

Abstract
Background
Phosphoinositide-dependent protein kinase-1 (PDK-1), which functions downstream of phosphoinositide 3-kinase (AGE-1) and activates protein kinases of the AGC family, plays critical roles in regulating biology processes, such as metabolism, growth, development and survival. In the free-living nematode Caenorhabditis elegans, PDK-1 is a key component of the insulin-like signalling pathway, regulating the entry into and exit from dauer (arrested development). Although it is proposed that similar molecular mechanisms control the transition from the free-living to the parasitic stages of nematodes, nothing is known about PDK-1 in Haemonchus contortus, a socioeconomically important gastric nematode of ruminants.
Methods
Here, we isolated and characterized the pdk-1 gene (Hc-pdk-1) and its inferred product (Hc-PDK-1) from H. contortus. Using in vitro and in vivo methods, we then studied the transcriptional profiles of Hc-pdk-1 and anatomical gene expression patterns of Hc-PDK-1 in different developmental stages of C. elegans.
Results
In silico analysis of Hc-PDK-1 displayed conserved functional domains, such as protein kinase and pleckstrin homology (PH) domains and two predicted phosphorylation sites (Thr226/Tyr229), which are crucial for the phosphorylation of downstream signalling. The Hc-pdk-1 gene is transcribed in all of the main developmental stages of H. contortus, with its highest transcription in the infective third-stage larvae (iL3) compared with other stages. Transgene constructs, in which respective promoters were fused to the coding sequence for green fluorescent protein (GFP), were used to transform C. elegans, and to localize and compare the expression of Hc-pdk-1 and Ce-pdk-1. The expression of GFP under the control of the Hc-pdk-1 promoter was localized to the intestine, and head and tail neurons, contrasting somewhat the profile for the C. elegans ortholog, which is expressed in pharynx, intestine and head and tail neurons.
Conclusions
This is the first characterization of pdk-1/PDK-1 from a trichostrongyloid nematode. Taken together, the findings from this study provide a first glimpse of the involvement of Hc-pdk-1 in the insulin-like signalling pathway in H. contortus.
Electronic supplementary material
The online version of this article (doi:10.1186/s13071-016-1351-6) contains supplementary material, which is available to authorized users.
Keywords: Haemonchus contortus, Transgenesis, Development, pdk-1 gene
Background
Haemonchus contortus is an important strongylid nematode infecting millions of small ruminants (including sheep and goats) globally. The disease (haemonchosis) caused by this nematode leads to major economic losses associated with morbidity, reduced production and mortality in these livestock species. H. contortus feeds on blood in the stomach (abomasum), and causes anaemia, oedema and, in extreme cases, death. Anthelmintic treatment has been an essential part of the control of H. contortus and related parasites. However, drug-resistance is now widespread in many countries, due to an excessive and often uncontrolled use of anthelmintics [1–4]. Therefore, it is important to work toward developing new anthelmintics and/or vaccines, built on a sound understanding of key molecules in biological pathways of H. contortus and related parasites [5, 6].
The infective, third-stage larva (iL3) of H. contortus is a motile, free-living stage that is arrested in its development until it enters the host animal, after which it becomes parasitic. Like iL3 of H. contortus, the dauer stage of Caenorhabditis elegans is also arrested in its development; this stage ceases to feed under unfavourable conditions, such as overcrowding, limited food availability and high environmental temperature, and has an extended lifespan [7]. Interestingly, when environmental conditions improve, C. elegans can exit dauer, and continues to develop and reproduce normally [8].
Evolutionarily, both C. elegans and H. contortus belong to clade V nematodes [9]. Therefore, it has been postulated that the exit from dauer in the free-living nematode (C. elegans) and activation of iL3 in the parasitic nematode (H. contortus) are governed by analogous molecular mechanisms [10–13]. Dauer is regulated principally by four signalling pathways, one of which is the insulin-like signaling pathway which involves several proteins, including DAF-2 [14], AGE-1 [15], PDK-1 [16], AKT-1/2 [17] and DAF-16 [18–20]. Signalling via DAF-2, AGE-1 and PDK-1 activates AKT-1/2 by phosphorylation, which, in turn, phosphorylates and, thus, negatively regulates DAF-16, a core regulator of multiple biological processes, such as longevity, stress-resistance and developmental arrest [21, 22].
Although advances have been made in understanding the molecular mechanisms of the dauer state, very little is known about the analogous process of iL3 development in parasitic nematodes. The genomes and transcriptomes of H. contortus [23, 24] provide a solid foundation for exploring key molecules in this transition process, but a lack of effective genetic and in vitro culture methods restricts somewhat the study of molecular mechanisms of these nematodes [25–27]. In contrast, C. elegans has been used as a surrogate system to explore molecular functions in some parasitic nematodes, such as H. contortus [28–30], Ancylostoma caninum [31, 32] and Strongyloides stercoralis [33, 34]. In the present study, we isolated and characterized the pdk-1 ortholog from H. contortus (Hc-pdk-1). Using in vitro and in vivo methods, we studied the transcriptional profiles of Hc-pdk-1 and anatomical gene expression patterns of Hc-PDK-1 in different developmental stages of C. elegans.
Methods
Ethics statement
All of the experimental animals used in this project were treated in strict accordance with Guidelines for the Use of Experimental Animals in the People’s Republic of China. The production of H. contortus in goats was approved by the Animal Ethics Committee of Hubei Province (permit SYXK-0029).
Nematode strains and their maintenance
The H. contortus Haecon 5 strain was maintained by serial passage in goats (helminth-free), which were infected intra-ruminally with 8000 iL3. Eggs, first-stage larvae (L1s), second-stage larvae (L2s) and iL3s were harvested or cultured from the faeces from infected goats, as described previously [35, 36]. L4s and adults were collected from the abomasa from infected goats euthanized at 8 and 30 days, respectively. These two developmental stages were washed extensively in phosphate-buffered saline (PBS; pH 7.4), and male and female worms were separated prior to storage at −80 °C. The N2 strain of C. elegans was obtained from the Caenorhabditis Genetics Center (CGC, University of Minnesota, USA) and maintained using standard procedures [37].
