

Abstract
Background
Doxorubicin (brand name: Adriamycin®) is used to treat solid tissue cancer but it also affects noncancerous tissues. Its mechanism of cytotoxicity is probably related to increased oxidation, mitochondrial dysfunction, and apoptosis. Melatonin is reported to have antiapoptotic and antioxidative effects. The aim of this study was to determine whether melatonin would counteract in vitro cytotoxicity of doxorubicin in mouse fibroblasts and determine the pathway of its action against doxorubicin-induced apoptosis.
Material/Methods
We measured markers of apoptosis (cytochrome-c, mitochondrial membrane potential, and apoptotic cell number) and oxidative stress (total oxidant and antioxidant status) and calculated oxidant stress index in 4 groups of fibroblasts: controls, melatonin-treated, doxorubicin-treated, and fibroblasts concomittantly treated with a combination of melatonin and doxorubicin.
Results
Melatonin given with doxorubicin succesfully countered apoptosis generated by doxorubicin alone, which points to its potential as a protective agent against cell death in doxorubicin chemotherapy. This also implies that patients should be receiving doxorubicin treatment when their physiological level of melatonin is at its highest, which is early in the morning.
Conclusions
This physiological level may not be high enough to overcome doxorubicin-induced oxidative stress, but adjuvant melatonin treatment may improve quality of life. Further research is needed to verify our findings.
MeSH Keywords: Apoptosis; Doxorubicin; Melatonin; Membrane Potential, Mitochondrial
Background
Doxorubicin (DOX) is an efficient anthracycline group chemotherapy drug used to treat hematologic and solid tumors [1], as it induces oxidative stress, inhibits topoisomerase II, and eventually leads to cell death, mainly by apoptosis [1–5]. It also inhibits cell propagation [2]. DOX causes alteration of mitochondrial function and structure due to DOX accumulation in mitochondria [6–10]. It has been suggested that the undesired effect of DOX is related to increase oxidative stress in the cell, triggering apoptosis [9,11,12].
There are strategies to attenuate the toxic effects of doxorubicin, such as dose regulation or combined therapy with antioxidants [13]. However, no effective combination has been found so far. One of the antiapoptotic agents with some promise is melatonin. Its antiapoptotic action has been evidenced in the brain, kidney, pancreas, and liver [14], but findings to the contrary have also been reported in vitro [15]). Because it is a small lipophilic molecule, it is distributed throughout the cell, with significant deposition in the mitochondria, where it can attenuate ischemia-reperfusion and indomethacin-induced damage [16]. It can also modulate mitochondrial function and homeostasis. Aziriova [17] reported that melatonin even reversed anxiety induced by doxorubicin in rats. One of its unique properties is that its metabolites also have antioxidative effects, even at low concentrations [18]. Furthermore, melatonin has low toxicity, which implies that it may be used safely in patient therapy [19].
Several studies have shown that melatonin can counteract doxorubicin-induced cytotoxicity [18,19] but they have not clarified the mechanism of its action. Therefore, with our study we wanted to determine if the mechanism of melatonin action against doxorubicin cytotoxicity was via its antioxidative and/or antiapoptotic pathway. We tested it in the mouse embryonic fibroblast cell line (NIH 3T3) because fibroblasts, which compose 70% of the heart as nanocardiomyocytes [20], are very responsive to chemical, electric, and mechanical signals, and changes made to these signals can affect other cardiac cells [21].
Material and Methods
Experimental design of study
Four groups of NIH3T3 cells were created: the control (Con) group received DMEM alone; the melatonin (MEL) group received melatonin in the dose of 1 μM for 24 h; the doxorubicin (DOX) group received doxorubicin hydrochloride (Adriblastina vial 10 mg, Pharmacia) in the dose of 2.6 μM for 24 h, and the MEL+DOX group received a combination of the doses corresponding to the MEL and DOX alone group. The dose of melatonin may be the key to achieving antiapoptotic effects. At high concentrations, melatonin activates the mitochondrial pathway by affecting the Bax/Bcl protein balance and the expressions of caspase-9 and caspase-3 in pancreatic carcinoma cell line [22]. At low concentrations, however, HSP27, HSP70, and HSP90 induction prevent the activation of caspase-3 [22]. One study reported that melatonin might inhibit cell proliferation at physiological (nanomolar and lower) and pharmacological concentrations (10 μM) but not at concentrations in between [23]. We therefore opted for 1 μM, which is much higher than the physiological (nanomolar) plasma concentrations. Furthermore, we opted to test melatonin effects when given with doxorubicin at the same time, thus extending an earlier study by Oz et al. [24], who reported that melatonin was less effective against doxorubicin when given as pretreatment than when given after doxorubicin.
