

Abstract
Increased mortality in antiretroviral (ARV)-treated, HIV-infected individuals has been attributed to persistent immune dysfunction, in part due to abnormalities at the gastrointestinal barrier. In particular, the poor reconstitution of gastrointestinal TH17 cells correlates with residual translocation of dysbiotic, immunostimulatory microflora across a compromised intestinal epithelial barrier. We have previously demonstrated that oral probiotics promote increased intestinal CD4+ T-cell reconstitution during ARV treatment in a non-human primate model of HIV infection; however, essential mucosal T-cell subsets, such as TH17 cells, had limited recovery. Here, we sought to promote TH17 cell recovery by administering IL-21 to a limited number of ARV-treated, probiotic-supplemented, SIV-infected pigtailed macaques. We demonstrate that probiotic and IL-21 supplementation of ARVs is associated with enhanced polyfunctional TH17 expansion and reduced markers of microbial translocation and dysbiosis as compared to infected controls receiving ARVs alone. Importantly, treatment resulted in fewer morbidities compared to controls, and was independent of increased immune activation or loss of viral suppression. We propose that combining ARVs with therapeutics aimed at restoring intestinal stasis may significantly improve disease prognosis of ARV-treated, HIV-infected, individuals.
Introduction
Progressive HIV infection is characterized by persistent innate and adaptive immune activation that is predictive of disease progression and that is not resolved with effective ARV therapy1, 2. Although mediators of immune activation are multi-faceted, it is evident that even with effective viral suppression, persistent immune dysfunction at the intestinal epithelial barrier permits the translocation of immunostimulatory luminal contents into intestinal tissues and systemic circulation3. In non-human primate (NHP) models of HIV infection, microbial translocation distinguishes progressive from non-progressive models of SIV infection, with progressive infection models such as rhesus (Macaca mulatta) and pigtailed (Macaca nemistrina) macaques displaying evidence of microbial translocation and immune activation in stark contrast to non-progressive hosts such as sooty mangabeys (Cercocebus atys)4–6. Importantly, although both progressive and non-progressive infected hosts exhibit some loss of intestinal CD4+ T-cells, progressive models exhibit a preferential loss of TH17 cells, implicating TH17 cell loss as a primary contributor to microbial translocation and disease progression6–8.
TH17 cells are a subset of differentiated, IL-17-expressing memory CD4+ T-cells important for control of the microbiota at mucosal barrier sites9. TH17 cells develop from naïve CD4+ T-cells in response to TGF-β and pro-inflammatory cytokine signaling induced by microbial antigens10–12. Although TH17 differentiation can be calibrated by the microbiome13, 14, the effects of TH17 cells on gastrointestinal immunity extend beyond interplay with commensal species. Thus, while commensal species are sufficient for the generation of TH17 cells13, 14, the ability of TH17 cells to induce production of antimicrobial peptides and chemotactic cytokines from the intestinal epithelium contributes to resistance against colonization with Salmonella typhimurium, Streptococcus pneumoniae, and Candida albicans8, 9. Implicit in this counter-regulation, microbial dysbiosis – an imbalance in the relative frequencies of commensal and pathogenic species – is associated with a dysregulation of TH17 frequencies and function15. This, in turn, is implicated in inflammation in a multitude of chronic autoimmune and infectious diseases. In HIV-infected individuals and SIV-infected macaques, microbial dysbiosis has been described as a contributor to TH17 loss and microbial translocation, with some dysbiotic species shown to be enriched in both treated and untreated subjects, and to be associated with markers of disease progression16, 17. Given the contributions of dysbiotic and translocating bacterial species to disease progression in treated, HIV-infected individuals, we previously considered that augmenting ARV-therapy in chronically SIV-infected macaques with probiotics might promote a restoration of the commensal microbiota, initiating a sequence of immune restorative processes. Although probiotic supplementation was associated with intestinal CD4+ T-cell restoration and enhanced TH17 functionality, we did not observe a significant improvement in the frequency of TH17 cells nor was there any discernible impact on the relative frequencies of fecal microbial communities18.
Subsequent to priming by APCs, TH17 development is augmented by autocrine production of the common γ-chain cytokine, IL-2119, 20. Though not essential for the differentiation of TH17 cells14, 21, blockade of IL-21 during TH17 development limits the expansion and functional capacity of TH17 cells19, 20. In progressive SIV infection, a paucity of IL-21-producing CD4+ T-cells correlates with TH17 loss22. Moreover, while IL-21 treatment during acute SIV infection has been shown to delay TH17 loss independent of ARV therapy, cessation of IL-21 administration results in a rapid decline in TH17 frequencies23.
Herein, we considered that augmenting ARV therapy with combination probiotic and IL-21 therapy might enhance the recovery and maintenance of TH17 cells in chronic SIV infection. As described, we observed that this regimen is associated with increased frequencies of intestinal polyfunctional TH17 cells as compared to ARV-only controls, and that supplementation is further associated with improvements in microbial translocation, microbial dysbiosis and clinical status. Importantly, as our therapy did not promote immune activation or negate viral suppression, we believe that our findings suggest that probiotic and IL-21 supplementation of ARVs may translate into a safe and effective therapeutic for the reduction of co-morbidities suffered by ARV-treated, HIV-infected individuals.
Results
Experimental Design
We previously demonstrated that treatment of SIV-infected, virus-suppressed pigtailed macaques (PTM) with probiotics improved gastrointestinal integrity and immunity; however, probiotic supplementation alone did not promote a restoration of TH17 cells nor was there evidence for a shift towards a significantly less inflammatory microbiome18. Here, we sought to determine if treatment with IL-2119, 20 enhances probiotic treatment in SIV-infected, ARV- and probiotic-treated PTMs by promoting TH17 recovery and reducing microbial translocation and dysbiosis. As depicted in Fig. 1, we infected 11 PTM with SIVmac239 and, at day 98 post-infection (p.i.), all animals began daily treatment with the reverse transcriptase inhibitors, PMPA and FTC, and the integrase inhibitor, L’812. Of these animals, 6 were simultaneously treated with the daily probiotic, VLS#3® (consisting of 1011 live Bifidobacterium, Lactobacillus and Streptococcus species) and with early and late administration courses of IL-21 to assess refractoriness to continued IL-21 therapy. We routinely sampled the blood and stool of all animals, sampled the pulmonary airways through bronchoalveolar lavage (BAL) and obtained jejunal (Jej), lymph node (LN), and rectal (RB) biopsies.