DNA and RNA preparation
Genomic DNA samples were extracted from mixed stages of C. elegans or L3s of H. contortus using the EasyPure Genomics DNA Kit (TransGen Biotech, China). Total RNA samples were isolated separately from egg, L1, L2, iL3, female and male fourth-stage larvae (L4s), and female and male adult stages of H. contortus using the TRIzol Plus Purification kit (Life Technologies, USA). RNA yields and quality were verified by spectrophotometric (NanoDrop Technologies) and by electrophoretic analysis, respectively. RNA was treated with RQ1-RNase-Free DNase (Promega, USA). Following isolation, nucleic acid samples were immediately frozen and stored at −80 °C.
Isolation of the Hc-pdk-1 gene and its upstream region
Guided by genomic and transcriptomic data for H. contortus (see [24]; GenBank accession no. AUUS00000000; SRA; accession nos. SRP027504 and SRP026668), we isolated the full-length Hc-pdk-1 gene and its cDNA (GenBank accession no. KU522003). The coding region was amplified and sequenced using the primer pair Hc-pdk-F and Hc-pdk-R (Additional file 1), and then cloned into the pMD-19 T vector (Takara, Japan). The gene sequence was obtained from the H. contortus genome [24], and exon-intron boundaries established. Part of the 5′-flanking region of the Hc-pdk-1 coding sequence was amplified from genomic DNA of H. contortus using the primers Hc-pdk-gw-1R and Hc-pdk-gw-2R (Additional file 1) using the GenomeWalker Universal Kit (Clontech, USA; according to the protocol provided) and sequenced (GenBank accession no. KU522003). Subsequently, the entire upstream sequence was amplified using the primer set Hc-pdk-pro-F/Hc-pdk-gfp-R (Additional file 1), cloned into the pMD-19T vector and sequenced in both directions (Sangon Biotech, Shanghai).
Bioinformatic analyses
Nucleotide sequences were assembled using the program CAP3 (http://bio.ifom-ieo-campus.it/) and compared with those in non-redundant databases using the BLAST v.2.0 suite of programs from the National Center for Biotechnology Information (NCBI) (http://www.ncbi.nlm.nih.gov/BLAST), the Sanger Centre (www.sanger.ac.uk) and the Parasite Genome database (www.ebi.ac.uk) to confirm the identity of genes isolated. Individual cDNAs were conceptually translated using the selection “translate”, available at http://bioinformatics.org/. Protein motifs were identified by scanning the databases Pfam (www.sanger.ac.uk/Software/Pfam) and PROSITE (www.expasy.ch/tools/scnpsit1.html). Signal sequences were predicted using SignalP v.2.0 [38], available at the Center for Biological Sequence Analysis (www.cbs.dtu.dk/services/SignalP). Amino acid sequences were aligned using the program Clustal W [39] and adjusted manually. Promoter elements in the 5′-genomic region upstream of the start codon (ATG) of Hc-pdk-1 were predicted using the transcription element search system (PLACE; available at http://www.dna.affrc.go.jp/PLACE/signalscan.html).
The predicted amino acid sequences of Hc-PDK-1 and homologues from other invertebrates (nematodes and Drosophila melanogaster) and vertebrates (human and Xenopus laevis) were aligned and subjected to phylogenetic analyses. These analyses were conducted using the neighbor-joining (NJ), maximum parsimony (MP) and maximum likelihood (ML) methods, respectively, based on the Jones-Taylor-Thornton (JTT) model [40]. Confidence limits were assessed using a bootstrap procedure employing 1000 pseudo-replicates for NJ, MP and ML trees and other settings were set according to default values in MEGA v.6.0 [40]. A 50 % cut-off value was implemented for the consensus tree.
Transcript abundance based on RNA-seq analysis
The abundances of Hc-pdk-1 transcripts in different developmental stages of H. contortus were assessed using publicly available RNA-seq data (Haecon 5 strain, Australia; [24]). Stages evaluated were eggs, L1, L2, L3, L4 (female and male) and adults (female and male) [24]. Levels of transcription in these stages were estimated and expressed as fragments per kilobase of coding exon per million mapped reads (FPKM), as described previously [24].
Transformation constructs
Two constructs Hc-pdk-1p (2975 bp)::Hc-pdk-1 (24 bp)::gfp::Ce-unc-54 t and Ce-pdk-1p (2958 bp)::Ce-pdk-1 (80 bp)::gfp::Ce-unc-54 t (designated pL-Hcpdk and pL-Cepdk, respectively) were made (Additional file 2). In brief, the putative promoter (2999 bp) was amplified from genomic DNA of H. contortus and cloned into the pPV199 vector [34] using the NovoRec PCR One-Step Directed Cloning kit (Novoprotein Scientific Inc., China), based on homologous recombination, employing primers Hc-pdk-199pro-F and Hc-pdk-gfp-R (Additional file 1). The homologous promoter region (3038 bp) was also amplified from genomic DNA of C. elegans and cloned into the pPV199 vector (BamH1 = B and Age1 = A sites) employing primer pair Ce-pdk-bam-3F and Ce-pdk-bam-4R (Additional file 1).
DNA transformation of C. elegans
A standard gonad microinjection method was performed in C. elegans, as described previously [41]. Briefly, the test constructs (pL-Hcpdk and pL-Cepdk) and plasmid pRF4 containing the marker gene, rol-6, were co-injected at final concentrations of 20 ng/μl and 80 ng/μl, respectively. Microinjected worms were reared on Nematode Growth Medium (NGM) plates on Escherichia coli OP50 lawns and maintained at 20 °C. Transformants were picked from F1 progeny, based on “right-roller” and green fluorescence protein (GFP) phenotypes, and re-plated. For the detection of spatio-temporal gene expression, transformants were anaesthetized with 10 mM levamisole, steadied on a 2 % agar pad, and assessed for GFP expression using a stereomicroscope with co-axial fluorescence, and a compound fluorescence microscope equipped with differential interference contrast (DIC) optics and a camera (Olympus BX51 Japan).
