

Abstract
Over decades it has been unclear how individual migratory songbirds cross large ecological barriers such as seas or deserts. By deploying light-level geolocators on four songbird species weighing only about 12 g, we found that these otherwise mainly nocturnal migrants seem to regularly extend their nocturnal flights into the day when crossing the Sahara Desert and the Mediterranean Sea. The proportion of the proposed diurnally flying birds gradually declined over the day with similar landing patterns in autumn and spring. The prolonged flights were slightly more frequent in spring than in autumn, suggesting tighter migratory schedules when returning to breeding sites. Often we found several patterns for barrier crossing for the same individual in autumn compared to the spring journey. As only a small proportion of the birds flew strictly during the night and even some individuals might have flown non-stop, we suggest that prolonged endurance flights are not an exception even in small migratory species. We emphasise an individual’s ability to perform both diurnal and nocturnal migration when facing the challenge of crossing a large ecological barrier to successfully complete a migratory journey.
Twice a year billions of birds undertake a migratory journey of several thousand kilometres to their non-breeding sites and back. The Palearctic-African flyway represents probably the largest avian migration system on earth1. An estimated 2.1 billion songbirds and near-passerines move from Europe to Africa each autumn2. At some point in time, nearly all of them have to cross a major ecological barrier, the 1500–2000 km wide Sahara Desert.
A crucial, and to date debated, issue is which migratory strategy individual songbirds use to reach their destination. In his seminal work, Moreau3 suggested “it does seem that an ability to maintain flight for 50–60 hours without food or water is essential for those birds which regularly migrate across the Sahara”. However, since the 1980s, an accumulating number of studies brought evidence in favour of the alternative intermittent migratory strategy4,5,6,7. In this scenario songbirds cross the Sahara in small steps: flying at night and resting and/or refuelling during the day. The picture, however, seems to be complex, as radar studies from various sites across the globe have brought evidence of nocturnal migrants regularly prolonging flight into the day when crossing large-scale barriers8,9,10,11. The proportion of diurnal migratory traffic in these studies was just a fraction of typical night-time migration, suggesting that landing occurs shortly around sunrise and that only some birds are capable of prolonged daytime flights. This means that some individuals show flexible migratory behaviour and they can switch from nocturnal to partially diurnal migration. In general, nocturnal migration is the prevailing pattern in small birds within the Palaearctic African migration system12. With our own survey we estimate that about 63% of species (44 out of 70 trans-Saharan migrants for which we have collated data; Supplementary Table S1) are expected to migrate during the night when crossing continental Europe. Another 16% migrate solely during the daytime, so the pool of individuals that might theoretically switch to temporary daytime migration is considerable. While we already know that daytime migration and prolonged flights across vast barriers do happen in small songbirds13, the magnitude, temporal and seasonal (spring vs. autumn passage) effects at the individual and species level remain little known for the large barriers between Europe and Africa.
Recent technical development of satellite transmitters and small light intensity data loggers (geolocators) has enabled research showing that at least in some larger migratory species, extreme endurance flights of several thousands of kilometres are possible14,15,16. Here we tracked several small songbird species, weighing about 12 g, and for the first time investigated their individual migratory patterns while crossing the Sahara Desert and the Mediterranean Sea. By the analysis of anomalies in the light patterns recorded during barrier crossing periods we aim to evaluate i) the occurrence and the timing of prolonged flights into the day by typical nocturnal migrants, ii) the day-to-day diversity in migratory patterns, and iii) to compare autumn and spring migration periods.
Results
Spatial and temporal occurrence of light anomalies
All four species in this study, i.e. the collared flycatcher Ficedula albicollis, pied flycatcher Ficedula hypoleuca, Eurasian reed warbler Acrocephalus scirpaceus and aquatic warbler Acrocephalus paludicola (see Supplementary Table S2 for details) are widely thought to migrate at night and rest during daytime. This behaviour is reflected in geolocator data with a zigzagging light pattern during the day, caused by variable exposure of the light sensor during foraging, preening, resting and moving through vegetation (Fig. 1). In both spring and autumn migration periods, however, there was a noticeable pattern of continuous full light intensity (full light pattern; FLP) during the daytime in all four species, i.e. the sensor recorded maximum light levels for uninterrupted periods of several hours (for an overview of FLP anomalies see Fig. 2). This FLP anomaly lasted for 1–3 days. In the collared flycatcher it occurred in 12 out of 13 birds in autumn and in all birds in spring (11 individuals for which data have been recorded until spring). All pied flycatchers showed FLP in both autumn and spring (4 and 2 individuals, respectively). Similarly, all aquatic warblers had FLP (5 and 2 individuals). In reed warblers FLP occurred in 4 out of 12 birds in autumn and in all 8 birds tracked in spring. Summarising across species, all individuals showed FLP in spring and the absence of FLP was observed in autumn only (9 out of 34 tracked individuals). Seven of 9 birds without FLP still had a distinct zigzag pattern of increased light intensities.
Figure 1. Representative examples of full light pattern anomalies (FLP, grey area) in a collared flycatcher (CF), pied flycatcher (PF) and barn swallow (BS) recorded by light-level geolocators on days while crossing the Sahara desert.
