

Abstract
Global change, especially land‐use intensification, affects human well‐being by impacting the delivery of multiple ecosystem services (multifunctionality). However, whether biodiversity loss is a major component of global change effects on multifunctionality in real‐world ecosystems, as in experimental ones, remains unclear. Therefore, we assessed biodiversity, functional composition and 14 ecosystem services on 150 agricultural grasslands differing in land‐use intensity. We also introduce five multifunctionality measures in which ecosystem services were weighted according to realistic land‐use objectives. We found that indirect land‐use effects, i.e. those mediated by biodiversity loss and by changes to functional composition, were as strong as direct effects on average. Their strength varied with land‐use objectives and regional context. Biodiversity loss explained indirect effects in a region of intermediate productivity and was most damaging when land‐use objectives favoured supporting and cultural services. In contrast, functional composition shifts, towards fast‐growing plant species, strongly increased provisioning services in more inherently unproductive grasslands.
Keywords: Biodiversity–ecosystem functioning, ecosystem services, global change, land use, multifunctionality
Introduction
Understanding and managing ecosystem‐service delivery is of key importance for human well‐being (Millennium Ecosystem Assessment 2005). In grasslands, land‐use intensification is the most important global change affecting a range of ecosystem functions and services (Thébault et al. 2014). Intensive land use promotes a small set of provisioning services but at the cost of other ecosystem functions or services, which are reduced due to trade‐offs. In particular, land‐use intensification can strongly reduce biodiversity (Foley et al. 2005; Allan et al. 2014), which might threaten ecosystem services that depend on biodiversity for their delivery (Isbell et al. 2011; Cardinale et al. 2012).
A significant body of experimental evidence demonstrates that more diverse model ecosystems have higher levels of functioning than less diverse systems (Cardinale et al. 2012). Furthermore, high biodiversity has been shown to be especially important for simultaneously sustaining multiple ecosystem functions and services at high levels, a property known as multifunctionality (Hector & Bagchi 2007; Isbell et al. 2011). This has led to predictions that biodiversity loss caused by anthropogenic global change is affecting ‘real‐world’ ecosystem services (Cardinale et al. 2012). However, we lack strong evidence for negative effects of biodiversity loss on services in natural and semi‐natural ecosystems (Balvanera et al. 2014). Observational studies have found weakly positive (Maestre et al. 2012; Gamfeldt et al. 2013), neutral (Grace et al. 2007) or negative (Creed et al. 2009) relationships. This difference from experiments may be due to the greater complexity of real‐world ecosystems (Hillebrand & Matthiessen 2009), where biodiversity loss is non‐random and is caused by global change drivers that also affect ecosystem services directly (Balvanera et al. 2014). For example land‐use intensification can directly alter productivity by changing soil chemistry or causing disturbance (Laliberté & Tylianakis 2011) and can indirectly affect functioning by strongly reducing the diversity of many taxa that provide ecosystem services (Foley et al. 2005; Allan et al. 2014).
Biodiversity loss is often accompanied by shifts in the functional composition of the vegetation: intensive grassland management favours fast‐growing, acquisitive species over slow‐growing, conservative ones (Lavorel et al. 2010; de Vries et al. 2012). This shift towards fast‐growing species can alter numerous ecosystem functions and services, with an increase in biomass production and nutrient cycling rates expected (Lavorel & Grigulis 2012). While all of these direct and indirect effects of land‐use intensity (LUI) occur simultaneously, to date they have either been examined separately, e.g. studies focussing only on biodiversity effects, or they were examined as a whole, e.g. studies looking at the overall impact of land use on ecosystem services. However, to understand the importance of biodiversity loss as a component of global change impacts on ecosystem multifunctionality, it is crucial to quantify the relative strengths of direct land‐use effects and indirect effects mediated by biodiversity and functional composition.
An additional obstacle to the transfer of results from biodiversity experiments to real‐world ecosystems comes from oversimplified definitions of multifunctionality. To date multifunctionality research (e.g. Hector & Bagchi 2007; Isbell et al. 2011) has assumed that all ecosystem processes are equally important. While this is a reasonable starting point, such multifunctionality measures have recently been criticised (Bradford et al. 2014) and they may have little relevance to land managers, who typically seek to maximise a subset of functions and services (Reed et al. 2009; Goldstein et al. 2012). For instance livestock farmers may favour forage production whereas recreation agencies would favour cultural services, such as the high aesthetic appeal provided by flower‐rich grasslands (Lindemann‐Matthies et al. 2010; Junge et al. 2015). In addition, ecosystem service trade‐offs are commonplace, making it impossible to maximise all services simultaneously (Lavorel et al. 2010; Raudsepp‐Hearne et al. 2010; Goldstein et al. 2012; Bradford et al. 2014; Martin et al. 2014). Strong trade‐offs also imply that the relative importance of direct and indirect effects of land‐use intensification on multifunctionality depends on which services are desired.
The strength and direction of direct and indirect effects might also depend on environmental conditions and previous land‐use impacts on biodiversity and functional composition. Indirect effects can only occur where land‐use intensification alters plant communities, which may not be the case in very productive grasslands or in regions that have a long history of intensive land use (Cousins & Eriksson 2002). Land‐use intensification may also cause stronger losses of biodiversity, and larger shifts in functional composition, in less productive grasslands than in more productive ones (Rudmann‐Maurer et al. 2008). Understanding such context‐dependence in relationships between land use, biodiversity, functional composition and ecosystem services is essential if we are to accurately predict effects of global change on ecosystem services.
