

Abstract
Prolactinomas are the most frequently observed pituitary adenomas and most of them respond well to conventional treatment with dopamine agonists. However, a subset of prolactinomas fails to respond to such therapies and is considered as dopamine agonist-resistant prolactinomas (DARPs). New therapeutic approaches are necessary for these tumors. TGFβ1 is a known inhibitor of lactotroph cell proliferation and prolactin secretion, and it partly mediates dopamine inhibitory action. TGFβ1 is secreted to the extracellular matrix as an inactive latent complex, and its bioavailability is tightly regulated by different components of the ‘TGFβ1 system including latent binding proteins (LTBPs), local activators (Thrombospondin-1, matrix metalloproteases, integrins, among others), and TGFβ receptors. Pituitary TGFβ1 activity and the expression of different components of the TGFβ1 system, are regulated by dopamine and estradiol. Prolactinomas (animal models and humans) present reduced TGFβ1 activity as well as reduced expression of several components of the TGFβ1 system. Therefore, restoration of TGFβ1 inhibitory activity represents a novel therapeutic approach to bypass dopamine action in DARPs. The aim of this review is to summarize the large literature supporting TGFβ1 important role as a local modulator of pituitary lactotroph function; as well to provide recent evidence of the restoration of TGFβ1 activity as an effective treatment in experimental prolactinomas.
Keywords: resistant prolactinomas, dopamine, estradiol, TGFβ1
Pituitary tumors
Pituitary tumors are commonly benign, slow growing adenomas and account for 10–15% of all intracranial neoplasms (Farrell 2006; Melmed 2015). The prevalence of these tumors is relatively high in the general population, with approximately 77 cases per 100.000 (Daly et al. 2009; Fernandez et al. 2010), and studies of autopsy specimens identified up to a 20% prevalence of clinically occult pituitary adenomas (Ezzat et al. 2004).
Despite their benign features pituitary tumors can cause considerably morbidity due to both hypersecretion of pituitary trophic hormones and excessive tumor growth that can affect surrounding tissue. Common symptoms of a pituitary tumor compressive ‘mass effect’ include visual impairment, headaches, neurological disorders and hypopituitarism caused by disruption of the hypothalamic-pituitary axis (Arafah & Nasrallah 2001; Melmed 2011). Based on their size, pituitary adenomas are classified as microadenomas (< 10mm), macroadenomas (>10mm) or giant adenomas (>40mm).
Pituitary tumors usually present with monoclonal growth and can also be classified according to their cell type origin and hormone secretion. Thus, somatotropinomas secrete growth hormone (GH), prolactinomas secrete prolactin (PRL), thyrotropinomas secrete thyroid-stimulating hormone (TSH), and corticotropinomas secrete adrenocorticotropin hormone (ACTH). In contrast, the nonfunctioning pituitary adenomas do not produce any hormone and usually derive from gonadotropes (Kovacs et al. 2001; Syro et al. 2015).
Prolactinomas
Among functioning pituitary tumors, prolactinomas are the most frequently observed in the clinic (40%) (Ciccarelli et al. 2005). Excessive PRL secretion by these tumors leads to hyperprolactinemia, which primarily affects gonadal/reproductive function, causing hypogonadism, galactorrhea, decreased libido and infertility both in men and women. Large macroprolactinomas can also cause neurological symptoms due to compression of adjacent tissues.
Prolactinomas are usually benign, and although some tumors show invasion into the parasellar compartment and/or sphenoid sinuses, malignant transformation and metastatic spread are extremely rare. Macroprolactinomas tend to be more aggressive and resistant to therapies than microprolactinomas (Wong et al. 2015a).
Differences in prolactinoma incidence, tumor size and behavior have been described among genders. The prevalence of prolactinomas is higher in women during the fertile period (20–50 years), while the frequency is similar between sexes after the fifth decade of life (Colao et al. 2003; Gillam et al. 2006). Also, women usually present with microprolactinomas whereas men more often present with macroprolactinomas ( Delgrange et al. 1997; Nishioka et al. 2003). These differences have been associated to the earlier diagnosis in woman due to the readily detection of symptoms caused by high prolactin (amenorrea/galactorrea) (Delgrange et al. 1997;Colao et al. 2003; Nishioka et al. 2003; Gillam et al. 2006). However, delayed diagnosis in men may not be the only explanation for the differences in tumor size, since young men also present with macroprolactinomas, and prolactinomas in men tend to be more aggressive, with higher proliferative indexes and lower rates of surgical cure, suggesting a sex-specific behavior for these tumors (Delgrange et al. 1997; Gillam MP & Molitch ME 2015).
Prolactinoma treatment
The major goals of treatment in patients with prolactinomas are to normalize serum PRL levels, to restore gonadal function, to reduce tumor size and to preserve or improve residual pituitary function. Prolactin secretion in the normal pituitary is tonically inhibited by hypothalamic dopamine through dopamine D2 receptors (Drd2) expressed on lactotroph cell membranes (Ben Jonathan & Hnasko 2001). The majority of prolactinomas retain an intact response to dopamine inhibition, therefore medical treatment with dopamine agonists (DAs), such as cabergolina and bromocriptine, represents the first line therapy for this tumors, including microprolactinomas, macroprolactinomas and giant prolactinomas (Wong et al. 2015b). Dopamine agonists are highly effective in achieving therapeutic aims with a favorable benefit/risk balance compared with surgical treatment.
Dopamine agonist-resistant prolactinomas (DARPs)
Despite the universal use of dopamine agonists and their high efficiency in reducing PRL levels and decreasing tumor size, there is a subset of prolactinomas (10–15%) that do not respond appropriately to the treatment, even at high doses of DA (Vroonen et al. 2012). These tumors represent a major challenge for clinical management. DARPs are more prevalent in men than woman, occur most frequently as macroprolactinomas and tend to be invasive, exhibiting extension to the cavernous sinuses.
The molecular mechanisms underlying the escape from dopaminergic regulation in DARPS are not fully elucidated. The main candidate thought to be responsible for resistance is the Drd2 itself. However, to date, no point mutation in the Drd2 gene has been identified in DARPs (Friedman et al. 1994; Molitch 2003; Gillam et al. 2006; Vroonen et al. 2012; Molitch 2014). Nevertheless, several mechanisms that lead to reduced Drd2 sensitivity were described in resistant prolactinomas, including evidence of decreased Drd2 mRNA expression, and differential expression of short and long Drd2 isoforms (Caccavelli et al. 1994; Vasilev et al. 2011; Shimazu et al. 2012); reduced Drd2 density and reduced dopamine binding sites in plasma membranes of DARP cells (Pellegrini et al. 1989). Alterations in dopamine signaling, such as decreased expression of the inhibitory alpha G protein subunit (Gαi2) have also been described (Caccavelli et al. 1996); as well as decreased expression of the nerve growth factor receptor (NGFR), which indirectly modulates Drd2 expression (Passos et al. 2009). Histological studies on DARPs also revealed increased angiogenesis, cellular atypia (multinucleated cells, irregular nuclei) and increased proliferation index measured by Ki67 staining, indicating an overall increase invasiveness (reviewed in (Gurlek et al. 2007)).
Alternative treatments for DARPs
At present, there is no alternative medical treatment for DARPs, and transsphenoidal surgery is indicated if the tumor is still resectable (Primeau et al. 2012; Smith et al. 2015). However, some aggressive prolactinomas recur post-operatively and show progressive growth, in which case, radiotherapy is the next therapeutic option, but with limited efficiency (Molitch 2014).
The chemotherapy agent temozolomide (TMZ) has been recently used as a last resort therapy and showed a moderately successful response in large aggressive DARPs (Whitelaw et al. 2012; Liu et al. 2015; McCormack et al. 2011). However the efficacy of TMZ therapy in aggressive pituitary adenomas remains controversial (Bruno et al. 2015) and clinical trials are now necessary to establish the indications, doses, and duration of TMZ administration to more accurately determine the efficacy of this agent.
New therapeutic approaches are necessary for those prolactinomas that are resistant to conventional treatments. Few reports of experimental treatments can be found in the literature and show variable effectiveness in pre-clinical and in vitro models. For instance, treatment with somatostatin receptor (SSTR) analogs failed to inhibit prolactin secretion by cultured cells derived from DARPs (Fusco et al. 2008) despite the expression of all subtypes of SSTR in human prolactinomas (Jaquet et al. 1999).
