

Abstract
Rosemary (Rosmarinus officinalis L.) is one of the most economically important species of the family Lamiaceae. Native to the Mediterranean region, the plant is now widely distributed all over the world mainly due to its culinary, medicinal, and commercial uses including in the fragrance and food industries. Among the most important group of compounds isolated from the plant are the abietane-type phenolic diterpenes that account for most of the antioxidant and many pharmacological activities of the plant. Rosemary diterpenes have also been shown in recent years to inhibit neuronal cell death induced by a variety of agents both in vitro and in vivo. The therapeutic potential of these compounds for Alzheimer's disease (AD) is reviewed in this communication by giving special attention to the chemistry of the compounds along with the various pharmacological targets of the disease. The multifunctional nature of the compounds from the general antioxidant-mediated neuronal protection to other specific mechanisms including brain inflammation and amyloid beta (Aβ) formation, polymerisation, and pathologies is discussed.
1. Introduction
Rosmarinus officinalis L. (family, Lamiaceae), commonly known as rosemary, is one of the most popular perennial culinary herbs cultivated all over the world. Both fresh and dried leaves of rosemary have been used for their characteristic aroma in food cooking or consumed in small amount as herbal tea, while rosemary extracts are routinely employed as natural antioxidant to improve the shelf life of perishable foods. In the latter case, the European Union has approved rosemary extract (E392) as a safe and effective natural antioxidant for food preservation [1]. The plant is also known to be employed in traditional medicines in many countries even far beyond its native Mediterranean region where it grows wild. Among the pharmacologically validated medicinal uses of rosemary are antibacterial [2], anticancer [3, 4], antidiabetic [5], anti-inflammatory and antinociceptive [6–8], antioxidant [5, 9], antithrombotic [10], antiulcerogenic [11, 12], improving cognitive deficits [13], antidiuretic [14], and hepatoprotective [15, 16] effects. The other major use of rosemary is in the perfumery industry where the essential oils are employed as natural ingredients of fragrances.
The culinary, medicinal, and fragrance uses of rosemary are attributed to the vast arrays of chemical constituents collectively known as plant secondary metabolites. Of these, one group are small molecular weight aromatic compounds called essential oils which play vital role in the fragrance and culinary properties of the plant. Essential oils of rosemary dominated by 1,8-cineole, α-pinene, camphene, α-terpineol, and borneol as principal constituents [17, 18] are also responsible for various pharmacological effects of the general antioxidant [8] and antimicrobial [2, 19–25] properties known for many essential oils, as well as other effects including anticarcinogenic activities [26]. The other group of secondary metabolites of rosemary are polyphenolic compounds including the flavonoids (e.g., homoplantaginin, cirsimaritin, genkwanin, gallocatechin, nepetrin, hesperidin, and luteolin derivatives) and phenolic acid derivatives (e.g., rosmarinic acid) [27–29]. By far the most important group of rosemary compounds that gain significant attention in recent years, however, are the unique class of polyphenolic diterpenes. In this review, the chemistry and pharmacology of rosemary diterpenes are scrutinised by giving special emphasis to their therapeutic potential for Alzheimer's disease (AD).
Accounting for an estimated 60 to 80 percent of dementia cases in the elderly populations, AD has become one of the major global health challenges of the century. The worldwide prevalence of dementia is now estimated to exceed 36 million cases with a further projection of 115 million by 2050 [30–32]. One of the current well-accepted pathologies of AD is the “amyloid hypothesis” that puts the accumulation and aggregation of amyloid-beta (Aβ) as the major cause of the progressive neuronal cells deaths in the brain. Neuronal deletion particularly in the cortex region is now known to lead to cognitive impairment including acquired learning skills and memory. The hosts of behavioural symptoms arising from AD include agnosia, aphasia, apraxia, erratic emotion, sleep disorders, and interpersonal/social deterioration [33, 34]. Numerous studies have shown that these clinical symptoms of AD are associated with the loss of cholinergic neurons induced by toxicants such as Aβ, reactive oxygen species (ROS), inflammatory cascades, and excitotoxicity mechanisms. Critical to the AD pathology is the basal forebrain region from where cortical cholinergic neurons originate. The loss of neurons in the basal forebrain has been shown to correlate with the degree and severity of clinical symptoms of AD [34]. To date, the handful of drugs available to treat AD are the acetyl cholinesterase (ACHE) inhibitors (e.g., rivastigmine, galantamine, tacrine, and donepezil) and N-methyl-D-aspartate (NMDA) receptor antagonist (memantine) which have some benefit in alleviating the clinical symptoms of AD [35]. Drug of cure for AD is, however, neither available nor within sight, and the search of new drugs from natural sources should be considered as a viable strategy for the future control of the disease. One group of compounds of interest are the rosemary diterpenes which are comprehensively assessed in this communication for their therapeutic potential for AD. Special emphasis is given to the structural features of the compounds with respect to their effects against specific AD target.
2. Overview of Rosemary Diterpenes
2.1. Biosynthetic Perspective
Biosynthetically, diterpenes are derived from the terpenoids or mevalonate pathway and hence composed of repeating 5-carbon backbone skeleton, isoprene unit(s). The two known isoprene building blocks are isopentenyl pyrophosphate (IPP, 1, Figure 1) and dimethylallyl pyrophosphate (DMAPP, 2) that polymerises in head-to-tail fashion to form the 20-carbon diterpene precursor (4 isoprene units) called geranylgeranyl pyrophosphate (GGPP, 3). The processing of the GGPP through reactions including cyclization, aromatisation, rearrangements, and a series of reaction steps emanating from the loss of the phosphate group, including removal of the carbonium ion, results in the formation of the diterpene subgroups. The class of diterpenes in rosemary identified so far is the abietane type (5–7) which is composed of six–membered tricyclic ring system of which one is aromatic (e.g., 7) [36]. Biosynthetically, abietane-type diterpenes are known to derive from their immediate precursor, labdane subclass (4), as shown in Figure 1. The labdane group of diterpenes on their own are diverse natural products that have been shown to include compounds of novel structural and biological significances [37–40].
Figure 1.
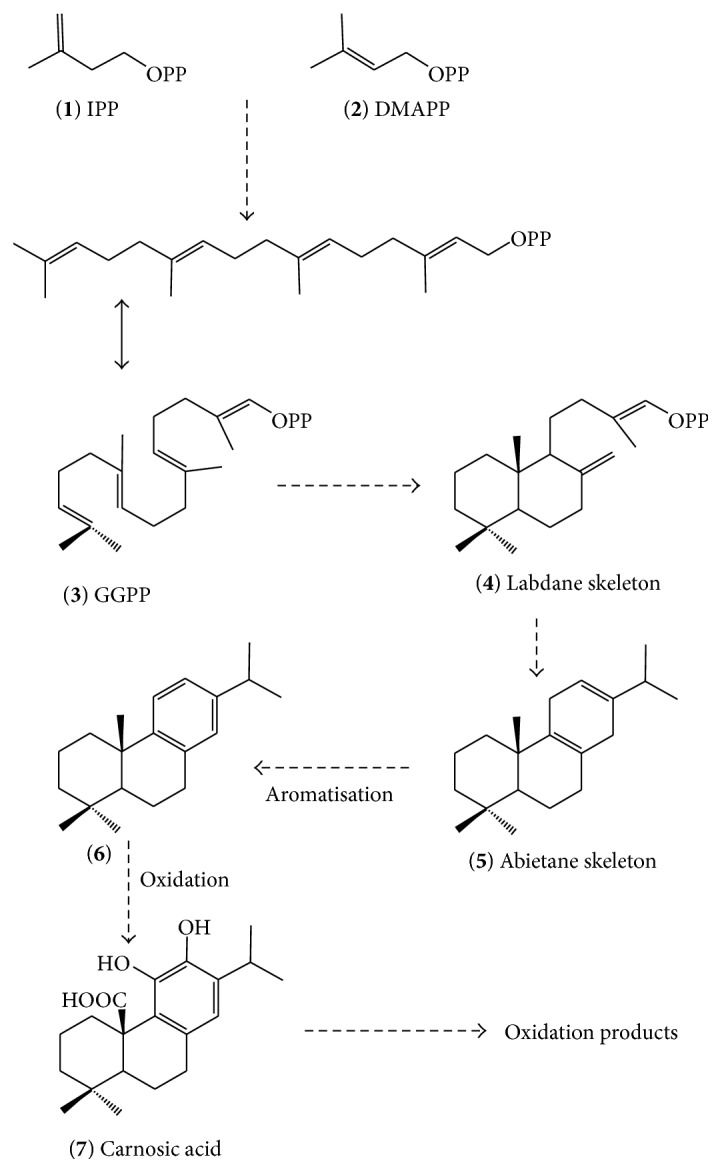
Schematic presentation of the biosynthetic pathway of rosemary diterpenes.
2.2. Diversity of Rosemary Diterpenes
The various types of diterpenes isolated from rosemary are shown in Figure 2. The basic skeleton of all of these diterpenes in rosemary appears to be carnosic acid (7) which was first isolated from the plant by Wenkert et al. [41] in 1965. It is now well known that this compound is the major constituent of rosemary that accounts to 1.5–2.5% of the dried leaves though even higher amounts have been reported [41]. Like many other secondary metabolites, the concentration of carnosic acid (7) and other diterpenes in rosemary could vary due to a host of environmental factors (e.g., sun light intensity and water stress) and growth conditions [42–46] as well as genetic factors as there are now several varieties that could yield the compound in up to 10% yield by dry weight [47]. Carnosic acid (7) is not unique to rosemary and its distribution in sage and other taxonomically related species has been revived recently by Birtić et al. [48]. Other taxonomically unrelated plants such as Premna species have also shown to synthesise pharmacologically significant abietane-type diterpenoids with even more aromatisation than those shown for rosemary diterpenoids in Figure 2 [49].
Figure 2.

Carnosic acid and related abietane-type diterpenes of rosemary.
Although carnosic acid (7) is the principal constituent of rosemary extracts, it is not a very stable compound once extracted and may undergo oxidation to form the γ-lactone diterpene, carnosol (8). In fact, the conversion of (7) to (8) in extracts of R. officinalis and Salvia officinalis has been well documented [50], and the latter was considered as the principal constituent of the plant in earlier studies. In addition to carnosol (8), the oxidation of (7) is also known to yield rosmanol (9) which differs from carnosol by possessing a free hydroxyl group at C-7 position and the γ-lactone formed via the C-20-C-6 route [50–53]. The epimeric form of rosmanol with stereochemistry difference at C-7 position has also been demonstrated by the identification of (11) (epirosmanol [54]). An enzyme catalysed conversion of carnosic acid (7) to lactone derivatives via singlet oxygen-mediated reactions has been suggested as a possible mechanism of these diterpene lactones formation [55–57]. Enzymatic dehydrogenation and free radical attack are now also generally considered as a common route for the formation of various oxidation products of (7) [55, 58]. An alternative structure, isorosmanol (12) [57], where the lactone ring is formed via the C-6 instead of the C-7 hydroxyl position, has also been identified in rosemary extract. The further route of structural diversification in rosemary diterpenes comes through methoxylation and hence the 12-methoxyl derivative of carnosic acid (14) and 11,12-dimethoxy isorosmanol (15) have been identified. Methoxylation at the 7-position is also evident as 7-methoxy-rosmanol (10) has been identified from rosemary [51]. All these diterpenes are relatively polar and are not found in the essential oil of rosemary [59].