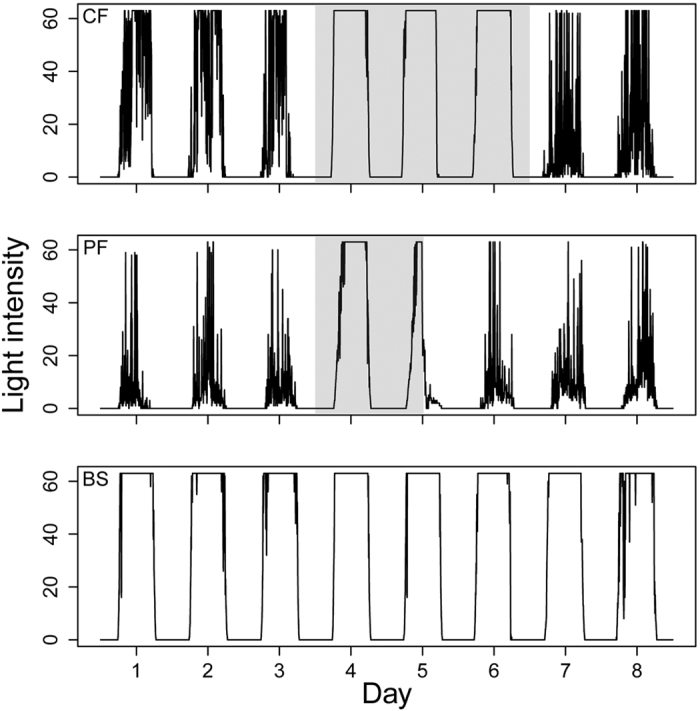
The middle plot (PF) depicts an example of FLP that abruptly ends on the second day at 12:00 UTC. Tick marks on x-axis denote noon. The zigzag light pattern (shading caused by habitat and bird behaviour) a few days before and after the full light pattern in collared and pied flycatchers represents typical geolocator data for most birds throughout their annual cycle. The barn swallow data represent typical geolocator data for an aerial forager.
Figure 2. Detailed seasonal overview of daytime light pattern anomalies (FLP) recorded by geolocators while the birds were crossing the Sahara Desert.

Each category is accompanied by a representative figure of recorded light intensities, % of occurrence (numbers of individuals are in parentheses), description of the anomaly and our most plausible interpretation (categories A–H).
In all four species the occurrence of FLP coincides with the migration time between Europe and Africa (Supplementary Table S2). The majority of the stopover sites preceding and following FLPs are located North and South of the Sahara (Fig. 3), demonstrating that FLP occurred while the birds were crossing the Sahara and/or the Mediterranean Sea.
Figure 3.
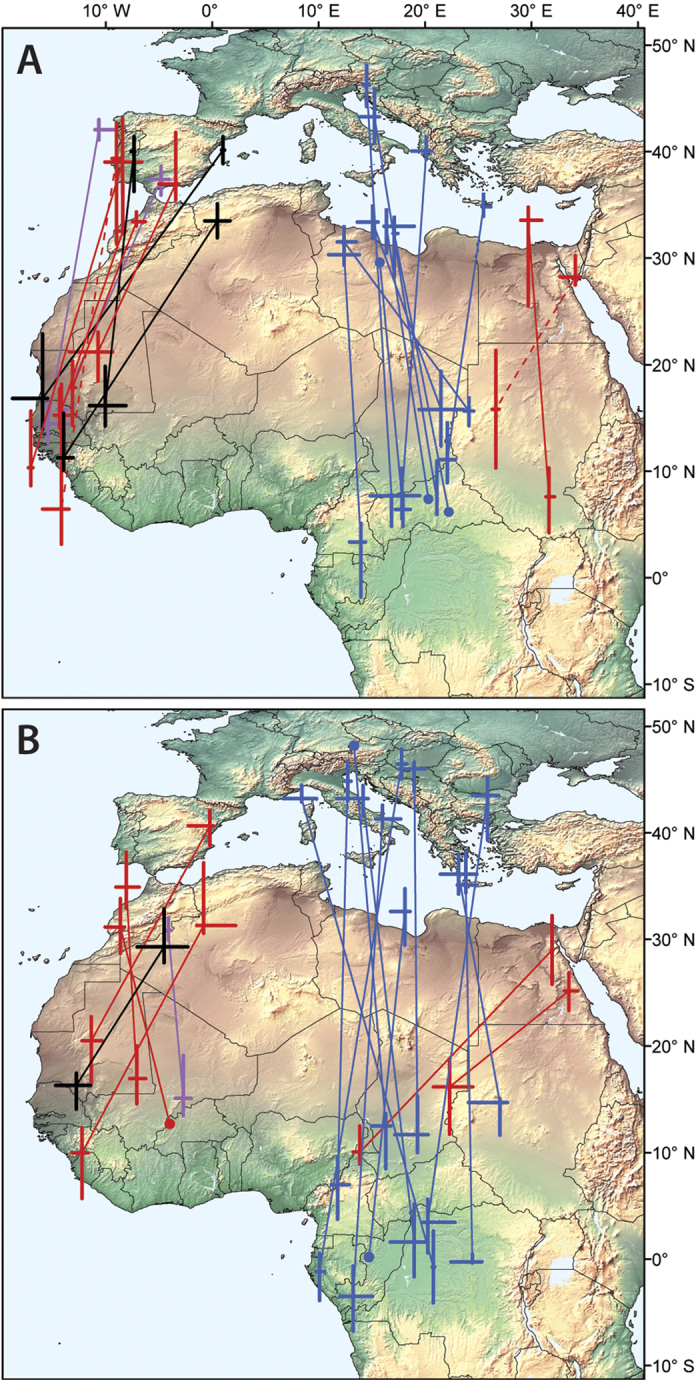
Autumn (A) and spring (B) stationary sites of birds (median ± 25th and 75th percentiles of location estimates) just prior and after the occurrence of the full light pattern (blue = collared flycatcher, black = pied flycatcher, red = Eurasian reed warbler, purple = aquatic warbler). The two dashed lines connect stationary sites for birds without FLP. The background map was made with Natural Earth public domain free vector and raster map data@naturalearthdata.com. The stationary sites were depicted in ArcGIS.