Based on the counteracting effects of estradiol on dopamine action in lactotrophs, targeting of the estrogen receptor with tamoxifen was evaluated in the pre-cabergoline era in patients with bromocriptine-resistant prolactinomas, but only a moderated reduction in PRL levels was observed (Volker et al. 1982). A novel anti-estrogen agent, fulvestrant, also reduced PRL secretion in pituitary cell lines, and decreased tumor growth and serum PRL in estrogen-induced prolactinomas in rats (Cao et al. 2014).
In the search for new therapeutic targets for DARPs
Studies in animal models of prolactinomas with altered sensitivity to DA, such as the estrogen induced prolactinomas in rats and the Drd2 knock-out mice (Drd2−/−) have been very helpful to identify molecular pathways altered in these tumors and to test potential future therapies. Many of these studies suggest that the deregulation of local growth factors and extracellular matrix remodeling participate in the pathogenesis of prolactinomas by promoting cell proliferation, angiogenesis and invasiveness (Paez-Pereda et al. 2005; Cristina et al. 2005; Cristina et al. 2007; Recouvreux et al. 2013).
Transforming growth factor beta 1 (TGFβ1), a well-known inhibitor in lactotroph physiology, has been recently identified as a novel target for the development of new therapies in resistant prolactinomas.
The complexity of the TGFβ system and biology
TGFβs are multifunctional cytokines known to play crucial regulatory roles in cellular proliferation and differentiation, angiogenesis, extracellular matrix modification and immunomodulation (Yoshinaga et al. 2008), and have powerful effects on embryogenesis, development and tissue homeostasis (Heldin et al. 2009; Galvin-Burgess et al. 2013; Itoh et al. 2014). The TGFβ family comprises more than 30 highly pleiotropic molecules including activins, inhibins, nodal, bone morphogenetic proteins (BMPs), the anti-muellerian hormone (AMH), and several growth and differentiation factors (GDFs) among others (Derynck & Akhurst 2007). Three isoforms of TGFβ have been identified (TGFβ1, 2, and 3).
The importance of TGFβ1 is clearly demonstrated by the fact that TGFβ1 null mutation causes excessive inflammatory response and early death (Kulkarni et al. 1993). On the contrary, an excess of TGFβ1 activity is associated to connective tissue diseases, fibrosis and inflammation, cirrhosis, arthritis and sclerosis, cardiovascular diseases and cancer, making TGFβ an interesting target for therapeutic research (Pohlers et al. 2009; Doyle et al. 2012; Akhurst & Hata 2012).
Nearly all cell types are sensitive to TGFβ1, but TGFβ action is highly dependent on cell type, developmental stage, physiological-pathological conditions, interaction with components of the extracellular matrix and, once bond to its receptor, interaction with other signaling pathways.
The potent biological activity of TGFβ1 is tightly regulated at different levels, including its synthesis, secretion, storage and activation. The three TGFβ isoforms are synthesized as homodimeric precursor molecules that contain a pro-peptide sequence, called latency associated peptide (LAP), and the functional mature TGFβ sequence (Figure 1. 1). After proteolytic processing by furin within the trans-golgi, LAP remains associated with the mature TGFβ by non-covalent interactions in a small latent complex (Figure 1. 2). While in the endoplasmic reticulum, LAP is linked, by disulfide bonds, with a latent TGFβ binding protein (LTBP) (Figure 1. 3). LTBPs belong to a family of large secretory extracellular matrix (ECM) glycoproteins. Although LTBPs are not required for maintenance of TGFβ latency, they facilitate the secretion, storage, and activation of the TGFβ–LAP complex (Rifkin 2005).
Figure 1. The biology of TGFβ system.
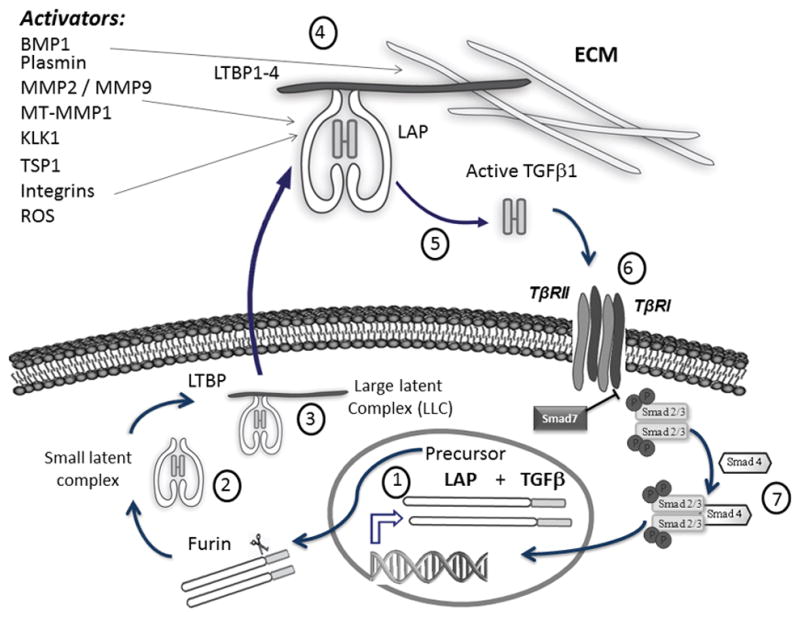
TGFβ is synthesized as homodimeric precursor containing a pro-peptide sequence LAP, (1), and then processed by furin. LAP remains associated with the mature TGFβ by non-covalent interactions in a small latent complex (2), which in turn is linked by disulfide bonds to one of the latent TGFβ binding proteins (LTBP1–4) (3). TGFβ is secreted as part of this large latent TGFβ complex (LLC) (3), and it is incorporated as component of the extracellular matrix (ECM), which acts as a reservoir of the cytokine (4). TGFβ must undergo a highly regulated activation process by which mature cytokine is released (5) to enable binding to its receptor complex (TβRI and TβRII) (6) and signal through Smad2/Smad3 pathway (7). Known TGFβ activators are listed in the upper left.
TGFβ is secreted as part of this large latent TGFβ complex (LLC), (Figure 1. 3), and is incorporated as component of the ECM, which acts as a cytokine reservoir (Figure 1. 4). Trapped in the ECM, TGFβ remains latent because of persistent binding of LAP and must undergo a highly regulated activation process by which mature cytokine is released from its latent complex to enable the active form to bind and signal through its receptor (Figure 1. 5). Latent TGFβ activation is a crucial event in governing the cytokine biological function and availability in the ECM (Annes et al. 2003; Annes et al. 2004; Rifkin 2005).
Several latent TGFβ1 activators have been described, including proteases (such as plasmin, matrix metalloproteinase 2 (MMP2), matrix metalloproteinase 9 (MMP9), BMP-1, thrombospondin-1 (TSP-1), kallikrein 1 (KLK1), integrins αvβ6 and αvβ8, and reactive oxygen species (ROS) or pH changes in the local environment, among others. However, their individual biological importance in releasing TGFβ1 from its latent complex and their local regulation in different tissues are not fully understood (Annes et al. 2003; Annes et al. 2004; Yoshinaga et al. 2008). Since all these factors are related to ECM perturbations, the latent TGFβ complex has been postulated as a “sensor” of environment disturbances (Annes et al. 2003).
Once TGFβ1 is released from the ECM, the active cytokine binds to its transmembrane receptor, the type II TGFβ receptor (TβRII), a constitutively active kinase that recruits and phosphorylates TβRI (type I TGFβ receptor) forming a heterotetrameric complex of serine/threonine kinase receptors containing two type I and two type II subunits (Figure 1. 6). Next, TβRI phosphorylates the downstream receptor-associated Smads (R-Smads: Smad2/Smad3), which form a heteromeric complex with Smad4, and translocate to the nucleus to regulate the transcription of target genes (Figure 1. 7). Additionally, an inhibitory Smad, Smad7, competes with the Smad2/3 for binding to the activated TβRI, thereby exerting a negative effect on TGFβ/Smad signaling (Shi & Massague 2003; Han et al. 2015).