The other structurally interesting group of rosemary diterpene derivatives are diterpene quinones (16)–(19) (Figure 3). Mahmoud et al. [60] reported the isolation and structural elucidation of two new abietane-type diterpenoid O-quinones, rosmaquinone A (16) and rosmaquinone B, (17) along with another known diterpene quinone, royleanonic acid (18) and rosmanol. Another example of diterpene quinone identified from rosemary was rosmariquinone (19) [61].
Figure 3.

Glycosylation is the common route of structural diversification in natural products. The study by Zhang et al. [54] has resulted in the identification of polar diterpene glycosides named as officinoterpenosides A1 (20) and A2 (21) (Figure 4). These polar compounds also differ from the carnosic acid derivatives (7–15) not only by their glycosylation and different oxygenation pattern but also by having an altered side chain whereby the 16-methyl group has migrated to the C17 position.
Figure 4.

Munné-Bosch and Alegre [44] have analysed the relative concentrations of diterpenes in rosemary tissues. In general, the level of carnosic acid (7) was about 6-fold higher than other derivatives such as 12-O-methylcarnosic acid (14) and carnosol (8), which (the latter two) were found in similar concentrations. On the other hand, isorosmanol (11) was found at slightly lower concentrations than carnosol (8) while the 11,12-di-O-methylisorosmanol (15) was about 10 times less abundant than isorosmanol (11). The rosmanol (9) concentration is regarded as a trace amount [44]. The most important diterpenes in terms of biological significance of the rosemary however remain to be carnosic acid (7) and carnosol (8) which are most abundant (~5% the dry weight) and shown to account for over 90% of rosemary's antioxidant effects [48, 62]. Dried rosemary could contain about 0.2–1% carnosol (8) [63] while many commercially available extracts may be optimised to contain approximately 10.3% carnosol (8) [64].
Bioavailability. Doolaege et al. [65] have studied the absorption, distribution, and elimination of carnosic acid (7) in rats following administration via the intravenous (20.5 ± 4.2 mg/kg) and oral (64.3 ± 5.8 mg/kg) routes. Their study revealed that the bioavailability of (7) after 360 min following the intravenous dosage was 40.1%. The study also showed that traces of (7) were found in various organs in its free form while elimination in the faeces after 24 h after oral administration was 15.6 ± 8.2% [65]. Another study by Vaquero et al. [66] emphasised on the oral route of (7) where the glucuronide conjugates were found to be the main metabolites detected in the gut, liver, and plasma. The other metabolites identified were the 12-methyl ether and 5,6,7,10-tetrahydro-7-hydroxyrosmariquinone of (7) [66]. Since these metabolites were detected as early as 25 min following oral administration, it was reasonable to conclude that rosemary diterpenes are bioavailable. Interestingly, the free form of (7) as well as its metabolites was detected in the brain [66] suggesting possible effect in this vital organ.
3. Pharmacological Targets of Rosemary Diterpenes Related to AD Therapy
3.1. General Pharmacological Effect of Rosemary Diterpenes on the Brain and Memory
In an attempt to investigate the effect of rosemary tea consumption on brain function, Ferlemi et al. [67] have recently tested the potential anxiolytic- and antidepressant-like behaviour effect on adult male mice. The result showed that oral intake of rosemary tea for 4 weeks has shown a positive effect without altering memory/learning when assessed by passive avoidance, elevated plus maze and forced swimming tests. In an olfactory bulbectomy procedure in mice, MacHado et al. [68] have also demonstrated that rosemary extract possesses antidepressant-like effect and is also able to abolish ACHE alterations although the spatial learning deficit induced by the procedure was not altered. Carnosic acid (7) has also shown to have neuroprotective effects on cyanide-induced brain damage in cultured rodent and human-induced pluripotent stem cell-derived neurons in vitro and in vivo in various brain areas of a non-Swiss albino mouse model [69]. As discussed in the later sections, this effect is likely to be mediated via upregulation of transcriptional pathways related to antioxidant and anti-inflammatory mechanisms [69]. Protective effects of carnosol (8) on rotenone-induced neurotoxicity in cultured dopaminergic cells were also observed in vitro in parallel with downregulation of apoptotic mechanisms [70]. It is also worth noting that other components of rosemary, such as essential oil constituents, are known to alter brain function at therapeutic doses. For example, the cognitive enhancing power of rosemary component, 1,8-cineole, has been well documented [71]. In agreement with these observed effects of the isolated compounds (7, 8), the crude extract of rosemary has been shown to improve memory impairment when tested in vivo using the scopolamine-induced dementia model of AD [72].
3.2. Antioxidant Mechanisms
A number of simple in vitro experiments where the antioxidant potential of rosemary diterpenes is demonstrated include lipid peroxidation and protection of cells from oxidative cell death [73, 74]. Readers must however bear in mind that the antioxidant potential of rosemary extracts and diterpenes on food preservation and various biological models have been established up to the level of large-scale commercial exploitations. The emphasis in this communication is therefore limited to highlighting mechanisms relevant to neurodegenerative diseases. In this respect, Hou et al. [75] have shown that carnosic acid (7) protects neuronal cells from ischemic injury by scavenging ROS. The antioxidant mechanisms of (7) and carnosol (8) are dependent on the loss of hydrogen from their phenolic hydroxyl groups leading to formation of quinone derivatives [76, 77]. Through this antioxidant mechanism, (7) can protect neuronal cells from oxidative damage both in vitro and in vivo. Numerous reports during the last few decades including ours have shown that the antioxidant mechanism and/or radical scavenging effect of polyphenolic natural products is exceptionally prominent when the compounds possess the catechol functional group [78–89]. The formation of the various diterpene derivatives as the oxidation products of (7) is also inherently related to its ability to interact with ROS [50, 90].
The induction of phase II detoxifying enzymes is an important defence mechanism for the removal of xenobiotics and other toxicants of internal and external origin. A large body of evidence to date indicates that the erythroid derived 2-related factor 2 (Nrf-2) is involved in the antioxidant response elements- (AREs-) mediated induction of genes for a variety of antioxidant enzymes, including phase II detoxifying enzymes [91–93]. The expression of many thiol-regulating enzymes, such as glutathione S-transferase, glutamylcysteine synthetase, and thioredoxin reductase, has also shown to be dependent on Nrf-2 [94]. Of the various mechanisms described for these antioxidant effects is direct S-alkylation of the cysteine thiol of the Kelch-like ECH-associated protein 1 (Keap1) protein by the “electrophilic” quinone derivative of (7) [95]. Keap1 is a regulatory protein associated with the transcriptional factor Nrf2 that binds to the ARE [96]. The binding of electrophiles compounds with the cysteine residues on Keap1 protein and the subsequent S-alkyl adduct formation will allow the migration of the Nrf2 to the nucleus. Nrf2 can then promote genes expression by binding to AREs of phase II genes. Through this mechanism, the application of electrophile compounds as antioxidant and neuroprotective agents has been well documented in the various literature [95, 97–99].
Carnosol (8) possesses high electrophilic activity and has been reported to activate Nrf2, phase II detoxifying enzyme genes, and antioxidant enzymes [99, 100]. Direct interaction of (8) with cysteine residues of the nuclear factor kappa B (NF-κB) has also been demonstrated [101–103]. In a similar manor, carnosic acid (7) has been shown to protect neuronal HT22 cells through activation of the antioxidant-responsive element [104]. The free carboxylic acid and catechol hydroxyl moieties have been shown to play critical role in these effects [104]. All the available evidence now therefore suggests that the major rosemary constituents (7 and 8) protect neurons against oxidative stress by activating the Keap1/Nrf2 pathway. Xiang et al. [104], for example, have demonstrated that (7) and (8) could protect HT22 cells against oxidative glutamate toxicity through mechanisms involving activation of the transcriptional ARE of phase II genes including heme oxygenase-1, NADPH-dependent quinone oxidoreductase, and γ-glutamyl cysteine ligase, all of which provide neuroprotection by regulating the cellular redox system. Through antioxidant mechanism, (7) does also protect the lipopolysaccharide- (LPS-) induced liver injury through enhancement of the body's cellular antioxidant defence system as the levels of superoxide dismutase, glutathione peroxidase, and glutathione in serum and liver after the LPS challenge were restored [105]. Pretreatments of RAW264.7 macrophages with (7) also resulted in a significant reduction of the hydrogen peroxide- or LPS-induced generation of ROS and nitric oxide while the heme oxygenase-1 (HO-1) protein expression was time- and dose-dependently upregulated [76]. Moreover, carnosol (8) has been shown to enhance the glutathione S-transferase (GST) and quinone reductase activity in vivo [105].
The therapeutic potential of rosemary diterpenes for AD must be seen in conjunction with the role of oxidant-antioxidant mechanisms in the pathology of the disease. A number of studies have clearly outlined the direct association between ROS-mediated macromolecular cell damage and neuronal cell death in AD, particularly in brain regions where Aβ is highly prevalent [106, 107]. Interestingly, neuronal cells in the brain appear to be more susceptible to ROS-mediated cell damage than any other cell types for numerous reasons including high oxygen consumption [108], high level of polyunsaturated fatty acids content of cell membrane [109], association of the NMDA receptor activation with ROS-induced neuronal apoptosis [110], and poor level of antioxidant defences including the catalase, glutathione peroxidase, and vitamin E contents [111]. Furthermore, antioxidant defences in AD have been found to be highly suppressed as low level of SOD [112] and reduced form of glutathione (GSH) [113, 114] as well as mitochondrial dysfunction [115] are all common features of AD. Hence, the numerous reports on the antioxidant effects of rosemary diterpenes along with their specific effect on neuronal cells through the abovementioned antioxidant mechanisms imply that they should be considered for further development as anti-Alzheimer's agents.
Metal Chelation. High level of metal ions such as copper, zinc, and iron have been found in the amyloid plaques of AD brains [116–118]. Higher millimolar level of unregulated metal ions in the brain has also been shown to arise due to age related deterioration of the blood-brain-barrier leading to unchecked access of the brain to metal ions [119]. As described in the later section, these metal ions play critical role in Aβ-induced neurotoxicity in AD. Hence, a potent metal chelative effect of a drug is an important feature of anti-AD therapy. Our own study on polyphenolic compounds in the last two decades has revealed that their biological effect including enzyme inhibition could be partly explained by their ability to chelate iron and other redox metals and, for such effect, one of the best structural features in a molecule is the orthodihydroxyl functional moiety [78–89].
The structural features of (7) and (8) are in favour of strong metal chelation properties. Carnosol (8) has been shown to inhibit Cu2+-induced LDL oxidation [120] but, most importantly, metal (e.g., iron) chelation is one of the known mechanisms of antioxidant effects. Furthermore, iron absorption from the gut is strongly suppressed by rosemary extract [121].