The estimated flight duration between stationary sites before and after the FLP was positively related to the estimated travel distance that the birds had to cross (b = 0.033 ± 0.009 SE, t = 3.9, P = 0.004; Fig. 4), while controlling for the non-significant effect of season (t = 1.5, P = 0.129; model means ± SE: autumn 52.1 ± 7.7 h, spring 47.7 ± 7.6 h; random effects variance: bird identity 49.3 (7.0 SD), species 170.4 (13.1 SD), residual variance 67.6 (8.2 SD)).
Figure 4. Relationship between travel distance (width of the Sahara desert on the individual crossing course) and the estimated duration of flight based on cumulative length of FLP (including night lengths).
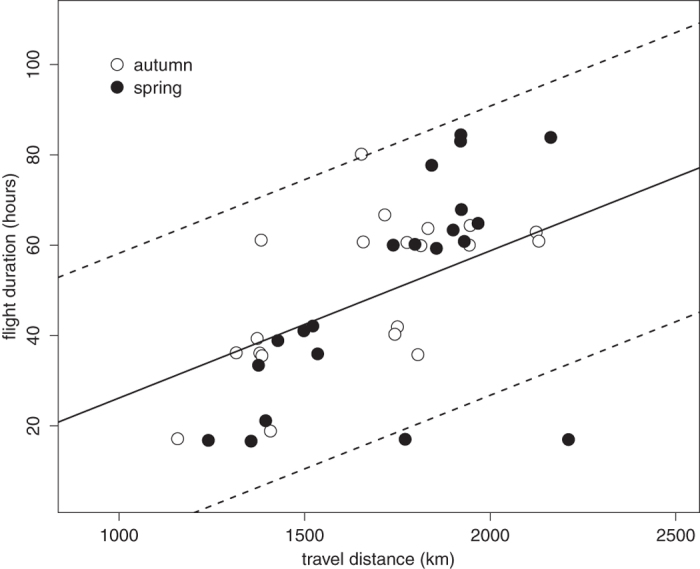
Fitted line ± 95% CI (dashed lines) is from a linear mixed-effect model. Note that there was no seasonal effect in the relationship (see Results). The point in the lower right corner was excluded as an outlier.
Strategies of barrier crossing
In both seasons FLPs often ended abruptly during daytime on the last day of the presumed Sahara crossing (see middle plot of Fig. 1). We interpret this as prolonged flight into the day and the sudden change in light intensities as landing time (Fig. 2). In autumn FLP occurred in 73.5% (25 out of 34) of tracked birds and 47.1% (16) had an abrupt ending of FLP. Abrupt FLP endings mostly occurred on the last day and for the remaining birds various patterns were detected (Fig. 2). In spring all 23 tracked birds had FLP and abrupt ending occurred in 65.2% (15) of individuals, and as in autumn, the abrupt FLP ending occurred often on the last day (Fig. 2). We found no statistical difference in the number of FLPs occurring in autumn compared to spring (Chi-squared test with Yates’ correction χ2 = 0.34, df = 1, P = 0.561). Eight individuals prolonged their flights into the day during the two- or three-day period of barrier crossing but the durations of diurnal flights within individuals varied considerably (Fig. 2 and Supplementary Table S2). Four out of 22 birds tracked during both migratory periods showed the same pattern of barrier crossing (Supplementary Table S3). If we consider categories B and C (Fig. 2) as a variation on the same pattern, then 9 out of 22 birds showed the same pattern.
Consistent with radar measurements (Fig. 5b 6) we found FLP with Tmax < 92 min (threshold time it took for each sunrise event to reach the maximum light intensity) and abrupt FLP endings to occur up to 12 hours after sunrise. The frequency of prolonged flights into the day (i.e. the number of FLPs with abrupt endings during daytime) was higher in spring (50% out of 28 cases) than in autumn (39.5% out of 38 cases; Fig. 5a) but this difference is not statistically significant (Kolmogorov-Smirnov test, D = 0.402, P = 0.31). The flights into the day ceased on average 5.8 ± 3.0 (SD) hours (n = 15 cases) after sunrise in autumn and 6.8 ± 2.1 hours (n = 14 cases) after sunrise in spring.
