

Abstract
The primary clinical motor symptoms of Parkinson’s disease (PD) result from loss of dopaminergic (DA) neurons in the substantia nigra (SN). Consequently, neurogenesis of this group of neurons in the adult brain has drawn considerable interest for the purpose of harnessing endogenous neurogenerative potential as well as devising better strategies for stem cell therapy for PD. However, the existence of adult neurogenesis for DA neurons within the SN remains controversial. To overcome technical and design limitations associated with previous studies, our group has developed a novel genetic mouse model for assessing adult nigral DA neurogenesis. This system utilizes transgenic mice that express a tamoxifen-activatable Cre recombinase (CreERT2) under the control of the neuronal progenitor cell promoters nestin or Sox2 leading to suppression of the DA neuron marker tyrosine hydroxylase (TH) via excision of exon 1 by flanking loxP sites in adult animals. This study reports that six months following initiation of a six week treatment with tamoxifen mice with nestin-mediated Th excision displayed a significant reduction in TH+ neurons in the SN. This finding indicates that nestin-expressing cells regenerate DA neurons within the SN of adult animals. Interestingly, no reduction was observed in TH+ cells following Sox2-mediated Th excision suggesting that a nestin+/SOX2- precursor cell population drives DA neurogenesis in the adult SN. This information represents a substantial leap in current knowledge of adult DA neurogenesis, will enable improved in vitro and in vivo modeling, as well as facilitate the harnessing of this process for therapeutic intervention for PD.
Keywords: adult neurogenesis, dopaminergic neurons, substantia nigra, mouse model, Parkinson’s disease
Graphical Abstract
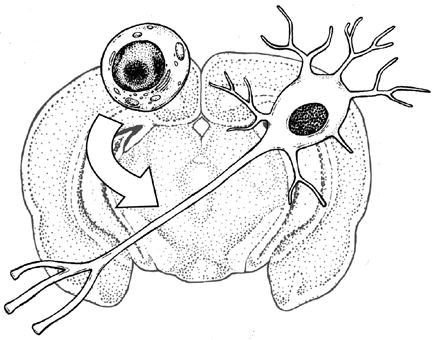
Nestin+/SOX2- cells give rise to dopaminergic neurons in the adult mouse substantia nigra.
Introduction
Parkinson’s disease (PD) is the most common motor disorder and the second most prevalent neurodegenerative disease. PD motor dysfunction (rigidity, tremor, bradykinesia, and postural instability) results from loss of dopaminergic (DA) neurons in the substantia nigra (SN). However, a basic understanding of the mechanism for DA neuron loss remains elusive. Consequently, only symptomatic treatments exist for PD and none that address the underlying neurodegeneration. Determining whether DA neurons are replenished in the adult SN is fundamental to understanding the loss of these neurons during PD. One possibility is that suppression of adult DA neurogenesis could be a driving force toward PD. Additionally, exploiting endogenous neurogenesis could offer potential avenues for therapy as well as better inform stem cell transplantation efforts.
Deficiencies in the process of adult neurogenesis have been strongly associated with Alzheimer’s disease (AD), a neurodegenerative disorder sharing many epidemiological features with PD. Hippocampal neurons responsible for memory processing are preferentially lost in AD and evidence suggests this may be due, in part, to decreased neurogenesis (reviewed in [1]). Hippocampal adult neurogenesis occurs at a high rate throughout life but appears to decrease sharply in human AD as well as mouse models of the disease. Interventions that promote adult neurogenesis improve preclinical model outcomes for histopathology as well as functionality. Neurogenesis of hippocampal neurons in adult humans and rodents begin from stem cell (SC) populations residing in the subgranular zone (SGZ) of the dentate gyrus. SC populations are relatively less differentiated and possess replicative capacity. Therefore, these cells can regenerate the SC pool as well as contribute progeny that can differentiate into neuronal precursor cells (NPCs) that are post-mitotic and committed to neuronal fates.