3.3. Anti-Inflammatory Mechanisms
The roles of Nrf2 and the antioxidant protein HO-1 in neuroinflammatory response have been well established. The search for effective Nrf2/HO-1 activators that modulate the microglia inflammatory response in AD would therefore have significant therapeutic value. A recent study has further revealed that Nrf2 activation inhibits inflammatory gene expression [122] through mechanisms involving HO-1 [123]. Lian et al. [124] have also shown that carnosol (8) and rosemary essential oils inhibit the adhesion of tumour necrosis factor-α- (TNF-α-) induced monocytes to endothelial cells and suppress the expression of intercellular adhesion molecule (ICAM-1) at the transcriptional level in vitro. The anti-inflammatory effect of (8) via inhibition of the TNF-α-induced protein expression of ICAM-1 was also shown to be extended to other cell surface molecules such as the vascular cell adhesion molecule- (VCAM-) 1 and E-selectin in endothelial cells as well as interleukin- (IL-) 8 and the monocyte chemoattractant protein- (MCP-) 1 [125]. Moreover, Foresti et al. [126] have shown that (8) inhibits the TNF-α-induced signaling pathways through inhibition of inhibitor of nuclear factor kappa-B (IKK-β) activity as well as the upregulation of HO-1 expression. At the concentration of 5–20 μM, (8) was demonstrated to upregulate Nrf2 and HO-1 leading to downregulation of the inflammatory response (TNF-α, prostaglandin E-2, and nitrite) [126]. Carnosic acid (7) was similarly shown to inhibit the expression of cytokine-induced adhesion molecules on endothelial cells surface leading to inhibition of monocyte-cell adhesions [127]. It does also potently inhibit the LPS-induced rise in serum levels of the proinflammatory cytokines (TNF-α and IL-6) in vivo [128]. Both (7) and (8) have also shown to inhibit the phorbol 12-myristate 13-acetate- (PMA-) induced ear inflammation in mice with EC50 of 10.20 μg/cm2 and 10.70 μg/cm2, respectively. This activity was coupled with reduced level of expression of IL-1β and TNF-α and cyclooxygenase-2 (COX-2). In another study [129], both (7) and (8) inhibited the formation of proinflammatory leukotrienes in cells with IC50 of 7–20 μM as well as purified recombinant 5-lipoxygenase (IC50 = 0.1–1 μM). The study also showed that both (7) and (8) potently antagonise intracellular Ca2+ mobilisation induced by a chemotactic stimulus, coupled with inhibition of ROS generation [129]. The LPS-induced nitric oxide production in Raw 264.7 cells was also shown to be inhibited by (8) with IC50 of 9.4 μM [130]. In an in vitro model of brain inflammation, (7) inhibited the LPS-induced activation of cells of the mouse microglial cell line MG6 [131], releasing inflammatory cytokines such as IL-1β and IL-6. The nitric oxide production associated with a decrease in the level of inducible nitric oxide synthase has also been reported for (7) [131].
Glial cells are the major inflammatory cells of the brain which produce massive amount of proinflammatory cytokines (e.g., IL-1β, IL-6, and TNF-α) upon activation. Numerous studies have highlighted the fact that high levels of these inflammatory cytokines are critical in the coordination of brain inflammation in AD [132, 133]. Moreover, both microglia and astrocytes have been shown to be highly regulated in AD brains [133, 134]. The potent anti-inflammatory activity of rosemary diterpenes in both the microglial cells [126, 135] and other inflammatory models therefore suggests their potential in tackling AD.
3.4. Aβ Mechanisms
Generally, amyloid plaques and neurofibrillary tangles (NFT), which are closely linked to the formation of toxic insoluble aggregates of Aβ, have shown to be the two most common pathological hallmarks of AD [136–138]. The Aβ is formed from the neuronal transmembrane glycoprotein (100–130 kDa) called the amyloid precursor protein (APP). The α-, β-, and γ-secretases are the three major proteolytic enzymes that process APP [139] through two major pathways: the amyloidogenic and nonamyloidogenic pathways. The non-amyloidogenic-dependent pathway involves APP processing through α-secretase leading to the generation of nonpathogenic amyloid products. In the amyloidogenic pathway, β-secretase processes APP at the N-terminus of the Aβ domain to generate the membrane-attached fragment, C99, and the sAPPβ fragment [140]. Further cleavage of the C99 fragment by γ-secretase leads to the formation of the two most common forms of Aβ peptides, Aβ1–40 (90%) and Aβ1–42 (10%), along with other fragments. To date, a number of therapeutic agents that inhibit APP processing have been identified and some appear to be in clinical trials [141]. Of these, inhibitors of β-secretase 1 (BACE1) appear to be most important as this enzyme takes the first rate limiting step in APP processing [142]. To the best of the author's knowledge, an inhibitory effect of rosemary diterpenes on β-secretase activity has not been demonstrated but a promising effect on α-secretase has been reported by Meng et al. [143]. In their study using the SH-SY5Y human neuroblastoma cells, carnosic acid (7) showed 61% suppression of Aβ42 secretion when tested at the concentration of 30 μM. The effect was also coupled with enhanced mRNA expressions of α-secretase but not the β-secretase BACE1. Hence, the mechanism of action of (7) for APP processing inhibition appears to be through promotion of the normal non-amyloidogenic-dependent pathway. Similar results were also demonstrated by Yoshida et al. [144] where Aβ peptides (1–40, 1–42, and 1–43) production in U373MG human astrocytoma cells was suppressed by (7) (50 μM). The study also revealed a 55 to 71% inhibition of Aβ release coupled with effect on mRNA expressions of an α-secretase, but once again not the β-secretase BACE1 [144].
Once Aβ is formed, it undergoes a serious of polymerisation processes leading to the formation of insoluble precipitates. It has been shown that small soluble oligomers as well as amyloid fibril aggregates induce toxicity to neuronal cells in AD [145–149]. Hence, various classes of natural and synthetic compounds that inhibit the polymerisation and stability of Aβ aggregates can be employed as viable therapeutic agents for AD. Some of these agents identified to date include chrysamine G [150], oligopeptides [151–155], and plant polyphenols such as curcumin, myricetin, morin, quercetin, kaempferol (+)-catechin, (−)-epicatechin, nordihydroguaiaretic acid and tannic acid [156–158], antibiotics (e.g., rifampicin [159]), and aspirin [160]. In this connection, Meng et al. [143] have recently investigated the effect of carnosic acid (7) on the viability of cultured SH-SY5Y human neuroblastoma cells challenged by Aβ42 or Aβ43. The cellular deletion in these cells treated with Aβ42 or Aβ43 (monomer, 10 μM each) was reported to be partially reversed by treatment with (7) (10 μM). The observed effect was also coupled with reduced level of cellular oligomers of Aβ42 and Aβ43 suggesting inhibition of oligomerisation as the possible mechanism of action [143]. These data were also in agreement with the in vivo observation where (7) has been demonstrated to show beneficial effect in AD models [161]. Rasoolijazi et al. [162] also provided direct evidence to demonstrate the therapeutic potential of (7) for AD by using Aβ toxicity in vivo. When Aβ (1–40) was injected into the Ca1 region of the hippocampus of rats, neurodegeneration and cognitive impairment were evident as assessed by the passive avoidance learning and spontaneous alternation behaviour tests. Treatment by (7) appears to reverse these Aβ (1–40) mediated changes suggesting the therapeutic potential of this compound for AD [162]. The association between Aβ formation and aggregation with metal ions such as copper has been reviewed in many literatures [163–166]. In agreement with this finding, metal chelators have been shown to decrease Alzheimer Aβ plaques [167]. It is now also known that Aβ is a redox-active peptide that reduces transition metals like Cu2+ and Fe3+ leading to the generation of ROS [168]. Both the polymerisation and toxicity of Aβ are therefore intimately linked to metal ions and ROS [169]. The polymerisation of Aβ itself is shown to be enhanced when the antioxidant defence is diminished [112, 170]. The multifunctional nature of rosemary diterpenes in metal chelation and ROS scavenging is thus likely to contribute to their effect against Aβ polymerisation and toxicity.
3.5. ACHE Activity
The impairment of memory and cognitive power in AD has been shown to be associated with the loss of cholinergic neurons in the cortex [171–174]. Under this circumstance where the acetyl choline (ACH) activity in this region is below the normal level, one approach of therapeutic intervention in AD is to minimise the degradation of ACH by its enzyme, ACHE. Even though such drugs have limitation due to their undesirable side effects, an overall beneficial effect in cognitive improvement and behavioural symptoms have been clinically observed [175]. Szwajgier [176] has studied the effect of carnosic acid (7) against ACHE along with 35 other phenolic compounds. Interestingly, CA was identified as the most potent. In silico molecular interaction study approach on AChE inhibitors has also resulted in the identification of (7) as a potential lead drug candidate [177]. The memory enhancing effect of rosemary extract (200 mg/kg, p.o.) in the scopolamine-induced dementia model of AD has also been shown to be linked with direct effect on ACHE activity [72]. While the mRNA expression of butyrylcholinesterase (BuChE) in the cortex was inhibited, its expression in the hippocampus was enhanced by rosemary extract [72]. These effects on the expression of enzymes however could be mediated through indirect effect via other mechanisms.
4. General Summary and Conclusion
The industrial scale exploitation of rosemary for food preservation and as natural antioxidant additives is attributed to its phenolic constituents. The predominant phenolic compounds that accounts for such effects as well as the various in vitro and in vivo pharmacological properties of the plant are the abietane type of diterpenes. Structurally, these groups of compounds are based on the steroidal-like terpenoid skeleton but have added pharmacophore of a phenolic structure. The rosemary diterpenoids of pharmacological relevance are represented by (7) and (8) where the diorthohydroxyl/catecholic functional group is evident. Through these structural features, these compounds display a vast array of pharmacological effects ranging between antioxidant, metal chelation, and anti-inflammatory properties. These very mechanisms do also appear to be involved in the potential therapeutic effect of the compounds for AD. The further effect of rosemary diterpenes in Aβ formation, aggregation, and toxicity accounts for their additional benefit in tackling AD. Given that AD is a complex disease involving many pathological processes, treatment with multifunctional drugs like those demonstrated by rosemary diterpenes constitutes a viable therapeutic approach. The cascade of neurodegeneration process in AD has lots of similarities with other diseases like Parkinson's disease. Interestingly, some of the rosemary diterpenes such as carnosic acid (7) have been shown to have beneficial effect in Parkinson's disease model [178, 179]. It is also worth noting that only (7) and (8) have been extensively investigated for their possible therapeutic effect related to AD. Other interesting diterpenes including the glycosidic forms could have different bioavailability and therapeutic profile. Further research in this field will therefore provide more evidence on the therapeutic potential of rosemary diterpenes. All the available date to date however suggest that their effect on AD is very promising and further research including clinical trials is well warranted.
Conflict of Interests
The author declares that there is no conflict of interests regarding the publication of this paper.
References
- 1.Food Standards Agency. Current EU approved additives and their E Numbers. https://www.food.gov.uk/science/additives/enumberlist.