To investigate the relative strengths of direct and indirect (biodiversity and functional composition mediated) land‐use effects on multifunctionality, we analyse data on land use, biodiversity and 14 ecosystem functions and services from 150 grasslands in three regions, differing in the favourability of growing conditions (Table S1). We measured indicators of the final benefits of provisioning, regulating and cultural services as well as ecosystem functions (previously supporting services) that underlie the sustainability of these benefits (Millennium Ecosystem Assessment 2005) (Fig. 1 and Table S2). Hereafter, we refer to these collectively as ecosystem services. To address five scenarios representing realistic and contrasting management objectives (ranging from a focus on production to a focus on cultural services) we calculated five multifunctionality metrics, with different weightings for the 14 services (Fig. 1). We then adopted a three‐step approach to decompose land‐use effects on multifunctionality. Firstly, we examined overall LUI effects on multifunctionality. Next, we partitioned these overall effects into direct effects and indirect effects mediated by biodiversity (plant species richness). We looked initially at biodiversity loss because this has been the focus of numerous experimental studies (Cardinale et al. 2012). However, in a third step, we tested whether indirect effects were better explained by biodiversity loss or by shifts in plant functional composition, i.e. the abundance of conservative (slow growing) relative to acquisitive (fast‐growing) species. We also tested whether the nature of the indirect effects varied regionally and hypothesised that indirect effects would be better explained by biodiversity or functional composition depending on which responded most strongly to land‐use intensification.
Figure 1.
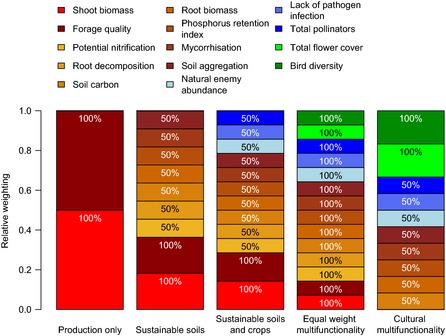
Ecosystem service weightings used to produce five measures of ecosystem multifunctionality according to scenarios representing different land‐use objectives. Scenarios are: (1) ‘production only’ with only provisioning services (forage production and quality); (2) ‘sustainable soils’ includes other functions promoting sustainable grass production; (3) ‘sustainable soils and crops’ additionally includes services benefitting surrounding croplands (pest and pathogen control and pollination); (4) ‘equal weight’ weights all processes equally and corresponds to classic multifunctionality measures; (5) ‘cultural multifunctionality’ weights cultural services highly, includes ecosystem functions associated with ecosystem health but does not include provisioning services (forage production and quality) or functions directly supporting production (potential nitrification and root decomposition). Weightings of 50% or 100% are indicated. Where we used different weightings, processes that were measures of final benefits for a particular scenario (e.g. forage production or aesthetic value) received weightings of 100% and functions which supported these were weighted at 50%.
Methods
Study design
The grassland plots are located in three regions of Germany and are part of the Biodiversity Exploratories project (www.biodiversity-exploratories.de; Fischer et al. 2010). The south‐west region is the UNESCO Biosphere Area Schwäbische Alb; the central region is in and around the National Park Hainich and the north‐east region is the UNESCO Biosphere Reserve Schorfheide‐Chorin. Plots in each region span the whole gradient of LUI found among Central European grasslands, whereas the regions cover a range of almost 3 °C in mean annual temperature and 500 to 1000 mm annual precipitation. The three regions differ substantially in geology and topography (Fischer et al. 2010): the south‐west grasslands are at the highest elevation, have the lowest mean annual temperature and are on steeper slopes, with shallower soils, which are more prone to drying in the summer and have lower phosphorus availability (see Table S1 for details). The north‐east region has the deepest, most fertile soils. The three regions therefore form a gradient in the favourability of growing conditions, from the least favourable in the south‐west to the most favourable in the north‐east (Table S1). The land‐use history also differs: the north‐east has the longest history of intensive land use and here many grasslands with current low intensity management are recovering from a period of historically high LUI. Historically grassland management in the south‐west has been the least intensive and intensification has only occurred more recently.
In each region 50 permanent grassland plots, 50 × 50 m, were established (150 in total). All plots had been grassland for at least 20 years prior to the start of the project in 2006.
Land use
Grasslands in all three regions were selected to cover a similarly large gradient in LUI, whilst minimising variation in confounding factors such as soil type and spatial distribution (Fischer et al. 2010). Land‐use intensification comprises increased fertilisation, mowing and grazing and was quantified using a questionnaire submitted annually to farmers and landowners (Fischer et al. 2010; Blüthgen et al. 2012). Grasslands could be grazed by cattle, horses or sheep and farmers reported the number of animals (converted to livestock units based on each species’ per capita impact) and the duration of grazing in each plot (G) (Fischer et al. 2010). Farmers also reported the number of annual mowing events (M) and the rate of nitrogen fertiliser addition (F; organic and inorganic). We used land‐use data from 2006 to 2010, averaged across years. For each plot an individual LUI component (F, M, G) was standardised relative to its mean across all three regions and all 5 years (standardising within regions gives very similar results Blüthgen et al. 2012). These data were used to generate a compound, continuous index of LUI, by summing the standardised components (F + M + G), see (Blüthgen et al. 2012) and Supporting information.
Process and diversity data
Data on 14 different ecosystem services (or service indicators) were collected. These included aboveground productivity (shoot biomass, g m−2), forage quality (index based on crude protein concentration and relative forage value), carbon sequestration (soil organic carbon concentration, g kg−1 soil), belowground productivity (root biomass, g m−2), root decomposition (mass loss from root litter bags after 6 months; note that data were not available for leaf litter decomposition), nitrification rate (potential nitrification based on enzyme activity), phosphorus (P) retention index ((shoot P stock + microbial P stock)/(shoot P stock + microbial P stock + soil extractable P [NaHCO3])), mycorrhisation (AMF hyphal length, g−1 soil), soil aggregation (proportion water stable soil aggregates), pest control (number of trap nesting wasps predating pest insects), pathogen regulation (log(1/total cover of foliar fungal pathogens)), pollination (number of flower visitors), aesthetic appeal to people using the grassland for recreation (total flower cover % was chosen as the public prefer flower rich landscapes (Junge et al. 2015; Lindemann‐Matthies et al. 2010)) and cultural/conservation value (species richness of birds), see Table S2 for more details. The ecosystem service measures were all collected on the same plots but sampling covered different areas: all were collected at scales appropriate for the particular measure (see Table S1).