In addition to hippocampal neurons, olfactory neurons are replenished throughout the life of primates and rodents. SCs responsible for this process reside in the subventricular zone (SVZ). Once born, NPCs migrate a considerable distance to the olfactory bulb before integration and final differentiation into functional neurons. Currently, the SGZ and SVZ are the only known locations for SC generation in the adult mammalian brain. However, whether NPC pools reside elsewhere or transdifferentiation to NPCs from existing cell types occur remains to be determined. The evidence for a contribution of adult neurogenesis to PD is inconclusive. Reduced proliferation of SVZ cells has been reported in human PD [2,3]. Additionally, a number of studies have reported that wide-spread overexpression of a protein known to be a monogenic cause of rare forms of PD, alpha-synuclein (WT, A53T, E46K, and A30P), in mice results in inhibition of neurogenesis in the SGZ and SVZ [4-8]. Moreover, transgenic mouse overexpression of PD-causing mutations in leucine-rich repeat kinase 2 (LRKK2) hinder proliferation and survival of SCs in the SVZ and SGZ [9]. However, a major caveat is that none of these mouse models induce DA neuron loss in the SN so the relevance to DA neurogenesis or even PD is debated. Administering the PD model neurotoxins 6-OHDA or MPTP in rats or mice, respectively, does target DA neurons and inhibition of SC generation in the SVZ has been reported [2,10,11]. Conversely, other groups have reported increased neurogenesis following MPTP treatment [12]. In any event, the DA neuron loss in these models is rapid, easily achieving 80-100% loss within two weeks. Such a speedy loss does not mirror human progressive PD and cannot be explained by suppression of neurogenesis making connections between PD and this process using these neurotoxins dubious.
Attempts have been made to monitor adult DA neurogenesis in the SN directly using various cell lineage tracing methods with conflicting results [13-15]. Lack of compelling evidence for DA neurogenesis has reinforced the prevailing notion that the presence of a toxic stimulus or trophic factor withdrawal induces mature DA neurons to undergo death in a slow and progressive manner. Therefore, while still controversial, the predominate viewpoint is that stem cell replacement of adult DA neurons in the SN does not occur at appreciable levels [13]. The most prevalent method of DA neuron lineage tracing utilizes DNA incorporation of the thymidine analog bromodeoxyuridine (Brd-U) or similar reagent to monitor for cell division of SCs. This method is problematic for a number of reasons. Firstly, Brd-U is quite toxic to the organism and to dividing cells. Secondly, Brd-U can yield false-positive results for cells undergoing DNA repair. Additionally, these regimens of Brd-U use are adopted from studies focused in regions of the brain exhibiting high levels of neurogenesis such as the hippocampus. Moreover, use of this chemical requires double-immunolabeling for the DA neuron marker tyrosine hydroxylase (TH) and Brd-U. Resolution of double-positive cells requires great care using confocal microscopy and issues have been reported [15]. This method is also not readily amenable to large sample numbers. Another common method of lineage tracing is by injection of retrovirus to permanently label SCs and NPCs. However, this requires knowledge of the precursor cell location which is currently unknown. The aim of this study was to overcome these limitations by employing a novel system that utilizes a drug that could be administered for 6-8 weeks without severe health complications, extends the post-labeling duration, and allows for accurate quantitation of DA neurons in the SN of mice by single-labeling.
Methods
All husbandry and study procedures involving mice were performed in accordance with Boise Veterans Affairs Medical Center Institutional Animal Care and Use Committee guidelines. Thlox mice [16] were kindly provided by Drs. Richard Palmiter and Martin Darvas at The University of Washington. The nestin-CREERT2 [17] and Sox2-CREERT2 [18] mouse lines were obtained from The Jackson Laboratory. Mice were fed ad libitum and maintained on 12hr light/dark cycles. For tamoxifen treatment, three-month-old mice were provided 400mg/kg tamoxifen citrate chow (Envigo) as sole food source for 6 weeks. Mice were then placed back on standard rodent chow for the remainder of the study.