Figure 1. Experimental Design.
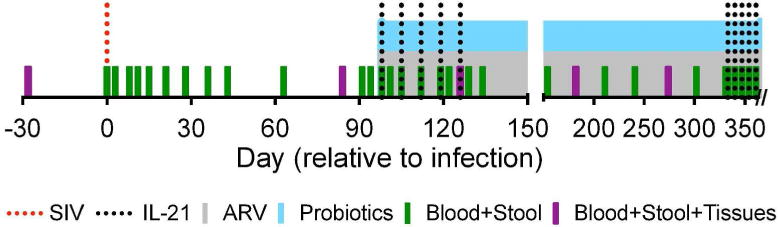
Eleven PTM were infected with SIVmac239 at day 0 (dotted red line) and daily ARV therapy (grey shaded region) was initiated at day 98 p.i. Of these animals, 6 PTM were additionally treated with daily probiotic therapy (blue shaded region) as well as two rounds of IL-21 therapy, administered once weekly for five weeks (dotted black lines). One ARV-only PTM was euthanized at day 126 p.i. per protocol specified clinical endpoint. Blood, stool and tissues (BAL, Jej, LN, and RB) were collected at pre-determined intervals, as indicated (green and purple bars).
IL-21 and Probiotic Supplementation Does Not Alter Viral Suppression in ARV-Treated Macaques
IL-21 is a pleiotropic cytokine known to promote the development of TH17 cells from naïve CD4+ T-cell precursors19, 20. Since mature CD4+ T-cells are the preferential targets for HIV and SIV infection, we assessed the impact of the IL-21 and probiotic regimen on viral suppression. With the initiation of therapy, all animals exhibited a rapid decline in plasma viral RNA (vRNA) with all but one probiotic, IL-21-supplemented animal (PTA2P031) achieving suppression to below 100 copies/mL within 30 days of treatment initiation (Fig. 2A). In each group, all but one animal exhibited at least one instance of breakthrough viremia; however, measurable virus in these animals did not exceed 100 copies/mL. Importantly, breakthrough viremia in supplemented animals was not associated with increased activation of systemic CD4+ or CD8+ memory T-cells (TM), as assessed by HLA-DR and Ki67 expression, or with increases in plasma soluble CD14 (sCD14) or LPS-binding protein (LBP) (Figure S1 and data not shown).
Figure 2. IL-21 and Probiotic Therapy does not Alter Viral Load or Cellular Distribution in ARV-Treated PTMs.

a. Longitudinal, log copies/mL of plasma vRNA from animals treated with ARVs only (open squares) or with ARVs, probiotic VSL#3, and IL-21 (closed circles). Grey shading indicates treatment initiation and duration. Dotted lines denote IL-21 administration timepoints. Symbols and shading are consistent throughout the manuscript unless otherwise noted. b. Copies of vDNA per 100 colonic CD4+ TM at the time of necropsy. Triangles indicate samples normalized to the limit of detection
IL-21 has been shown to promote the expansion of antiviral CD8+ T-cells23–25; however, we considered that CD4+ T-cell reconstitution might counteract ARV− and CD8+ T cell-mediated antiviral effects by expanding the viral reservoir. Indeed, the addition of IL-21 to probiotic and ARV therapy was associated with enhanced intestinal CD4+ T-cell recovery as compared ARV-only controls (Figure S2), similar to what we previously reported using probiotic-supplementation alone18. To quantify the viral reservoir within intestinal tissues – which exhibit the greatest loss of CD4+ T-cells after infection – we sorted and measured cell-associated viral DNA (vDNA) within colonic, CD4+ memory T-cells (TM; CD95+) at necropsy. Despite a trend for higher cell-associated vDNA in IL-21- and probiotic-supplemented animals, no significant differences were noted in residual vDNA between ARV-only (0.0188 ±0.00566 copies/100 cells; mean±SEM) and IL-21- and probiotic-supplemented (0.0364 ±0.0219) animals (Fig. 2B). Thus, probiotic- and IL-21-supplementation does not appear to dramatically alter virus production or infectivity within intestinal tissues in SIV-infected, ARV-treated PTMs.
Improved Clinical Status in ARV-Treated Macaques Supplemented with Il-21 and Probiotics
Effective ARV-therapy significantly improves disease progression and clinical prognosis in chronically HIV-infected individuals; however, prolonged therapy remains fraught with non-AIDS related co-morbidities1, 2, especially in situations in which dysbiosis is likely to be present26. To determine if probiotic and IL-21 supplementation might improve the prognosis of our animals, we compared the incidence of clinical complications between our experimental groups, as diagnosed by positive bacterial culture or altered blood chemistries (Fig. 3). Interestingly, although no overt complications were noted in the probiotic and IL-21 supplemented group, opportunistic infection (respiratory Enterobacter cloacae) and renal injury (azotemia as determined by elevated plasma levels of urea nitrogen or creatinine) were evident among 3 of the 5 ARV-only animals27, 28. While low platelet counts have been shown to be associated with disease progression in SIV-infected PTM29, we found similar counts between experimental groups throughout the study period (data not shown). Of animals that exhibited complications, all displayed subsequent instances of severe diarrhea with one animal requiring unplanned euthanasia, per protocol-specified humane clinical endpoints. To quantitatively assess disease progression, we compared the time to the first diagnosed clinical complication in ARV-only (median day 128 p.i.) and probiotic, IL-21-supplemented (no complication through study termination) animals and determined that supplementation was associated with a significant difference in the time to first complication (p=0.0316, Mantel-Cox). These results indicate that probiotic and IL-21 supplementation of ARV-therapy decreased the incidence of non-AIDS morbidities.