- 2.Bozin B., Mimica-Dukic N., Samojlik I., Jovin E. Antimicrobial and antioxidant properties of rosemary and sage (Rosmarinus officinalis L. and Salvia officinalis L., Lamiaceae) essential oils. Journal of Agricultural and Food Chemistry. 2007;55(19):7879–7885. doi: 10.1021/jf0715323. [DOI] [PubMed] [Google Scholar]
- 3.Cheung S., Tai J. Anti-proliferative and antioxidant properties of rosemary Rosmarinus officinalis . Oncology Reports. 2007;17(6):1525–1531. [PubMed] [Google Scholar]
- 4.Yesil-Celiktas O., Sevimli C., Bedir E., Vardar-Sukan F. Inhibitory effects of rosemary extracts, carnosic acid and rosmarinic acid on the growth of various human cancer cell lines. Plant Foods for Human Nutrition. 2010;65(2):158–163. doi: 10.1007/s11130-010-0166-4. [DOI] [PubMed] [Google Scholar]
- 5.Bakirel T., Bakirel U., Keleş O. Ü., Ülgen S. G., Yardibi H. In vivo assessment of antidiabetic and antioxidant activities of rosemary (Rosmarinus officinalis) in alloxan-diabetic rabbits. Journal of Ethnopharmacology. 2008;116(1):64–73. doi: 10.1016/j.jep.2007.10.039. [DOI] [PubMed] [Google Scholar]
- 6.González-Trujano M. E., Peña E. I., Martínez A. L., et al. Evaluation of the antinociceptive effect of Rosmarinus officinalis L. using three different experimental models in rodents. Journal of Ethnopharmacology. 2007;111(3):476–482. doi: 10.1016/j.jep.2006.12.011. [DOI] [PubMed] [Google Scholar]
- 7.Nogueira de Melo G. A., Grespan R., Fonseca J. P., et al. Rosmarinus officinalis L. essential oil inhibits in vivo and in vitro leukocyte migration. Journal of Medicinal Food. 2011;14(9):944–949. doi: 10.1089/jmf.2010.0159. [DOI] [PubMed] [Google Scholar]
- 8.Estévez M., Ramírez R., Ventanas S., Cava R. Sage and rosemary essential oils versus BHT for the inhibition of lipid oxidative reactions in liver pâté. LWT—Food Science and Technology. 2007;40(1):58–65. doi: 10.1016/j.lwt.2005.07.010. [DOI] [Google Scholar]
- 9.Laura P.-F., Garzón M. T., Vicente M. Relationship between the antioxidant capacity and effect of rosemary (Rosmarinus officinalis L.) polyphenols on membrane phospholipid order. Journal of Agricultural and Food Chemistry. 2010;58(1):161–171. doi: 10.1021/jf9026487. [DOI] [PubMed] [Google Scholar]
- 10.Yamamoto J., Yamada K., Naemura A., Yamashita T., Arai R. Testing various herbs for antithrombotic effect. Nutrition. 2005;21(5):580–587. doi: 10.1016/j.nut.2004.09.016. [DOI] [PubMed] [Google Scholar]
- 11.Gülçin I., Küfrevioğlu Ö. I., Oktay M., Büyükokuroğlu M. E. Antioxidant, antimicrobial, antiulcer and analgesic activities of nettle (Urtica dioica L.) Journal of Ethnopharmacology. 2004;90(2-3):205–215. doi: 10.1016/j.jep.2003.09.028. [DOI] [PubMed] [Google Scholar]
- 12.Corrêa Dias P., Foglio M. A., Possenti A., De Carvalho J. E. Antiulcerogenic activity of crude hydroalcoholic extract of Rosmarinus officinalis L. Journal of Ethnopharmacology. 2000;69(1):57–62. doi: 10.1016/s0378-8741(99)00133-6. [DOI] [PubMed] [Google Scholar]
- 13.Kennedy D. O., Scholey A. B. The psychopharmacology of European herbs with cognition-enhancing properties. Current Pharmaceutical Design. 2006;12(35):4613–4623. doi: 10.2174/138161206779010387. [DOI] [PubMed] [Google Scholar]
- 14.Haloui M., Louedec L., Michel J.-B., Lyoussi B. Experimental diuretic effects of Rosmarinus officinalis and Centaurium erythraea . Journal of Ethnopharmacology. 2000;71(3):465–472. doi: 10.1016/s0378-8741(00)00184-7. [DOI] [PubMed] [Google Scholar]
- 15.Abdel-Wahhab K. G. E.-D., El-Shamy K. A., El-Beih N. A. E.-Z., Morcy F. A., Mannaa F. A. E. Protective effect of a natural herb (Rosmarinus officinalis) against hepatotoxicity in male albino rats. Comunicata Scientiae. 2011;2(1):9–17. [Google Scholar]
- 16.Sotelo-Félix J. I., Martinez-Fong D., De la Torre P. M. Protective effect of carnosol on CCl4-induced acute liver damage in rats. European Journal of Gastroenterology & Hepatology. 2002;14(9):1001–1006. doi: 10.1097/00042737-200209000-00011. [DOI] [PubMed] [Google Scholar]
- 17.Atti-Santos A. C., Rossato M., Pauletti G. F., et al. Physico-chemical evaluation of Rosmarinus officinalis L. essential oils. Brazilian Archives of Biology and Technology. 2005;48(6):1035–1039. doi: 10.1590/s1516-89132005000800020. [DOI] [Google Scholar]
- 18.Touafek O., Nacer A., Kabouche A., et al. Chemical composition of the essential oil of Rosmarinus officinalis cultived in the Algerian Sahara. Chemistry of Natural Compounds. 2004;40(1):28–29. doi: 10.1023/b:conc.0000025460.78222.69. [DOI] [Google Scholar]
- 19.Valgimigli L. Essential Oils As Natural Food Additives: Composition, Applications, Antioxidant and Antimicrobial Properties. New York, NY, USA: Nova Science Publishers; 2012. [Google Scholar]
- 20.Djeddi S., Bouchenah N., Settar I., Skaltsa H. D. Composition and antimicrobial activity of the essential oil of Rosmarinus officinalis from Algeria. Chemistry of Natural Compounds. 2007;43(4):487–490. doi: 10.1007/s10600-007-0172-4. [DOI] [Google Scholar]
- 21.Dimitrijević S. I., Mihajlovski K. R., Antonović D. G., Milanović-Stevanović M. R., Mijin D. Ž. A study of the synergistic antilisterial effects of a sub-lethal dose of lactic acid and essential oils from Thymus vulgaris L., Rosmarinus officinalis L. and Origanum vulgare L. Food Chemistry. 2007;104(2):774–782. doi: 10.1016/j.foodchem.2006.12.028. [DOI] [Google Scholar]
- 22.Gachkar L., Yadegari D., Rezaei M. B., Taghizadeh M., Astaneh S. A., Rasooli I. Chemical and biological characteristics of Cuminum cyminum and Rosmarinus officinalis essential oils. Food Chemistry. 2007;102(3):898–904. doi: 10.1016/j.foodchem.2006.06.035. [DOI] [Google Scholar]
- 23.Issabeagloo E., Kermanizadeh P., Forughi R. T. Antimicrobial effects of rosemary (Rosmarinus officinalis L.) essential oil against Staphylococcus spp. African Journal of Microbiology Research. 2012;6(23):5039–5042. [Google Scholar]
- 24.Ojeda-Sana A. M., van Baren C. M., Elechosa M. A., Juárez M. A., Moreno S. New insights into antibacterial and antioxidant activities of rosemary essential oils and their main components. Food Control. 2013;31(1):189–195. doi: 10.1016/j.foodcont.2012.09.022. [DOI] [Google Scholar]
- 25.Zaouali Y., Bouzaine T., Boussaid M. Essential oils composition in two Rosmarinus officinalis L. varieties and incidence for antimicrobial and antioxidant activities. Food and Chemical Toxicology. 2010;48(11):3144–3152. doi: 10.1016/j.fct.2010.08.010. [DOI] [PubMed] [Google Scholar]
- 26.Başer K. C., Buchbauer G. Handbook of Essential Oils: Science, Technology, and Applications. New York, NY, USA: CRC Press, Taylor & Francis Group; 2010. [Google Scholar]
- 27.Bai N., He K., Roller M., et al. Flavonoids and phenolic compounds from Rosmarinus officinalis . Journal of Agricultural and Food Chemistry. 2010;58(9):5363–5367. doi: 10.1021/jf100332w. [DOI] [PubMed] [Google Scholar]
- 28.Del Baño M. J., Lorente J., Castillo J., et al. Flavonoid distribution during the development of leaves, flowers, stems, and roots of Rosmarinus officinalis. Postulation of a biosynthetic pathway. Journal of Agricultural and Food Chemistry. 2004;52(16):4987–4992. doi: 10.1021/jf040078p. [DOI] [PubMed] [Google Scholar]
- 29.Okamura N., Haraguchi H., Hashimoto K., Yagi A. Flavonoids in Rosmarinus officinalis leaves. Phytochemistry. 1994;37(5):1463–1466. doi: 10.1016/s0031-9422(00)90434-5. [DOI] [PubMed] [Google Scholar]
- 30.Alzheimer's Society UK. Alzheimers Disease. Demography. October 2015, http://www.alzheimers.org.uk/site/scripts/documents_info.php?documentID=412.