To determine the effects of these treatments on oxidation and apoptosis, we made the measurements described below.
Cell culture
NIH3T3 cells were grown in poly-L-lysine-coated flasks (~2.0–2.5×106 cells/mL) in DMEM medium supplemented with 10% heat-inactivated fetal bovine serum in a humidified incubator with 5% CO2/95% air mixture at 37°C.
Cell viability
NIH3T3 cells were seeded into 24-well culture plates at 1×106 cells/well. Viability was assessed by trypan blue exclusion using a Vi-Cell XR cell viability analyzer (Beckman Coulter, Brea, CA), which is an automated hemocytometer. Basically, cell samples were mixed 1:1 with trypan blue, and 50 images were taken for calculation of cell number.
Total oxidant status
Total oxidant status (TOS) was measured with a BioTek μQuant microplate spectrophotometer using a Rel Assay kit (Cat No: RL0024; Gaziantep, Turkey) according to the manufacturer’s instructions. The results are expressed as a μmol H2O2 Equiv./L.
Total antioxidant status
Total antioxidant status (TAS) was measured with a BioTek μQuant microplate spectrophotometer using a Rel Assay kit (Cat No: RL0017; Gaziantep, Turkey) according to the manufacturer’s instructions. The results are expressed as mmol Trolox Equiv./L.
Oxidative stress index
Oxidative stress index (OSI) is the ratio between TOS and TAS levels measured in 6 samples per group.
Protein levels
The mechanism of apoptosis in fibroblasts was established by determining protein levels. To prepare cell lysates, NIH3T3 cells were centrifuged at 3000 rpm for 8 min, washed several times with PBS, resuspended in RIPA buffer (Cat. No. sc-24948; Santa Cruz Biotechnology), and incubated on ice for 10 min. The lysates were stored at −20°C until used. Protein levels were measured using a Qubit fluorometer and quantitation assays: 100 μg protein sample was mixed with 4×SDS loading buffer and then separated on a 4–12% Bis-Tris gel. After running, proteins were transferred to a PVDF membrane. We used primary antibodies against the following proteins: AIF, β-actin, ERK ½, SAPK/JNK (Cell Signaling), PPAR-γ, AMPK, and cytochrome-c (Abcam, Cambridge, UK). Then we used alkaline phosphatase-conjugated secondary antibodies (anti-mouse and anti-rabbit 1/3000). Signals were detected using the NCIP/BNP kit (Cat. No. ES0006; Millipore, Boston, MA, USA).
F-actin filaments
We looked for actin filaments because doxorubicin has been reported to inhibit cardiac genes, including α-actin, troponin, and myosin light chain-2 [25,26]. NIH 3T3 fibroblasts grown on glass coverslips (~80 000 cells) were left unstimulated (control) or stimulated with 1 μM melatonin, 2.6 μM doxorubicin, and melatonin with doxorubicin. They were washed with PBS and fixed in 5% paraformaldehyde at room temperature for 60 min. Fixed cells were permeabilized in 0.1% (vol/vol) Triton X-100 in PBS at room temperature for 10 min and blocked in 5% BSA in PBS at room temperature for 60 min. Actin filaments were visualized using Alexa fluor 595 phalloidin, and nuclei with 4″,6-diamidino-2-phenylindole dichloride (DAPI) (Invitro Molecular Probes, OR, USA). All images were obtained on an Olympus BX51 microscope equipped with a DP72 camera controlled by Olympus DP2-TWAIN software.
Mitochondrial membrane potential measurement
Mitochondria are targets of doxorubicin, and play an important role in initiating apoptosis. NIH 3T3 cells (~80 000) were harvested and allowed to attach on coverslips over the night. Then the cells were exposed to melatonin, DOX, and melatonin with DOX for 24 h. Mitochondrial membrane potential (MMP) was measured using a Sigma-Aldrich kit, Cat. No. CS0390 according the manufacturer’s instructions. Briefly, the cells were incubated with JC-1 dye with a 5% CO2/95% O2 air mixture at 37°C for 20 min. JC-1 dye is specific for the mitochondrial membrane. Healthy mitochondria have a high electrochemical potential, and JC-1 dye crosses the membrane to form red aggregates on fluorescence. Unhealthy mitochondria with low potential prevent JC-1 dye from accumulating, and fluorescence monomers are green.
After incubation the cells were rinsed with staining buffer. The coverslips were directly observed under a fluorescence microscope (Olympus BX51 Microscope equipped with a DP72 camera, controlled by Olympus DP2-TWAIN software).