Vegetation data were collected annually between mid‐May and mid‐June in a 4 × 4 m area in the centre of each plot, where the percentage cover of all vascular plant species was recorded. As our biodiversity measure we used mean plant species richness between 2009 and 2012. We chose it because it is the most widely used diversity measure, making our results comparable with those of the many experiments which manipulated species richness (e.g. Isbell et al. 2011). Functional or phylogenetic diversity have been suggested as superior predictors of ecosystem functioning (Flynn et al. 2011). However, these metrics have not been used as widely and very few experiments have explicitly manipulated them (Cadotte 2013).
We used four measures to characterise the resource strategies of the plant communities. Three traits were used: high specific leaf area (SLA) and leaf nitrogen contents (leaf N) and low leaf dry matter contents (LDMC) (Reich 2014) indicate an acquisitive strategy. We calculated community weighted mean (CWM) values by weighting the trait value of each species by its abundance (relative cover). CWMs were calculated for each year between 2009 and 2012 and then averaged across years to produce a single value per plot. The fourth metric was an integrated measure based on the three traits (RES; see Fig. S1). Traits were obtained from the LEDA database (Knevel et al. 2003) but were not available for some species, so these were excluded from CWM estimation. However, at least 88% (mean of 99%) of the total cover (averaged across years) was represented by an SLA value. For the other traits these values were lower (minimum of 74% (mean of 98%) for LDMC and 61% (mean of 88%) for Leaf N).
Calculation of multifunctionality
We used a threshold‐based approach to calculate multifunctionality. This is conceptually superior to averaging functions because it does not assume that they are substitutable (Byrnes et al. 2013). In each plot, we scored each service depending on whether it passed a threshold, defined as a certain fraction of the maximum level across all plots. This assumes that land managers can accept a certain loss of service delivery but below a threshold the loss is unacceptable and functioning is impaired. The maximum was a mean of the highest five values, thus reducing the influence of outliers (Zavaleta et al. 2010). We focus on defining the maximum across all plots because we are interested in measuring the functionality of the grasslands relative to the maximum across Germany. Defining separate thresholds per region could be misleading as ecosystem services at low levels in a given region would still seem high if the regional maximum was low. However, using regional thresholds did not change our conclusions, Fig. S2. Here we present results based on a 50% threshold but we also explored other thresholds, see Fig. S3. As the range of values for some services did not include 0, we first rescaled all services to a minimum of 0, to prevent some exceeding the threshold in almost all plots. This scaling did not affect our conclusions because responses were similar when multifunctionality was calculated as a mean of z‐scores (Fig. S2). Multifunctionality was calculated as the proportion of processes measured that exceeded the threshold. Processes not measured on a plot were therefore excluded, however, this did not affect multifunctionality values, see Fig. S4. Multifunctionality was calculated using the function ‘multidiv’, available from github.com/eric‐allan/multidiversity.
To calculate multifunctionality according to a range of land‐use objective scenarios we weighted the services according to the values shown in Fig. 1. The five objectives form a gradient from scenario 1 prioritising agricultural production, via scenarios 2–4 that include an increasing number of supporting and regulating services, to scenario 5 in which cultural services are favoured. We used weightings of 50% or 100%: two ecosystem services with a weighting of 50% passing the threshold are equivalent to a single service with a weighting of 100%. To calculate multifunctionality for scenario 1, we scaled biomass and forage quality between 0 and 1 (the maximum was again a mean of the top five values) and averaged the scaled values because using a threshold with only two values would produce misleading results. Calculating multifunctionality as a mean of scaled values for the other scenarios did not affect our conclusions (Fig. S2). For more on the multifunctionality scenarios see Supporting information (extended methods).
Statistical analysis
All statistical analysis was done using the R statistical programme (R Core Team 2012). In the first step of our analysis we analysed the relationships between LUI and multifunctionality, using separate linear models for each multifunctionality measure and region. Relationships between LUI, species richness and functional composition were assessed by calculating Pearson correlation coefficients. Regions were analysed separately because they differ substantially in biodiversity and environmental conditions.
In the second step, and to estimate direct and biodiversity‐mediated indirect effects of LUI on multifunctionality, we used structural equation modelling (SEM). We fitted the SEM shown in Fig. 3a; in addition to quantifying direct and indirect effects of LUI, our SEM accounts for effects of two environmental covariates, soil pH and soil depth, that may drive plant species richness or functional composition (e.g. Grime et al. 1988; Dornbush & Wilsey 2010) and may also directly affect services, particularly biogeochemical cycling measures. These soil parameters could be correlated with each other and with LUI, so covariances between them were estimated (Fig. 3a). We fitted separate SEMs for each region and multifunctionality scenario, using the lavaan package (Rosseel 2012). We estimated direct and indirect effects as standardised path coefficients (SPC), thus allowing for comparisons between regions and multifunctionality scenarios. We calculated the fit of each SEM to the data using a Chi‐squared test (Table S3).
Figure 3.
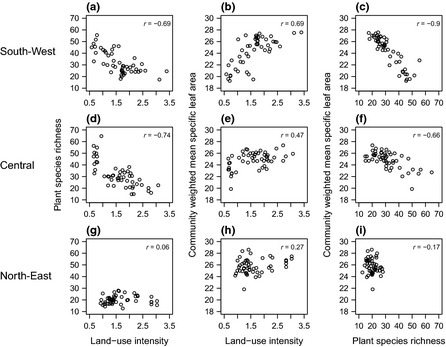
The relationships between land‐use intensity (LUI), plant species richness and community‐weighted mean specific leaf area (SLA). (a, d, g) LUI and plant species richness (b, e, h) LUI and community‐weighted mean of SLA and (c, f, i) species richness and community weighted mean (CWM) SLA. The relationship is shown for the three regions (a–c) south‐west (d–f) central and (g–i) north‐east. Pearson correlation coefficients, r, are shown for each relationship.