- 31.Alzheimer's Association. October 2015, http://www.alz.org/facts/
- 32.Wortmann M. Dementia: a global health priority—highlights from an ADI and World Health Organization report. Alzheimer's Research and Therapy. 2012;4(5, article 40) doi: 10.1186/alzrt143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Cummings J. L., Back C. The cholinergic hypothesis of neuropsychiatric symptoms in Alzheimer's disease. The American Journal of Geriatric Psychiatry. 1998;6(2, supplement 1):S64–S78. doi: 10.1097/00019442-199821001-00009. [DOI] [PubMed] [Google Scholar]
- 34.Whitehouse P. J., Price D. L., Struble R. G., Clark A. W., Coyle J. T., DeLong M. R. Alzheimer's disease and senile dementia: loss of neurons in the basal forebrain. Science. 1982;215(4537):1237–1239. doi: 10.1126/science.7058341. [DOI] [PubMed] [Google Scholar]
- 35.Raschetti R., Albanese E., Vanacore N., Maggini M. Cholinesterase inhibitors in mild cognitive impairment: a systematic review of randomised trials. PLoS Medicine. 2007;4(11):1818–1828. doi: 10.1371/journal.pmed.0040338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Brückner K., Božić D., Manzano D., et al. Characterization of two genes for the biosynthesis of abietane-type diterpenes in rosemary (Rosmarinus officinalis) glandular trichomes. Phytochemistry. 2014;101:52–64. doi: 10.1016/j.phytochem.2014.01.021. [DOI] [PubMed] [Google Scholar]
- 37.Habtemariam S., Gray A. I., Lavaud C., et al. ent-12-Oxolabda-8,13(16)-dien-15-oic acid and ent-8β,12α-epidioxy-12β-hydroxylabda-9(11),13-dien-15-oic acid γ-lactone: two new diterpenes from the aerial parts of Premna oligotricha . Journal of the Chemical Society, Perkin Transactions 1. 1991;(4):893–896. doi: 10.1039/P19910000893. [DOI] [Google Scholar]
- 38.Habtemariam S., Gray A. I., Waterman P. G. Antibacterial diterpenes from the aerial parts of Premna oligotricha . Planta Medica. 1992;58(1):109–110. doi: 10.1055/s-2006-961404. [DOI] [PubMed] [Google Scholar]
- 39.Habtemariam S. Cytotoxicity of diterpenes from Premna schimperi and Premna oligotricha . Planta Medica. 1995;61(4):368–369. doi: 10.1055/s-2006-958105. [DOI] [PubMed] [Google Scholar]
- 40.Habtemariam S. In vitro antileishmanial effects of antibacterial diterpenes from two Ethiopian Premna species: P. schimperi and P. oligotricha . BMC Pharmacology. 2003;3, article 6 doi: 10.1186/1471-2210-3-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Wenkert E., Fuchs A., Mcchesney J. D. Chemical artifacts from the family Labiatae. Journal of Organic Chemistry. 1965;30(9):2931–2934. doi: 10.1021/jo01020a012. [DOI] [Google Scholar]
- 42.Hidalgo P. J., Ubera J. L., Tena M. T., Valcárcel M. Determination of the carnosic acid content in wild and cultivated Rosmarinus officinalis . Journal of Agricultural and Food Chemistry. 1998;46(7):2624–2627. doi: 10.1021/jf970974j. [DOI] [Google Scholar]
- 43.Kosaka K., Mimura J., Itoh K., et al. Role of Nrf2 and p62/ZIP in the neurite outgrowth by carnosic acid in PC12h cells. Journal of Biochemistry. 2010;147(1):73–81. doi: 10.1093/jb/mvp149. [DOI] [PubMed] [Google Scholar]
- 44.Munné-Bosch S., Alegre L. Subcellular compartmentation of the diterpene carnosic acid and its derivatives in the leaves of rosemary. Plant Physiology. 2001;125(2):1094–1102. doi: 10.1104/pp.125.2.1094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Munné-Bosch S., Alegre L., Schwarz K. The formation of phenolic diterpenes in Rosmarinus officinalis L. under Mediterranean climate. European Food Research and Technology. 2000;210(4):263–267. doi: 10.1007/s002179900108. [DOI] [Google Scholar]
- 46.Tounekti T., Munné-Bosch S. Enhanced phenolic diterpenes antioxidant levels through non-transgenic approaches. Critical Reviews in Plant Sciences. 2012;31(6):505–519. doi: 10.1080/07352689.2012.696457. [DOI] [Google Scholar]
- 47.Wellwood C. R. L., Cole R. A. Relevance of carnosic acid concentrations to the selection of rosemary, Rosmarinus officinalis (L.), accessions for optimization of antioxidant yield. Journal of Agricultural and Food Chemistry. 2004;52(20):6101–6107. doi: 10.1021/jf035335p. [DOI] [PubMed] [Google Scholar]
- 48.Birtić S., Dussort P., Pierre F.-X., Bily A. C., Roller M. Carnosic acid. Phytochemistry. 2015;115:9–19. doi: 10.1016/j.phytochem.2014.12.026. [DOI] [PubMed] [Google Scholar]
- 49.Habtemariam S., Varghese G. K. A novel diterpene skeleton: identification of a highly aromatic, cytotoxic and antioxidant 5-methyl-10-demethyl-abietane-type diterpene from Premna serratifolia . Phytotherapy Research. 2015;29(1):80–85. doi: 10.1002/ptr.5229. [DOI] [PubMed] [Google Scholar]
- 50.Schwarz K., Ternes W. Antioxidative constituents of Rosmarinus officinalis and Salvia officinalis—II. Isolation of carnosic acid and formation of other phenolic diterpenes. Zeitschrift für Lebensmittel-Untersuchung und Forschung. 1992;195(2):99–103. doi: 10.1007/bf01201766. [DOI] [PubMed] [Google Scholar]
- 51.Arisawa M., Hayashi T., Ohmura K., et al. Chemical and pharmaceutical studies on medicinal plants in Paraguay: studies on ‘Romero,’ part 2. Journal of Natural Products. 1987;50(6):1164–1166. doi: 10.1021/np50054a030. [DOI] [PubMed] [Google Scholar]
- 52.Takenaka M., Watanabe T., Sugahara K., Harada Y., Yoshida S., Sugawara F. New antimicrobial substances against Streptomyces scabies from Rosemary (Rosmarinus officinalis L.) Bioscience, Biotechnology and Biochemistry. 1997;61(9):1440–1444. doi: 10.1271/bbb.61.1440. [DOI] [Google Scholar]
- 53.Cuvelier M.-E., Richard H., Berset C. Antioxidative activity and phenolic composition of pilot-plant and commercial extracts of sage and rosemary. Journal of the American Oil Chemists' Society. 1996;73(5):645–652. doi: 10.1007/bf02518121. [DOI] [Google Scholar]
- 54.Zhang Y., Adelakun T. A., Qu L., et al. New terpenoid glycosides obtained from Rosmarinus officinalis L. aerial parts. Fitoterapia. 2014;99:78–85. doi: 10.1016/j.fitote.2014.09.004. [DOI] [PubMed] [Google Scholar]
- 55.Luis J. G., Quinones W., Grillo T. A., Kishi M. P. Diterpenes from the aerial part of Salvia columbariae . Phytochemistry. 1994;35(5):1373–1374. doi: 10.1016/s0031-9422(00)94859-3. [DOI] [Google Scholar]
- 56.Nakatani N., Inatani R. A new diterpene lactone, rosmadial, from Rosemary (Rosmarinus officinalis L.) Agricultural and Biological Chemistry. 1983;47(2):353–358. doi: 10.1271/bbb1961.47.353. [DOI] [Google Scholar]
- 57.Nakatani N., Inatani R. Two antioxidative diterpenes from Rosemary (Rosmarinus officinalis L.) and a revised structure for rosmanol. Agricultural and Biological Chemistry. 1984;48(8):2081–2085. doi: 10.1080/00021369.1984.10866436. [DOI] [Google Scholar]
- 58.Luis J. G. Chemistry, biogenesis and chemotaxonomy of the diterpenoids of Salvia. In: Marborne J. B., editor. Ecological Chemistry and Biochemistry of Plant Terpenoids. Oxford, UK: Clarendon Press; 1991. pp. 63–82. [Google Scholar]
- 59.Jiang Y., Wu N., Fu Y.-J., et al. Chemical composition and antimicrobial activity of the essential oil of Rosemary. Environmental Toxicology and Pharmacology. 2011;32(1):63–68. doi: 10.1016/j.etap.2011.03.011. [DOI] [PubMed] [Google Scholar]
- 60.Mahmoud A. A., Al-Shihry S. S., Son B. W. Diterpenoid quinones from Rosemary (Rosmarinus officinalis L.) Phytochemistry. 2005;66(14):1685–1690. doi: 10.1016/j.phytochem.2005.04.041. [DOI] [PubMed] [Google Scholar]
- 61.Houlihan C. M., Ho C.-T., Chang S. S. The structure of rosmariquinone—a new antioxidant isolated from Rosmarinus officinalis L. Journal of the American Oil Chemists' Society. 1985;62(1):96–98. doi: 10.1007/bf02541500. [DOI] [Google Scholar]
- 62.Santos-Gomes P. C., Seabra R. M., Andrade P. B., Fernandes-Ferreira M. Determination of phenolic antioxidant compounds produced by calli and cell suspensions of sage (Salvia officinalis L.) Journal of Plant Physiology. 2003;160(9):1025–1032. doi: 10.1078/0176-1617-00831. [DOI] [PubMed] [Google Scholar]
- 63.Del Baño M. J., Lorente J., Castillo J., et al. Phenolic diterpenes, flavones, and rosmarinic acid distribution during the development of leaves, flowers, stems, and roots of Rosmarinus officinalis. Antioxidant activity. Journal of Agricultural and Food Chemistry. 2003;51(15):4247–4253. doi: 10.1021/jf0300745. [DOI] [PubMed] [Google Scholar]
- 64.Frankel E. N., Huang S.-W., Aeschbach R., Prior E. Antioxidant activity of a rosemary extract and its constituents, carnosic acid carnosol and rosmarinic acid in bulk oil and oil-in-water emulsion. Journal of Agricultural and Food Chemistry. 1996;44(1):131–135. doi: 10.1021/jf950374p. [DOI] [Google Scholar]
- 65.Doolaege E. H. A., Raes K., De Vos F., Verhé R., De Smet S. Absorption, distribution and elimination of carnosic acid, a natural antioxidant from Rosmarinus officinalis, in rats. Plant Foods for Human Nutrition. 2011;66(2):196–202. doi: 10.1007/s11130-011-0233-5. [DOI] [PubMed] [Google Scholar]
- 66.Vaquero M. R., Villalba R. G., Larrosa M., et al. Bioavailability of the major bioactive diterpenoids in a rosemary extract: metabolic profile in the intestine, liver, plasma, and brain of Zucker rats. Molecular Nutrition & Food Research. 2013;57(10):1834–1846. doi: 10.1002/mnfr.201300052. [DOI] [PubMed] [Google Scholar]
- 67.Ferlemi A. V., Katsikoudi A., Kontogianni V. G., et al. Rosemary tea consumption results to anxiolytic- and anti-depressant-like behavior of adult male mice and inhibits all cerebral area and liver cholinesterase activity; phytochemical investigation and in silico studies. Chemico-Biological Interactions. 2015;237:47–57. doi: 10.1016/j.cbi.2015.04.013. [DOI] [PubMed] [Google Scholar]
- 68.MacHado D. G., Cunha M. P., Neis V. B., et al. Rosmarinus officinalis L. hydroalcoholic extract, similar to fluoxetine, reverses depressive-like behavior without altering learning deficit in olfactory bulbectomized mice. Journal of Ethnopharmacology. 2012;143(1):158–169. doi: 10.1016/j.jep.2012.06.017. [DOI] [PubMed] [Google Scholar]