Apoptotic cell count
The cells were cultured in 6-well plates with coverslips overnight. After they were divided into groups, the coverslips were washed twice with PBS and fixed in 4% paraformaldehyde solution for 1 h at room temperature. The coverslips were then rinsed with PBS, incubated in permeabilization solution (0.1% Triton-X-100 in 0.1% sodium citrate, freshly prepared), and the apoptotic cells labelled with a terminal deoxynucleotidyl-transferase-mediated dUTP nick end-labeling (TUNEL) kit from Roche (Cat. No. 1684795; Mannheim, Germany) following the manufacturer’s instructions. All images were obtained on an Olympus BX51 microscope equipped with a DP72 camera and controlled with Olympus DP2-TWAIN software. The apoptotic (TUNEL-positive) cells were quantified using the automated cell counting in ImageJ with ITCN plugin (Automatic Nuclei Counter plugin) (NIH). Summarly, images were converted to a binary image for 8-bit. Dark areas represent cells. Then, ITCN was used for cell counter for 15 images for every groups, and the total areas of groups were the same.
Statistical analysis
For statistical analysis we used Excel and the SPSS version 16.0. All results are expressed as mean ±SEM. Normal distribution was tested using Levene’s test. To compare the results between the groups, we used multiple analyses of variance (ANOVA), followed by a post hoc protected Tukey test. p<0.05 was considered to be significant for all data.
Results
Cell viability
Cell viability is expressed as the percentage of control cell viability (taken to be 100%). We found no significant differences between the groups (Figure 1).
Figure 1.
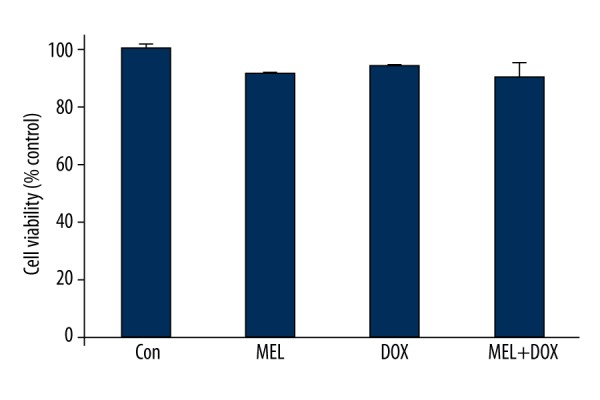
Cell viability across the groups. Con – control group; MEL – melatonin group; DOX – doxorubicin group; MEL+DOX – melatonin plus doxorubicin group. All data are expressed as mean ±SEM.
The effect of melatonin on oxidative stress induced by doxorubicin
Figure 2 shows the effects of melatonin on oxidative stress in fibroblast cells across the groups. Even though it lowered TOS in the combination group (Mel+DOX) in respect to the DOX group (or other groups for that matter), the differences in TOS and OSI were not statistically significant. MEL groups of OSI, TOS, and TAS levels were not different from Con.
Figure 2.
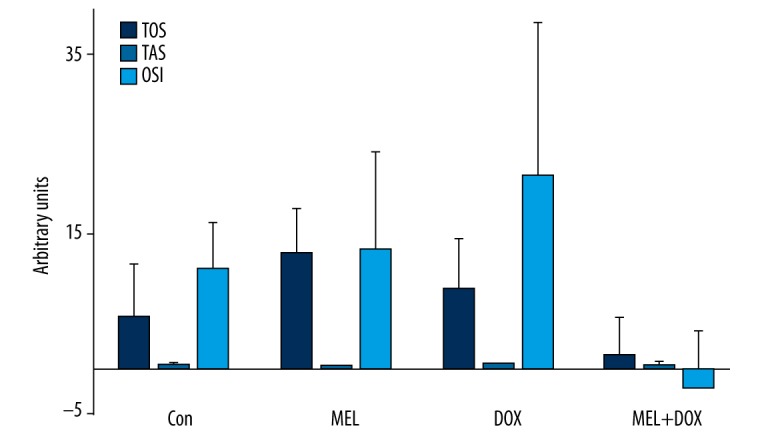
The effect of melatonin on oxidative stress in fibroblast cells treated with doxorubicin. There were no significant differences in TOS and OSI between treatment groups in Figure 2. TOS – total oxidant status; TAS – total antioxidant status, OSI – oxidative stress index. Con – control group; MEL – melatonin group; DOX – doxorubicin group; MEL+DOX – melatonin plus doxorubicin group. All data are expressed as mean ±SEM.
The effect of melatonin on doxorubicin-induced dissipation of mitochondrial membrane potential
The control and MEL group had normal healthy mitochondria, represented by red fluorescence (Figure 3A and B). Although DOX dissipated the MMP, resulting in green fluorescence (Figure 3C), MEL given with DOX restored the MMP (Figure 3D).
Figure 3.