Figure 5. Estimates of proportions of birds with diurnal flights while crossing the Sahara desert and their timing of landing (inset in the upper right corner).
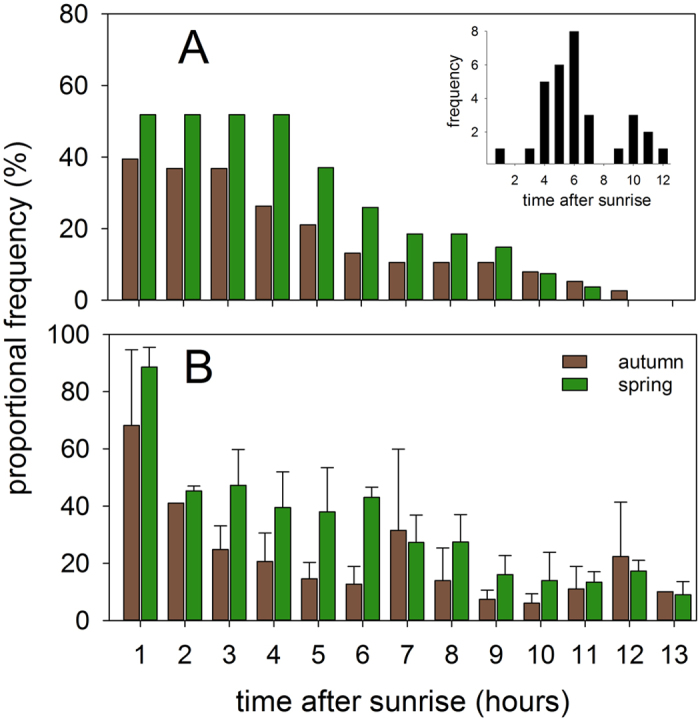
The upper plot (A) shows frequencies of migratory flights into the day estimated from this study (excluding cases when Tmax >92 min) and the lower panel (B) those (mean ± SD) recalculated from a radar field study in the Mauritanian Sahara6.
FLP without abrupt ending or any gaps by shading occurred in 26.5% (9) birds in autumn and 34.8% (8) birds in spring. Among them only one bird had perfect FLP and a low Tmax on both FLP days in autumn (i.e. 2.9% of 34 checked individuals; pied flycatcher, Tmax = 61 and 53 min) and another bird had perfect FLP for one day in spring (i.e. 4.3% of 23 individuals; aquatic warbler, Tmax = 34 min). All remaining birds showed a combination of perfect FLP and gaps by shading.
Our simulation approach showed that, depending on the assumed flight speeds, between 20–80% of individuals fly into the day, to cover the distance for crossing the Sahara desert (Supplementary Fig. S3). The simulation shows that for spring migration a slightly higher percentage of daytime flights are necessary for crossing the desert (Supplementary Fig. S3).
Discussion
High light intensities are typically recorded by geolocators in aerial foragers such as martins or swallows that stay airborne during large parts of the day (Fig. 1). In these “classic” diurnal migrants the light sensor is consistently exposed to the sun during the day. As a result, FLP is recorded regularly. We show here that four songbird species, which have been considered to be mainly nocturnal migrants, also show FLP during a limited migration period that coincides with their crossing of the Sahara desert. We suggest that migratory birds can flexibly switch from typical nocturnal migration to a prolonged flight into the day when facing the task of crossing a major ecological barrier. We found, however, that in three individuals (one aquatic warbler and two collared flycatchers) FLP was also detected outside the main migratory period (Supplementary Table S2). In two birds it occurred in November–December in sub-Saharan Africa (presumably effect of open habitat or perching behaviour) and in one bird at the breeding site in open habitat (marshes). Hence, at the moment we do not have any evidence for diurnal movements north of the Mediterranean.
In 9 individuals we did not find FLP while crossing the Sahara in autumn but for 7 of them we could still observe elevated light intensities that were above the average of typical light data recorded at times prior and after the desert crossing. We suggest that these birds behaved as typical nocturnal migrants that landed before dawn and rested during the day. In contrast, very few individuals (one in autumn and one in spring) showed perfect FLP either during the entire period of the Sahara crossing (or at least on some of the days needed for the crossing). Our very conservative suggestion is that these might be the non-stop flying individuals. An alternative view would be that the birds were resting on the ground without hiding in the shade. This seems unlikely, however, as the majority of observations of grounded birds show that they were actively hiding during daytime hours17. In addition, our simulation approach showed that the birds would have to fly at extreme speeds to cross the recorded distances by nocturnal flight only. Finally the diverse patterns of FLPs (Fig. 2) emphasise that birds are able to perform both diurnal and nocturnal migration even within one and the same journey.
