

Abstract
We demonstrate here that functional NMDAR1 receptors and NMDAR2 receptors are expressed by Mcf-7 and SKBR3 breast cancer cell lines, and possibly by most or all high-grade breast tumors, and that these receptors are important for the growth of human breast cancer xenografts in mice. RT-PCR demonstrated mRNA for both NMDAR1 and NMDAR2 receptors are expressed in both Mcf-7 and SKBR3 cell lines, and these messages likely have sequences identical to those reported for human mRNAs. Proteins of the expected respective sizes 120 and 170 KD are generated from these mRNAs by the tumor cells. Cell growth was found to be significantly (p<0.0001) impaired down to 10% of normal growth by the irreversible NMDAR1 antagonists MK-801 and memantine with IC 50s ranging from 600–>800 μM and from 200–300 μM for the two lines. Paradoxically, memantine with a lower binding affinity had the greater influence of the two inhibitors on cell viability. Immunohistochemical examination of high-grade invasive ductal and lobular breast cancer with our polyclonal antibodies against a peptide (-Met-Ser-Ile-Tyr-Ser-Asp-Lys-Ser-Ile-His-) in the extracellular domain of the NMDAR1 receptor gave specific positive staining for the receptor in all 10 cases examined. Positive staining was chiefly concentrated at the membranes of these tumor tissues. No staining with these antibodies was found for normal breast and kidney tissues. When Mcf-7 cells were grown as tumor xenografts in nu/nu mice, the growth of these tumors was completely arrested by daily treatments with MK-801 over five days. All of these data point to active NMDAR receptors being expressed by most breast cancers, and having an important influence on their survival.
Introduction
Functional NMDA receptors were found by us to be expressed by human neuroblastoma cells (LA-N2) [1], and then by small-cell lung cancer (SCLC) cells and SCLC tumors (presented elsewhere). NMDA receptors types 1 and 2 are activated by glycine and glutamate, and together, as heterodimers, constitute an important calcium channel for neurons. The proper functioning of these receptors gives rise to stimulatory impulses between neurons, and seems to be responsible for long-term potentiation, an essential ingredient of short-term memory consolidation in hippocampal and neo-cortical regions of the brain [2]. For some non-neuronal tissues, NMDA receptors are also known to lead to proliferation by activating growth promoting cascades, such as the MAP kinase and ERK pathways [3]. Alternatively, overstimulation of NMDA receptors can lead to cell death through influx of excessive calcium and apoptosis, a destructive process that has been proposed as a factor involved in a number of neurodegenerative diseases, such as Alzheimer’s disease [1, 4]. Recently, the NMDAR1 receptor antagonist memantine, which appears to selectively target overstimulated open NMDA channels, has been successfully introduced for therapy in the treatment of Alzheimer’s disease [5]. Memantine significantly slows loss in cognition in individuals who suffer from this disease, with few side-effects [6, 7]. Because it is well tolerated, this drug is now being considered for the treatment of other neurodegenerative diseases, such as Parkinson’s disease, because of the improvements of motor behavior it produces in MPTP-treated mice, a laboratory model for this disease [8]. The current study shows that both NMDAR1 and NMDAR2B receptors are expressed in breast cancer cells and tumors. Impairment of the function of these NMDA receptors on cells in vitro with NMDAR1 antagonists prevents proliferation and significantly decreases cell number, while treatment of human breast cancer xenografts in mice with an NMDAR1 antagonist stops growth of these tumors.
Materials and Methods
Cell culture and human tissue
Human breast adenocarcinoma cell lines MCF-7, and SKBR-3 were obtained from AmericanType Tissue Culture Collection (ATCC, Rockland, MD). Cells were grown and maintained in DMEM-F12 medium (Mediatech, Inc., Herndon, VA) supplemented with 10% fetal bovine serum (Sigma Chemical Co., St. Louis, MO) at cell densities of 1×105 to 5×105/ml in a humidified atmosphere of 5% CO2 at 37°C. Human breast tissue samples were obtained through the Cooperative Human Tissue Network (University of Alabama at Birmingham), and sections of formaldehyde-fixed breast cancers were from an archival tissue bank of the department of Pathology at Dartmouth Medical School. Sections were prescreened for cancer sub-type.