- 69.Zhang D., Lee B., Nutter A., et al. Protection from cyanide-induced brain injury by the Nrf2 transcriptional activator carnosic acid. Journal of Neurochemistry. 2015;133(6):898–908. doi: 10.1111/jnc.13074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Kim S.-J., Kim J.-S., Cho H.-S., et al. Carnosol, a component of rosemary (Rosmarinus officinalis L.) protects nigral dopaminergic neuronal cells. NeuroReport. 2006;17(16):1729–1733. doi: 10.1097/01.wnr.0000239951.14954.10. [DOI] [PubMed] [Google Scholar]
- 71.Moss M., Oliver L. Plasma 1,8-cineole correlates with cognitive performance following exposure to rosemary essential oil aroma. Therapeutic Advances in Psychopharmacology. 2012;2(3):103–113. doi: 10.1177/2045125312436573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Ozarowski M., Mikolajczak P. L., Bogacz A., et al. Rosmarinus officinalis L. leaf extract improves memory impairment and affects acetylcholinesterase and butyrylcholinesterase activities in rat brain. Fitoterapia. 2013;91:261–271. doi: 10.1016/j.fitote.2013.09.012. [DOI] [PubMed] [Google Scholar]
- 73.Wijeratne S. S. K., Cuppett S. L. Potential of rosemary (Rosemarinus officinalis L.) diterpenes in preventing lipid hydroperoxide-mediated oxidative stress in Caco-2 cells. Journal of Agricultural and Food Chemistry. 2007;55(4):1193–1199. doi: 10.1021/jf063089m. [DOI] [PubMed] [Google Scholar]
- 74.Haraguchi H., Saito T., Okamura N., Yagi A. Inhibition of lipid peroxidation and superoxide generation by diterpenoids from Rosmarinus officinalis . Planta Medica. 1995;61(4):333–336. doi: 10.1055/s-2006-958094. [DOI] [PubMed] [Google Scholar]
- 75.Hou C.-W., Lin Y.-T., Chen Y.-L., et al. Neuroprotective effects of carnosic acid on neuronal cells under ischemic and hypoxic stress. Nutritional Neuroscience. 2012;15(6):257–263. doi: 10.1179/1476830512Y.0000000021. [DOI] [PubMed] [Google Scholar]
- 76.Satoh T., Izumi M., Inukai Y., et al. Carnosic acid protects neuronal HT22 Cells through activation of the antioxidant-responsive element in free carboxylic acid- and catechol hydroxyl moieties-dependent manners. Neuroscience Letters. 2008;434(3):260–265. doi: 10.1016/j.neulet.2008.01.079. [DOI] [PubMed] [Google Scholar]
- 77.Masuda T., Inaba Y., Takeda Y. Antioxidant mechanism of carnosic acid: structural identification of two oxidation products. Journal of Agricultural and Food Chemistry. 2001;49(11):5560–5565. doi: 10.1021/jf010693i. [DOI] [PubMed] [Google Scholar]
- 78.Habtemariam S. Investigation into the antioxidant and antidiabetic potential of Moringa stenopetala: identification of the active principles. Natural Product Communications. 2015;10(3):475–478. [PubMed] [Google Scholar]
- 79.Habtemariam S., Varghese G. K. Extractability of rutin in herbal tea preparations of Moringa stenopetala leaves. Beverages. 2015;1(3):169–182. doi: 10.3390/beverages1030169. [DOI] [Google Scholar]
- 80.Habtemariam S., Varghese G. K. The antidiabetic therapeutic potential of dietary polyphenols. Current Pharmaceutical Biotechnology. 2014;15(4):391–400. doi: 10.2174/1389201015666140617104643. [DOI] [PubMed] [Google Scholar]
- 81.Roselli M., Lentini G., Habtemariam S. Phytochemical, antioxidant and anti-α-glucosidase activity evaluations of Bergenia cordifolia . Phytotherapy Research. 2012;26(6):908–914. doi: 10.1002/ptr.3655. [DOI] [PubMed] [Google Scholar]
- 82.Habtemariam S. Methyl-3-O-methyl gallate and gallic acid from the leaves of Peltiphyllum peltatum: isolation and comparative antioxidant, prooxidant, and cytotoxic effects in neuronal cells. Journal of Medicinal Food. 2011;14(11):1412–1418. doi: 10.1089/jmf.2010.0257. [DOI] [PubMed] [Google Scholar]
- 83.Juan-Badaturuge M., Habtemariam S., Thomas M. J. K. Antioxidant compounds from a South Asian beverage and medicinal plant, Cassia auriculata . Food Chemistry. 2011;125(1):221–225. doi: 10.1016/j.foodchem.2010.08.065. [DOI] [Google Scholar]
- 84.Juan-Badaturuge M., Habtemariam S., Jackson C., Thomas M. J. K. Antioxidant principles of Tanacetum vulgare L. aerial parts. Natural Product Communications. 2009;4(11):1561–1564. [PubMed] [Google Scholar]
- 85.Habtemariam S., Dagne E. Comparative antioxidant, prooxidant and cytotoxic activity of sigmoidin A and eriodictyol. Planta Medica. 2010;76(6):589–594. doi: 10.1055/s-0029-1240604. [DOI] [PubMed] [Google Scholar]
- 86.Habtemariam S., Jackson C. Antioxidant and cytoprotective activity of leaves of Peltiphyllum peltatum (Torr.) Engl. Food Chemistry. 2007;105(2):498–503. doi: 10.1016/j.foodchem.2007.03.073. [DOI] [Google Scholar]
- 87.Habtemariam S. Flavonoids as inhibitors or enhancers of the cytotoxicity of tumor necrosis factor-α in L-929 tumor cells. Journal of Natural Products. 1997;60(8):775–778. doi: 10.1021/np960581z. [DOI] [PubMed] [Google Scholar]
- 90.Habtemariam S. Modulation of tumour necrosis factor-α-induced cytotoxicity by polyphenols. Phytotherapy Research. 1997;11(4):277–280. doi: 10.1002/(SICI)1099-1573(199706)11:4<277::AID-PTR90>3.0.CO;2-8. [DOI] [Google Scholar]
- 89.Habtemariam S. Catechols and quercetin reduce MTT through iron ions: a possible artefact in cell viability assays. Phytotherapy Research. 1995;9(8):603–605. doi: 10.1002/ptr.2650090816. [DOI] [Google Scholar]
- 90.Richheimer S. L., Bailey D. T., Bernart M. W., et al. Antioxidant activity and oxidative degradation of phenolic compounds isolated from rosemary. Recent Research Developments in Oil Chemistry. 1999;3:45–58. [Google Scholar]
- 91.Itoh K., Chiba T., Takahashi S., et al. An Nrf2/small Maf heterodimer mediates the induction of phase II detoxifying enzyme genes through antioxidant response elements. Biochemical and Biophysical Research Communications. 1997;236(2):313–322. doi: 10.1006/bbrc.1997.6943. [DOI] [PubMed] [Google Scholar]
- 92.Alam J., Stewart D., Touchard C., Boinapally S., Choi A. M. K., Cook J. L. Nrf2, a Cap‘n’Collar transcription factor, regulates induction of the heme oxygenase-1 gene. The Journal of Biological Chemistry. 1999;274(37):26071–26078. doi: 10.1074/jbc.274.37.26071. [DOI] [PubMed] [Google Scholar]
- 93.McMahon M., Itoh K., Yamamoto M., et al. The Cap'n'Collar basic leucine zipper transcription factor Nrf2 (NF-E2 p45-related factor 2) controls both constitutive and inducible expression of intestinal detoxification and glutathione biosynthetic enzymes. Cancer Research. 2001;61(8):3299–3307. [PubMed] [Google Scholar]
- 94.Nguyen T., Yang C. S., Pickett C. B. The pathways and molecular mechanisms regulating Nrf2 activation in response to chemical stress. Free Radical Biology and Medicine. 2004;37(4):433–441. doi: 10.1016/j.freeradbiomed.2004.04.033. [DOI] [PubMed] [Google Scholar]
- 95.Satoh T., Kosaka K., Itoh K., et al. Carnosic acid, a catechol-type electrophilic compound, protects neurons both in vitro and in vivo through activation of the Keap1/Nrf2 pathway via S-alkylation of targeted cysteines on Keap1. Journal of Neurochemistry. 2008;104(4):1116–1131. doi: 10.1111/j.1471-4159.2007.05039.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Itoh K., Tong K. I., Yamamoto M. Molecular mechanism activating Nrf2-Keap1 pathway in regulation of adaptive response to electrophiles. Free Radical Biology and Medicine. 2004;36(10):1208–1213. doi: 10.1016/j.freeradbiomed.2004.02.075. [DOI] [PubMed] [Google Scholar]
- 97.Satoh T., Lipton S. A. Redox regulation of neuronal survival mediated by electrophilic compounds. Trends in Neurosciences. 2007;30(1):37–45. doi: 10.1016/j.tins.2006.11.004. [DOI] [PubMed] [Google Scholar]
- 98.Satoh T., Okamoto S.-I., Cui J., et al. Activation of the Keap1/Nrf2 pathway for neuroprotection by electrophillic phase II inducers. Proceedings of the National Academy of Sciences of the United States of America. 2006;103(3):768–773. doi: 10.1073/pnas.0505723102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Satoh T., McKercher S. R., Lipton S. A. Nrf2/ARE-mediated antioxidant actions of pro-electrophilic drugs. Free Radical Biology and Medicine. 2013;65:645–657. doi: 10.1016/j.freeradbiomed.2013.07.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Martin D., Rojo A. I., Salinas M., et al. Regulation of heme oxygenase-1 expression through the phosphatidylinositol 3-kinase/Akt pathway and the Nrf2 transcription factor in response to the antioxidant phytochemical carnosol. The Journal of Biological Chemistry. 2004;279(10):8919–8929. doi: 10.1074/jbc.m309660200. [DOI] [PubMed] [Google Scholar]
- 101.Heiss E., Herhaus C., Klimo K., Bartsch H., Gerhauser C. Nuclear factor-kappa B is a molecular target for sulforaphane-mediated anti-inflammatory mechanisms. The Journal of Biological Chemistry. 2001;276(34):32008–32015. doi: 10.1074/jbc.M104794200. [DOI] [PubMed] [Google Scholar]
- 102.Rossi A., Kapahi P., Natoli G., et al. Anti-inflammatory cyclopentenone prostaglandins are direct inhibitors of IkappaB kinase. Nature. 2000;403(6765):103–108. doi: 10.1038/47520. [DOI] [PubMed] [Google Scholar]
- 103.Cernuda-Morollón E., Pineda-Molina E., Cañada F. J., Pérez-Sala D. 15-Deoxy-Δ12,14-prostaglandin J2 inhibition of NF-κB-DNA binding through covalent modification of the p50 subuni. The Journal of Biological Chemistry. 2001;276(38):35530–35536. doi: 10.1074/jbc.m104518200. [DOI] [PubMed] [Google Scholar]
- 104.Xiang Q., Wang Y., Wu W., et al. Carnosic acid protects against ROS/RNS-induced protein damage and upregulates HO-1 expression in RAW264.7 macrophages. Journal of Functional Foods. 2013;5(1):362–369. doi: 10.1016/j.jff.2012.11.007. [DOI] [Google Scholar]