The duration of FLP (i.e. the sum of nocturnal and diurnal periods) was positively related to the distance we calculated that the birds have to fly over the Sahara. For birds that crossed longer distances over the Sahara, multiple and/or longer durations of FLP were recorded. In contrast, birds that crossed the Sahara at its narrowest points had FLP usually only on one day. Interestingly, spring and autumn desert crossing times were similar. Hence it seems that desert crossing is optimized independently from seasonal time pressures; crossing the inhospitable region as fast as possible might be the main aim for every individual. And indeed, currently available geolocator studies showed fast crossing of the Sahara desert7,18,19.
Earlier field studies demonstrated that nocturnally migrating birds of unknown provenience prolong their flights into the day3,11,20,21. It was generally believed that these prolonged flights had just a very short duration and most birds tended to land shortly after dawn6,11,22 (but see10). Based on abrupt endings of the FLP during daytime, we found that the birds on average prolong the flights until noon during both autumn and spring migration. Our estimates of diurnal flights (as estimated from the durations of FLPs and Tmax < 92 min) showed a gradual decline over the day. This pattern was remarkably similar to the one derived from the radar study in the western Sahara6 (recalculated in Fig. 5b). In spring, however, a higher proportion of tracked birds (i.e. with FLP and abrupt ending) prolonged their flights into the day than in autumn. Similarly, this seasonal pattern strongly resembled findings of the radar study by Schmaljohann et al.6. In addition, we found slightly higher occurrence of FLP in spring (25 out of 34 birds in autumn, while in all 23 individuals in spring). These two facts, however, do not match with estimates of desert crossing times which were similar for the two periods (see above). Faster total migration in spring is the general pattern across bird species23 but our data indicate that this might not be the case during desert crossing. That we found slightly more FLPs in spring might result from frequent tailwinds which prevail at higher flight altitudes at this time of the year24. Tailwinds are similar in autumn but at lower flight altitudes, in hotter and dry air24, which might explain the lower proportion of flights into the day. Accordingly, more birds prolonged their migration into the day under tailwind conditions as shown by radar11. Our data suggest that landing or searching for shade can occur nearly at any time of the day, most probably depending on when the bird reaches a suitable destination. This suggests an individually flexible prolongation of nocturnal migration into the day based on the bird’s needs and environmental conditions.
Another striking pattern we found was that the prolonged flights into day occurred most frequently on the last day of barrier crossing. Abrupt FLP endings were followed by much lower and variable light levels (i.e. a typical zigzag pattern) for the rest of the daytime than was typically observed on regular days without FLP. Field observations from the Sahara show that grounded fat birds (i.e. those that do not need to stop for refuelling) were often found resting in shade (e.g. single rocks, wadis, depressions or mountain ridges4,17) and our data seem to be in line with this. By simulation we estimated that depending on the flight speed between 20–80% of birds extended their nocturnal flights into the day in order to travel the distance they did.
To summarise, earlier studies detected diurnal flights of nocturnal migrants3,9,10 but were supposing that a small number of birds were doing so and that landing occurred shortly after dawn6. Here we emphasise that, at least in the four songbird species studied, prolonging flights into the day may be a common migratory pattern during barrier crossing. Such flights might be more common also in other barrier crossing systems, as has been recently shown for blackpoll warblers13. Based on the diverse patterns of FLP and its absence in some individuals, we emphasise the ability of birds to appropriately switch between diurnal and nocturnal migration when facing the challenge of crossing a large ecological barrier. There is accumulating evidence in larger-sized birds of considerable spatial but low temporal variability in migratory behaviour25,26,27. Often we found the same individuals to show different patterns of barrier crossing in autumn compared to spring journeys. This might be in line with the hypothesis of individually optimized migration schedules28. Such an assessment should be possible in the future by using larger data sets containing data from both sexes and repeated tracks of individuals across several migratory seasons.
Methods
Study species and the detection of light anomalies
From 2011 to 2012 individuals of four songbird species were equipped with geolocators (SOI-GDL 2.0, weight approx. 0.6 g, manufactured by the Swiss Ornithological Institute) at their European breeding grounds. After a year we retrieved 34 functional loggers: 13 from collared flycatchers, 4 from pied flycatchers, 12 from Eurasian reed warblers and 5 from aquatic warblers (Supplementary Table S2). The geolocators used an SMD photodiode EPD-470-1-0.9-1 (EPIGAP Optoelektronik, Germany) for light intensity measurements with a sensor wave length between 380 to 555 nm and a maximum range of about 3500 lux (corresponding to 63 arbitrary units). The SOI-GDL 2.0 geolocators recorded ambient light intensity in 5 min intervals.
The geolocators, conventionally used for positioning of migratory birds are also suitable for documenting changes in behaviour over the annual cycle29,30,31,32. When inspecting geolocator data in the four focal species we detected an obvious pattern of continuous full light intensity (hereafter full light pattern–FLP) with regular occurrence twice a year at times that coincide with the migratory period in many species (Fig. 1). We classified FLP as an uninterrupted period of >5 h (or >1 h on days with abrupt FLP ending, see below) during daytime where maximum light intensity (63 in arbitrary scale) was recorded.