RT-PCR
The expression of NMDAR1 and NMDAR2B transcripts in MCF-7 and SKBR3 breast cancer cell lines were evaluated by reverse transcription-polymerase chain reaction (RT-PCR). Total RNA was first extracted from cells using TRIzol reagent (Invitrogen, Carlsbad CA) following manufacturer recommendations. The synthesis of the cDNA was performed utilizing an oligo(dT) primer and reverse transcriptase (SuperScriptIII Invitrogen, Carlsbad CA). The cDNA synthesis was performed following the recommendations of the manufacturer. The RNA was first at denatured at 65° C for 5 min, then put on ice for 3 min. Then the reaction mixture was incubated at 50° C for 45 min, and 70° C 15 min. The PCR with the Platinum Taq DNA Polymerase (Invitrogen, Carlsbad CA) was performed in a Mastercycler gradient thermocycler (Eppendorf, Westbury NY). The reaction mixture was subjected to an initial denaturation at 94° C for 3 min, followed by 30 cycles comprised of a denaturation step at 94°C for 45 sec, a step for annealing the primers to the template at 56°C for 45 sec, and an extension step at 72°C for 1.0 min. At the completion of the cycling reaction, an additional extension step at 72°C for 5 min was performed. NMDAR1 primers used in PCR analysis comprised forward primer 1, 5′-ATC TAC TCG GAC AAG AGC ATC C-3′, corresponding to nucleotide 369 to 417 (Genbank L05666); reverse primer 1, 5′-AGC TCT TTC GCC TCC ATC AG-3′, corresponding to compliment nucleotide 658 to 639; forward primer 2, 5′-AAG TAT GCG GAT GGG GTG ACT-3′ (1002 to 1022), reverse primer 2, 5′-CAA AAG CCG TAG CAA CAC TGA -3′ (1393 to 1373). NMDAR2B primers included forward primer 1, 5′-TCA AGG ATG CCC ACG AGA AAG -3′, corresponding to nucleotide residues 358 to 378 (Genbank NM-000834); reverse primer 1, 5′-GTG GCT TCT TCC TTG TGA CAG –3 (922 to 902); forward primer 2, 5′-CCA AAG AGC ATC ATC ACC C -3′ (441 to 459); reverse primer 2, 5′-TGT AGC CAT AGC CAG TCA G -3′ (972 to 954). These primers were obtained from Integrated DNA Technologies, Inc(Coralville, IA). The PCR products were purified by washing once with an equal volume of chloroform and examined on 2% agarose gels. To validate that the products of PCR did not result from initial DNA contamination, DNase-free RNase and RNase-free DNase (Gibco/BRL) was used in some reactions.
Cloning and Sequencing
Direct sequencing was performed on most RT-PCR products without cloning. However, in some cases fresh PCR product (1 μl, 4–12 ng) was cloned into a pCR™ vector and transformed in One Shot™ Competent Cells using a TA Cloning Kit (Invitrogen, San Diego, CA). Cloned cDNAs were prepared by employing a Wizard Mini-preps DNA purification system (Promega), and positive clones were screened with EcoR1 restriction endonuclease (Gibco/BRL) digestion and evaluation on 2% agarose gels. At least two positive clones of each PCR product were chosen for double-strand cDNA sequencing with a Taq DyeDeoxy™ Terminator Cycle Sequencing kit (Applied Biosystems, Foster City, CA). The primers designed for PCR amplifications, as described above, and universal primers (M13 Forward, M13 Reverse and T7) were engaged as sequencing primers. The protocol for DNA sequencing was modified as follows:97°C for 2 min; 25 cycles at 95°C and 30 sec; 58°C for 1.5 min; and 72°C for 1.5 min, with a 72°C extension for 10 min. The products were purified (x2) by phenol/chloroform extraction and precipitation with 100% ethanol. Sequencing was performed using a Model 373 DNA Sequencer (Applied Biosystems, Instrument 865) and sequence analysis performed using the BLAST network service.
Western Blot Analysis
Cell lysates were prepared by sonication using a RIPA Buffer solution (1%NP-40, 1%sodium deoxycholate, 0.1%SDS, 150mM NaCl, 25mM Tris/HCl, PH 7.4) with protease inhibitor (Roche, Indianapolis). Extract were centrifuged at 12,000×g, and the supernatant retained. The protein content of these extracts was assessed by differential absorbance measurements at 215 nm and 225 nm. Aliquots were reduced with 2-mercaptoethanol, heated in a boiling water bath for 5 min and samples of approximately 100 μg of protein were separated on 12% gels by SDS-PAGE using Tris/Glycine/SDS buffer [25 mM Tris, 192 mM glycine, 0.1% SDS (pH 8.3)] at a voltage of 50 V for 10 minutes, then at 75 V for approx 2 h at ambient temperature or at 100V for 1 h at 4°C. The proteins were transferred onto Immobilon-P polyvinylidene difluoride membrane (Millipore, Bedford, MA) in Tris/glycine/SDS buffer with 20% methanol, using the MiniProtean 3 system (Bio-Rad, Hercules, CA). The membrane was blocked by drying using a Model 583 Gel Dryer (Bio-Rad) and by incubation with 5% bovine serum albumen or 5% nonfat milk/0.1% Tween 20, followed by incubation with primary antibodies. PANN1 is a rabbit polyclonal antiserum produced in this laboratory and directed against an N-terminal segment of NMDAR1 (see below). Antibodies from this antiserum were first isolated by affinity chromatography. Commercially available NMDAR1 antiserum was obtained from Cell Signaling, and NMDAR2 antiserum was purchased from Santa Cruz (NMDAe2, Santa Cruz Biochemicals, Santa Cruz, CA). The latter antiserum recognizes all NMDAR2 subunits. The blots were visualized using a chemiluminescence method that employed a horseradish peroxidase-labeled goat antirabbit antibody (Pierce or Cell Signaling), a Lumi-Light Western Blotting Substrate (Roche, Indianapolis, IL) or SuperSignal West Pico Chemiluminescent Substrate(Pierce), and exposure of X-ray film. They were scanned and evaluated using an AlphaEaseFc Stand Alone Image Machine with PC Software(Alpha Innotech, San Leandro CA). The blots were stripped and incubated with anti-GAPDH (Chemicon), or scanned with anti-β-actin (Sigma), to ensure equal protein loading.