For Immunohistochemistry (IHC), mice (n = 6 per group) were anesthetized with 5% isoflurane and transcardially perfused with 10mL 0.1M phosphate buffer (PB) (pH 7.2) with 1mM EDTA followed by 10mL 4% paraformaldehyde (PFA) in PB. Brains were harvested and then fixed in 4% PFA for 24h at 4°C. Next, brains were cryoprotected in 30% sucrose/PB at 4°C until sunk. The brains were then frozen in OCT media and a Leica CM1950 crysostat used to cut 35 μm sections. Free-floating IHC was performed with three PB washes between each step. Endogenous peroxidase activity was removed with H2O2 (3%) and methanol (10%) for 30min followed by blocking and permeabilization with 0.5% bovine serum albumin/0.2% triton X100/PB solution for 1h. To label DA neurons a primary antibody against tyrosine hydroxylase (EMD Millipore; AB152) (1:2000 dilution) was used overnight at 4°C. A secondary antibody conjugated to biotin (Jackson ImmunoResearch) (1:500 dilution) was then applied for 1h at room temperature followed by ABC staining (Vector Labs) (A and B solutions at 1:100 dilutions) for 1h. DA neurons were visualized using 3,3′ diaminobenzidine (DAB) (Sigma Aldrich) substrate addition at 1mg/mL (pH 7.2) and allowed to develop for 4 min. The reaction was stopped in ddH2O, sections placed on slides, dried, and coverslips mounted using Vectamount.
All DA neuron somas in the SN for all SN-containing sections in the right hemisphere (average of 38.6 sections per hemisphere) were counted using bright field microscopy. The left hemisphere was punctured with a 30 gauge needle at the time of cryosectioning to identify hemispheres. To correct for bisected DA neuron somas appearing in adjacent sections, the Abercrombie factor was determined and applied to counts [19]. Data is presented as mean ± SEM for groups. Statistical significance (p < 0.05) between groups was determined using multiple-way ANOVA followed by a post hoc Tukey’s test using GraphPad Prism 6 software.
Results
A genetic approach was developed to remove the DA neuron marker tyrosine hydroxylase (TH) from precursor cells in adult mice (Fig. 1). Consequently, if DA neurons were replenished in adult mice, there should be a gradual loss of TH positive neurons in the SN over time following removal of the Th gene from precursor cells. Tamoxifen-inducible Th excision was chosen for this drug’s ability to readily cross the blood-brain barrier and good tolerability. To target NPCs for Th excision, nestin and Sox2 promoters were selected to drive the expression of a tamoxifen-inducible CRE recombinase (CREERT2) in SCs and/or NPCs. Nestin and Sox2 are two of the most well-described neural progenitor markers and are not expressed in mature neurons making them good initial candidates to target DA neuron precursors. Both nestin-CREERT2 [17] and Sox2-CREERT2 [18] mouse lines were crossed with transgenic mice possessing the endogenous Th gene engineered with loxP sites flanking exon 1 [16] thereby allowing for the excision and silencing of Th in the presence of CRE activity. Three-month-old double or single (control) transgenic mice (nestinCRE-ERT2:Thlox/lox, Sox2CRE-ERT2:Thlox/lox, or Thlox/lox) were treated with or without tamoxifen-laden chow for six weeks (Fig. 2A). Six months following initiation of tamoxifen treatment brains were harvested and DA neurons in the SN counted. Remarkably, tamoxifen treated nestinCRE-ERT2:Thlox/lox mice exhibited significantly fewer TH+ neurons in the SN than controls (Fig. 2B, Fig S1). In addition, no reduction in TH+ cells was observed in the SN of untreated nestinCRE-ERT2:Thlox/lox mice or tamoxifen-treated Sox2CRE-ERT2:Thlox/lox or Thlox/lox mice indicating that the loss in TH signal was due to Th gene excision and not the result of non-specific downregulation of Th expression by tamoxifen or the presence of the CRE transgene. Therefore, this result indicates that a nestin-positive population of precursors replenishes adult DA neurons in mice. However, it was surprising to find that Sox2CRE-ERT2:Thlox/lox mice showed no TH+ cell loss given that Sox2 has been widely reported to be a ubiquitous neural SC marker [20,21].