The effect of melatonin on mitochondrial membrane potential in fibroblast cells treated with doxorubicin. Red fluorescence represents healthy mitochondria having a high electrochemical potential and green fluorescence represents unhealthy mitochondria having a low electrochemical potential (100× magnification). Con – control group (A); MEL – melatonin group (B); DOX – doxorubicin group (C); MEL+DOX – melatonin plus doxorubicin group (D).
The effect of melatonin on doxorubicin-induced changes in protein levels
Cytochrome-c levels
Cytochrome-c regulates the apoptosis-dependent mitochondrial pathway. DOX increased its level but melatonin in the MEL+DOX group restored it to lower levels (Figure 4A).
Figure 4.
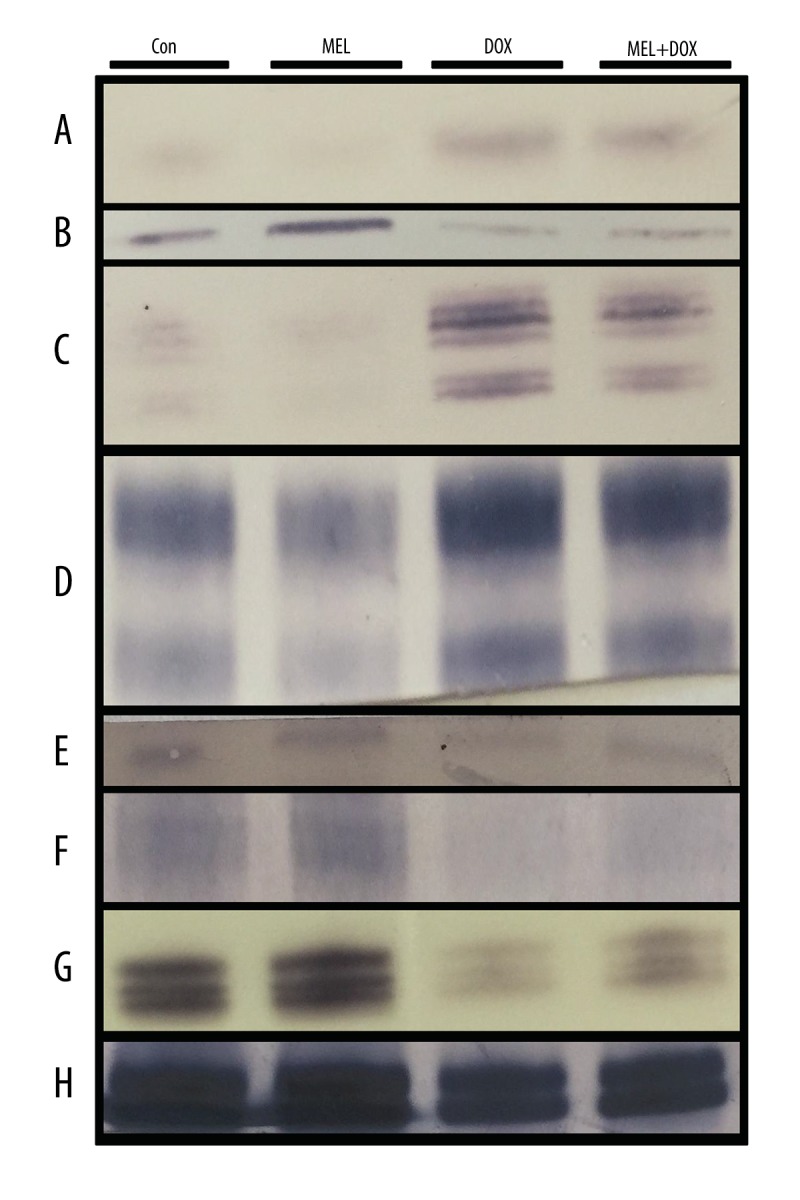
The effect of melatonin on protein affected by oxidative stress in fibroblast cells treated with doxorubicin. Con – control group; MEL – melatonin group; DOX – doxorubicin group; MEL+DOX – melatonin plus doxorubicin group. (A) Cytochrome-c; (B) AIF – apoptosis inducing factor; (C) ERK1/2 – extracellular signal-regulated kinases; (D) SAPK/JNK – stress-activated kinases; (E) PPAR-γ – peroxisome proliferator-activated receptor gamma; (F) AMPK – AMP-activated protein kinase; (G) PCNA – proliferating cell nuclear antigen; (H) β-actin.
Apoptosis-inducing factor levels
Apoptosis-inducing factor (AIF) plays a crucial role in caspase-independent apoptosis. It is located in the inner membrane of mitochondria. DOX caused its release into the cytosol (Figure 4), but MEL counteracted its release (Figure 4B).