Results
Characterization of cDNA and phylogenetic analysis of amino acid sequence data
The Hc-pdk-1 cDNA is 1731 bp in length and encodes a protein (Hc-PDK-1) of 576 amino acids, which has 35–52 % similarity to homologs from C. elegans, Ascaris suum, Loa loa, S. stercoralis and Trichinella spiralis as well as Homo sapiens and Drosophila melanogaster. Hc-PDK-1 consists of two functional elements, namely the catalytic and pleckstrin homology (PH) domains (Fig. 1). Additionally, phospho ELM BLAST analysis (http://phospho.elm.eu.org/pELMBlastSearch.html) of Hc-PDK-1 predicted two phosphorylation sites (Thr226/Tyr229), inferred to play a critical role in cell signalling by phosphorylation [42]. The NetNES 1.1 Server (http://www.cbs.dtu.dk/services/NetNES/) predicted that Hc-PDK-1 contained a nuclear export sequence (NES) consensus, L-x(2,3)-[LIVFM]-x(2,3)-L-x-[LI], with four hydrophobic amino acid residues, which is required for the export of PDK-1 from the nucleus to the cytoplasm via nuclear transport (Fig. 1) [43]. For Hc-PDK-1 and other PDK-1s, an invariant amino acid (= Trp513) is shared by sequences of members of the PH domain family (see Fig. 1). The predicted Hc-PDK-1 protein sequence was aligned with 11 PDK-1 homologs from eight nematodes and three eukaryotes, and then subjected to phylogenetic analyses (Fig. 2). There was concordance in topology among the MP, ML and NJ trees, which showed that Hc-PDK-1 has a close relationship with homologs from C. elegans and C. briggsae (Fig. 2; nodal support: 98 %).
Fig. 1.

Alignment of the predicted amino acid sequence of Haemonchus contortus 3-phosphoinositide-dependent protein kinase (Hc-PDK-1) with those of Caenorhabditis elegans (Ce-PDK-1; NP_001024742), Drosophila melanogaster (DSTPK61; Y07908) and Homo sapiens (Hs-PDK-1; AF017995). Functional domains are boxed, with coloured backgrounds: catalytic domain (light blue), nuclear export sequence (yellow) and pleckstrin homology domain (PH; light orange). An invariant tryptophan (W513) is coloured in grey, which is highly conserved in the PH domain and might be involved in interaction(s) with downstream signalling proteins [42]
Fig. 2.
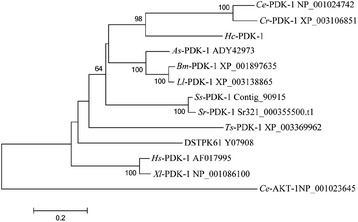
Neighbor-joining tree showing the relationship of Haemonchus contortus 3-phosphoinositide-dependent protein kinase (Hc-PDK-1) with related kinases. The tree was constructed using the Jones-Tayloe-Thornton model in the program MEGA v.6.0. Bootstrap values (>50 %) are shown above or below the branches (1000 iterations). The PDK-1 s of eight nematodes (Caenorhabditis elegans, Ce-PDK-1; C. remani, Cr-PDK-1; Ascaris sum, As-PDK-1; Brugia malayi, Bm-PDK-1; Loa loa, Ll-PDK-1; Strongyloides stercoralis, Ss-PDK-1; S. ratti, Sr-PDK-1; Trichinella spiralis, Ts-PDK-1) and three organisms other than nematodes (Drosophila melanogaster, DSTPK61; Homo sapiens, Hs-PDK-1; Xenopus laevis, Xl-PDK-1). Caenorhabditis elegans Ce-AKT-1 was used as an outgroup. GeneBank accession numbers are listed to the right of individual species names
Genomic organization, putative promoter elements and transcription
We located the full-length gene of Hc-pdk-1 in the H. contortus genome [24] and identified two orthologs (designated Hc-pdk-1 and Hc-pdk-2). The gene Hc-pdk-1 was 14,488 bp in length and had 16 exons (48–174 bp) that abided by the GT-AG rule [44]. However, Hc-pdk-2 had no introns and very limited transcription, and was thus interpreted to represent a pseudogene or the result of a genomic mis-assembly. Compared with homologs from C. elegans and C. briggase, Hc-pdk-1 had a complex exon/intron structure (Fig. 3). The 5′-genomic region upstream of the initiation codon ATG of the Hc-pdk-1 coding sequence was 2975 bp in length and included various predicted promoter elements, such as seven E-box (CANNTG) motifs, four TATA boxes, three inverse GATA (TTATC) motifs, one CAAT (CCAAT) and five inverse CAAT (ATTGG) motifs. Study of egg, L1, L2, L3, female L4, male L4, as well as female and male adult stages of H. contortus (Fig. 4) revealed a high level of transcription in L3 compared with other developmental stages, indicating that Hc-pdk-1 likely plays a core regulatory role in this stage.
Fig. 3.

Genomic structure of pdk-1 of Haemonchus contortus. Schematic diagram showing the genomic organization of pdk-1 of H. contortus (Hc-pdk-1), Strongyloides stercoralis (Ss-pdk-1) [54] and Caenorhabditis elegans (Ce-pdk-1) [16]. Black boxes represent exons. Lines between the exons represent introns. Start (ATG) and stop (TGA/TAA) codons are indicated
Fig. 4.