Figure 1. Genetic model to assess adult DA neurogenesis.

Tamoxifen (Tam) treatment activates CRE activity in nestin or Sox2-expressing cells resulting in Th gene silencing.
Figure 2. Adult DA neurogenesis by nestin positive cells.

Six week treatment with tamoxifen (Tam) citrate chow (400mg/kg chow) was used to activate CREERT2 activity in 3-month-old transgenic mice (A). NestinCRE-ERT2:Thlox/lox mice showed decreased DA neurons in the SN following Tam treatment (B). DA neurons in the SN were visualized by IHC (DAB) using a TH antibody. All sections containing SN from the right hemisphere were counted and neuron totals were corrected using the Abercrombie factor. Multiple-way ANOVA was performed followed by posthoc Tukey’s test (6 mice per group; error bars = SEM; * = p<0.05; ns = not significant).
Discussion
This study provides compelling evidence for DA neurogenesis in the SN of adult mice by utilizing a novel cell lineage tracing model. Discovery of a nestin+ pool of DA progenitor cells will empower future studies to focus on the process of adult neurogenesis for DA neurons as well as enable locating these cells within the mammalian brain. Interestingly, this study found that Sox2 was not expressed in progenitors that gave rise to DA neurons in the adult SN. Sox2 expression is largely restricted to neural SCs, being turned down following cell cycle exit [22]. Additionally, Sox2 expression is closely linked to SC function, being one of four transcription factors (Oct3/4, Sox2, c-Myc, and Klf4) whose overexpression in concert can induce adult somatic cells to become embryo-like SCs [23]. Nestin is an intermediate filament protein expressed by SCs and NPCs in vivo and in vitro and may persist for a longer period of time than Sox2 expression (reviewed in [24]). Thus, targeting nestin-expressing cells will affect SOX2+ SCs in addition to a more differentiated progenitor population. Taken together, this may suggest that SOX2- cells giving rise to DA neurons in the adult SN exhibit greater differentiation and are not renewed by SCs. The implication would be that there is a limited supply of DA neuron progenitors available for adult neurogenesis. Therefore, depletion of this progenitor pool by normal turnover or by deleterious factors would result in an eventual loss of mature DA neurons. Another possibility is that SCs are not needed due to transdifferentiation of progenitors from another cell type. Investigation of additional neural SC and NPC markers by cell lineage tracing in vivo might address this.
The cause of DA neuron loss in PD has remained a mystery despite several decades of intense investigation. Over this time, considerable effort has been placed in identifying factors that induce death of mature DA neurons in vitro and in vivo. However, if DA neurogenesis is a natural ongoing homeostatic mechanism in the brain as evidence presented here suggests, inhibition of this process could be responsible, at least in part, for the progressive loss of DA neurons observed in PD. Reports of heightened sensitivity for SC and NPC populations in the adult brain to enhanced inflammatory response and other toxic stimuli support this notion [25-27]. Interestingly, the rate for DA neuron loss in the SN using a purely inflammatory model of PD reported by two groups mirrors TH+ cell loss by nestin-mediated Th excision shown in Fig. 2B. Extrapolating for single brain hemispheres, data presented by Frank-Cannon et al. [28] and Morrison et al. [29] indicate an approximate rate of 14.3 and 12.3 DA neuron loss/day, respectively. If the total TH+ cell loss in the SN observed in Fig. 2B is divided by the tamoxifen treatment duration of six weeks, a rate of 13.9 ± 1.2 is yielded. This is an intriguing correlation that would also suggest that the enhanced inflammatory model might mediate neurodegeneration by impeding adult neurogenesis of DA neurons. Future studies are warranted to further substantiate this association.