Figure 3. IL-21 and Probiotic Therapy Improves Clinical Status in ARV-Treated PTMs.
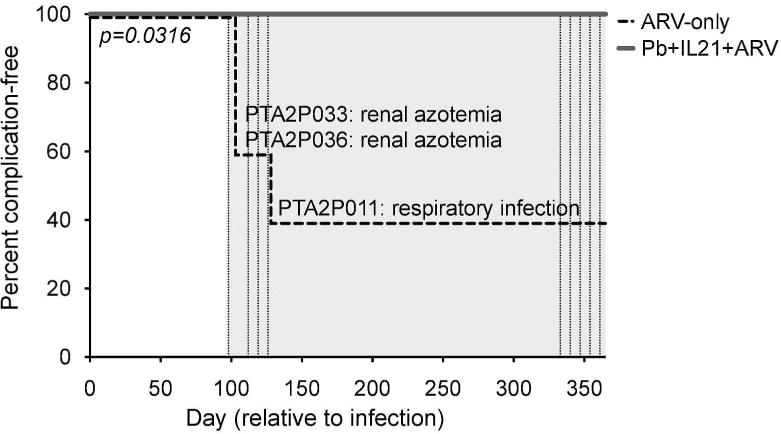
Percent of animals remaining free from clinical complications over time. Initial complications per animal are noted alongside changes in the incidence-free curve, with ARV-only animals represented by the dashed line and supplemented animals by the solid line. Significance in the time to the first reported clinical complication between groups was assessed by the Mantel-Cox Log-Rank test.
IL-21 and Probiotic Supplementation Promote Polyfunctional TH17 Recovery In ARV-Treated Macaques
In ARV-treated individuals and macaques, persistent microbial translocation and dysbiosis are associated with continued immune activation, thereby possibly exacerbating lentiviral-mediated TH17 losses16, 17. As our previous findings indicated that probiotic supplementation of ARV-therapy alone was unable to promote significant restoration of TH17 cells, we considered that additional supplementation with IL-21 might promote sustained restoration of these cells. We assessed the frequency of IL-17 expressing memory CD4+ T-cells (TH17) longitudinally in the PBMCs, BAL, Jej, LN, and RB. Among BAL, LN and PBMC CD4+ T-cells, probiotic and IL-21 supplemented animals exhibited no significant differences in TH17 frequencies as compared to ARV-only controls or as compared to the pre-treatment timepoint of day 84 p.i. (data not shown). Indeed, preferential loss of TH17 within chronically HIV/SIV-infected individuals is not observed within these specific anatomic sites7, 30. However, modest to significant improvements in the frequencies of intestinal TH17 cells were observed in supplemented animals as compared to the pre-treatment baseline timepoint (Fig. 4A–B and S3A). Coupled with increased intestinal CD4+ T-cell frequencies (Fig. S2), the improved relative frequencies of intestinal TH17 cells led to significantly improved absolute frequencies of intestinal TH17 cells (% IL-17-expressing CD4+TM of CD3+ T-cells). We observed increases in the absolute frequency of jejunal TH17 cells from day 84 p.i. to days 274 and 365 p.i. (p=0.016, 0.031) and in rectal TH17 cells from day 84 p.i. to days 126, 274, and 365 p.i (p=0.031, 0.031, 0.016; Fig. 4A–B) within the probiotic and IL-21 supplemented animals. No significant differences were evident in ARV-only controls (Table S1). Furthermore, at day 182 p.i. (98 days post-intervention), ARV-only controls exhibited a significantly lower absolute frequency of jejunal TH17 cells as compared to probiotic and IL-21 supplemented animals (p=0.0286).
Figure 4. IL-21 and Probiotic Therapy Promotes the Absolute Expansion of Polyfunctional TH17 Cells in ARV-Treated PTMs.
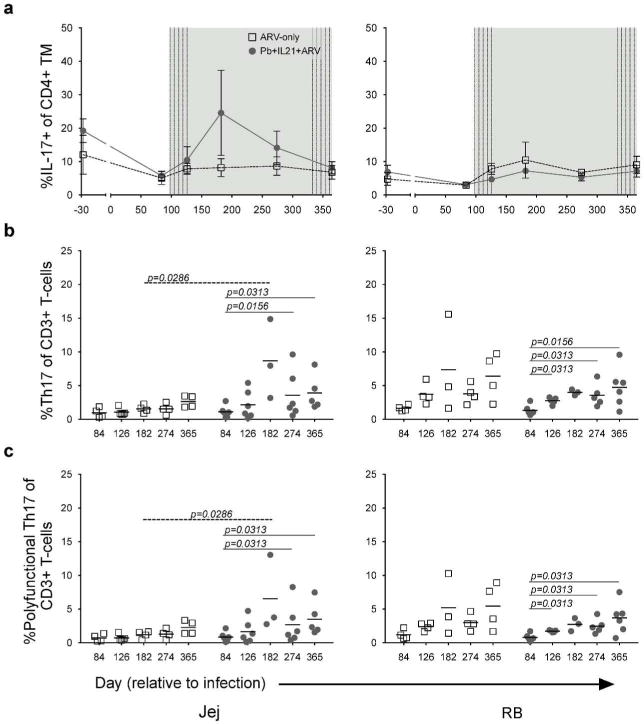
a. Longitudinal mean (±SEM) percent IL-17+ of CD4+ TM (TH17) from the Jej (left) and RB (right). b. Absolute percent TH17 (i.e., the percent of IL-17+ CD4+ TM found within CD3+ T cells) from the Jej and RB at the indicated days p.i. c. Absolute percent polyfunctional TH17 from the Jej and RB at the indicated days p.i. Line through samples represents mean; lines above samples span timepoints of significance. Significance within groups was assessed by Wilcoxon Signed-Rank test. Significance across groups was assessed by Mann-Whitney U test.