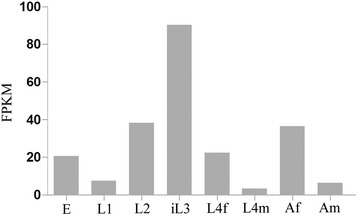
The abundance of Hc-pdk-1 transcripts in eight developmental stages or sexes of Haemonchus contortus. eggs (E), first-stage larvae (L1), second-stage larvae (L2), the infective third-stage larvae (iL3), female fourth-stage larvae (L4f), male fourth-stage larvae (L4m), adult females (Af) and adult males (Am). Transcript abundance is represented as fragments per kilobase of coding exon per million mapped reads (FPKM)
Expression pattern of the putative Hc-pdk-1 promoter in transgenic C. elegans
GFP expression driven by the Ce-pdk-1 promoter was localized to the head and tail neurons, intestine and pharynx, consistent with a pattern reported previously [16]. Despite some minor variation among individual transgenic lines, representative gene expression driven by the Hc-pdk-1 promoter was in head and tail neurons and the intestine (Fig. 5). Although the expression patterns for Ce-pdk-1 and Hc-pdk-1 were not identical, the anatomical localization of the proteins was similar (i.e. to intestine and head/tail neurons). This pattern of GFP expression was observed in all developmental stages of C. elegans transformed with p L-Hcpdk, including eggs, larvae and adults (data not shown).
Fig. 5.
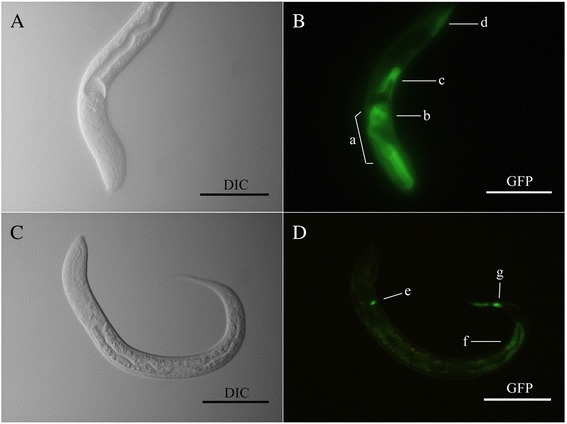
Representative expression patterns displayed in Caenorhabditis elegans using two GFP constructs, pL-Cepdk and pL-Hcpdk. Panels a and b Differential interference contrast (DIC) and fluorescence images of a N2 (wide type) L3 using the construct Ce-pdk-1 p::gfp (pL-Cepdk), respectively. GFP reporter expression was present in head neurons (a), pharynx (b), intestine (c) and hypodermal cells (d). Panels c and d DIC and fluorescence images showing the expression of construct Haemonchus contortus Hc-pdk-1 p::gfp (pL-Hcpdk) in the third larval stage (L3) of C. elegans (N2 strain). GFP reporter expression was present in head and tail neurons (e and g), and intestine (f). Scale bar = 50 μm
Discussion
The 3-phosphoinositide-dependent kinase 1 (PDK-1) is a member of the AGC family of serine and threonine kinases, and plays critical roles in physiological processes, such as metabolism, growth, proliferation and/or survival [45]. The deletion of pdk-1 is lethal in yeast [46], D. melanogaster [47] and mice [48, 49], showing that this molecule is essential for the normal embryonic development. In C. elegans, an insulin-like signalling pathway involving PDK-1 regulates development, metabolism and longevity [16]. Loss-of-function mutation of pdk-1 in C. elegans results in constitutive dauer arrest and increased lifespan [16], and the dauer stage shows similar characteristics to iL3 of parasitic nematodes [12].
Recently, homologs of genes and transcripts encoding intermediates of the insulin-like signalling pathway have been identified in parasitic nematodes. These components include the insulin-like receptor DAF-2 [29, 50], the PI3Ks protein kinase AGE-1 [30, 51] and the forkhead transcription factor DAF-16 [28, 34], suggesting that similar molecular mechanisms exist in both C. elegans and some parasitic nematodes. In the present study, the Hc-pdk-1 gene was identified in genomic and transcriptomic data sets for H. contortus. By contrast, the Hc-pdk-2 sequence (without introns) was found only in the genome, but not in the transcriptome, suggesting that it is a pseudogene or a misassembled genomic sequence tract.
Sequence and structural analyses showed that Hc-PDK-1 possesses an N-terminal catalytic domain, a nuclear export sequence and a C-terminal pleckstrin homology (PH) domain [16, 45]. PDK-1 is constitutively associated with a homo-dimeric complex through a PH domain interaction of two PDK-1 monomers; the PH domain interaction likely has significant roles in the regulation of Akt phosphorylation [52]. Hc-PDK-1 contains two predicted phosphorylation sites within the catalytic domain, which might be subjected to autophosphorylation [53]. Phylogenetic analyses of inferred amino acid sequence data grouped Hc-PDK-1 with the homologs from C. elegans and C. remanei. Additionally, PDK-1 homologs exist widely in parasitic nematodes, including A. suum, L. loa and S. stercoralis (confirmed by searching the National Center for Biotechnology Information databases; results not shown). These findings suggest that PDK-1 is relatively conserved and has functional similarity in various parasitic nematodes.
During key developmental transitions, including recovery of dauer larvae of C. elegans, and the switch from the free-living to the parasitic stage of H. contortus, many genes, such as Ce-daf-7 (encoding TGF-β ligands) and the insulin-like peptide-encoding genes, are transcriptionally regulated. The current study showed that Hc-pdk-1 transcripts are present throughout the life cycle, but have a considerably higher abundance in iL3 than other key developmental stages, which is consistent with the transcription profile of Ss-pdk-1 in Strongyloides stercoralis [54]. This up-regulation is interpreted to relate to the switch from the free-living to the parasitic stage, in accordance with previously transcriptional evidence for Hc-daf-2 and Hc-age-1 [29, 30], and with a reduced metabolic rate in the L3 stage [55].
The expression of Hc-pdk-1 predominated in the neurons and intestine of transgenic C. elegans larvae, which is similar to the spatio-temporal distribution of Ce-PDK-1 [16], where GFP was expressed under the Ce-pdk-1 promoter in pharynx, intestine and head/tail neurons [16]. This knowledge not only contributes to understanding gene expression in time and in space, but might also assist in the prediction of protein-protein interactomes [56]. Therefore, the similar gene expression profiles of Hc-pdk-1- and Ce-pdk-1-based reporters in head/tail neurons and the intestine suggest similar regulatory functions for Hc-pdk-1 and Ce-pdk-1 [16]. Amphidial neurons play crucial roles in regulating the entry into and exit from dauer in C. elegans [57], and arrest and developmental activation of iL3 in the parasite S. stercoralis [58, 59]. In addition, the intestine secretes important proteins for the regulation of oocyte development [60]; this information further supports some functional similarities between Ce-pdk-1 and Hc-pdk-1.