In the third step of our analysis we determined whether changes in functional composition provided a better explanation of indirect effects than changes in biodiversity did. We compared the fit of SEMs containing species richness (Fig. 3a) with SEMs containing CWM SLA (Fig. 3e), CWM leaf N, CWM LDMC or CWM RES, using Akaike's Information Criterion (AIC) and selected the model with the lowest AIC value (Table S3). Only models containing species richness or CWM SLA were ever selected as the best (Table S3), so we do not discuss other functional composition measures further.
Finally, we performed several tests that assessed the sensitivity of our results to the method used to calculate multifunctionality, see Supporting information.
Results
Overall effects of land‐use intensity
Effects of LUI on ecosystem multifunctionality depended strongly on the multifunctionality scenario. When the measure gave high weights to provisioning services (production and fodder quality, scenario 1), high LUI was associated with high multifunctionality (mean slope for LUI across regions = 0.13, Fig. 2a, f, k). However, this positive association weakened and became neutral as more ecosystem services were considered (mean slope 0.03, 0.02 and −0.007 for scenarios 2–4, respectively, Fig. 2b–d, g–i, l–n) and eventually became negative where the objective was to maximise cultural services (mean slope scenario 5 = −0.04, Fig. 2e, j, o). These patterns were consistent across regions but were only significant in the central and south‐west. In the north‐east effects were weaker.
Figure 2.
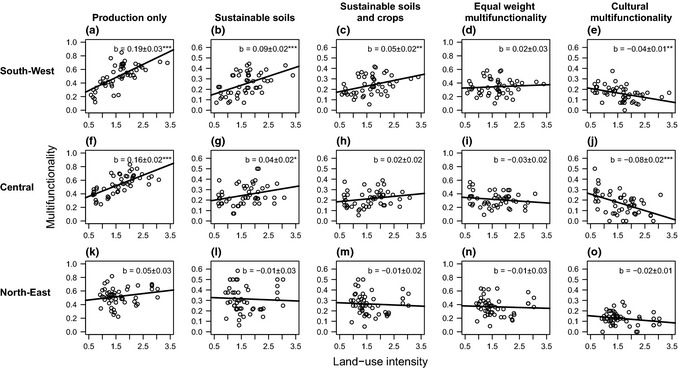
The overall relationship between land‐use intensity (LUI) and ecosystem multifunctionality calculated for five scenarios, representing different land‐use objectives (Fig. 1), and three regions. These graphs show total effects of LUI on multifunctionality, i.e. direct + indirect effects. Lines show fits from a linear regression and slopes ± 1 SE are shown. Asterisks indicate significance levels ***P < 0.001, **P < 0.01, *P < 0.05. Note that y‐axes have been scaled to the maximum multifunctionality value possible for a given scenario, see the extended methods in the Supporting information.
Responses of plant species richness and functional composition to LUI varied substantially between the three regions (Fig. 3). In the fertile grasslands of the north‐east, LUI was not related to plant species richness or CWM SLA (Fig. 3g–i). In the central region, land‐use intensification was related to decreased plant species richness (r = −0.74) and slightly increased CWM SLA (r = 0.47, Fig. 3d–f). In the south‐west land‐use intensification was related to both reduced plant species richness (r = −0.69) and increased CWM SLA (r = 0.69, Fig. 3a–c). Due to this common response, SLA and species richness were most closely correlated in the south‐west (r = −0.90). They were also correlated in the central region (r = −0.66) but were weakly correlated in the north‐east (r = −0.17).
Direct and biodiversity‐mediated indirect effects of land‐use intensity
Our SEM revealed that indirect effects mediated by species richness were often of similar or even greater importance than the direct effects of LUI. The mean absolute SPC for indirect effects via changing species richness (0.189), calculated by averaging values across all five multifunctionality measures and three regions, was very similar to that for direct effects (0.187; Fig. 4b–d and Fig. S6). Indirect effects were highly variable and could be positive or negative depending on the region and multifunctionality scenario. Negative indirect effects indicate biodiversity loss reduces multifunctionality, whereas positive effects indicate that it increases multifunctionality. In the north‐east, indirect effects were universally weak (Fig. 4d and h): mean direct SPC = 0.1 and indirect SPC = 0.02, whereas in the central region they were similar (SPC = 0.21) to direct effects (SPC = 0.28) and in the south‐west they were stronger than direct effects, see below.
Figure 4.
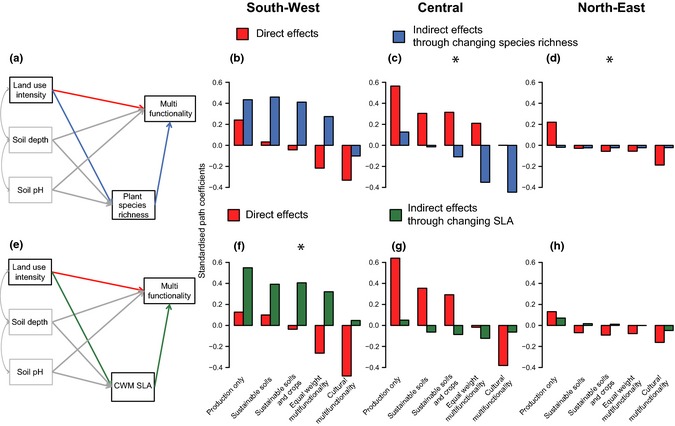
The strength of direct and indirect land‐use effects on ecosystem multifunctionality. The index of multifunctionality was calculated for five scenarios, representing different land‐use objectives (Fig. 1), and three regions. The structural equation model used to calculate the indirect effects (a) through changing species richness and (e) through changing specific leaf area (SLA). Indirect effects could be mediated through changing species richness (b–d) or through changes to the community weighted mean (CWM) of SLA (f–h). The best explanation for the indirect effects (either SLA or species richness) for each region is shown by an asterisk (*) (see, Methods and Table S3). Indirect effects are calculated by multiplying the path coefficient for the effect of land‐use intensity (LUI) on species richness/SLA with the path coefficient for the effect of species richness/SLA on multifunctionality, see Table S4 for the individual path coefficients.