An overview of individual FLP cases is given in Supplementary Table S2. The FLPs were classified into several categories in two steps based on a) the amount of shade of the daily light curves and b) time (Tmax) it took for each FLP sunrise event to reach the maximum light intensity (i.e. from 0/1 to 63 units). In the first step we fitted quadratic regressions to the sunrise data (delimited by the time of sunrise using the software Geolocator (SOI, Sempach) and the first consecutive data point which reached maximum light intensity) and sunset data (delimited by the last data point which reached maximum light intensity and sunset determined by the R-package GeoLight, version 1.0333) and summed up the absolute residuals. For the daytime period (delimited by the first and the last data point which reached maximum light intensity) we summed up all deviations from the maximal light intensity. These sums were used to assign every sunrise, day and sunset to the following categories: 1) perfect FLP, virtually no shading; 2) slight shading; 3) substantial shading. Additionally we assigned category 4 to FLPs with an abrupt start or end (Supplementary Fig. S1). In the second step, we calculated the Tmax for each sunrise FLP event. We assumed that during the flight the bird was at an unknown height above ground. This implies no shading by vegetation or by folded wings occasionally covering the light sensor and thus a rapid increase in the recorded light intensity from 0 at twilight to maximum values. To extract sunrises for potential flights prolonged into the day from other sunrises, we compared the data to the sunrise pattern recorded by a typical diurnal migrant and aerial forager. We used light-level logger data of barn swallows Hirundo rustica breeding in southern Switzerland and migrating along the central European-African flyway34. We selected 6 days during autumn (n = 10 birds) and spring (n = 7 birds) migration, when the birds moved between 16° and 35°N (southern borders of the Sahel and the Mediterranean Sea) and vice versa. This was at periods between 12–30 Sept and 10 March–17 April. The maximum Tmax value and its 95th percentile in barn swallows were 127 min and 91 min, respectively (Supplementary Fig. S2). The latter was used as a threshold for our conservative estimates of prolonged flights into the day. Hence, unless otherwise stated, for further analyses of flight into the day we considered only those FLP cases when Tmax < 92 min (68 FLP events, 27 excluded) and the FLP was classified as 1, 2 or 4.
Determining stationary periods
Data from autumn and spring were analysed independently using January 1 as a separator. We calculated stationary periods prior and after the occurrence of FLP using the changeLight function of the R-package GeoLight with minimum staging period set at 3 days. We filtered outlying positions that were >800 km from the median latitude of a given stationary site. We defined a stationary site to be the median of the geographic coordinates ± their 25th/75th percentiles within the particular stationary period. The same number of interquartile ranges (k = 2) of the loessFilter function was used for all individuals of the same species except for one bird (7OY, k = 1.1). To determine the first stationary period before and after the FLP the probability threshold of the changeLight function was adjusted for each bird individually. For autumn, geographic positions of the stationary periods before and after the FLP were calculated using sun-elevation angles derived from the in-habitat calibration in the breeding areas or Hill-Ekstrom calibration from data of the respective stationary period35. When one of the calibration techniques was not applicable or failed, the other was used instead. Please note that we were not able to determine stationary sites for all birds (available estimates are for 22 birds in autumn and 19 in spring). An example of light data profile used to determine the stationary periods before and after crossing the barrier is provided in Supplementary Fig. S4.
Duration of light anomalies
We considered two scenarios for estimating the duration of potential flight over the Sahara Desert at times when FLP occurred: nocturnal flight only or including prolonged flight into the daytime. When FLP ended abruptly during daytime, we took that abrupt change (accuracy to 5 min) as a termination of FLP and the assumed prolonged flight. We estimated the theoretical duration of the prolonged flight as nocturnal flight plus FLP. We assumed that the bird took off for the flight within an hour after sunset the day preceding the occurrence of FLP35,36,37,38. For cases when there were two or more periods of FLP separated by days without FLP, we excluded the daytime non-FLP period from the estimates of flight times. In those cases when an abrupt end of FLP occurred during the day, we added the time period between sunrise and the moment of abrupt decline of light data to the nocturnal flight duration. For cases without an abrupt end of FLP, landing time was estimated to be within an hour before the sunrise on the day that followed the FLP day11. Duration of nocturnal flights only was estimated as a sum of night lengths before, during and after the FLP.
We compared the frequency of potential flights into day based on abrupt FLP endings with those found in an empirical study provided by6. We recalculated the migration traffic rates from their original dataset by setting nocturnal migration traffic rates to 100 and calculated the declining proportion of traffic rates binned to hours after sunrise in autumn and spring.
Flight range estimates during FLP times
Distance between stationary sites was measured as the loxodromic distance between median positions of the last stationary site before the FLP and the first thereafter. For an approximation of barrier crossing distances, we estimated the width of the Sahara desert (minimum travel distance) at points where the bird presumably entered and exited the desert on the loxodromic line that connects the stationary sites just before and after the FLP. Northern and southern desert borders were derived from the land cover map from the GLC2000 database, European Commission Joint Research Centre, http://bioval. jrc.ec.europa.eu/products/glc2000/glc2000.php. We hypothesized that the duration of FLP was driven by the width of the desert and the barrier-crossing strategy of an individual. The relationship between travel distance and the estimated duration of flight during FLP was assessed by a linear mixed-effect model in the R-package lme4. We ran a model with flight duration as response variable that included our estimates of summed time for both nocturnal and diurnal migration (n = 40 cases after excluding one case, a reed warbler where a distance of 2211 km in 17 h was considered as an outlier, see Fig. 4. This individual would have to fly at speed of ca 130 km h−1 which is very unlikely). The fixed effect was travel distance, while season (autumn, spring) was taken as a covariate. Individual identity nested within species was entered as a random effect. We obtained similar results (not shown) when we ran the same analysis with travel distances between the stationary sites. All data analyses were conducted in R version 3.0.139.