This pathway is known as “the canonical” TGFβ signaling pathway. However, TGFβ can also signal through smad-independent pathways, including the mitogen-activated protein kinases (ERK1/2, JNK, p38), small GTP-binding proteins (Ras, RhoA, Rac1, CDC42, mTOR), the NF-κB pathway and Wnt/β-catenin pathway, the AKT/PKB pathway and phosphatidylinositol-4,5-bisphosphate 3-kinase (PI3K) ( Attisano & Wrana 2002; Derynck & Zhang 2003; Moustakas & Heldin 2005).
As a multifunctional cytokine with powerful effects on cell proliferation, cellular migration and inflammation, TGFβ signaling has been targeted for drug development and numerous strategies have proceeded through preclinical to clinical trials (reviewed in (Akhurst & Hata 2012)).
TGFβ1 in the pituitary: a brief history
The earliest publications on TGFβ1 action in the pituitary date from the late 80s and early 90s (Ying et al. 1986; Mueller & Kudlow 1991). Dr. Sarkar and col were the first to demonstrate local TGFβ1 mRNA and protein expression in the pituitary gland, and the inhibitory action of TGFβ1 on prolactin secretion and lactotrophic growth in 1992 (Sarkar et al. 1992). Although these first evidences were found in animal models (rat), TGFβ1 and TβRII expression were promptly found to be expressed in human pituitaries (Halper et al. 1992; Fujiwara et al. 1995), as well as in human pituitary adenomas (Fujiwara et al. 1995; Jin et al. 1997).
The main physiological modulators of lactotroph function are dopamine and estradiol, which exert inhibitory and stimulatory actions, respectively (Ben Jonathan & Hnasko 2001). The pro-mitotic effect of estradiol (pharmacological doses) and its role in prolactinoma induction is very well described in the literature (Heaney et al. 2002). However, estrogens also participate in the lactotroph cell turnover in normal pituitary glands, sensitizing lactotroph cells to apoptotic stimuli. Therefore the effect of estradiol on lactotroph function depends on the dose and normal/tumoral condition of the cells (Pisera et al. 2004; Zaldivar et al. 2009; Jaita et al. 2015). Interestingly, dopamine and estradiol also regulate the expression of both TGFβ1 and its receptor, but in opposite ways. Thus, while estrogen stimulation increases serum PRL levels and lactotroph proliferation, it decreases the expression of TGFβ1 in the anterior pituitary. On the contrary, dopamine, acting through the Drd2, up-regulates TGFβ1 expression and secretion in vivo and in vitro, with a concomitant reduction in the proliferation rate of lactotrophs. Moreover, it has been proposed that TGFβ1 partially mediates the inhibitory effect of dopamine on lactotroph proliferation (Sarkar et al. 2005). Our group has recently described that the amount of pituitary active TGFβ1 is also locally regulated by dopamine and estradiol treatment in mice, and moreover we found an inverse correlation between active TGFβ1 levels and serum PRL (Recouvreux et al. 2011). It is worth noting that less than 8% of total pituitary TGFβ1 was found in the active form, similar to what has been described in other tissues (Yoshinaga et al. 2008). This underscores the tightly regulation of the latent TGFβ activation process. Other important factor regulating lactotroph homeostasis is PRL itself, acting through the PRL receptor (PRLR). It has been shown that endogenous PRL exerts paracrine/autocrine antiproliferative and proapoptotic effects on lactotrophs, and moreover, knock-out mice lacking PRLR develop prolactinomas, further demonstrating the important role of PRL in the negative feedback on lactotroph function (Schuff et al. 2002; Ferraris et al. 2012; Ferraris et al. 2014).
Whether PRL can as well regulate TGFβ1 expression or function in the pituitary gland is an open question that has not yet been addressed.
Alterations in the components of the TGFβ1 system during prolactinoma development
Evidences of TGFβ1 alterations in estradiol-induced prolactinomas in rats
The estrogen-treated rat is a well-known model for prolactinoma development with increased pituitary weight, hyperprolactinemia, lactotroph hyperplasia and reduced dopaminergic action at the pituitary level (Heaney et al. 1999; Heaney et al. 2002b). Furthermore, estradiol treatment decreases pituitary TGFβ1 and TβRII mRNA and protein, together with an increase in PRL levels (Sarkar et al. 1992; Pastorcic et al. 1995; De et al. 1996) (Figure 2). Therefore, the inhibition of TGFβ1 and TβRII might cooperate in the development of prolactinomas induced by estradiol (Hentges & Sarkar 2001). In agreement with this idea, pituitary tumorigenesis induced by estrogen treatment is greatly accelerated in TβRII heterozygous knockout mice (TβRII+/−) where the expression of TβRII is markedly reduced (Shida et al. 1998).
Figure 2. Alterations of TGFβ1 system in prolactinomas.
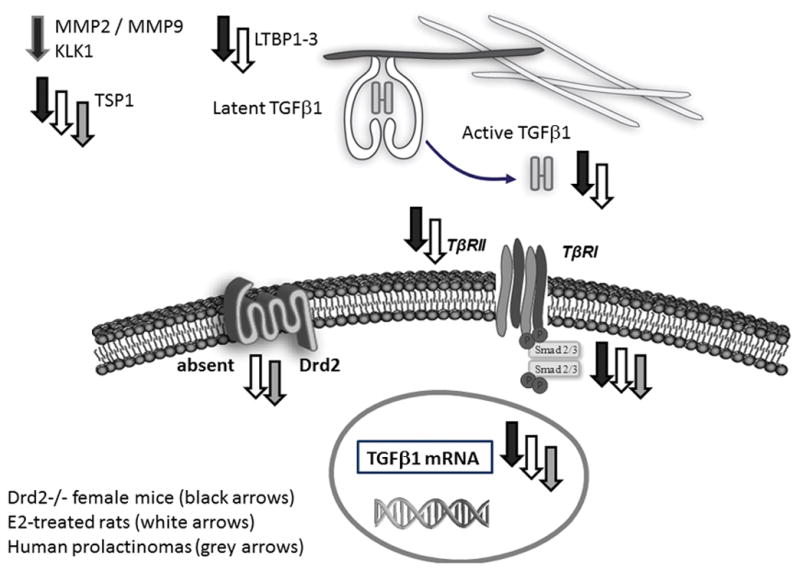
Decreased expression of different components of pituitary TGFβ1 system in prolactinoma models are represented with down-pointed arrows. Gray arrows indicate findings in human prolactinoma specimens; black arrows indicate findings in Drd2−/− mice; white arrows indicate findings in estradiol induced prolactinomas in rats.
TGFβ1 alterations in the prolactinoma development in Drd2 −/− mice
Another well characterized model to study prolactinoma development are the transgenic knock out mice lacking functional Drd2 (Drd2−/−). This model represents an excellent model to mimick dopamine agonist-resistance. Because of the absence of inhibitory dopaminergic control, these mice display chronic hyperprolactinemia and lactotroph hyperplasia (Kelly et al. 1997; Diaz-Torga et al. 2002; Cristina et al. 2006), but the loss of dopamine inhibition has deeper effect on pituitary function in female than in male mice (Saiardi et al. 1997; Diaz-Torga et al. 2002). In females the increase in serum prolactin levels is much more pronounced than in males, and females develop lactotroph hyperplasia from 6 month onwards, while age-matched Drd2-deficient males develop pituitary lactotroph adenomas at 17 to 20 months of age (Asa et al. 1999).
Interestingly, active and total TGFβ1 levels, as well as TβRII and LTBP1 expression are reduced in Drd2−/− pituitaries compared to controls (wild type or wt), highlighting the stimulatory role of dopamine on pituitary TGFβ1 system (Recouvreux et al. 2011) (Figure 2). On the other hand, the impact of the chronic loss of dopaminergic tone on the TGFβ1 system was also stronger in females, evidenced by the downregulation of several putative TGFβ1 activators (MMP2, MMP9, MT1-MMP, thrombospondin-1 and kallikrein) as well as the decreased expression of TGFβ1 target genes observed only in females (Drd2−/− vs. their wt counterpart). In this model we found sex differences in the regulation of the TGFβ1 system: males express higher levels of several components of the TGFβ1 system, and it could be due to the lower serum estradiol levels present in males, as estradiol negatively controls most of the components of the system (Recouvreux et al. 2013). We suggest that stronger pituitary TGFβ1 system could protect males from excessive lactotroph proliferation and prolactinoma development. Then, sex differences found in the regulation of the TGFβ1 system could explain sex differences found in the incidence of prolactinoma development in this model.