Cell Growth/Viability Assay
MCF-7 or SKBR3 breast cancer cells were treated with a 0.05% trypsin preparation, washed in PBS/glycine and plated onto 96 well plates at a density of 104 cells/well in medium for 24 hours to provide time for rectifying possible damage to membrane proteins from the treatment. The wells were washed and incubated with PBS/glycine containing albumin (1.25 mg/ml) in the presence or absence of differing concentrations (25 μM-800 μM) of the NMDA 1 antagonists, Memantine and Dizocilpine maleate (MK-801), or vehicle, and Alamar Blue (1:10 dilution following manufacturer’s recommendation). Fluorescence readings were taken at periods representing 24, 48, and 72 h of incubation with antagonist using a Synergy HT Multi-Detection Microplate Reader and excitation/emission wavelengths of 530 nm and 590 nm. Cell viability in the presence of different concentrations of antagonist was evaluated as % fluorescence of vehicle control at the corresponding incubation time.
Immunohistochemistry of Human Tumor Tissue
Immunohistochemical (IHC) staining for human NMDAR1 was performed on formalin-fixed preparations from ten high-grade invasive breast tumors representing primary, metastatic, and recurrent disease. This IHC followed manufacturer’s recommendations on antigen retrieval and use of purchased reagents. Breast cancer tissues were obtained from an archival tissue library of breast cancer at the Department of Pathology, Dartmouth-Hitchcock Medical Center. Tissue sections (4μm) of formalin-treated material were first treated by heating with citrate buffer to recover antigens, blocked by incubating them with 10% horse serum, and then incubated for two hours at room temperature with either affinity-purified PANN1 antibody preparation in PBS/5% bovine serum albumen alone, PANN1 in PBS/5% BSA containing an excess of peptide antigen (antigen-block negative control), or PBS/5% BSA (no primary antibody negative control). PANN1 represents rabbit polyclonal anti-NMDAR1 antibodies and were generated by us against an N-terminal extracellular peptide domain of the human receptor, in the form of the peptide amide coupled through glutaraldehyde to bovine thyroglobulin as antigen, and use of Fruend’s complete and incomplete adjuvant [9, 10, 11]. This peptide domain is represented in that portion of the mRNA message sequenced by RT-PCR (see above). Following washings (x3) in PBS/BSA, tissues were incubated for 1 h at ambient temperature with biotinylated horse anti-rabbit IgG, washed (x3), then incubated for 1 h at ambient temperature with Streptavidin coupled to peroxidase. Tissues were again washed (x3) and finally reacted with a diaminobenzidine/peroxide mixture and contrasted with hemotoxylin.
Treatment of Breast Tumor Xenografts in Mice
Human tumor xenografts of the breast cancer cell line MCF-7 were raised in estrogen-primed nu/nu mice, and the influence of the NMDAR1 receptor antagonist Dizocilpine maleate (MK-801) examined on the growth of these xenografts, once established, measured by multiplying depth, width, and length determined using a micrometer. Tumor growth in a control group of animals receiving i.p. PBS vehicle (n=4) was compared to tumor growth in animals (n=4) receiving Dizocilpine maleate (MK-801) over 10 days. Animals received two daily doses of 0.3 mg/Kg body weight of the NMDAR1 antagonist (MK-801). Results were summarized to determine the tumor growth rate for each mouse by the following formula: (FS-IS)/D where FS was the final tumor size (at the end of the study), IS the initial tumor size at the start of treatment, and D the length of the follow-up period in days. Results were found for each treatment group (mean, standard deviation, for the tumor growth rate). In addition, we modeled the longitudinal growth data using repeated measures ANOVA.