Extracellular signal-regulated kinase expression
ERK 1 and 2 participate in the regulation of cell adhesion, cell migration, cell survival, and differentiation [27]. The DOX-treated cells showed a very low expression of both ERK 1 and 2. Co-treatment with MEL recovered their expressions (Figure 4C).
Stress-activated protein kinases levels
SAPK participates in cellular stress response, including differentiation and proliferation [28]. DOX lowered its level while co-treatment with MEL restored it (Figure 4D).
Peroxisome proliferator-activated receptor gamma protein levels
Peroxisome proliferator-activated receptor gamma (PPAR-γ) plays a crucial role in fatty acid and glucose metabolism regulation. DOX lowered PPAR-γ levels, but MEL+DOX attenuated its effect (Figure 4E).
AMP-activated protein kinase levels
AMPK maintains cell energy homeostasis by balancing ATP production when energy changes. DOX lowered the AMPK level, but it was improved in the MEL+DOX group (Figure 4F).
Proliferating cell nuclear antigen level
Proliferating cell nuclear antigen (PCNA) assists in DNA replication and is a common marker of proliferation. As expected, co-treatment with MEL countered the PCNA-lowering effect of DOX (Figure 4G).
The effect of melatonin on doxorubicin-modulated distribution of F-actin
Figure 5 shows stained F-actin (red) and nucleus (blue) in NIH 3T3. DOX increased the formation of filopodia (Figure 5C), but co-treatment with MEL counteracted the stress, restoring F-actin to normal (Figure 5D). Only melatonin treatment also increased filopodia formation in the cells (Figure 5B).
Figure 5.

The effect of melatonin on F-actin distribution in fibroblasts treated with doxorubicin (100x magnification). Con – control group (A); MEL – melatonin group (B); DOX – doxorubicin group (C); MEL+DOX – melatonin plus doxorubicin group (D).
The effect of melatonin on doxorubicin-induced apoptosis
DOX significantly increased the number of apoptotic cells to 123±37 vs. control (23±6), whereas co-treatment with MEL decreased it to 30±6. Only MEL treated cells were not altered to apoptotic cells (22±5; Figures 6, 7).
Figure 6.

Apoptosis in fibroblasts treated with melatonin and doxorubicin (100× magnification). Green fluorescence represents healthier cells than red fluorescence (100× magnification). Con – control group (A); MEL – melatonin group (B); DOX – doxorubicin group (C); MEL+DOX – melatonin plus doxorubicin group (D).
Figure 7.
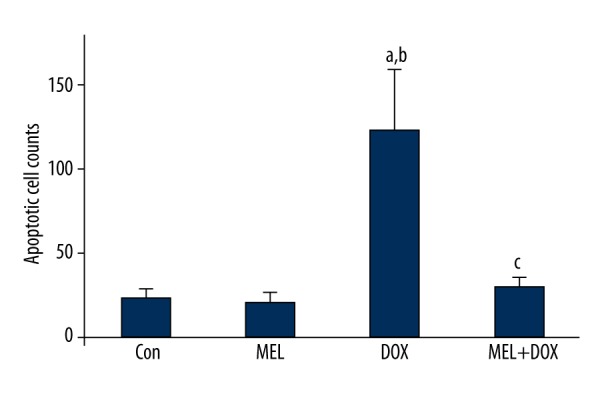
Apoptotic cell counts in fibroblasts treated with melatonin and doxorubicin. Con – control group; MEL – melatonin group; DOX – doxorubicin group; MEL+DOX – melatonin plus doxorubicin group. a p<0.05 vs. Con, b p<0.05 vs. MEL, c p<0.05 vs. DOX. All data are expressed as mean ±SEM.
Discussion
Our results have confirmed our hypothesis that melatonin would counteract the adverse effects of doxorubicin: it preserved mitochondrial membrane potential, and restored cytochrome-c and AIF levels as mechanisms of apoptosis and, therefore, decreased apoptotic cell count.
For the antioxidative action, our dosage was less efficient. An earlier study reported that oxidative stress induced by doxorubicin was higher in rats exposed to 24-h light than in rats kept in 24-h dark [29]. This finding suggests that melatonin could reverse oxidative stress induced by doxorubicin, even at physiological levels. However, our melatonin dose of 1 μM did not have a significant effect on OSI and has confirmed the findings by Santofimia-Castaño et al. [30]. Instead, it showed a dominant antiapoptotic effect. A previous study indicated that MEL cotreated with DOX in rats with hepatoma did not decrease the anticancer effect of DOX, and even decreased the effect of DOX on noncancerous tissue (e.g., heart) by decreasing apoptotic cell numbers [31].