Superior control of disease progression in HIV-infected individuals has been attributed to improved functionality (multiple cytokine expression, or polyfunctionality) among T-cells31–33. As such, we further investigated the frequency of intestinal, polyfunctional TH17 cells – as assessed here by the additional expression of IL-2, IL-22, or TNFα – in our animals. Although no significant differences were observed between groups in the expression of individual cytokines (data not shown), significant differences in the relative frequency of polyfunctional jejunal TH17 cells were noted exclusively among probiotic and IL-21 supplemented animals at necropsy as compared to days 84 and 274 p.i. (p=0.031, 0.031; Fig. S3B, Table S1). As compared to ARV-only controls, the relative frequencies of polyfunctional TH17 cells were lower in supplemented animals at day 274 p.i. within the Jej (p=0.033) and RB (p=0.032) but comparable at all other timepoints. However, an examination of absolute intestinal polyfunctional TH17 frequencies revealed significant increases specific to probiotic- and IL-21-supplemented animals after the initiation of therapy (Fig. 4C). Within jejunal tissues, significant differences in the absolute frequencies of polyfunctional TH17 cells were observed from day 84 p.i. to day 274 and 365 p.i. (p=0.031, 0.031) in supplemented animals, in whom a significantly higher absolute frequency of polyfunctional TH17 cells was observed as compared to ARV-only controls at day 182 p.i. (p=0.029). As compared to day 84 p.i., differences were additionally observed among probiotic and IL-21 supplemented animals in the absolute frequency of RB polyfunctional TH17 cells at days 126, 274, and 365 p.i. (p=0.031, all). No significant differences as compared to baseline were observed among the ARV-only controls at any timepoint (Table S1). Thus, probiotic and IL-21 supplementation of SIV-infected, ARV-treated macaques led to a significant improvement in the absolute frequencies of intestinal TH17 cells and in turn, the absolute frequencies of polyfunctional TH17 cells.
Since IL-21 similarly promotes the differentiation and maintenance of IL-17-expressing memory CD8+ T-cells (TC17), we additionally examined the longitudinal relative frequencies of TC17 cells. No significant differences between experimental groups were noted at any timepoint in any tissue examined (Fig. S3C and data not shown). However, a significant improvement in jejunal TC17 cells was noted in the probiotic- and IL-21-supplemented animals at day 274 p.i. as compared to the pre-treatment timepoint (p=0.0469) – no differences were noted in relative RB TC17 frequencies between or among groups. We similarly examined the relative frequencies of polyfunctional TC17 cells. No differences were noted between experimental groups and, again, significant changes were limited to jejunal TC17 cells in the probiotic and IL-21 supplemented animals (Fig. S3D). Significantly higher relative frequencies of polyfunctional, jejunal TC17 cells were noted in these animals at days 274 and 365 p.i. as compared to day 84 p.i. (p=0.031, 0.031). These data indicate that probiotic- and IL-21-supplementation of ARV therapy promotes a recovery of TH17 and TC17 cells during chronic SIV infection in macaques.
IL-21 and Probiotic Supplementation Reduce Microbial Translocation and Dysbiosis in ARV-Treated Macaques
Persistent immune activation, TH17 depletion, residual microbial translocation, and dysbiosis form a synergistic axis in ARV-treated individuals that likely results in the increased morbidities and mortality observed in ARV-treated subjects26. As our results indicate that probiotic and IL-21 supplementation of ARVs promotes the recovery of intestinal TH17 cells (Fig. 4), tempers some markers of immune activation (Fig. S1), and results in better clinical outcomes (Fig. 3), we next considered the possibility that such treatment might further be associated with a reduction in microbial translocation and dysbiosis. We assessed levels of microbial translocation in our animals by immunohistochemical (IHC) staining for Escherichia coli antigens in the colonic lamina propria, MLN paracortex, and liver tissue from samples obtained at necropsy. E. coli antigens were evident in all tissues and only within the liver of probiotic- and IL-21-supplemented animals was there a trend for lower levels of E. coli staining (p=0.0556; Fig. S4 A+B).
To assess whether probiotic and IL-21 supplementation influences the composition of microbial communities, we measured relative levels of commensal Bifidobacterium and Lactobacillus 16S DNA in the stool and rectal tissues of our experimental animals. No significant differences were observed in the ARV-only animals as compared to baseline with regards to Bifidobacterium levels (p=0.0625, all timepoints), but these animals exhibited significantly lower frequencies of Bifidobacterium as compared to probiotic and IL-21 supplemented animals at day 154 p.i. (p=0.0333; Fig. 5A). Importantly, probiotic- and IL-21-treated animals exhibited significant increases in the relative frequency of Bifidobacterium species as compared to the pre-treatment timepoint at days 154 and 365 p.i. (p=0.0156, 0.0313). Supplemented animals additionally exhibited significant increases in Lactobacillus species at day 154 p.i. as compared to the pre-treatment timepoint (p=0.0156; Figure 5B) – no significant differences were noted as compared to or among ARV-only animals (p=0.0625, all timepoints). Thus, probiotic and IL-21 supplementation of ARV-therapy during chronic SIV infection promoted a reduction in distal site microbial translocation as well as an increase in the level of probiotic species within the GI tract, reflecting a significant improvement in disease progression.
Figure 5. IL-21 and Probiotic Therapy Improve Commensal Colonization and Reduce Circulating Kyn.
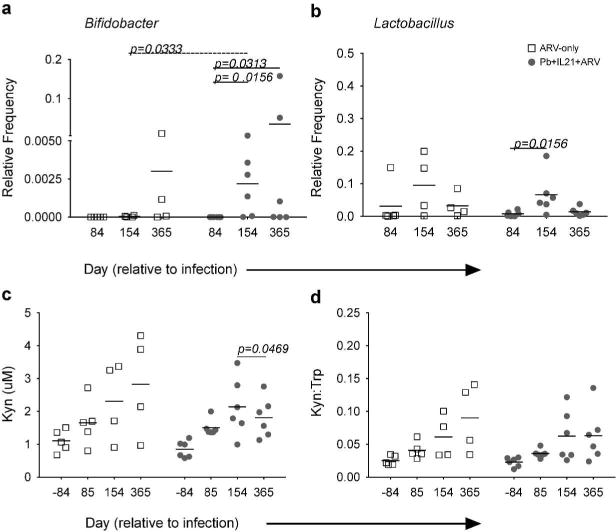
a–b. Relative frequency of Bifidobacter (a) and Lactobacillus (b) 16S DNA of total 16S DNA in rectal tissues at the indicated timepoints. c. Concentration of plasma Kyn (μM) at the indicated days p.i. d. Ratio of plasma Kyn to Trp concentrations at the indicated days p.i. Line through samples represents mean; lines above samples span timepoints of significance. Significance within groups was assessed by Wilcoxon Signed-Rank test. Significance across groups was assessed by Mann-Whitney U test.