Results
Expression of NMDA Receptors by Cultured Breast Cancer Cells and Tumor Tissues
RT-PCR of poly(A+)RNA preparations from both MCF-7 and SKBR3 breast cancer cell lines using selected forward and reverse primers, gave, in each case, a single product of size predicted from the structure of cDNA for human NMDAR1 from brain tissue, and reported earlier by us for human LA-N-2 neuroblastoma cells (13) and human small cell lung cancer (reported elsewhere). Cloning and nucleotide sequence analysis of these NMDA glutamate receptor RT-PCR overlapping products (488 bp and 263 bp), coding for portions of the extracellular domain, showed them to have exact sequence homology with position 208–695 of the brain receptor, and sequence identity in this portion of the NMDAR1 receptor from human neuroblastoma and small cell lung cancer cell lines. The region in the mRNA examined represents approximately 30% of the open reading frame for the extracellular N-terminal domain for this receptor sub-unit. As was found for the mRNA from LA-N-2 cells (and SCLC cells), there was no evidence for alternate splicing of the message as has been reported by Moriyoshi et al.[12] for NMDAR1 from rat brain. RT-PCR of poly(A+) RNA was unaffected by prior treatment of preparations with DNase, and no products were generated when initial treatment with RNase was performed or when reverse transcriptase was omitted from reaction mixtures. Nucleotide sequencing also revealed normal sequences for the two other regions of NMDAR1 mRNA amplified giving predicted RT-PCR products of 300 and 391 base pairs. In addition, RT-PCR of NMDAR2B message provided predicted products of 471 and 564 base pairs that upon nucleotide sequencing were shown to have sequences identical to those reported for human brain tissue [13].
There is now a consensus that the molecular weight of the NMDAR1A protein subunit is 116 kDaltons, and a major band of approximately this size was displayed by lysates from both cell lines examined in Western Analysis using affinity-purified PANN1 antibodies. A second minor band of approximately 55 kD was also apparent. This second band could be an N-terminal break-down product of the receptor, because PANN1 is directed against an N-terminal moiety of the protein (Figure 1). Alternatively, the NMDAR2 subunits are 165–180 kDaltons, and a major band of approximately 170 kDaltons (in addition to bands corresponding to smaller proteins) was dispayed in Westerns of both breast cancer cell lines by the Santa Cruz NMDAR2e polyclonal antibody, indicating that NMDAR2A, B, or C of normal size is expressed by all of the cell lines (data not shown). Coupled with RT-PCR data, we can infer that the breast cancer cell lines examined both express and translate at least the NMDAR2B form of the NMDAR2 receptor.
Figure 1.

Western Analysis showing the presence of NMDAR1 protein in MCF-7 and SKBR3 cells. NMDAR receptor proteins are present in breast cancer cell lysates. Lysates were examined by SDS-PAGE using 12.5% gels and Western blot analysis using our polyclonal antibody preparation recognizing NMDAR1 proteins. Shown are results obtained for, (A) MCF-7 cells and (B) SKBR3 cells with the positions of molecular mass markers (kDa) indicated on the right of figure. A major form protein of 116 kDa and a minor form of 55 kDa were apparent. Protein loading was the same for each cell line.
Immunohistochemical evaluation of breast cancer tumor sections with anti-NMDAR1 antibodies gave strong and clear positive staining for eight of ten tumors (Figure 2), and weak, questionable staining in the other two cases examined. All of the 6 cases of metastatic and recurrent disease were among those giving a strong (+++) positive reaction. Staining of cancer cells was principally located to the membrane of these cells. All staining of cancer cells with PANN1 antibodies was completely blocked by the presence of peptide antigen against which the antibodies were originally generated. Normal cells in each section did not react with the antibodies. No positive staining of normal breast, lung, and liver was apparent with PANN1 anti-NMDAR1 antibodies for the same conditions of IHC, except for Kufper cells in the liver and isolated macrophages in tissues.
Figure 2.

NMDAR1 protein is present in high grade breast cancer. (A), (B), Positive ABC immunohistochemical staining in two cases of ductal carcinoma using our polyclonal antibodies that recognize an N-terminal peptide fragment of the receptor. Staining was concentrated at the surface of cancer cells and absent from adjacent normal cells. (C) Absense of positive ABC immunohistochemical staining from asection of normal breast (magnification x 20, Olympus/Micropublisher QImager).