In our study, melatonin restored MMP reduced by doxorubicin. This result is consistent with earlier reports [32,33]. Melatonin stimulates the biogenesis of mitochondria and upregulates PPAR-gamma. It has been reported to enhance mitochondrial function and quantity [34]. In our study, doxorubicin lowered the PPAR-gamma protein level, which plays an important role in regulating mitochondrial energy production [35]. Wang et al. [36] reported that one of the PPAR-gamma agonists attenuated apoptosis by inhibiting NF-κB and MAPK P38 signaling pathways. A similar antiapoptotic mechanism through PPAR-gamma was reported by Mahmoud-Awny et al. [37].
Currently, however, less is known about the modulation of apoptosis by melatonin in fibroblasts exposed to doxorubicin. In our study, melatonin given with doxorubicin increased the PPAR-gamma and AMPK levels, which suggests that there is a link between these 2 proteins [38]. Also, AMPK is related to circadian rythm [39]. To the best of our knowledge, our study is the first to suggest a trimodal relationship between AMPK-PPAR-gamma and melatonin. Furthermore, we found a positive interaction between AMPK and ERK1/2. An earlier study reported that 1 μM of melatonin elevated ERK1/2 and SAP/JNK levels [30]. The observed changes could improve PCNA levels and cell proliferation of fibroblasts. All this suggests that melatonin counters doxorubicin’s cytotoxicity by increasing cell proliferation and inhibiting apoptosis. The AMPK-PPARγ pathway might be critical in the regulation of apoptosis induced by doxorubicin.
Apoptosis is one of the most important pathways of doxorubicin toxicity. This is why we felt it was worth seeking the exact mechanism of apoptosis induced by doxorubicin, as finding this mechanism could lead to new cardioprotective chemotherapy strategies.
However, our study has its limitations. First of all, the melatonin concentration we used is higher than the physiological level [34]. The other limitation is related to this study being in vitro, as we cannot tell how melatonin will behave in living organisms until in vivo studies are performed. Lastly, the protein level of melatonin’s receptors was not evaluated. We hope that future studies will overcome these limitations and investigate the relationship between melatonin and AMPK-PPARγ in doxorubicin cytotoxicity even more closely.
Conclusions
Our study has provided evidence that melatonin has potential as an adjuvant in doxorubicin chemotherapy, which could attenuate its adverse effects without reducing its effectiveness in cancer cells. If proved efficient, this could result in new cardioprotective strategies that may involve preserving the patient’s circadian rhythm. Future in vivo studies will be necessary to test the therapeutic value of melatonin administration with doxorubicin to determine how it behaves in living organisms.
Footnotes
Source of support: This work was supported by self-financing
References
- 1.Coldwell K, Cutts SM, Ognibene TJ, et al. Detection of adriamycin-DNA adducts by accelerator mass spectrometry. Methods Mol Biol. 2010;613:103–18. doi: 10.1007/978-1-60327-418-0_7. [DOI] [PubMed] [Google Scholar]
- 2.Sauter KA, Magun EA, Iordanov MS, Magun BE. ZAK is required for doxorubicin, a novel ribotoxic stressor, to induce SAPK activation and apoptosis in HaCaT cells. Cancer Biol Ther. 2010;10(3):258–66. doi: 10.4161/cbt.10.3.12367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Arif IS, Hooper CL, Greco F, et al. Increasing doxorubicin activity against breast cancer cells using PPARgamma-ligands and by exploiting circadian rhythms. Br J Pharmacol. 2013;169(5):1178–88. doi: 10.1111/bph.12202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ta HQ, Thomas KS, Schrecengost RS, Bouton AH. A novel association between p130Cas and resistance to the chemotherapeutic drug adriamycin in human breast cancer cells. Cancer Res. 2008;68(21):8796–804. doi: 10.1158/0008-5472.CAN-08-2426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Cao B, Li M, Zha W, et al. Metabolomic approach to evaluating adriamycin pharmacodynamics and resistance in breast cancer cells. Metabolomics. 2013;9(5):960–73. doi: 10.1007/s11306-013-0517-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Dursun N, Taskin E, Yerer Aycan MB, Sahin L. Selenium-mediated cardioprotection against adriamycin-induced mitochondrial damage. Drug Chem Toxicol. 2011;34(2):199–207. doi: 10.3109/01480545.2010.538693. [DOI] [PubMed] [Google Scholar]