The direction of indirect, biodiversity mediated, effects varied substantially between scenarios. In both the central and south‐west regions, indirect effects became increasingly negative as more services were included in the multifunctionality measure (Fig. 4b and c, mean indirect SPC = 0.18 for scenario 1 and −0.03 for scenario 4). In these regions, indirect effects on multifunctionality were most strongly negative when cultural services were the priority (mean indirect SPC for cultural multifunctionality −0.18), indicating that such services are more negatively affected by biodiversity loss. In the central region indirect effects on production orientated multifunctionality were weak. In contrast, in the south‐west they were strongly positive (Fig. 4), indicating that diversity loss boosted production‐focused multifunctionality. In the third step of our analysis, we tested whether this was because the indirect effects were better explained by changes in SLA, which is negatively correlated with species richness, see below.
Indirect effects of functional composition
Comparing models where indirect effects were mediated by either plant species richness or by SLA revealed that the nature of the indirect effects varied between the two regions where they were detected (central and south‐west). In the central region indirect effects were best explained by changes in plant species richness (Table S3) and were strongly negative for multifunctionality measures including many ecosystem functions and cultural services (see Methods above, Fig. 4c). In contrast, in the south‐west region indirect effects were best explained by changes in SLA (Table S3; Fig. 4 and Fig. S7a–c). Here, indirect effects, through changing SLA, were similar to those through species richness in that they were positive and stronger than the direct effects of LUI: the average SPC across all scenarios in the south‐west was 0.34 for indirect effects and 0.20 for direct effects. Positive indirect effects arise because land‐use intensification increases SLA (shift towards fast‐growing species) and this increase in SLA increases multifunctionality. SLA‐mediated indirect effects in the south‐west were particularly pronounced for production‐orientated scenarios: they were 1.8, 14.2 and 9.7 times greater than direct effects for scenarios 1–3 respectively (Fig. 4f).
Varying the threshold
Increasing the threshold above 50% of the maximum increased the strength of negative indirect effects in the central region: mean SPC for equal weight multifunctionality changed from 0.02 to −0.28 when the threshold increased from 50 to 80%, see Fig. S3. Indirect effects via species richness are therefore strongest when high levels of ecosystem services are desired. In contrast, increasing the threshold led to weaker effects of SLA in the south‐west and for scenarios 4 and 5, indirect effects via SLA were slightly negative at an 80% threshold, indicating some ecosystem functions respond negatively to increasing SLA.
Discussion
Land‐use intensity had strong overall effects on ecosystem multifunctionality. As expected, the strength and direction of these effects depended on the management scenario and the corresponding package of services which should be maintained. Effects of land‐use intensification were positive when the scenario focussed on agricultural production and were negative when it focussed on cultural ecosystem services. These results provide further evidence of a strong trade‐off between the delivery of provisioning and cultural services in agricultural systems (Lavorel et al. 2010; Raudsepp‐Hearne et al. 2010; Martin et al. 2014), which has been central to debates about the environmental impacts of intensification and their management (Foley et al. 2005).
Decomposing the overall effect of LUI into direct and indirect effects, revealed that changes to the plant community, including biodiversity loss and associated changes to functional composition, were a key mechanism by which land‐use intensification indirectly affected ecosystem multifunctionality. Examining first the indirect effects of biodiversity loss, we found that, across all multifunctionality scenarios and regions, indirect effects were on average as strong as direct effects. These indirect effects were therefore considerably larger than those observed in the two previous attempts to separate direct and indirect effects of global change on ecosystem functioning, both of which were experimental studies of effects of nitrogen enrichment on productivity or biogeochemical cycling (Manning et al. 2006; Isbell et al. 2013). This is especially remarkable, as our estimates of the strength of indirect effects were quite conservative. We assumed that a service was still provided when it declined by 50% relative to its maximum; however, land managers may often find a 50% loss of services unacceptable. Using thresholds > 50% revealed stronger negative effects of biodiversity loss. Biodiversity loss may therefore be particularly damaging where high levels of many ecosystem services are desired (Byrnes et al. 2013). In the south‐west region biodiversity loss was associated with increased levels of provisioning services, which further analysis revealed was because biodiversity loss is accompanied by an increased abundance of fast‐growing species and this shift in functional composition boosts production, see below. Overall, our results indicate that biodiversity loss can have important functional consequences in the real world, and not just in experimental ecosystems (Hector & Bagchi 2007; Isbell et al. 2011) and it is an important mechanism by which land‐use change affects ecosystem service delivery.
While we observed large biodiversity‐mediated effects on multifunctionality, their direction and strength varied considerably depending on the management objective. In particular, indirect effects became increasingly important as more services were included in the multifunctionality measure, which agrees with experimental results showing increasingly positive effects of biodiversity on multifunctionality as more functions are considered (Isbell et al. 2011). We also found that biodiversity loss had negative effects on cultural services and weak or positive effects on provisioning services. The trade‐off between provisioning and cultural services, found in many other studies (e.g. Lavorel et al. 2010; Raudsepp‐Hearne et al. 2010; Martin et al. 2014) may therefore be mediated by biodiversity loss: land‐use intensification increases forage production but also causes large loss of biodiversity, which reduces the delivery of cultural services. These results emphasise the importance of management scenarios for understanding effects on multifunctionality. The scenarios considered here are a first attempt to move away from the equal weighting of functions in multifunctionality measures. Future work could involve collaboration with social scientists to derive weightings from surveys and produce measures of multifunctionality specifically tailored to the needs of different stakeholders, along with overall multifunctionality measures that reflect consensus between them.