NMDA Receptor Antagonists Affect Cultured Breast Cancer Cell Viability
The Alamar Blue assay used in this study is reported to provide a similar evaluation of cell proliferation as the 3H-thymidine assay [14]. Significant effects (P<0.001) on this aspect of cell viability by different antagonists (reductions to 10–20% control) were found at all three incubation times of 24, 48, and 72 h. The changes induced following 48h and 72h of incubation were similar, and greater than those found following 24h of incubation Effects were also different for the different cell lines. MK-801 and memantine produced dramatic decreases (P<0.0001) in cell viability with IC50s at 48 hours of approximately 600 μM for both SKBR3 and MCF-7, with MK-801; and of approximately 250 μM for SKBR3 and 200 μM for MCF-7 with memantine. Results are depicted in two manners. Figure 3 represents changes in cell growth with selected MK-801 and memantine treatments of cell line MCF-7 as indexed by increases in fluorescence. Figures 5&6 allow all of the data to be presented as percentage reductions of control cell growth produced by different concentrations of the antagonists.
Figure 3.

NMDAR1 receptors appear to be important for in vitro proliferation of breast cancer cells. Reduction in cell viability of MCF-7 cells depicted as estimated cell growth (means ± SEM) produced in the presence of different concentrations (μM) of irreversible NMDAR1 receptor antagonists (A) Memantine, and (B) MK-801(dizocilpine maleate), after 24 h, 48 h and 72 h of treatment. Symbols for each concentration of antagonist used are provided in the figure. Slopes for control (0 μM) and 100 μM MK-801 coincide. Cell viability was assessed as relative increases (mean± SEM) in specific fluorescence of Alamar Blue and estimated growth was reduced to about 10% of control. Reductions were highly significant (p<0.0001) and IC50s were obtained with ~200 μM and ~500–600 μM doses of memantine and MK-801 at 48 and 72 hours of incubation.
Figure 5.

NMDAR1 receptors appear to be important for in vitro proliferation of breast cancer cells. Depiction of data as percentage reduction in cell viability (means ± SEM) produced by different concentrations of irreversible antagonist memantine on (A) MCF-7 cells, and (B) SKBR3 cells, after 24 h (□), 48 h (●) and 72 h (△) of treatment. Cell viability was assessed using Alamar Blue and was reduced to 10% of control. Reductions were highly significant (p<0.0001) and IC50s were obtained with ~200 μM doses of antagonist at 48 and 72 hours of incubation.
Figure 6.
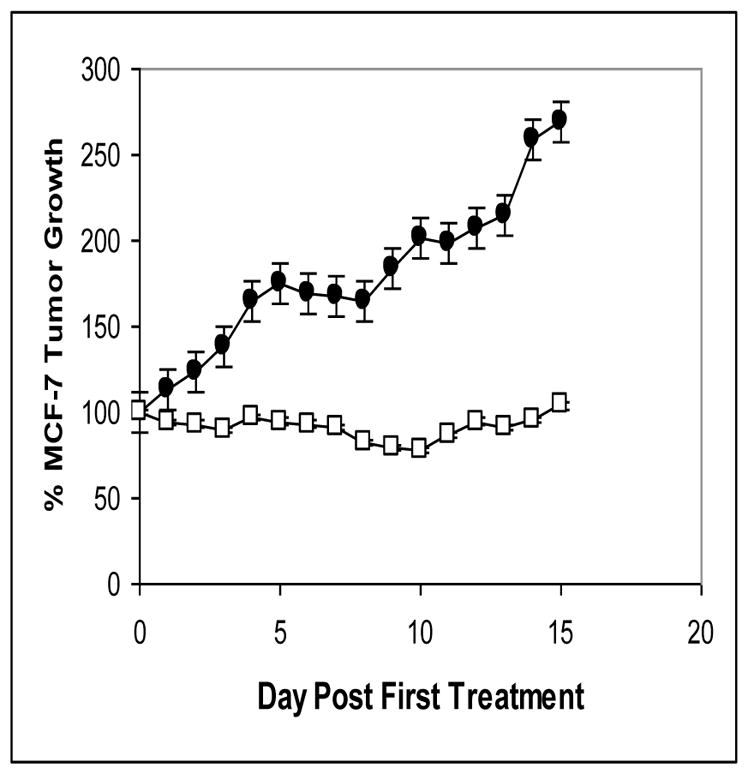
NMDAR1 receptors influence on the growth of human breast cancer tumor xenografts in nu/nu mice. Administration of MK-801 in doses of 0.3 mg/kg twice daily for 10 days (days 0 through 9) completely prevented growth of MCF-7 xenografts (p<0.0001) while having no apparent impact on animal health. Shown as means ±SEM are values for control (■) and treated (□) animals. Tumor volume measured by micrometer was expressed as the product of length x width x depth and was evaluated on a daily basis, as was animal body weight.