- 7.Ozdogan K, Taskin E, Dursun N. Protective effect of carnosine on adriamycin-induced oxidative heart damage in rats. Anadolu Kardiyol Derg. 2011;11(1):3–10. doi: 10.5152/akd.2011.003. [DOI] [PubMed] [Google Scholar]
- 8.Taskin E, Dursun N. The protection of selenium on adriamycin-induced mitochondrial damage in rat. Biol Trace Elem Res. 2012;147(1–3):165–71. doi: 10.1007/s12011-011-9273-9. [DOI] [PubMed] [Google Scholar]
- 9.Taskin E, Kindap EK, Ozdogan K, et al. Acute adriamycin-induced cardiotoxicity is exacerbated by angiotension II. Cytotechnology. 2016;68(1):33–43. doi: 10.1007/s10616-014-9748-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Taskin E, Ozdogan K, Kunduz Kindap E, Dursun N. The restoration of kidney mitochondria function by inhibition of angiotensin-II production in rats with acute adriamycin-induced nephrotoxicity. Ren Fail. 2014;36(4):606–12. doi: 10.3109/0886022X.2014.882737. [DOI] [PubMed] [Google Scholar]
- 11.Taskin E, Dursun N. Recovery of adriamycin induced mitochondrial dysfunction in liver by selenium. Cytotechnology. 2015;67(6):977–86. doi: 10.1007/s10616-014-9736-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Yapislar H, Taskin E, Ozdas S, et al. Counteraction of apoptotic and inflammatory effects of adriamycin in the liver cell culture by clinopitolite. Biol Trace Elem Res. 2015 doi: 10.1007/s12011-015-0476-3. [Epub ahead of print] [DOI] [PubMed] [Google Scholar]
- 13.Quiles JL, Huertas JR, Battino M, et al. Antioxidant nutrients and adriamycin toxicity. Toxicology. 2002;180(1):79–95. doi: 10.1016/s0300-483x(02)00383-9. [DOI] [PubMed] [Google Scholar]
- 14.Chen Z, Chua CC, Gao J, et al. Prevention of ischemia/reperfusion-induced cardiac apoptosis and injury by melatonin is independent of glutathione peroxdiase 1. J Pineal Res. 2009;46(2):235–41. doi: 10.1111/j.1600-079X.2008.00654.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Carbajo-Pescador S, Steinmetz C, Kashyap A, et al. Melatonin induces transcriptional regulation of Bim by FoxO3a in HepG2 cells. Br J Cancer. 2013;108(2):442–49. doi: 10.1038/bjc.2012.563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Brzozowska I, Ptak-Belowska A, Pawlik M, et al. Mucosal strengthening activity of central and peripheral melatonin in the mechanism of gastric defense. J Physiol Pharmacol. 2009;60(Suppl 7):47–56. [PubMed] [Google Scholar]
- 17.Aziriova S, Repova Bednarova K, Krajcirovicova K, et al. Doxorubicin-induced behavioral disturbances in rats: protective effect of melatonin and captopril. Pharmacol Biochem Behav. 2014;124:284–89. doi: 10.1016/j.pbb.2014.06.021. [DOI] [PubMed] [Google Scholar]
- 18.Govender J, Loos B, Engelbrecht AM. Melatonin: A protective role against doxorubicin-induced cardiotoxicity. Future Oncol. 2015;11(14):2003–6. doi: 10.2217/fon.15.48. [DOI] [PubMed] [Google Scholar]
- 19.Govender J, Loos B, Marais E, Engelbrecht AM. Mitochondrial catastrophe during doxorubicin-induced cardiotoxicity: A review of the protective role of melatonin. J Pineal Res. 2014;57(4):367–80. doi: 10.1111/jpi.12176. [DOI] [PubMed] [Google Scholar]
- 20.Camelliti P, Borg TK, Kohl P. Structural and functional characterisation of cardiac fibroblasts. Cardiovasc Res. 2005;65(1):40–51. doi: 10.1016/j.cardiores.2004.08.020. [DOI] [PubMed] [Google Scholar]
- 21.Souders CA, Bowers SL, Baudino TA. Cardiac fibroblast: the renaissance cell. Circ Res. 2009;105(12):1164–76. doi: 10.1161/CIRCRESAHA.109.209809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Jaworek J, Leja-Szpak A. Melatonin influences pancreatic cancerogenesis. Histol Histopathol. 2014;29(4):423–31. doi: 10.14670/HH-29.10.423. [DOI] [PubMed] [Google Scholar]