Reduced IDO Activity in IL-21-, Probiotic- and ARV-treated Macaques
The persistent depletion of TH17 cells in ARV-treated, HIV-infected individuals correlates with elevated expression and activity of indoleamine-2,3-dioxygenase (IDO), a cytokine-inducible, tryptophan (Trp) catabolizing enzyme16, 34. Trp metabolites are known to inhibit the generation and maintenance of TH17 cells34, and persistently elevated IDO activity has been postulated to contribute to poor TH17 recovery in HIV-infected individuals16, 34 while also predicting poor CD4+ T-cell recovery and increased mortality. To determine if probiotic and IL-21 supplementation reduced IDO expression and activity, we first quantified IDO expression by IHC in colon and mesenteric lymph node (MLN) tissue samples obtained at necropsy. Tissue staining revealed no significant differences in MLN IDO between the two groups and only a trend for reduced colonic IDO in probiotic and IL-21 supplemented animals as compared to ARV-controls (p=0.0857) (Fig S4C+D). We next considered whether TH17 recovery might correspond with a reduction in circulating levels of kynurenine (Kyn), the first immunoactive Trp catabolite generated in the IDO pathway. When we measured the concentration of plasma Kyn, we observed that whereas ARV-only animals exhibited a steady increase in Kyn throughout the study period, probiotic and IL-21 supplemented animals displayed reduced Kyn from day 154 to 365 p.i. (Fig. 5C). Although no significant differences were noted between experimental groups at the timepoints examined, significantly lower Kyn was apparent between these later timepoints within the supplemented group (p=0.0469). To ascertain whether the observed Kyn levels might result from reduced IDO expression and/or activity, we additionally measured the ratio of plasma Kyn:Trp. Similar to the pattern observed for Kyn itself, while ARV-only animals exhibited a steady increase in plasma Kyn:Trp throughout the experimental period, probiotic and IL-21 supplemented animals exhibited an increase only through the final examined pre-necropsy timepoint, day 154 p.i. (Fig. 5D). There was no significant difference between the final two timepoints, however, although supplemented animals exhibited a plateau in Kyn:Trp levels. Thus, probiotic and IL-21 supplementation of ARVs during chronic SIV infection is associated with a trend towards reduction in intestinal IDO and an apparent reduction in circulating levels of the Trp catabolite, Kyn.
Discussion
ARV-treatment significantly prolongs the length and quality of life of HIV-infected individuals; however, disease progression in these individuals remains evident even with decades of effective viral suppression1, 2. In treated HIV-infected individuals, morbidities and mortality have been attributed to a self-reinforcing cycle of microbial translocation and dysbiosis, immune activation, and a persistent, loss of antimicrobial TH17 cells3, 35. While we have previously demonstrated that probiotic supplementation of ARVs in SIV-infected macaques promotes a restoration of intestinal CD4+ T-cells18, in our prior study we did not observe significant TH17 expansion nor was there a measurable impact on microbial translocation or on the composition of the GI tract microbiome. Herein, we demonstrate that the supplementation of ARVs with probiotics and IL-21 promotes intestinal TH17 recovery, a reduction in measures of microbial translocation, and an enhanced microbiome. Importantly, our treatment regimen did not promote immune activation or a significant expansion of the viral reservoir. Although limited in size, these findings suggest that probiotic and IL-21 supplementation of ARV therapy should be further evaluated as a therapeutic in the treatment of non AIDS-related comorbidities.
The development of TH17 cells from naïve CD4+ T-cell precursors or Tregs is augmented by the autocrine production of IL-2119–21. In this capacity, IL-21 enhances RORγt transcription, FOXP3 suppression, and the downstream transcription of IL-17 and additional TH17-associated cytokines such as IL-2219, 20. Importantly, IL-21 alone is insufficient to induce TH17 development21, requiring additional signaling originating from the commensal microbiome. Thus, in murine models, although the presence of the commensal microbiome – specifically, segmented filamentous bacteria – is sufficient to drive the development of TH17 cells in vivo13, 14, an absence or blockade of IL-21 signaling diminishes TH17 differentiation19, 20. That the inclusion of IL-21 in our regimen promotes TH17 reconstitution (Fig. 4) where probiotic supplementation of ARVs alone does not18 indicates that therapeutically provided probiotic species alone are either insufficient or incorrectly parlayed into the development of TH17 cells during chronic SIV infection. We believe that the prolonged regeneration of TH17 cells in our animals subsequent to IL-21 withdrawal (approximately 7 months between early and late administration; Fig. 1) suggests that IL-21 functioned directly at the level of CD4+ T-cells to establish a basal threshold of TH17 cells necessary for self-propagation. Although a direct assessment of circulating IL-21 was not performed, attainment of a theoretical threshold is further supported by the observation that late IL-21 administration did not augment already enhanced TH17 frequencies (Figure 4). To this end, it would be of significant interest to determine whether increasing the concentration of administered IL-21 or lengthening early IL-21 therapy might not augment or accelerate TH17 recovery beyond what we describe here. Although we cannot discount alternative roles for IL-21 in improving disease progression in our supplemented animals – such as altered functionality among IL-21-responsive B-cells and NK-cells36 – the administration of IL-21 during acute SIV infection in macaques leads to a selective maintenance of TH17 cells that is strictly dependent on the continued administration of IL-2123, arguing against an ancillary role for IL-21 in sustaining TH17 frequencies. An assessment of the effects of IL-21 supplementation to ARV therapy independent of probiotic administration will significantly further our understanding of the conditions required for the generation and maintenance of TH17 cells in treated chronic lentiviral infections.