The “dauer hypothesis” proposes that the dauer larvae of the free-living nematode C. elegans are physiologically similar to the iL3s of parasitic nematodes [12]. Many scientists posited that this process might be regulated through similar signalling mechanisms. In parasitic nematodes, such as A. caninum, A. ceylanicum and S. stercoralis, LY294002, a specific inhibitor of PI3K, effectively blocks the resumption of feeding [51, 61], representing a phenotypic marker. In S. stercoralis, genes encoding the forkhead transcription factor Ss-DAF-16 have an anatomical expression pattern similar to their C. elegans orthologs. Furthermore, transgenes encoding Ss-DAF-16b with phospho-null and phospho-mimetic mutations at crucial AKT phosphorylation sites gave products with nuclear and cytoplasmic localizations, respectively [62]. Finally, mutant constructs of Ss-daf-16b generated a dominant-negative phenotype, including the developmental alterations of the larval intestine and pharynx, failing to arrest of transgenic larvae in the infective stage [62]. Results of studies employing C. elegans as a genetic surrogate indicate that central components of insulin-like signalling, such as daf-16, daf-2, age-1 and daf-12, from parasitic nematodes [28–30, 32, 34, 63] have similar functional characteristics to their orthologs in C. elegans. This evidence supports the “dauer hypothesis” or “daf-c paradigm”, in which the activation of the L3 stage in parasitic nematodes and the recovery from dauer in free-living nematodes are governed by relatively conserved molecular mechanisms. Significantly, the CRISPR/Cas9 technology has been applied in the “model parasitic nematode” Pristionchus pacificus [64]. This technology might allow developmental processes of parasitic nematodes to be addressed in the near future.
Conclusion
In the present study, we investigated a 3-phosphoinositide-dependent protein kinase-encoding gene, Hc-pdk-1, in the parasitic nematode H. contortus. We isolated and characterized the cDNA, genomic DNA and upstream (predicted) promoter elements of Hc-pdk-1, and assessed transcription levels of this gene in eight developmental stages/sexes. We also compared the anatomical expression patterns of Hc-pdk-1 and Ce-pdk-1, and predict similar functions for these genes/gene products. Taken together, these findings provide a first glimpse of the involvement of Hc-pdk-1 in the insulin-like signalling pathway in H. contortus.
Acknowledgements
This study was supported by “National Natural Science Foundation of China (NSFC)” (Grant no. 31172310), the National Key Basic Research Program (973 program) of China (Grant No. 2015CB150300) and “Special Fund for Agro-scientific Research in the Public Interest, China” (Grant no. 201303037) to MH, by a grant from The National Institute of Health (NIH), USA (AI-50688) to JBL and by funds from the National Health and Medical Research Council (NHMRC) and the Australian Research Council (ARC) of Australia to RBG. This study was also supported by a Victoria Life Sciences Computation Initiative (VLSCI; grant number VR0007) on its Peak Computing Facility at the University of Melbourne, an initiative of the Victorian Government, Australia (RBG).
Abbreviations
- DIC
Differential interference contrast optics
- GFP
Green fluorescence protein
- JTT
Jones-Taylor-Thornton model
- L3s
The third-stage larvae
- ML
Maximum likelihood
- MP
Maximum parsimony
- NGM
Nematode growth medium
- NJ
Neighbour-joining
- PCR
Polymerase chain reaction
- RACE
Rapid amplification of cDNA ends
- UTR
Untranslated region
Additional files
Primers used to isolate Hc-pdk-1 of Haemonchus contortus and to make constructs for green fluorescent protein (GFP) localization in Caenorhabditis elegans . (DOC 32 kb)
Cloning strategy for reporter constructs. The constructs containing the Caenorhabditis elegans Ce-pdk-1 and the Haemonchus contortus Hc-pdk-1 promoters (pL-Cepdk and pL-Hcpdk) were made in the vector pPV199 [34]. Briefly, the Ce-pdk-1 promoter region was cloned into pPV199 (BamH1 and Age1 sites). The Hc-pdk-1 promoter region was cloned into pPV199 by homologous recombination. (DOC 127 kb)
Footnotes
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
MH, JBL and FCL conceived the project. FCL carried out laboratory work. FCL, PKK and MH performed the data analyses. FCL and MH interpreted the data. RBG, JBL, LH, WDD, FYY, RZ, YQZ, JLZ contributed reagents/materials/analysis tools. FCL, RBG, JBL and MH wrote the manuscript, and all authors read and approved the final manuscript.
Contributor Information
Fa-Cai Li, Email: li78561270@163.com.
Robin B. Gasser, Email: robinbg@unimelb.edu.au
James B. Lok, Email: jlok@vet.upenn.edu
Pasi K. Korhonen, Email: pasi.korhonen@unimelb.edu.au
Li He, Email: 463771946@qq.com.
Wen-Da Di, Email: 732471543@qq.com.
Fang-Yuan Yin, Email: yinfangyuan96@126.com.
Rui Zhou, Email: rzhou@mail.hzau.edu.cn.
Yan-Qin Zhou, Email: yanqinzhou@mail.hzau.edu.cn.
Jun-Long Zhao, Email: zhaojunlong@mail.hzau.edu.cn.
Min Hu, Email: mhu@mail.hzau.edu.cn.