- 105.Singletary K. W. Rosemary extract and carnosol stimulate rat liver glutathione-S-transferase and quinone reductase activities. Cancer Letters. 1996;100(1-2):139–144. doi: 10.1016/0304-3835(95)04082-x. [DOI] [PubMed] [Google Scholar]
- 106.Butterfield D. A., Drake J., Pocernich C., Castegna A. Evidence of oxidative damage in Alzheimer's disease brain: central role for amyloid β-peptide. Trends in Molecular Medicine. 2001;7(12):548–554. doi: 10.1016/s1471-4914(01)02173-6. [DOI] [PubMed] [Google Scholar]
- 107.Pereira C., Agostinho P., Moreira P. I., Cardoso S. M., Oliveira C. R. Alzheimer's disease-associated neurotoxic mechanisms and neuroprotective strategies. Current Drug Target—CNS & Neurological Disorders. 2005;4(4):383–403. doi: 10.2174/1568007054546117. [DOI] [PubMed] [Google Scholar]
- 108.Halliwell B. Oxidative stress and neurodegeneration: where are we now? Journal of Neurochemistry. 2006;97(6):1634–1658. doi: 10.1111/j.1471-4159.2006.03907.x. [DOI] [PubMed] [Google Scholar]
- 109.Dyall S. C., Michael-Titus A. T. Neurological benefits of omega-3 fatty acids. NeuroMolecular Medicine. 2008;10(4):219–235. doi: 10.1007/s12017-008-8036-z. [DOI] [PubMed] [Google Scholar]
- 110.Mailly F., Marin P., Israël M., Glowinski J., Prémont J. Increase in external glutamate and NMDA receptor activation contribute to H2O2-induced neuronal apoptosis. Journal of Neurochemistry. 1999;73(3):1181–1188. doi: 10.1046/j.1471-4159.1999.0731181.x. [DOI] [PubMed] [Google Scholar]
- 111.Asha Devi S. Aging brain: prevention of oxidative stress by vitamin E and exercise. TheScientificWorldJOURNAL. 2009;9:366–372. doi: 10.1100/tsw.2009.46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Murakami K., Murata N., Noda Y., et al. SOD1 (copper/zinc superoxide dismutase) deficiency drives amyloid beta protein oligomerization and memory loss in mouse model of Alzheimer disease. The Journal of Biological Chemistry. 2011;286(52):44557–44568. doi: 10.1074/jbc.m111.279208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Bermejo P., Martín-Aragón S., Benedí J., et al. Peripheral levels of glutathione and protein oxidation as markers in the development of Alzheimer's disease from mild cognitive impairment. Free Radical Research. 2008;42(2):162–170. doi: 10.1080/10715760701861373. [DOI] [PubMed] [Google Scholar]
- 114.Liu H., Wang H., Shenvi S., Hagen T. M., Liu R.-M. Glutathione metabolism during aging and in Alzheimer disease. Annals of the New York Academy of Sciences. 2004;1019:346–349. doi: 10.1196/annals.1297.059. [DOI] [PubMed] [Google Scholar]
- 115.Moreira P. I., Carvalho C., Zhu X., Smith M. A., Perry G. Mitochondrial dysfunction is a trigger of Alzheimer's disease pathophysiology. Biochimicaa et Biophysica Acta (BBA)—Molecular Basis of Disease. 2010;1802(1):2–10. doi: 10.1016/j.bbadis.2009.10.006. [DOI] [PubMed] [Google Scholar]
- 116.Dong J., Atwood C. S., Anderson V. E., et al. Metal binding and oxidation of amyloid-β within isolated senile plaque cores: raman microscopic evidence. Biochemistry. 2003;42(10):2768–2773. doi: 10.1021/bi0272151. [DOI] [PubMed] [Google Scholar]
- 117.Grundke-Iqbal I., Fleming J., Tung Y.-C., Lassmann H., Iqbal K., Joshi J. G. Ferritin is a component of the neuritic (senile) plaque in Alzheimer dementia. Acta Neuropathologica. 1990;81(2):105–110. doi: 10.1007/bf00334497. [DOI] [PubMed] [Google Scholar]
- 118.Lovell M. A., Robertson J. D., Teesdale W. J., Campbell J. L., Markesbery W. R. Copper, iron and zinc in Alzheimer's disease senile plaques. Journal of the Neurological Sciences. 1998;158(1):47–52. doi: 10.1016/s0022-510x(98)00092-6. [DOI] [PubMed] [Google Scholar]
- 119.Tamano H., Takeda A. Dynamic action of neurometals at the synapse. Metallomics. 2011;3(7):656–661. doi: 10.1039/c1mt00008j. [DOI] [PubMed] [Google Scholar]
- 120.Hui-Hui Z., Peng-Fei T., Kan Z., Hui W., Bao-Huai W., Jing-Fen L. Antioxidant properties of phenolic diterpenes from Rosmarinus officinalis . Acta Pharmacologica Sinica. 2001;22(12):1094–1098. [PubMed] [Google Scholar]
- 121.Samman S., Sandström B., Toft M. B., et al. Green tea or rosemary extract added to foods reduces nonheme-iron absorption. The American Journal of Clinical Nutrition. 2001;73(3):607–612. doi: 10.1093/ajcn/73.3.607. [DOI] [PubMed] [Google Scholar]
- 122.Chen X.-L., Dodd G., Thomas S., et al. Activation of Nrf2/ARE pathway protects endothelial cells from oxidant injury and inhibits inflammatory gene expression. American Journal of Physiology—Heart and Circulatory Physiology. 2006;290(5):H1862–H1870. doi: 10.1152/ajpheart.00651.2005. [DOI] [PubMed] [Google Scholar]
- 123.Kapturczak M. H., Wasserfall C., Brusko T., et al. Heme oxygenase-1 modulates early inflammatory responses: evidence from the heme oxygenase-1-deficient mouse. The American Journal of Pathology. 2004;165(3):1045–1053. doi: 10.1016/s0002-9440(10)63365-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Lian K.-C., Chuang J.-J., Hsieh C.-W., et al. Dual mechanisms of NF-κB inhibition in carnosol-treated endothelial cells. Toxicology and Applied Pharmacology. 2010;245(1):21–35. doi: 10.1016/j.taap.2010.01.003. [DOI] [PubMed] [Google Scholar]
- 125.Yao H., Chen Y., Zhang L., et al. Carnosol inhibits cell adhesion molecules and chemokine expression by tumor necrosis factor-α in human umbilical vein endothelial cells through the nuclear factor-κB and mitogen-activated protein kinase pathways. Molecular Medicine Reports. 2014;9(2):476–480. doi: 10.3892/mmr.2013.1839. [DOI] [PubMed] [Google Scholar]
- 126.Foresti R., Bains S. K., Pitchumony T. S., et al. Small molecule activators of the Nrf2-HO-1 antioxidant axis modulate heme metabolism and inflammation in BV2 microglia cells. Pharmacological Research. 2013;76:132–148. doi: 10.1016/j.phrs.2013.07.010. [DOI] [PubMed] [Google Scholar]
- 127.Yu Y.-M., Lin C.-H., Chan H.-C., Tsai H.-D. Carnosic acid reduces cytokine-induced adhesion molecules expression and monocyte adhesion to endothelial cells. European Journal of Nutrition. 2009;48(2):101–106. doi: 10.1007/s00394-008-0768-x. [DOI] [PubMed] [Google Scholar]
- 128.Xiang Q., Liu Z., Wang Y., et al. Carnosic acid attenuates lipopolysaccharide-induced liver injury in rats via fortifying cellular antioxidant defense system. Food and Chemical Toxicology. 2013;53:1–9. doi: 10.1016/j.fct.2012.11.001. [DOI] [PubMed] [Google Scholar]
- 129.Poeckel D., Greiner C., Verhoff M., et al. Carnosic acid and carnosol potently inhibit human 5-lipoxygenase and suppress pro-inflammatory responses of stimulated human polymorphonuclear leukocytes. Biochemical Pharmacology. 2008;76(1):91–97. doi: 10.1016/j.bcp.2008.04.013. [DOI] [PubMed] [Google Scholar]
- 130.Lo A.-H., Liang Y.-C., Lin-Shiau S.-Y., Ho C.-T., Lin J.-K. Carnosol, an antioxidant in rosemary, suppresses inducible nitric oxide synthase through down-regulating nuclear factor-κB in mouse macrophages. Carcinogenesis. 2002;23(6):983–991. doi: 10.1093/carcin/23.6.983. [DOI] [PubMed] [Google Scholar]
- 131.Yanagitai M., Itoh S., Kitagawa T., Takenouchi T., Kitani H., Satoh T. Carnosic acid, a pro-electrophilic compound, inhibits LPS-induced activation of microglia. Biochemical and Biophysical Research Communications. 2012;418(1):22–26. doi: 10.1016/j.bbrc.2011.12.087. [DOI] [PubMed] [Google Scholar]
- 132.Prokop S., Miller K. R., Heppner F. L. Microglia actions in Alzheimer's disease. Acta Neuropathologica. 2013;126(4):461–477. doi: 10.1007/s00401-013-1182-x. [DOI] [PubMed] [Google Scholar]
- 133.Verri M., Pastoris O., Dossena M., et al. Mitochondrial alterations, oxidative stress and neuroinflammation in Alzheimer's disease. International Journal of Immunopathology and Pharmacology. 2012;25(2):345–353. doi: 10.1177/039463201202500204. [DOI] [PubMed] [Google Scholar]
- 134.Ito D., Imai Y., Ohsawa K., Nakajima K., Fukuuchi Y., Kohsaka S. Microglia-specific localisation of a novel calcium binding protein, Iba1. Molecular Brain Research. 1998;57(1):1–9. doi: 10.1016/S0169-328X(98)00040-0. [DOI] [PubMed] [Google Scholar]
- 135.Kuhlmann A., Röhl C. Phenolic antioxidant compounds produced by in vitro. Cultures of Rosemary (Rosmarinus officinalis.) and their anti-inflammatory effect on lipopolysaccharide-activated microglia. Pharmaceutical Biology. 2006;44(6):401–410. doi: 10.1080/13880200600794063. [DOI] [Google Scholar]
- 136.Braak H., de Vos R. A. I., Jansen E. N. H., Bratzke H., Braak E. Neuropathological hallmarks of Alzheimer's and Parkinson's diseases. Progress in Brain Research. 1998;117:267–285. doi: 10.1016/s0079-6123(08)64021-2. [DOI] [PubMed] [Google Scholar]
- 137.Armstrong R. A. The molecular biology of senile plaques and neurofibrillary tangles in Alzheimer's disease. Folia Neuropathologica. 2009;47(4):289–299. [PubMed] [Google Scholar]
- 138.Serrano-Pozo A., Frosch M. P., Masliah E., Hyman B. T. Neuropathological alterations in Alzheimer disease. Cold Spring Harbor Perspectives in Medicine. 2011;1(1) doi: 10.1101/cshperspect.a006189.a006189 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Portelius E., Mattsson N., Andreasson U., Blennow K., Zetterberg H. Novel abeta isoforms in Alzheimer's disease—their role in diagnosis and treatment. Current Pharmaceutical Design. 2011;17(25):2594–2602. doi: 10.2174/138161211797416039. [DOI] [PubMed] [Google Scholar]
- 140.Perneczky R., Alexopoulos P., Kurz A. Soluble amyloid precursor proteins and secretases as alzheimer's disease biomarkers. Trends in Molecular Medicine. 2014;20(1):8–15. doi: 10.1016/j.molmed.2013.10.001. [DOI] [PubMed] [Google Scholar]
- 141.Haass C. Take five—BACE and the γ-secretase quartet conduct Alzheimer's amyloid beta-peptide generation. The EMBO Journal. 2004;23(3):483–488. doi: 10.1038/sj.emboj.7600061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Guo T., Hobbs D. W. Development of BACE1 inhibitors for Alzheimer's disease. Current Medicinal Chemistry. 2006;13(15):1811–1829. doi: 10.2174/092986706777452489. [DOI] [PubMed] [Google Scholar]