TGFβ1 alterations in human pituitary tumors
In humans the expression of several components of the TGFβ signaling pathway was recently compared in five normal human anterior pituitaries, 29 invasive nonfunctioning pituitary adenomas (NFPAs) and 21 noninvasive NFPAs (Zhenye et al. 2014). This report demonstrated that TGFβ1 mRNA expression and p-Smad3 protein levels gradually decreased, while Smad7 mRNA levels gradually increased from normal anterior pituitaries to noninvasive NFPAs and invasive NFPAs. The authors concluded that the activity of TGFβ signaling would be limited during tumor development.
Recent work also described a significant down-regulation of the TGFβ1/Smad signaling cascade in 12 cases of DARPs compared to normal human anterior pituitaries. The authors showed that TGFβ1 mRNA levels, and Smad2 and Smad3 mRNA and protein expression were significantly decreased in human prolactinomas (Li et al. 2015) (Figure 2).
Overall, decreased TGFβ1 activity and decreased expression of different components of the TGFβ1 system have been described in animal models of prolactinomas as well as in human prolactinomas. Taking into account that TGFβ1 inhibits lactotroph proliferation and PRL synthesis and secretion, we speculate that recovering local TGFβ1 activity could contribute to revert the adenoma development and to normalize prolactinemia.
Recovery of local TGFβ1 activity. Successful treatment in an experimental model of prolactinoma
Thrombospondin-1 (TSP1) is one of the main physiologic latent TGFβ1 activators in vitro and in vivo (Schultz-Cherry et al. 1994). TSP1 is a large multifunctional matrix glycoprotein involved in cell growth, adhesion, migration (Lawler 2002). TSP1 also functions as an endogenous antiangiogenic factor, inhibiting the proliferation and migration of endothelial cells by interaction with its cell surface receptor CD36 and by antagonizing VEGF activity (Lawler & Lawler 2012).
Based on the CD36-binding peptide sequence from TSP-1, small-molecules were developed to mimic TSP-1 antiangiogenic properties (Haviv et al. 2005). Several of these new drugs were able to slow tumor growth in preclinical models (Anderson et al. 2007; Yang et al. 2007; Garside et al. 2010). Among them, ABT-510 and ABT-898 (Abbott Laboratories), two of such TSP1 analogs, were assayed in several solid tumors (Haviv et al. 2005). ABT-510 was evaluated in phase II clinical trials for the treatment of head and neck cancer, non-small cell lung cancer, lymphoma, and renal cell carcinoma (Haviv et al. 2005; Ebbinghaus et al. 2007; Markovic et al. 2007; Yang et al. 2007; Gordon et al. 2008; Nabors et al. 2010). The second-generation TSP-1 synthetic analogue, ABT-898, was found to have greater potency than ABT-510 and is expected to have greater efficacy than the other available TSP-1-mimetic peptides (Garside et al. 2010; Campbell et al. 2011) due to its lower clearance rate.
Immunoreactive TSP-1 is present in the anterior pituitary, particularly in endothelial cells (Burns & Sarkar 1993), and TSP-1 levels and its antiangiogenic effect are reduced in prolactinomas induced by estradiol in rats (Sarkar et al. 2007) and in the hyperplastic pituitaries of Drd2−/− mice (Recouvreux et al. 2013). TSP-1 expression was also found down-regulated in invasive vs. non-invasive prolactinomas in humans (Jiang et al. 2012).
Given that: 1) TSP1 is an antiangiogenic factor, 2) TSP1 expression is reduced during prolactinoma development, 3) TSP1 is a known TGFβ1 activator, 4) TGFβ1 activity is also reduced during the development of prolactinomas, and 5) TGFβ1 is an inhibitory factor of lactotroph proliferation and synthesis; we speculated that treatments that improve pituitary TSP-1 and/or TGFβ1 activities could reduce the progression of prolactinomas.
We first evaluated whether the TSP1 analogs were able to activate TGFβ1 in the pituitary. In fact, an in vivo short term treatment (100 mg/kg ABT-510, ip, 30 min) enhanced the biological activity of pituitary TGFβ1, with a concomitant reduction in serum prolactin levels (Recouvreux et al. 2012). Notably, same effect was observed after short term treatment with ABT-510 in female rats carrying prolactinoma induced by chronic estradiol treatment. We next evaluated whether an in vivo treatment for two weeks with the TSP1 analogs could counteract the development of estradiol-induced prolactinomas in rats. ABT-510 and ABT-898 treatment (100mg/kg ip, thrice a week for two weeks) significantly decreased pituitary tumor size, reduced tissue angiogenesis and pituitary proliferation markers, as well as serum prolactin levels, in female rats with prolactinomas induced by chronic treatment with estradiol (Recouvreux et al. 2012). Furthermore, ABT-510 and ABT-898 treatment markedly increased active TGFβ1 content, measured by ELISA within the tumors. The increase in cytokine activation was also reflected in the recovery of intrapituitary pSMAD2/3 expression (Figure 3). Besides from the well-known antiangiogenic effect of these TSP-1 mimetic peptides, the improvement of the local TGFβ1 biological activity, most likely contributed to the reduction in serum prolactin and in the inhibition of prolactinoma growth.
Figure 3. Recovery of TGFβ1 activity emerges as a novel therapeutic target for treatment of dopamine agonist resistant prolactinomas.
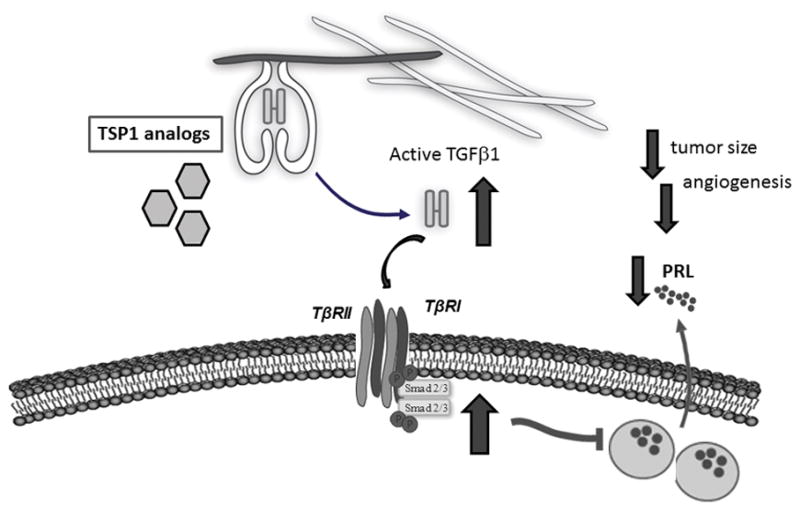
Treatment with Thrombospondin-1 analogs (ABT-510, ABT-898) recover pituitary TGFβ1 activity, reduce tumor size, tumor angiogenesis and proliferation markers, as well as serum
Overall conclusions and Perspectives
Prolactinomas are the most frequent pituitary tumors in adults accounting for 60% of all functioning pituitary tumors (Ciccarelli et al. 2005). Even though prolactinomas are usually benign and in most cases respond well to treatment with dopaminergic agents, 15% of these tumors are resistant to classical therapy, become invasive and aggressive, and require extirpation. The mechanisms underlying the escape from dopaminergic regulation in DARPS are not fully understood, and the main candidate to be responsible for resistance is the Drd2 itself. Since TGFβ1 mediates, at least partially, the inhibitory action exerted by dopamine on lactotrophs, and reduced TGFβ1 activity is a common feature of prolactinoma development, treatments that improve pituitary TGFβ1 activity represent a rational approach to develop alternative therapies for DARPs. Supporting this, we provide evidence of the effectiveness of a treatment with the small TSP1-analog peptides ABT-510 and ABT-898 to restore TGFβ1 activity and to counteract prolactinoma development in rats.
Taken together the data summarized here, the recovery of TGFβ1 activity emerges as a novel therapeutic target for treatment of dopamine agonist resistant prolactinomas.
Acknowledgments
We thank the National Institute of Diabetes and Digestive and Kidney Diseases National Hormone and Pituitary Program and Dr A. F. Parlow for prolactin RIA kits.
FUNDING: This work was supported by the Agencia Nacional de Promoción Científica y Técnica, Buenos Aires, Argentina (grant PICT 2013–2016 N2136 to G.D.T.), and the National Institutes of Health (grant R01 CA034282–25 to D.B.R.).
Footnotes
Disclosure Summary: The authors have nothing to disclose. There is no conflict of interest that could be perceived as prejudicing the impartiality of the research reported.
Reference List
- Akhurst RJ, Hata A. Targeting the TGFbeta signalling pathway in disease. Nat Rev Drug Discov. 2012;11:790–811. doi: 10.1038/nrd3810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson JC, Grammer JR, Wang W, Nabors LB, Henkin J, Stewart JE, Jr, Gladson CL. ABT-510, a modified type 1 repeat peptide of thrombospondin, inhibits malignant glioma growth in vivo by inhibiting angiogenesis. Cancer Biol Ther. 2007;6:454–462. doi: 10.4161/cbt.6.3.3630. [DOI] [PubMed] [Google Scholar]
- Annes JP, Chen Y, Munger JS, Rifkin DB. Integrin alphaVbeta6-mediated activation of latent TGF-beta requires the latent TGF-beta binding protein-1. J Cell Biol. 2004;165:723–734. doi: 10.1083/jcb.200312172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Annes JP, Munger JS, Rifkin DB. Making sense of latent TGFbeta activation. J Cell Sci. 2003;116:217–224. doi: 10.1242/jcs.00229. [DOI] [PubMed] [Google Scholar]
- Arafah B, Nasrallah M. Pituitary tumors: pathophysiology, clinical manifestations and management. Endocr Relat Cancer. 2001;8:287–305. doi: 10.1677/erc.0.0080287. [DOI] [PubMed] [Google Scholar]
- Asa SL, Kelly MA, Grandy DK, Low MJ. Pituitary lactotroph adenomas develop after prolonged lactotroph hyperplasia in dopamine D2 receptor-deficient mice. Endocrinology. 1999;140:5348–5355. doi: 10.1210/endo.140.11.7118. [DOI] [PubMed] [Google Scholar]
- Attisano L, Wrana JL. Signal transduction by the TGF-beta superfamily. Science. 2002;296:1646–1647. doi: 10.1126/science.1071809. [DOI] [PubMed] [Google Scholar]
- Ben Jonathan N, Hnasko R. Dopamine as a prolactin (PRL) inhibitor. Endocrine reviews. 2001;22:724–763. doi: 10.1210/edrv.22.6.0451. [DOI] [PubMed] [Google Scholar]
- Bruno OD, Juarez-Allen L, Christiansen SB, Manavela M, Danilowicz K, Vigovich C, Gomez RM. Temozolomide Therapy for Aggressive Pituitary Tumors: Results in a Small Series of Patients from Argentina. Int J Endocrinol. 2015;2015:587893. doi: 10.1155/2015/587893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burns G, Sarkar DK. Transforming growth factor-beta1-like immunoreactivity in the pituitary gland of the rat: effect of estrogen. Endocrinology. 1993;133:1444–1449. doi: 10.1210/endo.133.3.8365375. [DOI] [PubMed] [Google Scholar]
- Caccavelli L, Feron F, Morange I, Rouer E, Benarous R, Dewailly D, Jaquet P, Kordon C, Enjalbert A. Decreased Expression of the Two D2 Dopamine Receptor Isoforms in Bromocriptine-Resistant Prolactinomas. Neuroendocrinology. 1994;60:314–322. doi: 10.1159/000126764. [DOI] [PubMed] [Google Scholar]
- Caccavelli L, Morange-Ramos I, Kordon C, Jaquet P, Enjalbert A. Alteration of G alpha subunits mRNA levels in bromocriptine resistant prolactinomas. J Neuroendocrinol. 1996;8:737–746. doi: 10.1046/j.1365-2826.1996.04902.x. [DOI] [PubMed] [Google Scholar]
- Campbell N, Greenaway J, Henkin J, Petrik J. ABT-898 Induces Tumor Regression and Prolongs Survival in a Mouse Model of Epithelial Ovarian Cancer. Mol Cancer Ther. 2011;10:1876–1885. doi: 10.1158/1535-7163.MCT-11-0402. [DOI] [PubMed] [Google Scholar]
- Cao L, Gao H, Gui S, Bai G, Lu R, Wang F, Zhang Y. Effects of the estrogen receptor antagonist fulvestrant on F344 rat prolactinoma models. J Neurooncol. 2014;116:523–531. doi: 10.1007/s11060-013-1351-8. [DOI] [PubMed] [Google Scholar]
- Ciccarelli A, Daly AF, Beckers A. The epidemiology of prolactinomas. Pituitary. 2005;8:3–6. doi: 10.1007/s11102-005-5079-0. [DOI] [PubMed] [Google Scholar]
- Colao A, Sarno AD, Cappabianca P, Briganti F, Pivonello R, Somma CD, Faggiano A, Biondi B, Lombardi G. Gender differences in the prevalence, clinical features and response to cabergoline in hyperprolactinemia. European Journal of Endocrinology. 2003;148:325–331. doi: 10.1530/eje.0.1480325. [DOI] [PubMed] [Google Scholar]
- Cristina C, Diaz-Torga G, Baldi A, Gongora A, Rubinstein M, Low MJ, Becu-Villalobos D. Increased pituitary vascular endothelial growth factor-A in dopaminergic D2 receptor knockout female mice. Endocrinology. 2005;146:2952–2962. doi: 10.1210/en.2004-1445. [DOI] [PubMed] [Google Scholar]
- Cristina C, Diaz-Torga G, Gongora A, Guida MC, Perez-Millan MI, Baldi A, Becu-Villalobos D. Fibroblast Growth Factor-2 in hyperplastic pituitaries of D2R knockout female mice. American Journal of Physiology, Endocrinology and Metabolism. 2007;293:E1341–E1351. doi: 10.1152/ajpendo.00260.2007. [DOI] [PubMed] [Google Scholar]
- Cristina C, García-Tornadú I, Diaz-Torga G, Rubinstein M, Low MJ, Becu-Villalobos D. The dopaminergic D2 receptor knockout mouse: an animal model of prolactinoma. Frontiers of Hormone Research. 2006;35:50–63. doi: 10.1159/000094308. [DOI] [PubMed] [Google Scholar]
- Daly AF, Tichomirowa MA, Beckers A. The epidemiology and genetics of pituitary adenomas. Best Pract Res Clin Endocrinol Metab. 2009;23:543–554. doi: 10.1016/j.beem.2009.05.008. [DOI] [PubMed] [Google Scholar]
- De A, Morgan TE, Speth RC, Boyadjieva N, Sarkar DK. Pituitary lactotrope expresses transforming growth factor beta (TGF beta) type II receptor mRNA and protein and contains 125I-TGF beta 1 binding sites. Journal of Endocrinology. 1996;149:19–27. doi: 10.1677/joe.0.1490019. [DOI] [PubMed] [Google Scholar]
- Delgrange E, Trouillas J, Maiter D, Donckier J, Tourniaire J. Sex-related difference in the growth of prolactinomas: a clinical and proliferation marker study. Journal of Clinical Endocrinology and Metabolism. 1997;82:2102–2107. doi: 10.1210/jcem.82.7.4088. [DOI] [PubMed] [Google Scholar]
- Derynck R, Akhurst RJ. Differentiation plasticity regulated by TGF-beta family proteins in development and disease. Nat Cell Biol. 2007;9:1000–1004. doi: 10.1038/ncb434. [DOI] [PubMed] [Google Scholar]
- Derynck R, Zhang YE. Smad-dependent and Smad-independent pathways in TGF-beta family signalling. Nature. 2003;425:577–584. doi: 10.1038/nature02006. [DOI] [PubMed] [Google Scholar]
- Diaz-Torga G, Feierstein C, Libertun C, Gelman D, Kelly MA, Low MJ, Rubinstein M, Becu-Villalobos D. Disruption of the D2 dopamine receptor alters GH and IGF-I secretion and causes dwarfism in male mice. Endocrinology. 2002;143:1270–1279. doi: 10.1210/endo.143.4.8750. [DOI] [PubMed] [Google Scholar]
- Doyle JJ, Gerber EE, Dietz HC. Matrix-dependent perturbation of TGFbeta signaling and disease. FEBS Letters. 2012;586:2003–2015. doi: 10.1016/j.febslet.2012.05.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ebbinghaus S, Hussain M, Tannir N, Gordon M, Desai AA, Knight RA, Humerickhouse RA, Qian J, Gordon GB, Figlin R. Phase 2 study of ABT-510 in patients with previously untreated advanced renal cell carcinoma. Clinical Cancer Research. 2007;13:6689–6695. doi: 10.1158/1078-0432.CCR-07-1477. [DOI] [PubMed] [Google Scholar]
- Ezzat S, Asa SL, Couldwell WT, Barr CE, Dodge WE, Vance ML, McCutcheon IE. The prevalence of pituitary adenomas: a systematic review. Cancer. 2004;101:613–619. doi: 10.1002/cncr.20412. [DOI] [PubMed] [Google Scholar]
- Farrell WE. Pituitary tumours: findings from whole genome analyses. Endocr Relat Cancer. 2006;13:707–716. doi: 10.1677/erc.1.01131. [DOI] [PubMed] [Google Scholar]
- Fernandez A, Karavitaki N, Wass JA. Prevalence of pituitary adenomas: a community-based, cross-sectional study in Banbury (Oxfordshire, UK) Clinical Endocrinology (Oxford) 2010;72:377–382. doi: 10.1111/j.1365-2265.2009.03667.x. [DOI] [PubMed] [Google Scholar]
- Ferraris J, Boutillon F, Bernadet M, Seilicovich A, Goffin V, Pisera D. Prolactin receptor antagonism in mouse anterior pituitary: effects on cell turnover and prolactin receptor expression. Am J Physiol Endocrinol Metab. 2012;302:E356–E364. doi: 10.1152/ajpendo.00333.2011. [DOI] [PubMed] [Google Scholar]
- Ferraris J, Zarate S, Jaita G, Boutillon F, Bernadet M, Auffret J, Seilicovich A, Binart N, Goffin V, Pisera D. Prolactin induces apoptosis of lactotropes in female rodents. PLoS ONE. 2014;9:e97383. doi: 10.1371/journal.pone.0097383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friedman E, Adams HF, Hoog A, Gejman PV, Carson E, Larsson C, De Marco L, Werner S, Fahlbusch R, Nordenskjöld M. Normal structural dopamine type 2 receptor gene in prolactin-secreting and other pituitary tumors. Journal of Clinical Endocrinology and Metabolism. 1994;78:568–574. doi: 10.1210/jcem.78.3.7907340. [DOI] [PubMed] [Google Scholar]
- Fujiwara K, Ikeda H, Yoshimoto T. Immunohistochemical demonstration of TGF-beta-receptor type II in human pituitary adenomas. Acta Histochem. 1995;97:445–454. doi: 10.1016/S0065-1281(11)80071-1. [DOI] [PubMed] [Google Scholar]
- Fusco A, Gunz G, Jaquet P, Dufour H, Germanetti AL, Culler MD, Barlier A, Saveanu A. Somatostatinergic ligands in dopamine-sensitive and -resistant prolactinomas. European Journal of Endocrinology. 2008;158:595–603. doi: 10.1530/EJE-07-0806. [DOI] [PubMed] [Google Scholar]
- Galvin-Burgess KE, Travis ED, Pierson KE, Vivian JL. TGF-beta-superfamily signaling regulates embryonic stem cell heterogeneity: self-renewal as a dynamic and regulated equilibrium. Stem Cells. 2013;31:48–58. doi: 10.1002/stem.1252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garside SA, Henkin J, Morris KD, Norvell SM, Thomas FH, Fraser HM. A thrombospondin-mimetic peptide, ABT-898, suppresses angiogenesis and promotes follicular atresia in pre- and early-antral follicles in vivo. Endocrinology. 2010;151:5905–5915. doi: 10.1210/en.2010-0283. [DOI] [PubMed] [Google Scholar]
- Gillam MP, Molitch ME. Prolactinoma. In: Melmed Shlomo., editor. The Pituitary. 3. Elsevier Inc; 2015. [Google Scholar]
- Gillam MP, Molitch ME, Lombardi G, Colao A. Advances in the treatment of prolactinomas. Endocrine reviews. 2006;27:485–534. doi: 10.1210/er.2005-9998. [DOI] [PubMed] [Google Scholar]
- Gordon MS, Mendelson D, Carr R, Knight RA, Humerickhouse RA, Iannone M, Stopeck AT. A phase 1 trial of 2 dose schedules of ABT-510, an antiangiogenic, thrombospondin-1-mimetic peptide, in patients with advanced cancer. Cancer. 2008;113:3420–3429. doi: 10.1002/cncr.23953. [DOI] [PubMed] [Google Scholar]
- Gurlek A, Karavitaki N, Ansorge O, Wass JA. What are the markers of aggressiveness in prolactinomas? Changes in cell biology, extracellular matrix components, angiogenesis and genetics. European Journal of Endocrinology. 2007;156:143–153. doi: 10.1530/eje.1.02339. [DOI] [PubMed] [Google Scholar]
- Halper J, Parnell PG, Carter BJ, Ren P, Scheithauer BW. Presence of growth factors in human pituitary. Laboratory Investigation. 1992;66:639–645. [PubMed] [Google Scholar]
- Han J, Alvarez-Breckenridge CA, Wang QE, Yu J. TGF-beta signaling and its targeting for glioma treatment. Am J Cancer Res. 2015;5:945–955. [PMC free article] [PubMed] [Google Scholar]
- Haviv F, Bradley MF, Kalvin DM, Schneider AJ, Davidson DJ, Majest SM, McKay LM, Haskell CJ, Bell RL, et al. Thrombospondin-1 mimetic peptide inhibitors of angiogenesis and tumor growth: design, synthesis, and optimization of pharmacokinetics and biological activities. J Med Chem. 2005;48:2838–2846. doi: 10.1021/jm0401560. [DOI] [PubMed] [Google Scholar]
- Heaney AP, Fernando M, Melmed S. Functional role of estrogen in pituitary tumor pathogenesis. Journal of Clinical Investigation. 2002;109:277–283. doi: 10.1172/JCI14264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heaney AP, Horwitz GA, Wang Z, Singson R, Melmed S. Early involvement of estrogen-induced pituitary tumor transforming gene and fibroblast growth factor expression in prolactinoma pathogenesis. Nature Medicine. 1999;5:1317–1321. doi: 10.1038/15275. [DOI] [PubMed] [Google Scholar]
- Heldin CH, Landstrom M, Moustakas A. Mechanism of TGF-beta signaling to growth arrest, apoptosis, and epithelial-mesenchymal transition. Curr Opin Cell Biol. 2009;21:166–176. doi: 10.1016/j.ceb.2009.01.021. [DOI] [PubMed] [Google Scholar]
- Hentges S, Sarkar DK. Transforming growth factor-beta regulation of estradiol-induced prolactinomas. Frontiers in Neuroendocrinology. 2001;22:340–363. doi: 10.1006/frne.2001.0220. [DOI] [PubMed] [Google Scholar]
- Itoh F, Watabe T, Miyazono K. Roles of TGF-beta family signals in the fate determination of pluripotent stem cells. Semin Cell Dev Biol. 2014;32:98–106. doi: 10.1016/j.semcdb.2014.05.017. [DOI] [PubMed] [Google Scholar]
- Jaita G, Zarate S, Ferraris J, Gottardo MF, Eijo G, Magri ML, Pisera D, Seilicovich A. Estradiol Upregulates c-FLIPlong Expression in Anterior Pituitary Cells. Horm Metab Res. 2015 doi: 10.1055/s-0035-1565068. [DOI] [PubMed] [Google Scholar]
- Jaquet P, Ouafik L, Saveanu A, Gunz G, Fina F, Dufour H, Culler MD, Moreau JP, Enjalbert A. Quantitative and functional expression of somatostatin receptor subtypes in human prolactinomas. J Clin Endocrinol Metab. 1999;84:3268–3276. doi: 10.1210/jcem.84.9.5962. [DOI] [PubMed] [Google Scholar]
- Jiang M, Mou CZ, Han T, Wang M, Yang W. Thrombospondin-1 and transforming growth factor-beta1 levels in prolactinoma and their clinical significance. J Int Med Res. 2012;40:1284–1294. doi: 10.1177/147323001204000407. [DOI] [PubMed] [Google Scholar]
- Jin L, Qian X, Kulig E, Sanno N, Scheithauer BW, Kovacs K, Young WF, Jr, Lloyd RV. Transforming growth factor-beta, transforming growth factor-beta receptor II, and p27Kip1 expression in nontumorous and neoplastic human pituitaries. Am J Pathol. 1997;151:509–519. [PMC free article] [PubMed] [Google Scholar]
- Kelly MA, Rubinstein M, Asa SL, Zhang G, Saez C, Bunzow JR, Allen RG, Hnasko R, Ben-Jonathan N, Grandy DK, et al. Pituitary lactotroph hyperplasia and chronic hyperprolactinemia in dopamine D2 receptor-deficient mice. Neuron. 1997;19:103–113. doi: 10.1016/s0896-6273(00)80351-7. [DOI] [PubMed] [Google Scholar]
- Kovacs K, Horvath E, Vidal S. Classification of pituitary adenomas. J Neurooncol. 2001;54:121–127. doi: 10.1023/a:1012945129981. [DOI] [PubMed] [Google Scholar]
- Kulkarni AB, Huh CG, Becker D, Geiser A, Lyght M, Flanders KC, Roberts AB, Sporn MB, Ward JM, Karlsson S. Transforming growth factor beta 1 null mutation in mice causes excessive inflammatory response and early death. Proceedings of the National Academy of Science. 1993;90:770–774. doi: 10.1073/pnas.90.2.770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lawler J. Thrombospondin-1 as an endogenous inhibitor of angiogenesis and tumor growth. J Cell Mol Med. 2002;6:1–12. doi: 10.1111/j.1582-4934.2002.tb00307.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lawler PR, Lawler J. Molecular basis for the regulation of angiogenesis by thrombospondin-1 and -2. Cold Spring Harb Perspect Med. 2012;2:a006627. doi: 10.1101/cshperspect.a006627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li Z, Liu Q, Li C, Zong X, Bai J, Wu Y, Lan X, Yu G, Zhang Y. The role of TGF-beta/Smad signaling in dopamine agonist-resistant prolactinomas. Mol Cell Endocrinol. 2015;402:64–71. doi: 10.1016/j.mce.2014.12.024. [DOI] [PubMed] [Google Scholar]
- Liu JK, Patel J, Eloy JA. The role of temozolomide in the treatment of aggressive pituitary tumors. J Clin Neurosci. 2015;22:923–929. doi: 10.1016/j.jocn.2014.12.007. [DOI] [PubMed] [Google Scholar]
- Markovic SN, Suman VJ, Rao RA, Ingle JN, Kaur JS, Erickson LA, Pitot HC, Croghan GA, McWilliams RR, Merchan J, et al. A phase II study of ABT-510 (thrombospondin-1 analog) for the treatment of metastatic melanoma. Am J Clin Oncol. 2007;30:303–309. doi: 10.1097/01.coc.0000256104.80089.35. [DOI] [PubMed] [Google Scholar]
- McCormack AI, Wass JA, Grossman AB. Aggressive pituitary tumours: the role of temozolomide and the assessment of MGMT status. Eur J Clin Invest. 2011;41:1133–1148. doi: 10.1111/j.1365-2362.2011.02520.x. [DOI] [PubMed] [Google Scholar]
- Melmed S. Pathogenesis of pituitary tumors. Nat Rev Endocrinol. 2011;7:257–266. doi: 10.1038/nrendo.2011.40. [DOI] [PubMed] [Google Scholar]
- Melmed S. Pituitary tumors. Endocrinol Metab Clin North Am. 2015;44:1–9. doi: 10.1016/j.ecl.2014.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Molitch ME. Dopamine resistance of prolactinomas. Pituitary. 2003;6:19–27. doi: 10.1023/a:1026225625897. [DOI] [PubMed] [Google Scholar]
- Molitch ME. Management of medically refractory prolactinoma. J Neurooncol. 2014;117:421–428. doi: 10.1007/s11060-013-1270-8. [DOI] [PubMed] [Google Scholar]
- Moustakas A, Heldin CH. Non-Smad TGF-beta signals. J Cell Sci. 2005;118:3573–3584. doi: 10.1242/jcs.02554. [DOI] [PubMed] [Google Scholar]
- Mueller SG, Kudlow JE. Transforming growth factor-beta (TGF beta) inhibits TGF alpha expression in bovine anterior pituitary-derived cells. Molecular Endocrinology. 1991;5:1439–1446. doi: 10.1210/mend-5-10-1439. [DOI] [PubMed] [Google Scholar]
- Nabors LB, Fiveash JB, Markert JM, Kekan MS, Gillespie GY, Huang Z, Johnson MJ, Meleth S, Kuo H, Gladson CL, et al. A phase 1 trial of ABT-510 concurrent with standard chemoradiation for patients with newly diagnosed glioblastoma. Arch Neurol. 2010;67:313–319. doi: 10.1001/archneurol.2010.16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nishioka H, Haraoka J, Akada K. Growth potential of prolactinomas in men: is it really different from women? Surg Neurol. 2003;59:386–390. doi: 10.1016/s0090-3019(03)00012-0. [DOI] [PubMed] [Google Scholar]
- Paez-Pereda M, Kuchenbauer F, Arzt E, Stalla GK. Regulation of pituitary hormones and cell proliferation by components of the extracellular matrix. Braz J Med Biol Res. 2005;38:1487–1494. doi: 10.1590/s0100-879x2005001000005. [DOI] [PubMed] [Google Scholar]
- Passos VQ, Fortes MA, Giannella-Neto D, Bronstein MD. Genes differentially expressed in prolactinomas responsive and resistant to dopamine agonists. Neuroendocrinology. 2009;89:163–170. doi: 10.1159/000156116. [DOI] [PubMed] [Google Scholar]
- Pastorcic M, De A, Boyadjieva N, Vale W, Sarkar DK. Reduction in the expression and action of transforming growth factor-b1 on lactotropes during estrogen-induced tumorigenesis. Cancer Research. 1995;55:4892–4898. [PubMed] [Google Scholar]
- Pellegrini I, Rasolonjanahary R, Gunz G, Bertrand P, Delivet S, Jedynak CP, Kordon C, Peillon F, Jaquet P, Enjalbert A. Resistance to bromocriptine in prolactinomas. Journal of Clinical Endocrinology and Metabolism. 1989;69:500–509. doi: 10.1210/jcem-69-3-500. [DOI] [PubMed] [Google Scholar]
- Pisera D, Candolfi M, Navarra S, Ferraris J, Zaldivar V, Jaita G, Castro MG, Seilicovich A. Estrogens sensitize anterior pituitary gland to apoptosis. Am J Physiol Endocrinol Metab. 2004;287:E767–E771. doi: 10.1152/ajpendo.00052.2004. [DOI] [PubMed] [Google Scholar]
- Pohlers D, Brenmoehl J, Loffler I, Muller CK, Leipner C, Schultze-Mosgau S, Stallmach A, Kinne RW, Wolf G. TGF-beta and fibrosis in different organs - molecular pathway imprints. Biochim Biophys Acta. 2009;1792:746–756. doi: 10.1016/j.bbadis.2009.06.004. [DOI] [PubMed] [Google Scholar]
- Primeau V, Raftopoulos C, Maiter D. Outcomes of transsphenoidal surgery in prolactinomas: improvement of hormonal control in dopamine agonist-resistant patients. European Journal of Endocrinology. 2012;166:779–786. doi: 10.1530/EJE-11-1000. [DOI] [PubMed] [Google Scholar]
- Recouvreux MV, Camilletti MA, Rifkin DB, Becu-Villalobos D, Diaz-Torga G. Thrombospondin-1 (TSP-1) Analogs ABT-510 and ABT-898 Inhibit Prolactinoma Growth and Recover Active Pituitary Transforming Growth Factor-beta1 (TGF-beta1) Endocrinology. 2012;153:3861–3871. doi: 10.1210/en.2012-1007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Recouvreux MV, Guida MC, Rifkin DB, Becu-Villalobos D, Diaz-Torga G. Active and Total Transforming Growth Factor-{beta}1 Are Differentially Regulated by Dopamine and Estradiol in the Pituitary. Endocrinology. 2011;152:2722–2730. doi: 10.1210/en.2010-1464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Recouvreux MV, Lapyckyj L, Camilletti MA, Guida MC, Ornstein A, Rifkin DB, Becu-Villalobos D, Diaz-Torga G. Sex differences in the pituitary transforming growth factor-beta1 system: studies in a model of resistant prolactinomas. Endocrinology. 2013;154:4192–4205. doi: 10.1210/en.2013-1433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rifkin DB. Latent transforming growth factor-beta (TGF-beta) binding proteins: orchestrators of TGF-beta availability. Journal of Biological Chemistry. 2005;280:7409–7412. doi: 10.1074/jbc.R400029200. [DOI] [PubMed] [Google Scholar]