The overall effects of land‐use intensification on multifunctionality were similar across the different regions. The best explanation of the underlying mechanisms, however, varied substantially. This agrees with our hypothesis that the strength and nature of indirect effects should vary depending on land‐use impacts on plant communities. Where land‐use intensification does not reduce biodiversity or change functional composition, only direct effects can be expected. This was true in the most fertile region, in the north‐east (Fig. 4, also see Socher et al. 2012). In the other two regions, indirect effects were important but appeared to be mediated by different processes. In the central region indirect effects were best explained by biodiversity loss. In general, where exploitative species already dominate diverse communities, e.g. in productive mesotrophic grasslands (Ellenberg 1988), land‐use intensification may reduce biodiversity but have little impact on functional composition. In contrast, in the region with the least favourable growing conditions, the south‐west, extensively managed grasslands were diverse and dominated by slow growing species. Land‐use intensification was associated with large biodiversity losses and a strong shift to fast‐growing species. In line with our hypothesis, here the shifts in functional composition indirectly increased more production focussed measures of multifunctionality and were more important than biodiversity loss. Perhaps surprisingly, changes in functional composition had a larger effect on provisioning services than the direct effects of land‐use intensification. In general, fertilisation in intensively managed grasslands will cause a shift towards plants with an exploitative strategy, and tissues that turn‐over and decompose rapidly. This accelerates nutrient cycling rates (Lavorel et al. 2010; de Vries et al. 2012) and appears to play a greater role in increasing agricultural productivity than the direct effect of fertilisation. Overall, these strong regional differences emphasise the need to consider the context dependency of biodiversity and functional composition effects.
Our results have implications for mapping and managing ecosystem services at large spatial scales. They suggest that using simple measures of species richness and functional diversity as proxies of ecosystem function may be misleading (e.g. Maskell et al. 2013). Instead, we need to consider the dependence of relationships between biodiversity (or functional composition) and ecosystem services on environmental conditions. Similarly, according to our findings, the management of biodiversity and ecosystem services needs to be tailored to local environmental conditions. A strong trade‐off between biodiversity and provisioning services, as in our south‐west region, might call for a ‘land‐sparing’ strategy (Green et al. 2005), where diverse, low‐productivity grasslands provide cultural services, while separate, more intensely used grasslands are dedicated to forage production. In contrast, a positive relationship between biodiversity and multifunctionality, as in our central region, calls for a ‘land‐sharing’ strategy, where management simultaneously promotes both biodiversity and a range of ecosystem services.
SEM allows causal relationships to be inferred from observed data. However, as our study did not experimentally manipulate LUI we cannot rule out the possibility that other unmeasured variables are responsible for some of its effects. In particular, the diversity and functional composition of other taxa, such as soil microbes (de Vries et al. 2012) and invertebrates (Hillebrand & Matthiessen 2009; Cardinale et al. 2012) can also drive ecosystem functions. In our grasslands plant diversity is positively correlated with the diversity of many arthropod groups and with aboveground multidiversity (overall ecosystem diversity) (Manning et al. 2015). The diversities of belowground ecosystem service providers may also be strongly positively correlated with plant species richness (Scherber et al. 2010), making plant diversity a suitable proxy for their diversity as well. In contrast, where these groups are uncorrelated with plant diversity, their indirect effects on multifunctionality will be absorbed within the direct effects of land use, leading to underestimates of the importance of biodiversity in driving multifunctionality. Future analyses that compare the strength of plant diversity effects with those of the diversities of other ecosystem service providers are needed to test these ideas.
Conclusions
We introduced a new approach to understanding biodiversity–ecosystem function relationships in natural systems and were able to demonstrate that biodiversity loss and changes to functional composition are key mechanisms that underlie the impacts of land‐use intensification on ecosystem‐service delivery in managed grassland ecosystems. Restoration of biodiversity (Benayas et al. 2009) in more intensively managed grasslands might therefore offset some of the negative effects of intensification and promote ecosystems that deliver a wider range of services, especially cultural services. By using novel measures of multifunctionality, in which services were weighted according to management objectives, we were also able to show that the relationships between land use, biodiversity, functional composition and multifunctionality depend greatly on the services required. We therefore recommend the development of improved measures of multifunctionality that are specifically tailored to the needs of stakeholders. In addition, we emphasise that understanding ecosystem‐service provision requires a focus on the interaction between global change drivers and changes to both biodiversity and functional composition, if we are to successfully transfer conclusions from experimental biodiversity research to the real‐world and elucidate the true importance of biodiversity for providing ecosystem services.
Authorship
EA, PM and MF designed the analysis; EA analysed data; EA, PM and MF wrote the paper, all other authors provided data and substantially revised the paper.
Supporting information
Acknowledgements
We thank Robert Bagchi, Santiago Soliveres, Owen Petchey and Teja Tscharntke for comments on the manuscript. We also thank the managers of the three exploratories, Sonja Gockel, Kerstin Wiesner and Martin Gorke for their work in maintaining the plot and project infrastructure; Simone Pfeiffer and Christiane Fischer giving support through the central office, Birgitta König‐Ries and Michael Owonibi for managing the central database, and Eduard Linsenmair, Dominik Hessenmöller, Jens Nieschulze, François Buscot, Ernst‐Detle f Schulze, and the late Elisabeth Kalko for their role in setting up the Biodiversity Exploratories project. This work was funded by the Deutsche Forschungsgemeinschaft Priority Program 1374 ‘Infrastructure‐Biodiversity Exploratories.’ Fieldwork permits were given by the responsible state environmental offices of Baden‐Württemberg, Thüringen and Brandenburg (according to §72 Brandenburgisches Naturschutzgesetz).