Findings from this study may direct stem cell replacement therapy for PD. Growth of stem cell research in the late 1980’s generated great interest for use of this technology as a potential PD treatment. However, clinical trials using fetal stem cells to replace lost DA neurons have yielded promising but variable results [30,31]. The variability was believed to result from inconsistent source stem cells. Thorough characterization of neural stem cells in vivo may allow for the development of appropriate and consistent cell types for therapy. Therefore, identification and characterization of the newly discovered nestin+/Sox2- DA neuron precursor population will provide a foundation for these investigations.
In conclusion, this study reveals the existence of a nestin+/SOX2- DA progenitor pool that replenishes mature DA neurons in the adult mouse SN. This finding has broad implications for brain biology and PD pathology and serves as a basis for future investigation of these important cells.
Supplementary Material
Highlights.
Novel model developed for cell lineage tracing of dopaminergic neuron progenitors.
Nestin+ neural progenitors are responsible for replenishing nigral dopaminergic neurons.
Sox2 is not expressed by adult dopaminergic neuron progenitors.
Acknowledgments
This work was supported by resources from Boise State University and the Boise VA Medical Center, Boise, ID. Undergraduate student internship support was provided by Institutional Development Awards (IDeA) from the National Institute of General Medical Sciences of the National Institutes of Health under Grants #P20GM103408 and P20GM109095. The authors also acknowledge support from The Biomolecular Research Center at Boise State. In addition, gratitude is extended to Drs. Richard Palmiter and Martin Darvas at The University of Washington for providing Thlox transgenic mice.
Abbreviations
- PD
Parkinson’s disease
- AD
Alzheimer’s disease
- DA
dopaminergic
- TH
tyrosine hydroxylase
- Sox2
SRY (sex determining region Y)-box 2
- SGZ
subgranular zone
- SVZ
subventricular zone
- SC
stem cell
- NPC
neuronal precursor cell
- 6-OHDA
6-hydroxydopamine
- MPTP
1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Winner B, Kohl Z, Gage FH. Neurodegenerative disease and adult neurogenesis. Eur J Neurosci. 2011;33:1139–1151. doi: 10.1111/j.1460-9568.2011.07613.x. [DOI] [PubMed] [Google Scholar]
- 2.Höglinger GU, Rizk P, Muriel MP, Duyckaerts C, Oertel WH, Caille I, et al. Dopamine depletion impairs precursor cell proliferation in Parkinson disease. Nat Neurosci. 2004;7:726–735. doi: 10.1038/nn1265. [DOI] [PubMed] [Google Scholar]
- 3.O’Keeffe GC, Tyers P, Aarsland D, Dalley JW, Barker RA, Caldwell MA. Dopamine-induced proliferation of adult neural precursor cells in the mammalian subventricular zone is mediated through EGF. Proc Natl Acad Sci U S A. 2009;106:8754–8759. doi: 10.1073/pnas.0803955106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Winner B, Lie DC, Rockenstein E, Aigner R, Aigner L, Masliah E, et al. Human wild-type alpha-synuclein impairs neurogenesis. J Neuropathol Exp Neurol. 2004;63:1155–1166. doi: 10.1093/jnen/63.11.1155. [DOI] [PubMed] [Google Scholar]