- Saiardi A, Bozzi Y, Baik J-H, Borrelli E. Antiproliferative role of dopamine: loss of D2 receptors causes hormonal dysfunction and pituitary hyperplasia. Neuron. 1997;19:115–126. doi: 10.1016/s0896-6273(00)80352-9. [DOI] [PubMed] [Google Scholar]
- Sarkar AJ, Chaturvedi K, Chen CP, Sarkar DK. Changes in thrombospondin-1 levels in the endothelial cells of the anterior pituitary during estrogen-induced prolactin-secreting pituitary tumors. Journal of Endocrinology. 2007;192:395–403. doi: 10.1677/joe.1.06925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sarkar DK, Chaturvedi K, Oomizu S, Boyadjieva NI, Chen CP. Dopamine, dopamine D2 receptor short isoform, transforming growth factor (TGF)-beta1, and TGF-beta type II receptor interact to inhibit the growth of pituitary lactotropes. Endocrinology. 2005;146:4179–4188. doi: 10.1210/en.2005-0430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sarkar DK, Kim KK, Minami S. Transforming growth factor-beta1 messenger RNA and protein expression in the pituitary gland: its action on prolactin secretion and lactotropic growth. Molecular Endocrinology. 1992;6:1825–1833. doi: 10.1210/mend.6.11.1480172. [DOI] [PubMed] [Google Scholar]
- Schuff KG, Hentges ST, Kelly MA, Binart N, Kelly PA, Iuvone PM, Asa SL, Low MJ. Lack of prolactin receptor signaling in mice results in lactotroph proliferation and prolactinomas by dopamine-dependent and -independent mechanisms. Journal of Clinical Investigation. 2002;110:973–981. doi: 10.1172/JCI15912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schultz-Cherry S, Ribeiro S, Gentry L, Murphy-Ullrich JE. Thrombospondin binds and activates the small and large forms of latent transforming growth factor-beta in a chemically defined system. Journal of Biological Chemistry. 1994;269:26775–26782. [PubMed] [Google Scholar]
- Shi Y, Massague J. Mechanisms of TGF-beta signaling from cell membrane to the nucleus. Cell. 2003;113:685–700. doi: 10.1016/s0092-8674(03)00432-x. [DOI] [PubMed] [Google Scholar]
- Shida N, Ikeda H, Yoshimoto T, Oshima M, Taketo MM, Miyoshi I. Estrogen-induced tumorigenesis in the pituitary gland of TGF-beta(+/-) knockout mice. Biochim Biophys Acta. 1998;1407:79–83. doi: 10.1016/s0925-4439(98)00024-6. [DOI] [PubMed] [Google Scholar]
- Shimazu S, Shimatsu A, Yamada S, Inoshita N, Nagamura Y, Usui T, Tsukada T. Resistance to dopamine agonists in prolactinoma is correlated with reduction of dopamine D2 receptor long isoform mRNA levels. European Journal of Endocrinology. 2012;166:383–390. doi: 10.1530/EJE-11-0656. [DOI] [PubMed] [Google Scholar]
- Smith TR, Hulou MM, Huang KT, Gokoglu A, Cote DJ, Woodmansee WW, Laws ER., Jr Current indications for the surgical treatment of prolactinomas. J Clin Neurosci. 2015 doi: 10.1016/j.jocn.2015.06.001. [DOI] [PubMed] [Google Scholar]
- Syro LV, Rotondo F, Ramirez A, Di Ieva A, Sav MA, Restrepo LM, Serna CA, Kovacs K. Progress in the Diagnosis and Classification of Pituitary Adenomas. Front Endocrinol(Lausanne) 2015;6:97. doi: 10.3389/fendo.2015.00097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vasilev V, Daly AF, Vroonen L, Zacharieva S, Beckers A. Resistant prolactinomas. J Endocrinol Invest. 2011;34:312–316. doi: 10.1007/BF03347092. [DOI] [PubMed] [Google Scholar]
- Volker W, Gehring WG, Berning R, Schmidt RC, Schneider J, von zur MA. Impaired pituitary response to bromocriptine suppression: reversal after bromocriptine plus tamoxifen. Acta Endocrinol(Copenh) 1982;101:491–500. doi: 10.1530/acta.0.1010491. [DOI] [PubMed] [Google Scholar]
- Vroonen L, Jaffrain-Rea ML, Petrossians P, Tamagno G, Chanson P, Vilar L, Borson-Chazot F, Naves LA, Brue T, Gatta B, et al. Prolactinomas resistant to standard doses of cabergoline: a multicenter study of 92 patients. European Journal of Endocrinology. 2012;167:651–662. doi: 10.1530/EJE-12-0236. [DOI] [PubMed] [Google Scholar]
- Whitelaw BC, Dworakowska D, Thomas NW, Barazi S, Riordan-Eva P, King AP, Hampton T, Landau DB, Lipscomb D, Buchanan CR, et al. Temozolomide in the management of dopamine agonist-resistant prolactinomas. Clinical Endocrinology (Oxford) 2012;76:877–886. doi: 10.1111/j.1365-2265.2012.04373.x. [DOI] [PubMed] [Google Scholar]
- Wong A, Eloy JA, Couldwell WT, Liu JK. Update on prolactinomas. Part 1: Clinical manifestations and diagnostic challenges. J Clin Neurosci. 2015a doi: 10.1016/j.jocn.2015.03.058. [DOI] [PubMed] [Google Scholar]
- Wong A, Eloy JA, Couldwell WT, Liu JK. Update on prolactinomas. Part 2: Treatment and management strategies. J Clin Neurosci. 2015b doi: 10.1016/j.jocn.2015.03.059. [DOI] [PubMed] [Google Scholar]
- Yang Q, Tian Y, Liu S, Zeine R, Chlenski A, Salwen HR, Henkin J, Cohn SL. Thrombospondin-1 peptide ABT-510 combined with valproic acid is an effective antiangiogenesis strategy in neuroblastoma. Cancer Res. 2007;67:1716–1724. doi: 10.1158/0008-5472.CAN-06-2595. [DOI] [PubMed] [Google Scholar]
- Ying SY, Becker A, Baird A, Ling N, Ueno N, Esch F, Guillemin R. Type beta transforming growth factor (TGF-beta) is a potent stimulator of the basal secretion of follicle stimulating hormone (FSH) in a pituitary monolayer system. Biochemical and Biophysical Research Communications. 1986;135:950–956. doi: 10.1016/0006-291x(86)91020-x. [DOI] [PubMed] [Google Scholar]
- Yoshinaga K, Obata H, Jurukovski V, Mazzieri R, Chen Y, Zilberberg L, Huso D, Melamed J, Prijatelj P, Todorovic V, et al. Perturbation of transforming growth factor (TGF)-beta1 association with latent TGF-beta binding protein yields inflammation and tumors. Proceedings of the National Academy of Science. 2008;105:18758–18763. doi: 10.1073/pnas.0805411105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zaldivar V, Magri ML, Zarate S, Jaita G, Eijo G, Radl D, Ferraris J, Pisera D, Seilicovich A. Estradiol increases the Bax/Bcl-2 ratio and induces apoptosis in the anterior pituitary gland. Neuroendocrinology. 2009;90:292–300. doi: 10.1159/000235618. [DOI] [PubMed] [Google Scholar]
- Zhenye L, Chuzhong L, Youtu W, Xiaolei L, Lei C, Lichuan H, Hongyun W, Yonggang W, Fei W, Yazhuo Z. The expression of TGF-beta1, Smad3, phospho-Smad3 and Smad7 is correlated with the development and invasion of nonfunctioning pituitary adenomas. J Transl Med. 2014;12:71. doi: 10.1186/1479-5876-12-71. [DOI] [PMC free article] [PubMed] [Google Scholar]