Ecology Letters (2015) 18: 834–843
References
- Allan, E. , Bossdorf, O. , Dormann, C.F. , Prati, D. , Gossner, M.M. , Tscharntke, T. et al (2014). Interannual variation in land‐use intensity enhances grassland multidiversity. Proc. Natl. Acad. Sci. USA, 111, 308–313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Balvanera, P. , Siddique, I. , Dee, L. , Paquette, A. , Isbell, F. , Gonzalez, A. et al (2014). Linking biodiversity and ecosystem services: current uncertainties and the necessary next steps. Bioscience, 64, 49–57. [Google Scholar]
- Benayas, J.M.R. , Newton, A.C. , Diaz, A. & Bullock, J.M. (2009). Enhancement of biodiversity and ecosystem services by ecological restoration: a meta‐analysis. Science, 325, 1121–1124. [DOI] [PubMed] [Google Scholar]
- Blüthgen, N. , Dormann, C.F. , Prati, D. , Klaus, V.H. , Kleinebecker, T. , Hölzel, N. et al (2012). A quantitative index of land‐use intensity in grasslands: integrating mowing, grazing and fertilization. Basic Appl. Ecol., 13, 207–220. [Google Scholar]
- Bradford, M.A. , Wood, S.A. , Bardgett, R.D. , Black, Helaina I.J. , Bonkowski, M. , Eggers, T. et al (2014). Discontinuity in the responses of ecosystem processes and multifunctionality to altered soil community composition. Proc. Natl. Acad. Sci. USA, 111, 14478–14483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Byrnes, J.E.K. , Gamfeldt, L. , Isbell, F. , Lefcheck, J.S. , Griffin, J.N. , Hector, A. et al (2013). Investigating the relationship between biodiversity and ecosystem multifunctionality: challenges and solutions. Methods Ecol. Evol., 5, 111–124. [Google Scholar]
- Cadotte, M.W. (2013). Experimental evidence that evolutionarily diverse assemblages result in higher productivity. Proc. Natl. Acad. Sci. USA, 110, 8996–9000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cardinale, B.J. , Duffy, J.E. , Gonzalez, A. , Hooper, D.U. , Perrings, C. , Venail, P. et al (2012). Biodiversity loss and its impact on humanity. Nature, 486, 59–67. [DOI] [PubMed] [Google Scholar]
- Cousins, S.A.O. & Eriksson, O. (2002). The influence of management history and habitat on plant species richness in a rural hemiboreal landscape, Sweden. Landscape Ecol., 17, 517–529. [Google Scholar]
- Creed, R.P. , Cherry, R.P. , Pflaum, J.R. & Wood, C.J. (2009). Dominant species can produce a negative relationship between species diversity and ecosystem function. Oikos, 118, 723–732. [Google Scholar]
- Dornbush, M.E. & Wilsey, B.J. (2010). Experimental manipulation of soil depth alters species richness and co‐occurrence in restored tallgrass prairie. J. Ecol., 98, 117–125. [Google Scholar]
- Ellenberg, H. (1988). Vegetation Ecology of Central Europe. Cambridge University Press, Cambridge. [Google Scholar]
- Fischer, M. , Bossdorf, O. , Gockel, S. , Hänsel, F. , Hemp, A. , Hessenmöller, D. et al (2010). Implementing large‐scale and long‐term functional biodiversity research: the Biodiversity Exploratories. Basic Appl. Ecol., 11, 473–485. [Google Scholar]
- Flynn, D.F.B. , Mirotchnick, N. , Jain, M. , Palmer, M.I. & Naeem, S. (2011). Functional and phylogenetic diversity as predictors of biodiversity–ecosystem‐function relationships. Ecology, 92, 1573–1581. [DOI] [PubMed] [Google Scholar]
- Foley, J.A. , DeFries, R. , Asner, G.P. , Barford, C. , Bonan, G. , Carpenter, S.R. et al (2005). Global consequences of land use. Science, 309, 570–574. [DOI] [PubMed] [Google Scholar]
- Gamfeldt, L. , Snall, T. , Bagchi, R. , Jonsson, M. , Gustafsson, L. , Kjellander, P. et al (2013). Higher levels of multiple ecosystem services are found in forests with more tree species. Nat. Commun., 4, 1340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldstein, J.H. , Caldarone, G. , Duarte, T.K. , Ennaanay, D. , Hannahs, N. , Mendoza, G. et al (2012). Integrating ecosystem‐service tradeoffs into land‐use decisions. Proc. Natl. Acad. Sci. USA, 109, 7565–7570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grace, J.B. , Michael Anderson, T. , Smith, M.D. , Seabloom, E. , Andelman, S.J. , Meche, G. et al (2007). Does species diversity limit productivity in natural grassland communities? Ecol. Lett., 10, 680–689. [DOI] [PubMed] [Google Scholar]
- Green, R.E. , Cornell, S.J. , Scharlemann, J.P.W. & Balmford, A. (2005). Farming and the Fate of Wild Nature. Science, 307, 550–555. [DOI] [PubMed] [Google Scholar]
- Grime, J.P. , Hodgson, J.G. & Hunt, R. (1988). Comparative Plant Ecology. A Functional Approach to Common British Species. Unwin Hyman Ltd, London. [Google Scholar]
- Hector, A. & Bagchi, R. (2007). Biodiversity and ecosystem multifunctionality. Nature, 448, 188–190. [DOI] [PubMed] [Google Scholar]
- Hillebrand, H. & Matthiessen, B. (2009). Biodiversity in a complex world: consolidation and progress in functional biodiversity research. Ecol. Lett., 12, 1405–1419. [DOI] [PubMed] [Google Scholar]
- Isbell, F. , Calcagno, V. , Hector, A. , Connolly, J. , Harpole, W.S. , Reich, P.B. et al (2011). High plant diversity is needed to maintain ecosystem services. Nature, 477, 199–202. [DOI] [PubMed] [Google Scholar]