A Receptor Antagonist Stops Tumor Growth
The treatment of established MCF-7 tumor xenografts in nu/nu mice with MK-801 for the dosing used produced no noticeable behavioral effects in the animals and had no apparent effect on their well-being as exemplified by body weight in treated animals paralleling that for controls. At the time treatment commenced the volume of tumors was 867± 188 mm3 and 986 ± 65 mm3 for the control group and MK-801 group, respectively. Tumors of the control group had a doubling time of about 9 days over the a 15 day period of observation. MK-801 treatment resulted in a complete inhibition of tumor growth that was highly significant (p<0.001) over the course of the study (Figure 6).
Discussion
Most breast cancers have been described as adenocarcinomas and presumed to arise from breast epithelial or myoepithelial tissues [15]. Alternatively, Pierce in the APUD hypothesis [16] has proposed that neuroendocrine tumors arise from neuroendocrine cells, which in turn are derived during embryonic development from cell of the neural crest. Breast cancer might therefore not be expected to display markers generally associated with the central nervous system, such as functional NMDA receptors. However, we propose the findings presented here for breast cancer cell lines and tissues, provide strong evidence that NMDA receptors are components of at least high grade breast tumors, represented by tubular and ductal carcinomas. The breast cancer cells in culture, both of which are derived from estrogen-dependent disease, were shown to express mRNAs for NMDAR1 and NMDAR2B that appears to be the same as those found by others in the human brain, and, in the case of NMDAR1, by us, for a human neuroblastoma cell line [1] and for small-cell lung cancer. Partial sequencing of these messages show sequence identity with messages from these other sources. These mRNAs were shown by Western analysis to give rise to receptor proteins of approximately 120 kDaltons (NMDAR1) and 170 kDaltons (NMDAR2B), sizes similar to functional brain receptors [3]. However, the commercial anti-NMDAR2 antibodies used could not reveal which of the known NMDAR2 sub-types were present on these cells, so it is possible that they represent all or any one of these receptors. The presence of NMDA R1 and R2 receptors on breast cancer as markers could result from the oncogenetic process in formation of this disease, and, if so, would support the STEPS hypothesis advanced by us that emphasizes process over origin [17]. STEPS is seen as a needed alternative to the APUD concept of Pierce [16].
The effects of the non-reversible NMDAR1 antagonists Memantine and Dizocilpine (MK801) shown here demonstrate that NMDA receptors, as functional calcium channels, are playing an active role in breast cancer survival and growth. These effects were produced by amounts of MK-801 found to alter neuronal activity in culture [18, 19], and growth promoting cascades in non-small cell lung carcinoma cells in culture [20]. Both antagonists dramatically reduced cell viability of the two cell lines examined, with the most effective action being from memantine, which seems to be approximately twice as effective as MK801, despite it having a much lower binding affinity than the latter antagonist. As mentioned above, it has been proposed that unlike MK-801, memantine appears to selectively block over-stimulated (“pathological”) open channels and have very few side-effects through normally functioning channels [6, 7]. If verified in vivo, we can anticipate an agent with the same or similar actions could have a selective action on the NMDA receptors in patients with breast cancer.
A proof of the above principle that NMDA receptors are important for breast cancer growth and development is found in the influence MK-801 has on the growth of tumor xenografts of Mcf7 cells in nude mice. A complete cessation of growth was found at the end of treatment, at doses of antagonist that had no apparent negative influence on the health of animals. The ideal dose levels of the antagonist and duration of treatment to achieve maximal impact on tumor growth has yet to be determined, as has the influence of other NMDAR1 and NMDAR2 antagonists. The dose chosen is the same as that used by others to increase the survival time of mice implanted with lung adenocarcinoma cells [20]. It is similar to those amounts known to have some influence behavior in rodents [21, 22, 23]. Such an influence would impose a limitation in the use of small molecules such as MK-801 because they can readily cross the blood-brain barrier and perhaps disrupt neuronal activities. Since the levels of MK-801 antagonist shown to be effective in our studies had no obvious pathological effects reflected in adverse animal behavior while having ready access to the CNS, it is conceivable it could have therapeutic benefits is treating brain metastases. However, for treatment of all peripheral disease, a safer alternative could be use of NMDA receptor-blocking antibodies as targeting agents in patients. That such antibody approaches to treatment would not adversely affect individuals, despite the reported presence of NMDA receptors in some peripheral tissues [24], is suggested by the work of During and coworkers [25] who were able to perform safe oral vaccination in animals against NMDAR1 receptors.
Our findings demonstrate that functional NMDA receptors are expressed by breast cancer and are important agents for maintaining cell growth and viability. As such they can represent and opportunity for development of new and effective treatments for this disease.
Figure 4.