Although de novo TH17 development from naïve CD4+ T-cell precursors is largely reliant on the ability of APCs to integrate microbial sensing into pro-TH17 cytokine signaling10, 11, TH17 and Treg cells exist as a transcriptional dichotomy19, 20, 37, 38 which may further be reciprocally programmed by aryl hydrocarbon receptor (AHR) ligand signaling34, 39. As recent findings have demonstrated a role for the IDO-derived AHR-agonist Kyn in reducing the TH17:Treg during lentiviral infection34, reduced plasma Kyn (Fig. 5C) in our supplemented animals suggests that TH17 recovery may have included mechanisms adjunct to the direct influence of therapeutic IL-21. Given that species of the dysbiotic microbiota encode enzymatic homologues to primate Trp-catabolizing enzymes16, we cannot discount the possibility that an IL-21-mediated recuperation of TH17 cells preempted a reduction of dysbiotic species and hence, Kyn. Indeed, the prolonged recovery of commensal Bifidobacteria in our supplemented animals (Fig. 5B) in contrast to animals receiving probiotic supplementation alone18 suggests a role for IL-21 in promoting the colonization of probiotic species. However, as a replete commensal microbiome is itself necessary for TH17 maintenance14, prolonged TH17 recovery after IL-21 cessation may thus be dependent upon a reduction in the frequency of dysbiotic species.
Microbial translocation has been extensively described as a significant contributor to immune activation and disease progression in chronic lentiviral infections3. In HIV-infected humans and SIV-infected NHPs the influence of microbial translocation on immune activation has been causally associated by the correlation of disseminated LPS with systemic IFNα4, 5, and formally demonstrated by the prevention of immune activation with acute use of the LPS-sequestering drug sevelamer40. Intriguingly, TH17 recovery (Figure 4) and improved disease progression (Figure 3) occurred independent of systemically reduced E. coli antigen (albeit a trend for reduced E. coli in the liver; Figure S4A+B) or persistent immune activation (Figure S1). In this regard, immune activation in our supplemented animals may reflect improved immune function rather than a manifestation of innate cell refractoriness or lymphocyte exhaustion4, 41, 42. Indeed, as compared to ARV-only controls, supplemented animals exhibited control over rectal CD4+ T-cell proliferation (Fig S1A) concomitant with improved CD4+ T-cell reconstitution (Fig. S2), which is suggestive of homeostatic restoration. A more detailed assessment of immune function, particularly alterations in inflammatory biomarkers, will be necessary to address these hypotheses.
Increased mortality among HIV-infected individuals remains evident even with decades of effective viral suppression2, especially in conditions when the IDO-mediated kynurenine pathway is induced and microbial dysbiosis may be present26. Consistent with these observations in humans, co-morbidities were evident in our ARV-only animals where, despite continued viral suppression (Fig. 2), 3 of 5 animals displayed repeated complications. That none of our supplemented animals displayed complications despite co-housing with the ARV-only controls is particularly notable given the propensity of PTMs toward microbial translocation and systemic immune activation, even in the absence of SIV infection43. Although the recovery of TH17 cells in our supplemented animals (Figure 4) likely contributed to improved resistance to co-infection8, 9, an association between TH17 recovery and ARV tolerance is more tenuous. Indeed, ARV complications were reduced in animals supplemented with probiotics alone, where TH17 recovery was absent18. While the presence of commensal bacterial species has been demonstrated to improve the efficacy of anticancer therapeutics such as cyclophosphamide44, 45, the influence of probiotics and commensal species in mediating pharmaceutical toxicities has not been formally explored. A thorough characterization of the contributions of our individual therapeutic constituents – including individual probiotic species – to immune function in the presence and absence of ARVs will be necessary to fully uncover the mechanisms which govern disease progression in treated chronic lentiviral infections.
In summary, we demonstrate that probiotic and IL-21 supplementation of ARV therapy in chronically SIV-infected macaques improves clinical outcomes, as manifested by a reduction in non-AIDS associated morbidities. Importantly, improved clinical outcomes were accompanied with an increase in the absolute frequency of intestinal, polyfunctional TH17 cells, and reduced measures of intestinal microbial dysbiosis. A larger, mechanistic dissection of the immune processes associated with improved disease progression in our supplemented animals will significantly inform the design of future therapeutics for the treatment of HIV-infected individuals.
Methods
Animal Infections and Interventions
Eleven, healthy pigtail macaques (Macaca nemestrina) were accessed for this study and infected with 3,000 TCID50 SIVmac239. At day 98 p.i., all animals began a daily ARV regimen consisting of PMPA and FTC (30 mg/kg at 120 mg/mL in PBS s.c. once-daily) and L’812 (120 mg oral, twice-daily) – PMPA was reduced to 20 mg/kg after 30 days. Of these animals, 6 additionally received daily probiotic VSL#3® (oral, two capsules daily) for the entirety of the study and two courses of rMamu IL-21-IgFc (50 ug/kg at 1 mg/mL in H2O s.c., 5 once-weekly doses) initiated at days 98 and 333 p.i. Animals were assigned to treatment groups to evenly distribute animals by weight and MHC genotype (Mane-A10) and were sampled as a mixed population. The National Institute of Allergy and Infectious Diseases Division of Intramural Research Animal Care and Use Program, as part of the National Institutes of Health (NIH) Intramural Research Program, approved all of the experimental procedures (protocol LMM 12). Animals were sacrificed according to the recommendations set forth by the American Veterinary Medical Association Guidelines on Euthanasia.
Sample Collection
Blood, BAL, and biopsies from colon, liver, lymph nodes, jejunum, and rectum were collected longitudinally or at necropsy. Biopsies were maintained in RPMI prior to processing. For longitudinal LN biopsies, axillary or inguinal LNs were locally exposed and excised by blunt dissection. For longitudinal RBs, fecal material was removed from the rectum and biopsies obtained with biopsy forceps. Jejunal biopsies were obtained by video-guided endoscopy. For BAL collection, silicone tubing was directed into the trachea with the assistance of a laryngoscope, whereupon warmed normal PBS or saline was instilled and subsequently aspirated for collection. Biopsies collected at necropsy were directly accessed immediately post-mortem. Stool and luminal swabs were obtained as previously described17. Plasma was isolated from blood by centrifugation. Mononuclear cells were isolated from blood by Ficoll gradient centrifugation and from BAL and tissue biopsies by straining/grinding samples through a 0.22 μm cell strainer.
Determination of vRNA and vDNA
Plasma vRNA levels were determined by qRT-PCR (ABI Prism 7900 sequence detection system; Applied Biosystems). vRNA was reverse transcribed and cDNA amplified (45 cycles/default setting) with Ampli Taq Gold DNA polymerase (PCR core reagents kit; Perkin-Elmer/Roche) utilizing primer pairs corresponding to SIVmac239 gag gene sequences (forward, nucleotides 1181 to 1208, and reverse, nucleotides 1338 to 1317). Cell-associated viral DNA was measured as previously46. Briefly, qPCR was performed on sorted, lysed cells using the Taq DNA polymerase kit (Invitrogen) and the SIVmac239 forward primer GTCTGCGTCATYTGGTGCATTC, reverse primer CACTAGYTGTCTCTGCACTATRTGTTTTG, and probe sequence CTTCRTCAGTYTGTTTCACTTTCTCTTCTGCG. Results assessed below the limit of detection were normalized to the threshold cutoff.
Flow Cytometry and Immunophenotypic Analysis
Polychromatic flow cytometry and cell sorting were performed on stained mononuclear cells utilizing the BD LSRFortessa and FACS Aria, respectively, equipped with FACS DiVA software. The following Abs were used for staining at predetermined concentrations: CD3 (clone SP34-2) AL700, HLADR (G46-6) APC-H7, IL-2 (MQ1-17H12) FITC, and Ki67 (B56) FITC from BD; CD4 (OKT4) eFluor450, CD8 (SK1) PerCP-e710, IFNg (4S.B3) eFluor450, IL17A (eBio64DEC17) PE, IL22 (IL22JOP) APC, and TNFa (MAb11) PE-Cy7 from eBioscience; CD28 (CD28.2) ECD from Beckman Coulter; and CD95 (DX2) PE-Cy5 from Biolegend. Cell viability was assessed using the Live/Dead Aqua Fixable Dead Cell Stain (Invitrogen). Cells were permeabilized with Cytofix/Cytoperm (BD) prior to intracellular staining. Acquired data were analyzed using FlowJo software. To quantify cell subset frequencies, we used a threshold cutoff of 100 cells of the parent population. For TH17 functional analyses, cytokine gates were established on a threshold cutoff of 100 CD4+ or CD8+ TM and Boolean gates utilized to ascertain frequencies – populations not meeting this threshold were not considered for analysis.
IHC and quantitative image analysis
IHC for rabbit polyclonal anti-E. coli (Cat. # B0357; Dako) and rabbit monoclonal anti-IDO (Millipore; clone EPR1230Y) was performed using a biotin-free polymer approach (Rabbit Polink-1 for E. coli and Mouse Polink-2 for IDO, Golden Bridge International, Inc.) on 5-mm tissue sections mounted on glass slides, which were dewaxed and rehydrated with double-distilled H2O. Antigen retrieval was performed by heating slides in 1× DIVA Decloacker reagent (Biocare Medical) or 0.01% citraconic anhydride containing 0.05% Tween-20 in a pressure cooker (122°C for 30sec). Slides were stained with optimal conditions determined empirically on an IntelliPATH autostainer (Biocare Medical) that consisted of a blocking step using blocking buffer (TBS with 0.05% Tween-20 and 0.5% casein) for 10min and an endogenous peroxidase block using 1.5% (v/v) H2O2 in TBS (pH 7.4) for 10min. Primary Abs were diluted 1:5,000 in blocking buffer and incubated for 1h at room temperature. Tissue sections were washed, and either Rabbit Polink-1 or Polink-2 staining systems (Golden Bridge International, Inc) were applied for 30min at room temperature. Sections were developed with Impact™ 3,3′-diaminobenzidine (Vector Laboratories), counterstained with hematoxylin and mounted in Permount (Fisher Scientific). All stained slides were scanned at high magnification (400×) using the Aperio® AT2 System (Leica Biosystems) yielding high-resolution data for the entire tissue section. Representative high magnification (400×) images were acquired from these whole tissue scans.
Quantification of systemic inflammatory markers
Concentrations of Kyn and Trp were quantified from plasma by liquid-chromatography-tandem mass spectrometry, as previously described34. Levels of LBP and sCD14 were assessed from plasma using commercially available ELISA kits (Antibodies Online and R&D, respectively), according to the manufacturers’ protocols.
Assessment of bacterial 16S DNA
16S DNA was isolated and quantified from stool or from luminal swabs as previously described17.
Statistics
Statistical analyses were performed using Prism (v6.0; GraphPad Software Inc.). The MannWhitney U test was used for comparisons between groups; the Wilcoxon Signed-Rank test for within groups. The Mantel-Cox test was used to assess disease progression as measured by time to the first presentation of health complication. Averaged data are presented as arithmetic mean ± SEM. p values less than 0.05 was considered significant.
Supplementary Material
Acknowledgments
We thank F. Villinger (Emory University) for providing the IL-21; D. Hazuda (Gilead), M. Miller (Merck) and J. Lifson (NCI) for providing the ARVs; C. de Simon (Sigma Tau Pharmaceuticals) for providing the VSL#3; H. Cronise, J. Swerczek and R. Herbert for veterinary assistance; and the NIAID LMM-Core for assessing viral loads. Funding for this study was provided in part by the Division of Intramural Research/NIAID/NIH. The content of this publication does not necessarily reflect the views or policies of DHHS, nor does the mention of trade names, commercial products, or organizations imply endorsement by the U.S. Government.