- 5.Crews L, Mizuno H, Desplats P, Rockenstein E, Adame A, Patrick C, et al. Alpha-synuclein alters Notch-1 expression and neurogenesis in mouse embryonic stem cells and in the hippocampus of transgenic mice. J Neurosci Off J Soc Neurosci. 2008;28:4250–4260. doi: 10.1523/JNEUROSCI.0066-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Nuber S, Petrasch-Parwez E, Winner B, Winkler J, von Hörsten S, Schmidt T, et al. Neurodegeneration and motor dysfunction in a conditional model of Parkinson’s disease. J Neurosci Off J Soc Neurosci. 2008;28:2471–2484. doi: 10.1523/JNEUROSCI.3040-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Winner B, Rockenstein E, Lie DC, Aigner R, Mante M, Bogdahn U, et al. Mutant alpha-synuclein exacerbates age-related decrease of neurogenesis. Neurobiol Aging. 2008;29:913–925. doi: 10.1016/j.neurobiolaging.2006.12.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Marxreiter F, Nuber S, Kandasamy M, Klucken J, Aigner R, Burgmayer R, et al. Changes in adult olfactory bulb neurogenesis in mice expressing the A30P mutant form of alpha-synuclein. Eur J Neurosci. 2009;29:879–890. doi: 10.1111/j.1460-9568.2009.06641.x. [DOI] [PubMed] [Google Scholar]
- 9.Winner B, Melrose HL, Zhao C, Hinkle KM, Yue M, Kent C, et al. Adult neurogenesis and neurite outgrowth are impaired in LRRK2 G2019S mice. Neurobiol Dis. 2011;41:706–716. doi: 10.1016/j.nbd.2010.12.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Baker SA, Baker KA, Hagg T. Dopaminergic nigrostriatal projections regulate neural precursor proliferation in the adult mouse subventricular zone. Eur J Neurosci. 202004:575–579. doi: 10.1111/j.1460-9568.2004.03486.x. [DOI] [PubMed] [Google Scholar]
- 11.Winner B, Geyer M, Couillard-Despres S, Aigner R, Bogdahn U, Aigner L, et al. Striatal deafferentation increases dopaminergic neurogenesis in the adult olfactory bulb. Exp Neurol. 2006;197:113–121. doi: 10.1016/j.expneurol.2005.08.028. [DOI] [PubMed] [Google Scholar]
- 12.Peng J, Xie L, Jin K, Greenberg DA, Andersen JK. Fibroblast growth factor 2 enhances striatal and nigral neurogenesis in the acute 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine model of Parkinson’s disease. Neuroscience. 2008;153:664–670. doi: 10.1016/j.neuroscience.2008.02.063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Arias-Carrión O, Yamada E, Freundlieb N, Djufri M, Maurer L, Hermanns G, et al. Neurogenesis in substantia nigra of parkinsonian brains? J Neural Transm Suppl. 2009:279–285. doi: 10.1007/978-3-211-92660-4_23. [DOI] [PubMed] [Google Scholar]
- 14.Zhao M, Momma S, Delfani K, Carlen M, Cassidy RM, Johansson CB, et al. Evidence for neurogenesis in the adult mammalian substantia nigra. Proc Natl Acad Sci U S A. 2003;100:7925–7930. doi: 10.1073/pnas.1131955100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Frielingsdorf H, Schwarz K, Brundin P, Mohapel P. No evidence for new dopaminergic neurons in the adult mammalian substantia nigra. Proc Natl Acad Sci U S A. 2004;101:10177–10182. doi: 10.1073/pnas.0401229101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Jackson CR, Ruan G-X, Aseem F, Abey J, Gamble K, Stanwood G, et al. Retinal dopamine mediates multiple dimensions of light-adapted vision. J Neurosci Off J Soc Neurosci. 2012;32:9359–9368. doi: 10.1523/JNEUROSCI.0711-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Battiste J, Helms AW, Kim EJ, Savage TK, Lagace DC, Mandyam CD, et al. Ascl1 defines sequentially generated lineage-restricted neuronal and oligodendrocyte precursor cells in the spinal cord. Dev Camb Engl. 2007;134:285–293. doi: 10.1242/dev.02727. [DOI] [PubMed] [Google Scholar]