References
- 1.Wolstenholme AJ, Fairweather I, Prichard R, von Samson-Himmelstjerna G, Sangster NC. Drug resistance in veterinary helminths. Trends Parasitol. 2004;20:469–76. doi: 10.1016/j.pt.2004.07.010. [DOI] [PubMed] [Google Scholar]
- 2.Geary TG, Woo K, McCarthy JS, Mackenzie CD, Horton J, Prichard RK, et al. Unresolved issues in anthelmintic pharmacology for helminthiases of humans. Int J Parasitol. 2010;40:1–13. doi: 10.1016/j.ijpara.2009.11.001. [DOI] [PubMed] [Google Scholar]
- 3.Papadopoulos E, Gallidis E, Ptochos S. Anthelmintic resistance in sheep in Europe: a selected review. Vet Parasitol. 2012;189:85–8. doi: 10.1016/j.vetpar.2012.03.036. [DOI] [PubMed] [Google Scholar]
- 4.Kaplan RM, Vidyashankar AN. An inconvenient truth: global worming and anthelmintic resistance. Vet Parasitol. 2012;186:70–8. doi: 10.1016/j.vetpar.2011.11.048. [DOI] [PubMed] [Google Scholar]
- 5.Blaxter ML, Raghavan N, Ghosh I, Guiliano D, Lu W, Williams SA, et al. Genes expressed in Brugia malayi infective third stage larvae. Mol Biochem Parasitol. 1996;77:77–93. [DOI] [PubMed]
- 6.Cantacessi C, Campbell BE, Young ND, Jex AR, Hall RS, Presidente PJ, et al. Differences in transcription between free-living and CO2-activated third-stage larvae of Haemonchus contortus. BMC Genomics. 2010;11:266. doi: 10.1186/1471-2164-11-266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Hu PJ. Dauer. In The C. elegans Research Community. Edited by Wormbook. Available from: http://www.wormbook.org/chapters/www_dauer/dauer.html
- 8.Golden JW, Riddle DL. The Caenorhabditis elegans dauer larva: developmental effects of pheromone, food, and temperature. Dev Biol. 1984;102:368–78. doi: 10.1016/0012-1606(84)90201-X. [DOI] [PubMed] [Google Scholar]
- 9.Blaxter ML, De Ley P, Garey JR, Liu LX, Scheldeman P, Vierstraete A, et al. A molecular evolutionary framework for the phylum Nematoda. Nature. 1998;392:71–5. doi: 10.1038/32160. [DOI] [PubMed] [Google Scholar]
- 10.Blaxter M. Caenorhabditis elegans is a nematode. Science. 1998;282:2041–6. doi: 10.1126/science.282.5396.2041. [DOI] [PubMed] [Google Scholar]
- 11.Burglin TR, Lobos E, Blaxter ML. Caenorhabditis elegans as a model for parasitic nematodes. Int J Parasitol. 1998;28:395–411. doi: 10.1016/S0020-7519(97)00208-7. [DOI] [PubMed] [Google Scholar]
- 12.Crook M. The dauer hypothesis and the evolution of parasitism: 20 years on and still going strong. Int J Parasitol. 2014;44:1–8. doi: 10.1016/j.ijpara.2013.08.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hotez P, Hawdon J, Schad GA. Hookworm larval infectivity, arrest and amphiparatenesis: the Caenorhabditis elegans Daf-c paradigm. Parasitol Today. 1993;9:23–6. doi: 10.1016/0169-4758(93)90159-D. [DOI] [PubMed] [Google Scholar]
- 14.Kimura KD, Tissenbaum HA, Liu Y, Ruvkun G. daf-2, an insulin receptor-like gene that regulates longevity and diapause in Caenorhabditis elegans. Science. 1997;277:942–6. doi: 10.1126/science.277.5328.942. [DOI] [PubMed] [Google Scholar]
- 15.Vanhaesebroeck B, Leevers SJ, Panayotou G, Waterfield MD. Phosphoinositide 3-kinases: a conserved family of signal transducers. Trends Biochem Sci. 1997;22:267–72. doi: 10.1016/S0968-0004(97)01061-X. [DOI] [PubMed] [Google Scholar]
- 16.Paradis S, Ailion M, Toker A, Thomas JH, Ruvkun G. A PDK1 homolog is necessary and sufficient to transduce AGE-1 PI3 kinase signals that regulate diapause in Caenorhabditis elegans. Genes Dev. 1999;13:1438–52. doi: 10.1101/gad.13.11.1438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Paradis S, Ruvkun G. Caenorhabditis elegans Akt/PKB transduces insulin receptor-like signals from AGE-1 PI3 kinase to the DAF-16 transcription factor. Genes Dev. 1998;12:2488–98. doi: 10.1101/gad.12.16.2488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lin K, Dorman JB, Rodan A, Kenyon C. daf-16: An HNF-3/forkhead family member that can function to double the life-span of Caenorhabditis elegans. Science. 1997;278:1319–22. doi: 10.1126/science.278.5341.1319. [DOI] [PubMed] [Google Scholar]
- 19.Ogg S, Paradis S, Gottlieb S, Patterson GI, Lee L, Tissenbaum HA, et al. The Fork head transcription factor DAF-16 transduces insulin-like metabolic and longevity signals in C. elegans. Nature. 1997;389:994–9. doi: 10.1038/40194. [DOI] [PubMed] [Google Scholar]
- 20.Ogg S, Ruvkun G. The C. elegans PTEN homolog, DAF-18, acts in the insulin receptor-like metabolic signaling pathway. Mol Cell. 1998;2:887–93. doi: 10.1016/S1097-2765(00)80303-2. [DOI] [PubMed] [Google Scholar]
- 21.Mukhopadhyay A, Oh SW, Tissenbaum HA. Worming pathways to and from DAF-16/FOXO. Exp Gerontol. 2006;41:928–34. doi: 10.1016/j.exger.2006.05.020. [DOI] [PubMed] [Google Scholar]
- 22.Yen K, Narasimhan SD, Tissenbaum HA. DAF-16/Forkhead box O transcription factor: many paths to a single Fork(head) in the road. Antioxid Redox Signal. 2011;14:623–34. doi: 10.1089/ars.2010.3490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Laing R, Kikuchi T, Martinelli A, Tsai IJ, Beech RN, Redman E, et al. The genome and transcriptome of Haemonchus contortus, a key model parasite for drug and vaccine discovery. Genome Biol. 2013;14:R88. doi: 10.1186/gb-2013-14-8-r88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Schwarz EM, Korhonen PK, Campbell BE, Young ND, Jex AR, Jabbar A, et al. The genome and developmental transcriptome of the strongylid nematode Haemonchus contortus. Genome Biol. 2013;14:R89. doi: 10.1186/gb-2013-14-8-r89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Britton C, Murray L. Using Caenorhabditis elegans for functional analysis of genes of parasitic nematodes. Int J Parasitol. 2006;36:651–9. doi: 10.1016/j.ijpara.2006.02.010. [DOI] [PubMed] [Google Scholar]
- 26.Gilleard JS. The use of Caenorhabditis elegans in parasitic nematode research. Parasitology. 2004;128(Suppl 1):S49–70. doi: 10.1017/S003118200400647X. [DOI] [PubMed] [Google Scholar]
- 27.Hashmi S, Tawe W, Lustigman S. Caenorhabditis elegans and the study of gene function in parasites. Trends Parasitol. 2001;17:387–93. doi: 10.1016/S1471-4922(01)01986-9. [DOI] [PubMed] [Google Scholar]
- 28.Hu M, Lok JB, Ranjit N, Massey HC, Jr, Sternberg PW, Gasser RB. Structural and functional characterisation of the fork head transcription factor-encoding gene, Hc-daf-16, from the parasitic nematode Haemonchus contortus (Strongylida) Int J Parasitol. 2010;40:405–15. doi: 10.1016/j.ijpara.2009.09.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Li F, Lok JB, Gasser RB, Korhonen PK, Sandeman MR, Shi D, et al. Hc-daf-2 encodes an insulin-like receptor kinase in the barber’s pole worm, Haemonchus contortus, and restores partial dauer regulation. Int J Parasitol. 2014;44:485–96. doi: 10.1016/j.ijpara.2014.03.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Li FC, Gasser RB, Lok JB, Korhonen PK, Wang YF, Yin F, et al. Exploring the role of two interacting phosphoinositide 3-kinases of Haemonchus contortus. Parasit Vectors. 2014;7:498. doi: 10.1186/s13071-014-0498-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Gelmedin V, Brodigan T, Gao X, Krause M, Wang Z, Hawdon JM. Transgenic C. elegans dauer larvae expressing hookworm phospho null DAF-16/FoxO exit dauer. PloS One. 2011;6:e25996. doi: 10.1371/journal.pone.0025996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Gao X, Frank D, Hawdon JM. Molecular cloning and DNA binding characterization of DAF-16 orthologs from Ancylostoma hookworms. Int J Parasitol. 2009;39:407–15. doi: 10.1016/j.ijpara.2008.09.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Massey HC, Jr, Nishi M, Chaudhary K, Pakpour N, Lok JB. Structure and developmental expression of Strongyloides stercoralis fktf-1, a proposed ortholog of daf-16 in Caenorhabditis elegans. Int J Parasitol. 2003;33:1537–44. doi: 10.1016/S0020-7519(03)00205-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Massey HC, Jr, Bhopale MK, Li X, Castelletto M, Lok JB. The fork head transcription factor FKTF-1b from Strongyloides stercoralis restores DAF-16 developmental function to mutant Caenorhabditis elegans. Int J Parasitol. 2006;36:347–52. doi: 10.1016/j.ijpara.2005.11.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Cox DD, Todd AC. Survey of gastrointestinal parasitism in Wisconsin dairy cattle. J Am Vet Med Assoc. 1962;141:706–9. [PubMed] [Google Scholar]
- 36.Nikolaou S, Hartman D, Presidente PJ, Newton SE, Gasser RB. HcSTK, a Caenorhabditis elegans PAR-1 homologue from the parasitic nematode, Haemonchus contortus. Int J Parasitol. 2002;32:749–58. doi: 10.1016/S0020-7519(02)00008-5. [DOI] [PubMed] [Google Scholar]
- 37.Stiernagle T. Maintenance of C. elegans. In The C. elegans Research Community. Edited by Wormbook. Available from: http://www.wormbook.org/chapters/www_strainmaintain/strainmaintain.html
- 38.Nielsen H, Engelbrecht J, Brunak S, von Heijne G. A neural network method for identification of prokaryotic and eukaryotic signal peptides and prediction of their cleavage sites. Int J Neural Syst. 1997;8:581–99. doi: 10.1142/S0129065797000537. [DOI] [PubMed] [Google Scholar]
- 39.Thompson JD, Higgins DG, Gibson TJ. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994;22:4673–80. doi: 10.1093/nar/22.22.4673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol Biol Evol. 2013;30:2725–9. doi: 10.1093/molbev/mst197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Mello CC, Kramer JM, Stinchcomb D, Ambros V. Efficient gene transfer in C. elegans: extrachromosomal maintenance and integration of transforming sequences. EMBO J. 1991;10:3959–70. doi: 10.1002/j.1460-2075.1991.tb04966.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Alessi DR, Deak M, Casamayor A, Caudwell FB, Morrice N, Norman DG, et al. 3-Phosphoinositide-dependent protein kinase-1 (PDK1): structural and functional homology with the Drosophila DSTPK61 kinase. Curr Biol. 1997;7:776–89. doi: 10.1016/S0960-9822(06)00336-8. [DOI] [PubMed] [Google Scholar]
- 43.la Cour T, Kiemer L, Molgaard A, Gupta R, Skriver K, Brunak S. Analysis and prediction of leucine-rich nuclear export signals. Protein Eng Des Sel. 2004;17:527–36. doi: 10.1093/protein/gzh062. [DOI] [PubMed] [Google Scholar]
- 44.Breathnach R, Chambon P. Organization and expression of eucaryotic split genes coding for proteins. Annu Rev Biochem. 1981;50:349–83. doi: 10.1146/annurev.bi.50.070181.002025. [DOI] [PubMed] [Google Scholar]
- 45.Fyffe C, Falasca M. 3-Phosphoinositide-dependent protein kinase-1 as an emerging target in the management of breast cancer. Cancer Manag Res. 2013;5:271–80. doi: 10.2147/CMAR.S35026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Niederberger C, Schweingruber ME. A Schizosaccharomyces pombe gene, ksg1, that shows structural homology to the human phosphoinositide-dependent protein kinase PDK1, is essential for growth, mating and sporulation. Mol Gen Genet. 1999;261:177–83. doi: 10.1007/s004380050955. [DOI] [PubMed] [Google Scholar]
- 47.Cho KS, Lee JH, Kim S, Kim D, Koh H, Lee J, et al. Drosophila phosphoinositide-dependent kinase-1 regulates apoptosis and growth via the phosphoinositide 3-kinase-dependent signaling pathway. Proc Natl Acad Sci U S A. 2001;98:6144–9. doi: 10.1073/pnas.101596998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Artunc F, Rexhepaj R, Volkl H, Grahammer F, Remy C, Sandulache D, et al. Impaired intestinal and renal glucose transport in PDK-1 hypomorphic mice. Am J Physiol Regul Integr Com Physiol. 2006;291:R1533–8. doi: 10.1152/ajpregu.00024.2006. [DOI] [PubMed] [Google Scholar]
- 49.Rexhepaj R, Grahammer F, Volkl H, Remy C, Wagner CA, Sandulache D, et al. Reduced intestinal and renal amino acid transport in PDK1 hypomorphic mice. FASEB J. 2006;20:2214–22. doi: 10.1096/fj.05-5676com. [DOI] [PubMed] [Google Scholar]
- 50.Massey HC, Jr, Ranjit N, Stoltzfus JD, Lok JB. Strongyloides stercoralis daf-2 encodes a divergent ortholog of Caenorhabditis elegans DAF-2. Int J Parasitol. 2013;43:515–20. doi: 10.1016/j.ijpara.2013.01.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Stoltzfus JD, Massey HC, Jr, Nolan TJ, Griffith SD, Lok JB. Strongyloides stercoralis age-1: a potential regulator of infective larval development in a parasitic nematode. PLoS One. 2012;7:e38587. doi: 10.1371/journal.pone.0038587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Masters TA, Calleja V, Armoogum DA, Marsh RJ, Applebee CJ, Laguerre M, et al. Regulation of 3-phosphoinositide-dependent protein kinase 1 activity by homodimerization in live cells. Sci Signal. 2010;3:ra78. doi: 10.1126/scisignal.2000738. [DOI] [PubMed] [Google Scholar]
- 53.Wick MJ, Ramos FJ, Chen H, Quon MJ, Dong LQ, Liu F. Mouse 3-phosphoinositide-dependent protein kinase-1 undergoes dimerization and trans-phosphorylation in the activation loop. J Biol Chem. 2003;278:42913–9. doi: 10.1074/jbc.M304172200. [DOI] [PubMed] [Google Scholar]
- 54.Stoltzfus JD, Minot S, Berriman M, Nolan TJ, Lok JB. RNAseq analysis of the parasitic nematode Strongyloides stercoralis reveals divergent regulation of canonical dauer pathways. PLoS Negl Trop Dis. 2012;6:e1854. doi: 10.1371/journal.pntd.0001854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Nikolaou S, Gasser RB. Prospects for exploring molecular developmental processes in Haemonchus contortus. Int J Parasitol. 2006;36:859–68. doi: 10.1016/j.ijpara.2006.04.007. [DOI] [PubMed] [Google Scholar]
- 56.Dupuy D, Bertin N, Hidalgo CA, Venkatesan K, Tu D, Lee D, et al. Genome-scale analysis of in vivo spatiotemporal promoter activity in Caenorhabditis elegans. Nature Biotechnol. 2007;25:663–8. doi: 10.1038/nbt1305. [DOI] [PubMed] [Google Scholar]
- 57.Cornils A, Gloeck M, Chen Z, Zhang Y, Alcedo J. Specific insulin-like peptides encode sensory information to regulate distinct developmental processes. Development. 2011;138:1183–93. doi: 10.1242/dev.060905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Ashton FT, Bhopale VM, Holt D, Smith G, Schad GA. Developmental switching in the parasitic nematode Strongyloides stercoralis is controlled by the ASF and ASI amphidial neurons. J Parasitol. 1998;84:691–5. doi: 10.2307/3284571. [DOI] [PubMed] [Google Scholar]
- 59.Ashton FT, Zhu X, Boston R, Lok JB, Schad GA. Strongyloides stercoralis: Amphidial neuron pair ASJ triggers significant resumption of development by infective larvae under host-mimicking in vitro conditions. Exp Parasitol. 2007;115:92–7. doi: 10.1016/j.exppara.2006.08.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Kimble J, Sharrock WJ. Tissue-specific synthesis of yolk proteins in Caenorhabditis elegans. Dev Biol. 1983;96:189–96. doi: 10.1016/0012-1606(83)90322-6. [DOI] [PubMed] [Google Scholar]
- 61.Brand A, Hawdon JM. Phosphoinositide-3-OH-kinase inhibitor LY294002 prevents activation of Ancylostoma caninum and Ancylostoma ceylanicum third-stage infective larvae. Int J Parasitol. 2004;34:909–14. doi: 10.1016/j.ijpara.2004.04.003. [DOI] [PubMed] [Google Scholar]
- 62.Castelletto ML, Massey HC, Jr, Lok JB. Morphogenesis of Strongyloides stercoralis infective larvae requires the DAF-16 ortholog FKTF-1. PLoS Pathog. 2009;5:e1000370. doi: 10.1371/journal.ppat.1000370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Wang Z, Zhou XE, Motola DL, Gao X, Suino-Powell K, Conneely A, et al. Identification of the nuclear receptor DAF-12 as a therapeutic target in parasitic nematodes. Proc Natl Acad Sci U S A. 2009;106:9138–43. doi: 10.1073/pnas.0904064106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Witte H, Moreno E, Rodelsperger C, Kim J, Kim JS, Streit A, et al. Gene inactivation using the CRISPR/Cas9 system in the nematode Pristionchus pacificus. Dev Genes Evol. 2015;225:55–62. doi: 10.1007/s00427-014-0486-8. [DOI] [PubMed] [Google Scholar]