Ethical note
The field work was carried out in accordance with the current laws of Belarus, Czech Republic, Finland, Germany, Sweden and Ukraine. The procedures used to handle and fit the birds with geolocators were approved by Academy of Sciences of the Czech Republic (#38/2011), Varsinais-Suomi Centre for Economic Development, Transport and the Environment (#LOS-2009-L-308-259), Landratsamt Saale-Orla-Kreis (#16.075.364.622.0 SC/12), Landkreis Leipzig (364.620/15/7/4), Stockholms södra djurförsöksetiska nand (#S55-11), Ukrainian Ministry of Ecology and Natural Resources (1/2011) and by ethical committees of Palacký University and Czech Ministry of Education (#1/2011, licence #CZ00231).
Additional Information
How to cite this article: Adamík, P. et al. Barrier crossing in small avian migrants: individual tracking reveals prolonged nocturnal flights into the day as a common migratory strategy. Sci. Rep. 6, 21560; doi: 10.1038/srep21560 (2016).
Supplementary Material
Acknowledgments
We thank H. Schmidt for providing data on migratory strategies of birds, and M. Král, J. Träff, M. Požgayová, V. Jelínek, J. Koleček, B. Meister, J. Auerswald, K. Fletcher, J. Könönen, B.B. Babu, S. Zonneveld, E. YeXiong, M. Ahola, W. Velmala, M. Flade, O. Ilucha, G. Kiljan, I. Legeyda, U. Malashevich, A. Poluda and many other volunteers for field assistance. Funding was provided by Czech Science Foundation (grant #13-06451S), Swiss Federal Office for Environment (UTF-Nr. 254, 332, 363, 400), Royal Society for the Protection of Birds, O.M.H. Schmidt-Felsche Foundation, Swedish Research Council and FORMAS.
Footnotes
Author Contributions P.A., T.E., M.B. and S.H. conceived the study design, collected data, performed data analyses, wrote the manuscript. L.G., I.H., M.K., T.L., F.L., P.P. and V.S. contributed with data, commented on the manuscript. All authors approved the manuscript.
References
- Newton I. The Migration Ecology of Birds. Ch. 24, 699–727 (Academic Press, 2008). [Google Scholar]
- Hahn S., Bauer S. & Liechti F. The natural link between Europe and Africa–2.1 billion birds on migration. Oikos 118, 624–626 (2009). [Google Scholar]
- Moreau R. E. Problems of Mediterranean-Saharan migration. Ibis 103, 373–427, 580–623 (1961). [Google Scholar]
- Biebach H., Friedrich W. & Heine G. Interaction of bodymass, fat, foraging and stopover period in trans-Sahara migrating passerine birds. Oecologia 69, 370–379 (1986). [DOI] [PubMed] [Google Scholar]
- Bairlein F. How do migratory songbirds cross the Sahara? Trends Ecol. Evol. 3, 191–194 (1988). [DOI] [PubMed] [Google Scholar]
- Schmaljohann H., Liechti F. & Bruderer B. Songbird migration across the Sahara: the non-stop hypothesis rejected! Proc. R. Soc. B-Biological Sci. 274, 735–739 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eraud C. et al. Migration routes and staging areas of trans-Saharan Turtle Doves appraised from light-level geolocators. PLoS One 8, e59396 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gauthreaux S. A. In Anim. Migr. Navig. Homing (ed. Schmidt-Koenig K. & Keeton W. T.) 219–227 (Springer-Verlag, 1978). [Google Scholar]
- Nisbet I., McNair D., Post W. & Williams T. Transoceanic migration of the Blackpoll Warbler: summary of scientific evidence and response to criticisms by Murray. J. F. Ornithol. 66, 612–622 (1995). [Google Scholar]
- Biebach H. et al. Strategies of passerine migration across the Mediterranean Sea and the Sahara Desert: a radar study. Ibis 142, 623–634 (2000). [Google Scholar]
- Schmaljohann H., Liechti F. & Bruderer B. Daytime passerine migrants over the Sahara–are these diurnal migrants or prolonged flights of nocturnal migrants? Ostrich 78, 357–362 (2007). [Google Scholar]
- Dorka V. Das jahres- und tageszeitliche Zugmuster von Kurz- und Langstreckenziehern nach Beobachtungen auf den Alpenpässen Cou/Bretolet (Wallis). Der Ornithol. Beobachter 63, 165–223 (1966). [Google Scholar]
- DeLuca W. V et al. Transoceanic migration by a 12 g songbird. Biol. Lett. 11, 20141045 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gill R. E. et al. Extreme endurance flights by landbirds crossing the Pacific Ocean: ecological corridor rather than barrier? Proc. R. Soc. B-Biological Sci. 276, 447–458 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klaassen R. H. G., Alerstam T., Carlsson P., Fox J. W. & Lindström A. Great flights by great snipes: long and fast non-stop migration over benign habitats. Biol. Lett. 7, 833–5 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bairlein F. et al. Cross-hemisphere migration of a 25 g songbird. Biol. Lett. 8, 505–7 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haas W. & Beck P. Zum Frühjahrszug paläarktischer Vögel über die westliche Sahara. J. Ornithol. 120, 237–246 (1979). [Google Scholar]