NMDAR1 receptors appear to be important for in vitro proliferation of breast cancer cells. Depiction of data as percentage reduction in cell viability (means ± SEM) produced by different concentrations of irreversible NMDAR1 receptor antagonist MK-801(dizocilpine maleate) on (A) MCF-7 cells, and (B) SKBR3 cells, after 24 h (□), 48 h (●) and 72 h (△) of treatment. Cell viability was assessed using Alamar Blue and was reduced to about 10% of control. Reductions were highly significant (p<0.0001) and IC50s were obtained with ~500–600 μM doses of antagonist at 48 and 72 hours of incubation.
Acknowledgments
Launa Lynch was a postdoctoral fellow on NIH Training Grant T32 DK 07508 (William G. North, Director). Excellent technical assistance was provided by Alison North and Maudine Waterman.
References
- 1.North WG, Fay MJ, Du JL, Cleary M, Gallagher JD, McCann FV. Presence of functional NMDA receptors in a human neuroblastoma cell-line. Molecular and Chemical Neuropathology. 1997;30:77–94. doi: 10.1007/BF02815151. [DOI] [PubMed] [Google Scholar]
- 2.Danilczuk Z, Ossowska G, Lupina T, Ciesslik K, Zebrowska-Lupina I. Effect of NMDA receptor antagonists on behavioral impairment induced by chronic treatment with dexamethasone. Pharmacol Rep. 2005;57:47–54. [PubMed] [Google Scholar]
- 3.Carpenter D. NMDA receptors and the molecular mechanisms of excitotoxicity. In: Johnson P, Boldyrev AA, editors. Oxidative Stress at the Molecular, Cellular, and Organ Levels. Research Signpost; Trivandrum: 2002. pp. 77–88. [Google Scholar]
- 4.Hinoi ET, Takarada T, Ueshima T, Tsuchihashi Y, Yoneda Y. Glutamate signaling in peripheral tissues. Eur J Biochem. 2004;271:1–13. doi: 10.1046/j.1432-1033.2003.03907.x. [DOI] [PubMed] [Google Scholar]
- 5.Sonkusare SK, Kaul CL, Ramarao P. Dementia of Alzheimer’s disease and other neurodegenerative disorders-memantine, a new hope. Pharmacol Rev. 2005;51:1–17. doi: 10.1016/j.phrs.2004.05.005. [DOI] [PubMed] [Google Scholar]
- 6.Plosker GL, Lyseng-Williamson KA. Memantine:A pharmacoeconomic review of its use in moderate-to-severe Alzheimer’s disease. Pharmacoecon. 2005;23:193–206. doi: 10.2165/00019053-200523020-00010. [DOI] [PubMed] [Google Scholar]
- 7.Kashiwagi K, Masuko T, Nguyen CD, Kuno T, Tanaka I, Igarashi K, Williams K. Channel blockers acting at N-methyl-D-aspartate receptors:differential effects of mutations in the vestibule and ion channel pore. Molec Pharmacol. 2002;61:533–545. doi: 10.1124/mol.61.3.533. [DOI] [PubMed] [Google Scholar]
- 8.Fredriksson A, Danysz W, Quack G, Archer T. Co-administration of memantine and amantadine with sub/suprathreshold doses of L-dopa restores motor behaviour of MPTP-treated mice. J Neural Transm. 2001;108:167–187. doi: 10.1007/s007020170086. [DOI] [PubMed] [Google Scholar]
- 9.North WG, Maurer LHH, Valtin JF. O’Donnell Human neurophysins as potential tumor markers for small-cell carcinoma of the lung: Application of specific radioimmunoassays for vasopressin-associated and oxytocin-associated neurophysins. J Clin Endocrinol Metab. 1980;51:892–896. doi: 10.1210/jcem-51-4-892. [DOI] [PubMed] [Google Scholar]
- 10.North WG, Pai S, Friedmann AS, Fay MJ, Yu XM, Memoli VA. Vasopressin gene-related products are markers of human breast cancer. Breast Cancer Res Treat. 1995;34:229–235. doi: 10.1007/BF00689714. [DOI] [PubMed] [Google Scholar]
- 11.North WG. Neuropeptide production by small cell carcinoma: Vasopressin and oxytocin as plasma markers of disease. J Clin Endocrin Metab. 1991;73:1316–1320. doi: 10.1210/jcem-73-6-1316. [DOI] [PubMed] [Google Scholar]
- 12.Moriyoshi K, Masu M, Ishu T, Shigemoto R, Mizuno N, Nakanishi S. Molecular cloning and characterization of the rat NMDA receptor. Nature. 1991;354:31–37. doi: 10.1038/354031a0. [DOI] [PubMed] [Google Scholar]