- 143.Meng P., Yoshida H., Matsumiya T., et al. Carnosic acid suppresses the production of amyloid-β 1-42 by inducing the metalloprotease gene TACE/ADAM17 in SH-SY5Y human neuroblastoma cells. Neuroscience Research. 2013;75(2):94–102. doi: 10.1016/j.neures.2012.11.007. [DOI] [PubMed] [Google Scholar]
- 144.Yoshida H., Meng P., Matsumiya T., et al. Carnosic acid suppresses the production of amyloid-β 1-42 and 1-43 by inducing an α-secretase TACE/ADAM17 in U373MG human astrocytoma cells. Neuroscience Research. 2014;79(1):83–93. doi: 10.1016/j.neures.2013.11.004. [DOI] [PubMed] [Google Scholar]
- 145.Dahlgren K. N., Manelli A. M., Blaine Stine W., Jr., Baker L. K., Krafft G. A., Ladu M. J. Oligomeric and fibrillar species of amyloid-β peptides differentially affect neuronal viability. The Journal of Biological Chemistry. 2002;277(35):32046–32053. doi: 10.1074/jbc.m201750200. [DOI] [PubMed] [Google Scholar]
- 146.Hartley D. M., Walsh D. M., Ye C. P., et al. Protofibrillar intermediates of amyloid β-protein induce acute electrophysiological changes and progressive neurotoxicity in cortical neurons. The Journal of Neuroscience. 1999;19(20):8876–8884. doi: 10.1523/JNEUROSCI.19-20-08876.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Hoshi M., Sato M., Matsumoto S., et al. Spherical aggregates of β-amyloid (amylospheroid) show high neurotoxicity and activate tau protein kinase I/glycogen synthase kinase-3β . Proceedings of the National Academy of Sciences of the United States of America. 2003;100(11):6370–6375. doi: 10.1073/pnas.1237107100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Kayed R., Head E., Thompson J. L., et al. Common structure of soluble amyloid oligomers implies common mechanism of pathogenesis. Science. 2003;300(5618):486–489. doi: 10.1126/science.1079469. [DOI] [PubMed] [Google Scholar]
- 149.Lambert M. P., Barlow A. K., Chromy B. A., et al. Diffusible, nonfibrillar ligands derived from Aβ 1–42 are potent central nervous system neurotoxins. Proceedings of the National Academy of Sciences of the United States of America. 1998;95(11):6448–6453. doi: 10.1073/pnas.95.11.6448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Ishii K., Klunk W. E., Arawaka S., et al. Chrysamine G and its derivative reduce amyloid β-induced neurotoxicity in mice. Neuroscience Letters. 2002;333(1):5–8. doi: 10.1016/s0304-3940(02)00915-1. [DOI] [PubMed] [Google Scholar]
- 151.Adessi C., Frossard M.-J., Boissard C., et al. Pharmacological profiles of peptide drug candidates for the treatment of Alzheimer's disease. The Journal of Biological Chemistry. 2003;278(16):13905–13911. doi: 10.1074/jbc.m211976200. [DOI] [PubMed] [Google Scholar]
- 152.Gordon D. J., Tappe R., Meredith S. C. Design and characterization of a membrane permeable N-methyl amino acid-containing peptide that inhibits Aβ1-40 fibrillogenesis. Journal of Peptide Research. 2002;60(1):37–55. doi: 10.1034/j.1399-3011.2002.11002.x. [DOI] [PubMed] [Google Scholar]
- 153.Tjernberg L. O., Näslundt J., Lindqvist F., et al. Arrest of β-amyloid fibril formation by a pentapeptide ligand. Journal of Biological Chemistry. 1996;271(15):8545–8548. doi: 10.1074/jbc.271.15.8545. [DOI] [PubMed] [Google Scholar]
- 154.Tjernberg L. O., Lilliehöök C., Callaway D. J. E., et al. Controlling amyloid β-peptide fibril formation with protease-stable ligands. Journal of Biological Chemistry. 1997;272(19):12601–12605. doi: 10.1074/jbc.272.19.12601. [DOI] [PubMed] [Google Scholar]
- 155.Soto C., Sigurdsson E. M., Morelli L., Kumar R. A., Castaño E. M., Frangione B. β-Sheet breaker peptides inhibit fibrillogenesis in a rat brain model of amyloidosis: implications for Alzheimer's therapy. Nature Medicine. 1998;4(7):822–826. doi: 10.1038/nm0798-822. [DOI] [PubMed] [Google Scholar]
- 156.Ono K., Hasegawa K., Naiki H., Yamada M. Curcumin has potent anti-amiloidogenic effects for Alzheimer's β-amyloid fibrils in vitro . Journal of Neuroscience Research. 2004;75(6):742–750. doi: 10.1002/jnr.20025. [DOI] [PubMed] [Google Scholar]
- 157.Ono K., Hasegawa K., Naiki H., Yamada M. Anti-amyloidogenic activity of tannic acid and its activity to destabilize Alzheimer's β-amyloid fibrils in vitro. Biochimica et Biophysica Acta (BBA)—Molecular Basis of Disease. 2004;1690(3):193–202. doi: 10.1016/j.bbadis.2004.06.008. [DOI] [PubMed] [Google Scholar]
- 158.Ono K., Hasegawa K., Yoshiike Y., Takashima A., Yamada M., Naiki H. Nordihydroguaiaretic acid potently breaks down pre-formed Alzheimer's β-amyloid fibrils in vitro . Journal of Neurochemistry. 2002;81(3):434–440. doi: 10.1046/j.1471-4159.2002.00904.x. [DOI] [PubMed] [Google Scholar]
- 159.Tomiyama T., Asano S., Suwa Y., et al. Rifampicin prevents the aggregation and neurotoxicity of amyloid β protein in vitro . Biochemical and Biophysical Research Communications. 1994;204(1):76–83. doi: 10.1006/bbrc.1994.2428. [DOI] [PubMed] [Google Scholar]
- 160.Thomas T., Nadackal T. G., Thomas K. Aspirin and non-steroidal anti-inflammatory drugs inhibit amyloid-β aggregation. NeuroReport. 2001;12(15):3263–3267. doi: 10.1097/00001756-200110290-00024. [DOI] [PubMed] [Google Scholar]
- 161.Azad N., Rasoolijazi H., Joghataie M. T., Soleimani S. Neuroprotective effects of Carnosic acid in an experimental model of Alzheimer's disease in rats. Cell Journal. 2011;13(1):39–44. [PMC free article] [PubMed] [Google Scholar]
- 162.Rasoolijazi H., Azad N., Joghataei M. T., Kerdari M., Nikbakht F., Soleimani M. The protective role of carnosic acid against beta-amyloid toxicity in rats. The Scientific World Journal. 2013;2013:5. doi: 10.1155/2013/917082.917082 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Maynard C. J., Bush A. I., Masters C. L., Cappai R., Li Q.-X. Metals and amyloid-β in Alzheimer's disease. International Journal of Experimental Pathology. 2005;86(3):147–159. doi: 10.1111/j.0959-9673.2005.00434.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Duce J. A., Bush A. I., Adlard P. A. Role of Amyloid-β-metal Interactions in Alzheimer's Disease. Future Neurology. 2011;6(5):641–659. [Google Scholar]
- 165.Faller P., Hureau C., Berthoumieu O. Role of metal ions in the self-assembly of the Alzheimer's amyloid-β peptide. Inorganic Chemistry. 2013;52(21):12193–12206. doi: 10.1021/ic4003059. [DOI] [PubMed] [Google Scholar]
- 166.Hane F., Leonenko Z. Effect of metals on kinetic pathways of amyloid-β aggregation. Biomolecules. 2014;4(1):101–116. doi: 10.3390/biom4010101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Gouras G. K., Beal M. F. Metal chelator decreases Alzheimer β-amyloid plaques. Neuron. 2001;30(3):641–642. doi: 10.1016/s0896-6273(01)00330-0. [DOI] [PubMed] [Google Scholar]
- 168.Cuajungco M. P., Goldstein L. E., Nunomura A., et al. Evidence that the β-amyloid plaques of Alzheimer's disease represent the redox-silencing and entombment of Aβ by zinc. The Journal of Biological Chemistry. 2000;275(26):19439–19442. doi: 10.1074/jbc.c000165200. [DOI] [PubMed] [Google Scholar]
- 169.Tõugu V., Tiiman A., Palumaa P. Interactions of Zn(II) and Cu(II) ions with Alzheimer's amyloid-beta peptide. Metal ion binding, contribution to fibrillization and toxicity. Metallomics. 2011;3(3):250–261. doi: 10.1039/c0mt00073f. [DOI] [PubMed] [Google Scholar]
- 170.Ono K., Hamaguchi T., Naiki H., Yamada M. Anti-amyloidogenic effects of antioxidants: implications for the prevention and therapeutics of Alzheimer's disease. Biochimica et Biophysica Acta (BBA)—Molecular Basis of Disease. 2006;1762(6):575–586. doi: 10.1016/j.bbadis.2006.03.002. [DOI] [PubMed] [Google Scholar]
- 171.Bartus R. T., Dean R. L., III, Beer B., Lippa A. S. The cholinergic hypothesis of geriatric memory dysfunction. Science. 1982;217(4558):408–414. doi: 10.1126/science.7046051. [DOI] [PubMed] [Google Scholar]
- 172.Blennow K., de Leon M. J., Zetterberg H. Alzheimer's disease. The Lancet. 2006;368(9533):387–403. doi: 10.1016/s0140-6736(06)69113-7. [DOI] [PubMed] [Google Scholar]
- 173.Coyle J. T., Price D. L., DeLong M. R. Alzheimer's disease: a disorder of cortical cholinergic innervation. Science. 1983;219(4589):1184–1190. doi: 10.1126/science.6338589. [DOI] [PubMed] [Google Scholar]
- 174.Giacobini E. Cholinesterases: new roles in brain function and in Alzheimer's disease. Neurochemical Research. 2003;28(3-4):515–522. doi: 10.1023/a:1022869222652. [DOI] [PubMed] [Google Scholar]
- 175.Casey D. A., Antimisiaris D., O'Brien J. Drugs for Alzheimer's disease: are they effective? Pharmacy and Therpeutics. 2010;35(4):208–211. [PMC free article] [PubMed] [Google Scholar]
- 176.Szwajgier D. Anticholinesterase activity of phenolic acids and their derivatives. Zeitschrift für Naturforschung C. 2013;68(3-4):125–132. doi: 10.5560/znc.2013.68c0125. [DOI] [PubMed] [Google Scholar]
- 177.Merad M., Soufi W., Ghalem S., et al. Molecular interaction of acetylcholinesterase with carnosic acid derivatives: a neuroinformatics study. CNS & Neurological Disorders—Drug Targets. 2014;13(3):440–446. doi: 10.2174/18715273113126660157. [DOI] [PubMed] [Google Scholar]
- 178.Chen J.-H., Ou H.-P., Lin C.-Y., et al. Carnosic acid prevents 6-hydroxydopamine-induced cell death in SH-SY5Y cells via mediation of glutathione synthesis. Chemical Research in Toxicology. 2012;25(9):1893–1901. doi: 10.1021/tx300171u. [DOI] [PubMed] [Google Scholar]
- 179.Wu C.-R., Tsai C.-W., Chang S.-W., Lin C.-Y., Huang L.-C., Tsai C.-W. Carnosic acid protects against 6-hydroxydopamine-induced neurotoxicity in in vivo and in vitro model of Parkinson's disease: involvement of antioxidative enzymes induction. Chemico-Biological Interactions. 2015;225:40–46. doi: 10.1016/j.cbi.2014.11.011. [DOI] [PubMed] [Google Scholar]