- Isbell, F. , Reich, P.B. , Tilman, G.David , Hobbie, S.E. , Polasky, S. & Binder, S. (2013). Nutrient enrichment, biodiversity loss, and consequent declines in ecosystem productivity. Proc. Natl. Acad. Sci. USA, 110, 11911–11916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Junge, X. , Schüpbach, B. , Walter, T. , Schmid, B. & Lindemann‐Matthies, P. (2015). Aesthetic quality of agricultural landscape elements in different seasonal stages in Switzerland. Landsc. Urban Plan., 133, 67–77. [Google Scholar]
- Knevel, I.C. , Bekker, R.M. , Bakker, J.P. & Kleyer, M. (2003). Life‐history traits of the Northwest European flora: the LEDA database. J. Veg. Sci., 14, 611–614. [Google Scholar]
- Laliberté, E. & Tylianakis, J.M. (2011). Cascading effects of long‐term land‐use changes on plant traits and ecosystem functioning. Ecology, 93, 145–155. [DOI] [PubMed] [Google Scholar]
- Lavorel, S. & Grigulis, K. (2012). How fundamental plant functional trait relationships scale‐up to trade‐offs and synergies in ecosystem services. J. Ecol., 100, 128–140. [Google Scholar]
- Lavorel, S. , Grigulis, K. , Lamarque, P. , Colace, M.‐P. , Garden, D. , Girel, J. et al (2010). Using plant functional traits to understand the landscape distribution of multiple ecosystem services. J. Ecol., 99, 135–147. [Google Scholar]
- Lindemann‐Matthies, P. , Junge, X. & Matthies, D. (2010). The influence of plant diversity on people's perception and aesthetic appreciation of grassland vegetation. Biol. Conserv., 143, 195–202. [Google Scholar]
- Maestre, F.T. , Quero, J.L. , Gotelli, N.J. , Escudero, A. , Ochoa, V. , Delgado‐Baquerizo, M. et al (2012). Plant species richness and ecosystem multifunctionality in global drylands. Science, 335, 214–218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manning, P. , Gossner, M.M. , Bossdorf, O. , Allan, E. , Zhang, Y.‐Y. , Prati, D. et al (2015). Grassland management intensification weakens the associations among the diversities of multiple plant and animal taxa. Ecology, 96, 1492–1501. [Google Scholar]
- Manning, P. , Newington, J.E. , Robson, H.R. , Saunders, M. , Eggers, T. , Bradford, M.A. et al (2006). Decoupling the direct and indirect effects of nitrogen deposition on ecosystem function. Ecol. Lett., 9, 1015–1024. [DOI] [PubMed] [Google Scholar]
- Martin, L.M. , Polley, H.W. , Daneshgar, P. , Harris, M. & Wilsey, B. (2014). Biodiversity, photosynthetic mode, and ecosystem services differ between native and novel ecosystems. Oecologia, 175, 687–697. [DOI] [PubMed] [Google Scholar]
- Maskell, L.C. , Crowe, A. , Dunbar, M.J. , Emmett, B. , Henrys, P. , Keith, A.M. et al (2013). Exploring the ecological constraints to multiple ecosystem service delivery and biodiversity. J. Appl. Ecol., 50, 561–571. [Google Scholar]
- Millennium Ecosystem Assessment (2005). Ecosystems and Human Well‐being. Island Press Washington, DC. [Google Scholar]
- R Core Team (2012). R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. [Google Scholar]
- Raudsepp‐Hearne, C. , Peterson, G.D. & Bennett, E.M. (2010). Ecosystem service bundles for analyzing tradeoffs in diverse landscapes. Proc. Natl. Acad. Sci. USA, 107, 5242–5247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reed, M.S. , Graves, A. , Dandy, N. , Posthumus, H. , Hubacek, K. , Morris, J. et al (2009). Who's in and why? A typology of stakeholder analysis methods for natural resource management. J. Environ. Manage., 90, 1933–1949. [DOI] [PubMed] [Google Scholar]
- Reich, P.B. (2014). The world‐wide ‘fast–slow’ plant economics spectrum: a traits manifesto. J. Ecol., 102, 275–301. [Google Scholar]
- Rosseel, Y. (2012). lavaan: an R package for structural equation modeling. J. Stat., 48, 1–36. [Google Scholar]
- Rudmann‐Maurer, K. , Weyand, A. , Fischer, M. & Stocklin, J. (2008). The role of landuse and natural determinants for grassland vegetation composition in the Swiss Alps. Basic Appl. Ecol., 9, 494–503. [Google Scholar]
- Scherber, C. , Eisenhauer, N. , Weisser, W.W. , Schmid, B. , Voigt, W. , Fischer, M. et al (2010). Bottom‐up effects of plant diversity on multitrophic interactions in a biodiversity experiment. Nature, 468, 553–556. [DOI] [PubMed] [Google Scholar]
- Socher, S.A. , Prati, D. , Boch, S. , Müller, J. , Klaus, V.H. , Hölzel, N. et al (2012). Direct and productivity‐mediated indirect effects of fertilization, mowing and grazing on grassland species richness. J. Ecol., 100, 1391–1399. [Google Scholar]
- Thébault, A. , Mariotte, P. , Lortie, C.J. & MacDougall, A.S. (2014). Land management trumps the effects of climate change and elevated CO2 on grassland functioning. J. Ecol., 102, 896–904. [Google Scholar]
- de Vries, F.T. , Manning, P. , Tallowin, J.R.B. , Mortimer, S.R. , Pilgrim, E.S. , Harrison, K.A. et al (2012). Abiotic drivers and plant traits explain landscape‐scale patterns in soil microbial communities. Ecol. Lett., 15, 1230–1239. [DOI] [PubMed] [Google Scholar]
- Zavaleta, E.S. , Pasari, J.R. , Hulvey, K.B. & Tilman, G.David (2010). Sustaining multiple ecosystem functions in grassland communities requires higher biodiversity. Proc. Natl. Acad. Sci. USA, 107, 1443–1446. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials