research in Toxicology. 1999;12(12):1278–1283. doi: 10.1021/tx9901231. [DOI] [PubMed] [Google Scholar]
- 31.Radabaugh T. R., Aposhian H. V. Enzymatic reduction of arsenic compounds in mammalian systems: reduction of arsenate to arsenite by human liver arsenate reductase. Chemical Research in Toxicology. 2000;13(1):26–30. doi: 10.1021/tx990115k. [DOI] [PubMed] [Google Scholar]
- 32.Mazo V. K., Gmoshinskii I. V., Zilova I. S. Microalgae Spirulina in human nutrition. Voprosy Pitaniia. 2004;73(1):45–53. [PubMed] [Google Scholar]
- 33.Tang G., Suter P. M. Vitamin A, nutrition, and health values of Algae: spirulina, chlorella, and dunaliella. Journal of Pharmacy and Nutrition Sciences. 2011;1(2):111–118. doi: 10.6000/1927-5951.2011.01.02.04. [DOI] [Google Scholar]
- 34.Stivala L. A., Savio M., Cazzalini O., et al. Effect of beta-carotene on cell cycle progression of human fibroblasts. Carcinogenesis. 1996;17(11):2395–2401. doi: 10.1093/carcin/17.11.2395. [DOI] [PubMed] [Google Scholar]
- 35.Mueller L., Boehm V. Antioxidant activity of β-carotene compounds in different in vitro assays. Molecules. 2011;16(2):1055–1069. doi: 10.3390/molecules16021055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Garg M. C., Chaudhary D. P., Bansal D. D. Effect of vitamin E supplementation on diabetes induced oxidative stress in experimental diabetes in rats. Indian Journal of Experimental Biology. 2005;43(2):177–180. [PubMed] [Google Scholar]
- 37.El-Demerdash F. M. Effects of selenium and mercury on the enzymatic activities and lipid peroxidation in brain, liver, and blood of rats. Journal of Environmental Science and Health, Part B: Pesticides, Food Contaminants, and Agricultural Wastes. 2001;36(4):489–499. doi: 10.1081/pfc-100104191. [DOI] [PubMed] [Google Scholar]
- 38.Plazinski W. Binding of heavy metals by algal biosorbents, theoretical models of kinetics, equilibria and thermodynamics. Advances in Colloid and Interface Science. 2013;197-198:58–67. doi: 10.1016/j.cis.2013.04.002. [DOI] [PubMed] [Google Scholar]
- 39.Ali M. H., Schlidt S. A., Chandel N. S., Hynes K. L., Schumacker P. T., Gewertz B. L. Endothelial permeability and IL-6 production during hypoxia: role of ROS in signal transduction. American Journal of Physiology—Lung Cellular and Molecular Physiology. 1999;277(5):L1057–L1065. doi: 10.1152/ajplung.1999.277.5.L1057. [DOI] [PubMed] [Google Scholar]
- 40.Suematsu N., Tsutsui H., Wen J., et al. Oxidative stress mediates tumor necrosis factor-α-induced mitochondrial DNA damage and dysfunction in cardiac myocytes. Circulation. 2003;107(10):1418–1423. doi: 10.1161/01.cir.0000055318.09997.1f. [DOI] [PubMed] [Google Scholar]
- 41.Clancy R. M., Abramson S. B. Nitric oxide: a novel mediator of inflammation. Proceedings of the Society for Experimental Biology and Medicine. 1995;210(2):93–101. doi: 10.3181/00379727-210-43927aa. [DOI] [PubMed] [Google Scholar]
- 42.Del Punta K., Charreau E. H., Pignataro O. P. Nitric oxide inhibits leydig cell steroidogenesis. Endocrinology. 1996;137(12):5337–5343. doi: 10.1210/en.137.12.5337. [DOI] [PubMed] [Google Scholar]
- 43.Hong C. Y., Park J. H., Ahn R. S., et al. Molecular mechanism of suppression of testicular steroidogenesis by proinflammatory cytokine tumor necrosis factor alpha. Molecular and Cellular Biology. 2004;24(7):2593–2604. doi: 10.1128/mcb.24.7.2593-2604.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Ku C. S., Pham T. X., Park Y., et al. Edible blue-green algae reduce the production of pro-inflammatory cytokines by inhibiting NF-κB pathway in macrophages and splenocytes. Biochimica et Biophysica Acta (BBA)—General Subjects. 2013;1830(4):2981–2988. doi: 10.1016/j.bbagen.2013.01.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Dohle G. R., Smit M., Weber R. F. A. Androgens and male fertility. World Journal of Urology. 2003;21(5):341–345. doi: 10.1007/s00345-003-0365-9. [DOI] [PubMed] [Google Scholar]
- 46.Ai J., Zarifkar A., Takhshid M. A., Alavi J., Moradzadeh M. The effect of thyroid activity on adult rat spermatogenesis. Iranian Journal of Veterinary Research, University of Shiraz. 2007;8(2):155–160. [Google Scholar]
- 47.Buzzard J. J., Morrison J. R., O'Bryan M. K., Song Q., Wreford N. G. Developmental expression of thyroid hormone receptors in the rat testis. Biology of Reproduction. 2000;62(3):664–669. doi: 10.1095/biolreprod62.3.664. [DOI] [PubMed] [Google Scholar]
- 48.Aitken R. J. Free radicals, lipid peroxidation and sperm function. Reproduction, Fertility, and Development. 1995;7(4):659–668. doi: 10.1071/rd9950659. [DOI] [PubMed] [Google Scholar]
- 49.Vernet P., Aitken R. J., Drevet J. R. Antioxidant strategies in the epididymis. Molecular and Cellular Endocrinology. 2004;216(1):31–39. doi: 10.1016/j.mce.2003.10.069. [DOI] [PubMed] [Google Scholar]
- 50.Ramya T., Misro M. M., Sinha D., Nandan D. Sperm function and seminal oxidative stress as tools to identify sperm pathologies in infertile men. Fertility and Sterility. 2010;93(1):297–300. doi: 10.1016/j.fertnstert.2009.05.074. [DOI] [PubMed] [Google Scholar]
- 51.Rooney J. P. K. The role of thiols, dithiols, nutritional factors and interacting ligands in the toxicology of mercury. Toxicology. 2007;234(3):145–156. doi: 10.1016/j.tox.2007.02.016. [DOI] [PubMed] [Google Scholar]
- 52.Prasad A. S., Bao B., Beck F. W. J., Kucuk O., Sarkar F. H. Antioxidant effect of zinc in humans. Free Radical Biology and Medicine. 2004;37(8):1182–1190. doi: 10.1016/j.freeradbiomed.2004.07.007. [DOI] [PubMed] [Google Scholar]
- 53.Yousef M. I., Abdallah G. A., Kamel K. I. Effect of ascorbic acid and vitamin E supplementation on semen quality and biochemical parameters of male rabbits. Animal Reproduction Science. 2003;76(1-2):99–111. doi: 10.1016/s0378-4320(02)00226-9. [DOI] [PubMed] [Google Scholar]
- 54.Rezvanfar M. A., Sadrkhanlou R. A., Ahmadi A., et al. Protection of cyclophosphamide-induced toxicity in reproductive tract histology, sperm characteristics, and DNA damage by an herbal source; evidence for role of free-radical toxic stress. Human and Experimental Toxicology. 2008;27(12):901–910. doi: 10.1177/0960327108102046. [DOI] [PubMed] [Google Scholar]