- Tøttrup A. P. et al. The annual cycle of a trans-equatorial Eurasian-African passerine migrant: different spatio-temporal strategies for autumn and spring migration. Proc. R. Soc. B-Biological Sci. 279, 1008–16 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lemke H. W. et al. Annual cycle and migration strategies of a trans-Saharan migratory songbird: a geolocator study in the great reed warbler. PLoS One 8, e79209 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spina F. & Pilastro A. Strategy of sea and desert crossing in spring passerine migrants as suggested by the analysis of intra-and inter-specific variation of residual fat levels. In Proc. 22 Int. Ornithol. Congr., Durban (ed. Adams N. & Slotow R.) 1958–1976 (BirdLife South Africa, 1999). [Google Scholar]
- Grattarola A., Spina F. & Pilastro A. Spring migration of the Garden Warbler (Sylvia borin) across the Mediterranean Sea. J. Ornithol. 140, 419–430 (1999). [Google Scholar]
- Bruderer B. & Liechti F. Bird migration across the Mediterranean. in Proc. 22 Int. Ornithol. Congr., Durban (ed. Adams N. & Slotow R.) 1983–1999 (BirdLife South Africa, 1999). [Google Scholar]
- Nilsson C., Klaassen R. & Alerstam T. Differences in speed and duration of bird migration between spring and autumn. Am. Nat. 181, 837–845 (2013). [DOI] [PubMed] [Google Scholar]
- Liechti F. Birds: blowin’ by the wind? J. Ornithol. 147, 202–211 (2006). [Google Scholar]
- Stanley C. Q., MacPherson M., Fraser K. C., McKinnon E. A. & Stutchbury B. J. M. Repeat tracking of individual songbirds reveals consistent migration timing but flexibility in route. PLoS One 7, e40688 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vardanis Y., Klaassen R. H. G., Strandberg R. & Alerstam T. Individuality in bird migration: routes and timing. Biol. Lett. 7, 502–5 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- López-López P., García-Ripollés C. & Urios V. Individual repeatability in timing and spatial flexibility of migration routes of trans-Saharan migratory raptors. Curr. Zool. 60, 642–652 (2014). [Google Scholar]
- Battley P. F. Consistent annual schedules in a migratory shorebird. Biol. Lett. 2, 517–520 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eichhorn G., Afanasyev V., Drent R. H. & Jeugd H. P. Van Der. Spring stopover routines in Russian Barnacle Geese Branta leucopsis tracked by resightings and geolocation. Ardea 94, 667–678 (2006). [Google Scholar]
- Van Wijk R. E., Schaub M., Tolkmitt D., Becker D. & Hahn S. Short-distance migration of Wrynecks Jynx torquilla from Central European populations. Ibis 155, 886–890 (2013). [Google Scholar]
- Gow E., Wiebe K. & Fox J. Cavity use throughout the annual cycle of a migratory woodpecker revealed by geolocators. Ibis 157, 167–170 (2014). [Google Scholar]
- Burger J., Niles L., Porter R. & Dey A. Using geolocator data to reveal incubation periods and breeding biology in Red Knots Calidris canutus rufa. Wader Study Gr. Bull. 119, 26–36 (2014). [Google Scholar]
- Lisovski S. & Hahn S. GeoLight–processing and analysing light-based geolocator data in R. Methods Ecol. Evol. 3, 1055–1059 (2012). [Google Scholar]
- Liechti F. et al. Timing of migration and residence areas during the non-breeding period of barn swallows Hirundo rustica in relation to sex and population. J. Avian Biol. 46, 254–265 (2015). [Google Scholar]
- Lisovski S. et al. Geolocation by light: accuracy and precision affected by environmental factors. Methods Ecol. Evol. 3, 603–612 (2012). [Google Scholar]
- Alerstam T. Bird Migration. 227–350 (Cambridge University Press, 1990). [Google Scholar]
- Åkesson S., Alerstam T. & Hedenström A. Flight initiation of nocturnal passerine migrants in relation to celestial orientation conditions at twilight. J. Avian Biol. 27, 95–102 (1996). [Google Scholar]
- Smolinsky J. A., Diehl R. H., Radzio T. A., Delaney D. K. & Moore F. R. Factors influencing the movement biology of migrant songbirds confronted with an ecological barrier. Behav. Ecol. Sociobiol. 67, 2041–2051 (2013). [Google Scholar]
- R Development Core Team. R: A language and environment for statistical computing. (R Foundation for Statistical Computing, 2012). at http://www.r-project.org.
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.