- 13.Hess SD, Daggett LP, Crona J, Deal C, Lu C-C, Urrutia A, Chavez-Noriega L, Ellis SB, Johnson EC, Velicelebi G. Cloning and Functional Characterisation of Human Heteromeric N-Methyl-D-Aspartate Receptors. J Pharmacol Exper Therap. 1996;278:808–816. [PubMed] [Google Scholar]
- 14.Ahmed SA, Gogal RM, Jr, Walsh JE. A new rapid and simple non-radioactive assay to monitor and determine the proliferation of lymphocytes: an alternative to [3H]thymidine incorporation assay. J Immunol Methods. 1994 Apr 15;170(2):211–24. doi: 10.1016/0022-1759(94)90396-4. [DOI] [PubMed] [Google Scholar]
- 15.Silverstein MJ, et al. Image-Detected Breast Cancer: State of the Art Diagnosis and Treatment. The Breast Journal; International Breast Cancer Consensus Conference, Sponsored by the Keck School of Medicine, University of Southern California; 2002. pp. 70–76. [DOI] [PubMed] [Google Scholar]
- 16.Pierce AGE. The cytochemistry and ultrastructure of polypeptide hormone producing cells of the APUD series, and the embryologic, physiologic, and pathologic implications of the concept. J Histochem Cytochem. 1969;17:303–313. doi: 10.1177/17.5.303. [DOI] [PubMed] [Google Scholar]
- 17.North WG. Gene regulation of vasopressin and vasopressin receptors in cancer. Exper Physiol. 2000;85S:27–40. doi: 10.1111/j.1469-445x.2000.tb00005.x. [DOI] [PubMed] [Google Scholar]
- 18.Roos DH, Puntel RL, Santos MM, Souza DO, Farina M, Nogueira CW, Aschner M, Burger ME, Barbosa NB, Rocha JB. Guanosine and synthetic organoselenium compounds modulate methylmercury-induced oxidative stress in rat brain cortical slices: involvement of oxidative stress and glutamatergic system. Toxicol in Vitro. 2009;23:302–307. doi: 10.1016/j.tiv.2008.12.020. [DOI] [PubMed] [Google Scholar]
- 19.Longuemare MC, Keung EC, Chun S, Sharp FR, Chan PH, Swanson RA. MK801 reduces uptake and stimulates efflux of excitatory amino acids via membrane depolarization. Am J Physiol. 1996;270:C1398–C1404. doi: 10.1152/ajpcell.1996.270.5.C1398. [DOI] [PubMed] [Google Scholar]
- 20.Andrzej Stepulak A, Sifringer M, Rzeski W, Endesfelder S, Gratopp A, Pohl EE, Bittigau P, Felderhoff-Mueser U, Kaindl AM, Bührer C, Hansen HH, Stryjecka-Zimmer M, Turski L, Ikonomidou C. NMDA antagonist inhibits the extracellular signal-regulated kinase pathway and suppresses cancer growth. Proc Natl Acad Sci U S A. 2005;102:15605–15610. doi: 10.1073/pnas.0507679102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Tricklebank MD, Singh L, Oles RJC, Preston C, Iversen SD. The behavioural effects of MK-801: a comparison with antagonists acting non-competitively and competitively at the NMDA receptor. Eur J Pharmacol. 1989;167:127–135. doi: 10.1016/0014-2999(89)90754-1. [DOI] [PubMed] [Google Scholar]
- 22.Kawabe K, Yoshihara T, Ichitani Y, Iwasaki T. Intrahippocampal d-cycloserine improves MK-801-induced memory deficits: Radial arm maze performance in rats. Brain Research. 1998;814:226–230. doi: 10.1016/s0006-8993(98)01043-9. [DOI] [PubMed] [Google Scholar]
- 23.Gilbert ME. The NMDA-receptor antagonist MK-801 suppresses limbic kindling and kindled seizures. Brain Res. 1988;463:90–99. doi: 10.1016/0006-8993(88)90530-6. [DOI] [PubMed] [Google Scholar]
- 24.Hinoi ET, Takarada T, Ueshima T, Tsuchihashi Y, Yoneda Y. Glutamate signaling in peripheral tissues. Eur J Biochem. 2004;271:1–13. doi: 10.1046/j.1432-1033.2003.03907.x. [DOI] [PubMed] [Google Scholar]
- 25.During MJ, Symes CW, Lawlor PA, Lin J, Dunning J, Fitzsimons HL, Poulsen D, Leone P, Xu R, Dicker BL, Lipski J, Young D. An oral vaccine against NMDAR1 with efficacy in experimental stroke and epilepsy. Science. 2000:1453–1460. doi: 10.1126/science.287.5457.1453. [DOI] [PubMed] [Google Scholar]