- 23.Kim TK, Kleszczynski K, Janjetovic Z, et al. Metabolism of melatonin and biological activity of intermediates of melatoninergic pathway in human skin cells. FASEB J. 2013;27(7):2742–55. doi: 10.1096/fj.12-224691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Oz E, Erbas D, Surucu HS, Duzgun E. Prevention of doxorubicin-induced cardiotoxicity by melatonin. Mol Cell Biochem. 2006;282(1–2):31–37. doi: 10.1007/s11010-006-1153-9. [DOI] [PubMed] [Google Scholar]
- 25.Boucek RJ, Jr, Miracle A, Anderson M, et al. Persistent effects of doxorubicin on cardiac gene expression. J Mol Cell Cardiol. 1999;31(8):1435–46. doi: 10.1006/jmcc.1999.0972. [DOI] [PubMed] [Google Scholar]
- 26.Nakamura T, Ueda Y, Juan Y, et al. Fas-mediated apoptosis in adriamycin-induced cardiomyopathy in rats: In vivo study. Circulation. 2000;102(5):572–78. doi: 10.1161/01.cir.102.5.572. [DOI] [PubMed] [Google Scholar]
- 27.Roskoski R., Jr ERK1/2 MAP kinases: structure, function, and regulation. Pharmacol Res. 2012;66(2):105–43. doi: 10.1016/j.phrs.2012.04.005. [DOI] [PubMed] [Google Scholar]
- 28.Paul A, Wilson S, Belham CM, et al. Stress-activated protein kinases: Activation, regulation and function. Cell Signal. 1997;9(6):403–10. doi: 10.1016/s0898-6568(97)00042-9. [DOI] [PubMed] [Google Scholar]
- 29.Escribano BM, Diaz-Moreno A, Tasset I, Tunez I. Impact of light/dark cycle patterns on oxidative stress in an adriamycin-induced nephropathy model in rats. PLoS One. 2014;9(5):e97713. doi: 10.1371/journal.pone.0097713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Santofimia-Castano P, Garcia-Sanchez L, Ruy DC, et al. Melatonin induces calcium mobilization and influences cell proliferation independently of MT1/MT2 receptor activation in rat pancreatic stellate cells. Cell Biol Toxicol. 2015;31(2):95–110. doi: 10.1007/s10565-015-9297-6. [DOI] [PubMed] [Google Scholar]
- 31.Dziegiel P, Podhorska-Okolow M, Zabel M. Melatonin: Adjuvant therapy of malignant tumors. Med Sci Monit. 2008;14(5):RA64–70. [PubMed] [Google Scholar]
- 32.Song N, Kim AJ, Kim HJ, et al. Melatonin suppresses doxorubicin-induced premature senescence of A549 lung cancer cells by ameliorating mitochondrial dysfunction. J Pineal Res. 2012;53(4):335–43. doi: 10.1111/j.1600-079X.2012.01003.x. [DOI] [PubMed] [Google Scholar]
- 33.Teodoro BG, Baraldi FG, Sampaio IH, et al. Melatonin prevents mitochondrial dysfunction and insulin resistance in rat skeletal muscle. J Pineal Res. 2014;57(2):155–67. doi: 10.1111/jpi.12157. [DOI] [PubMed] [Google Scholar]
- 34.Kato H, Tanaka G, Masuda S, et al. Melatonin promotes adipogenesis and mitochondrial biogenesis in 3T3-L1 preadipocytes. J Pineal Res. 2015;59(2):267–75. doi: 10.1111/jpi.12259. [DOI] [PubMed] [Google Scholar]
- 35.Guo J, Guo Q, Fang H, et al. Cardioprotection against doxorubicin by metallothionein Is associated with preservation of mitochondrial biogenesis involving PGC-1alpha pathway. Eur J Pharmacol. 2014;737:117–24. doi: 10.1016/j.ejphar.2014.05.017. [DOI] [PubMed] [Google Scholar]
- 36.Wang R, Yan Z, Wu X, et al. Rosiglitazone attenuates renal injury caused by hyperlipidemic pancreatitis. Int J Clin Exp Pathol. 2015;8(5):4332–43. [PMC free article] [PubMed] [Google Scholar]
- 37.Mahmoud-Awny M, Attia AS, Abd-Ellah MF, El-Abhar HS. Mangiferin mitigates gastric ulcer in ischemia/reperfused rats: Involvement of PPAR-gamma, NF-kappaB and Nrf2/HO-1 signaling pathways. PLoS One. 2015;10(7):e0132497. doi: 10.1371/journal.pone.0132497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Cheang WS, Tian XY, Wong WT, et al. Metformin protects endothelial function in diet-induced obese mice by inhibition of endoplasmic reticulum stress through 5′ adenosine monophosphate-activated protein kinase-peroxisome proliferator-activated receptor delta pathway. Arterioscler Thromb Vasc Biol. 2014;34(4):830–36. doi: 10.1161/ATVBAHA.113.301938. [DOI] [PubMed] [Google Scholar]
- 39.Menassol JB, Tautou C, Collet A, et al. The effect of an intracerebroventricular injection of metformin or AICAR on the plasma concentrations of melatonin in the ewe: potential involvement of AMPK? BMC Neurosci. 2011;12:76. doi: 10.1186/1471-2202-12-76. [DOI] [PMC free article] [PubMed] [Google Scholar]