- 18.Arnold K, Sarkar A, Yram MA, Polo JM, Bronson R, Sengupta S, et al. Sox2(+) adult stem and progenitor cells are important for tissue regeneration and survival of mice. Cell Stem Cell. 2011;9:317–329. doi: 10.1016/j.stem.2011.09.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Abercrombie M. Estimation of nuclear population from microtome sections. Anat Rec. 1946;94:239–247. doi: 10.1002/ar.1090940210. [DOI] [PubMed] [Google Scholar]
- 20.Suh H, Consiglio A, Ray J, Sawai T, D’Amour KA, Gage FH. In vivo fate analysis reveals the multipotent and self-renewal capacities of Sox2+ neural stem cells in the adult hippocampus. Cell Stem Cell. 2007;1:515–528. doi: 10.1016/j.stem.2007.09.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kim JB, Zaehres H, Wu G, Gentile L, Ko K, Sebastiano V, et al. Pluripotent stem cells induced from adult neural stem cells by reprogramming with two factors. Nature. 2008;454:646–650. doi: 10.1038/nature07061. [DOI] [PubMed] [Google Scholar]
- 22.Graham V, Khudyakov J, Ellis P, Pevny L. SOX2 functions to maintain neural progenitor identity. Neuron. 2003;39:749–765. doi: 10.1016/s0896-6273(03)00497-5. [DOI] [PubMed] [Google Scholar]
- 23.Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006;126:663–676. doi: 10.1016/j.cell.2006.07.024. [DOI] [PubMed] [Google Scholar]
- 24.Hsieh J. Orchestrating transcriptional control of adult neurogenesis. Genes Dev. 2012;26:1010–1021. doi: 10.1101/gad.187336.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Ekdahl CT, Claasen JH, Bonde S, Kokaia Z, Lindvall O. Inflammation is detrimental for neurogenesis in adult brain. Proc Natl Acad Sci. 2003;100:13632–13637. doi: 10.1073/pnas.2234031100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Monje ML, Toda H, Palmer TD. Inflammatory blockade restores adult hippocampal neurogenesis. Science. 2003;302:1760–1765. doi: 10.1126/science.1088417. [DOI] [PubMed] [Google Scholar]
- 27.Fujioka H, Akema T. Lipopolysaccharide acutely inhibits proliferation of neural precursor cells in the dentate gyrus in adult rats. Brain Res. 2010;1352:35–42. doi: 10.1016/j.brainres.2010.07.032. [DOI] [PubMed] [Google Scholar]
- 28.Frank-Cannon TC, Tran T, Ruhn KA, Martinez TN, Hong J, Marvin M, et al. Parkin deficiency increases vulnerability to inflammation-related nigral degeneration. J Neurosci Off J Soc Neurosci. 2008;28:10825–10834. doi: 10.1523/JNEUROSCI.3001-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Morrison BE, Marcondes MCG, Nomura DK, Sanchez-Alavez M, Sanchez-Gonzalez A, Saar I, et al. Cutting edge: IL-13Rα1 expression in dopaminergic neurons contributes to their oxidative stress-mediated loss following chronic peripheral treatment with lipopolysaccharide. J Immunol Baltim Md 1950. 2012;189:5498–5502. doi: 10.4049/jimmunol.1102150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Freed CR, Greene PE, Breeze RE, Tsai WY, DuMouchel W, Kao R, et al. Transplantation of embryonic dopamine neurons for severe Parkinson’s disease. N Engl J Med. 2001;344:710–719. doi: 10.1056/NEJM200103083441002. [DOI] [PubMed] [Google Scholar]
- 31.Olanow CW, Goetz CG, Kordower JH, Stoessl AJ, Sossi V, Brin MF, et al. A double-blind controlled trial of bilateral fetal nigral transplantation in Parkinson’s disease. Ann Neurol. 2003;54:403–414. doi: 10.1002/ana.10720. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.