Footnotes
DISCLOSURE
The authors declare no known conflicts of interest.
References
- 1.Lederman MM, Funderburg NT, Sekaly RP, Klatt NR, Hunt PW. Residual immune dysregulation syndrome in treated HIV infection. Advances in immunology. 2013;119:51–83. doi: 10.1016/B978-0-12-407707-2.00002-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Volberding PA, Deeks SG. Antiretroviral therapy and management of HIV infection. Lancet. 2010;376(9734):49–62. doi: 10.1016/S0140-6736(10)60676-9. [DOI] [PubMed] [Google Scholar]
- 3.Marchetti G, Tincati C, Silvestri G. Microbial translocation in the pathogenesis of HIV infection and AIDS. Clinical microbiology reviews. 2013;26(1):2–18. doi: 10.1128/CMR.00050-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Brenchley JM, Price DA, Schacker TW, Asher TE, Silvestri G, Rao S, et al. Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Nat Med. 2006;12(12):1365–1371. doi: 10.1038/nm1511. [DOI] [PubMed] [Google Scholar]
- 5.Estes JD, Harris LD, Klatt NR, Tabb B, Pittaluga S, Paiardini M, et al. Damaged intestinal epithelial integrity linked to microbial translocation in pathogenic simian immunodeficiency virus infections. PLoS pathogens. 2010;6(8):e1001052. doi: 10.1371/journal.ppat.1001052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Favre D, Lederer S, Kanwar B, Ma ZM, Proll S, Kasakow Z, et al. Critical loss of the balance between Th17 and T regulatory cell populations in pathogenic SIV infection. PLoS Pathog. 2009;5(2):e1000295. doi: 10.1371/journal.ppat.1000295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Brenchley JM, Paiardini M, Knox KS, Asher AI, Cervasi B, Asher TE, et al. Differential Th17 CD4 T-cell depletion in pathogenic and nonpathogenic lentiviral infections. Blood. 2008;112(7):2826–2835. doi: 10.1182/blood-2008-05-159301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Raffatellu M, Santos RL, Verhoeven DE, George MD, Wilson RP, Winter SE, et al. Simian immunodeficiency virus-induced mucosal interleukin-17 deficiency promotes Salmonella dissemination from the gut. Nat Med. 2008;14(4):421–428. doi: 10.1038/nm1743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Peck A, Mellins ED. Precarious balance: Th17 cells in host defense. Infection and immunity. 2010;78(1):32–38. doi: 10.1128/IAI.00929-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Veldhoen M, Hocking RJ, Atkins CJ, Locksley RM, Stockinger B. TGFbeta in the context of an inflammatory cytokine milieu supports de novo differentiation of IL-17-producing T cells. Immunity. 2006;24(2):179–189. doi: 10.1016/j.immuni.2006.01.001. [DOI] [PubMed] [Google Scholar]
- 11.Goto Y, Panea C, Nakato G, Cebula A, Lee C, Diez MG, et al. Segmented filamentous bacteria antigens presented by intestinal dendritic cells drive mucosal Th17 cell differentiation. Immunity. 2014;40(4):594–607. doi: 10.1016/j.immuni.2014.03.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Yang Y, Torchinsky MB, Gobert M, Xiong H, Xu M, Linehan JL, et al. Focused specificity of intestinal TH17 cells towards commensal bacterial antigens. Nature. 2014;510(7503):152–156. doi: 10.1038/nature13279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ivanov II, Atarashi K, Manel N, Brodie EL, Shima T, Karaoz U, et al. Induction of intestinal Th17 cells by segmented filamentous bacteria. Cell. 2009;139(3):485–498. doi: 10.1016/j.cell.2009.09.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ivanov II, Frutos Rde L, Manel N, Yoshinaga K, Rifkin DB, Sartor RB, et al. Specific microbiota direct the differentiation of IL-17-producing T-helper cells in the mucosa of the small intestine. Cell host & microbe. 2008;4(4):337–349. doi: 10.1016/j.chom.2008.09.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Littman DR, Pamer EG. Role of the commensal microbiota in normal and pathogenic host immune responses. Cell host & microbe. 2011;10(4):311–323. doi: 10.1016/j.chom.2011.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Vujkovic-Cvijin I, Dunham RM, Iwai S, Maher MC, Albright RG, Broadhurst MJ, et al. Dysbiosis of the gut microbiota is associated with HIV disease progression and tryptophan catabolism. Science translational medicine. 2013;5(193):193ra191. doi: 10.1126/scitranslmed.3006438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Klase Z, Ortiz A, Deleage C, Mudd JC, Quinones M, Schwartzman E, et al. Dysbiotic bacteria translocate in progressive SIV infection. Mucosal immunology. 2015 doi: 10.1038/mi.2014.128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Klatt NR, Canary LA, Sun X, Vinton CL, Funderburg NT, Morcock DR, et al. Probiotic/prebiotic supplementation of antiretrovirals improves gastrointestinal immunity in SIV-infected macaques. J Clin Invest. 2013;123(2):903–907. doi: 10.1172/JCI66227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Korn T, Bettelli E, Gao W, Awasthi A, Jager A, Strom TB, et al. IL-21 initiates an alternative pathway to induce proinflammatory T(H)17 cells. Nature. 2007;448(7152):484–487. doi: 10.1038/nature05970. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Nurieva R, Yang XO, Martinez G, Zhang Y, Panopoulos AD, Ma L, et al. Essential autocrine regulation by IL-21 in the generation of inflammatory T cells. Nature. 2007;448(7152):480–483. doi: 10.1038/nature05969. [DOI] [PubMed] [Google Scholar]
- 21.Yeste A, Mascanfroni ID, Nadeau M, Burns EJ, Tukpah AM, Santiago A, et al. IL-21 induces IL-22 production in CD4+ T cells. Nature communications. 2014;5:3753. doi: 10.1038/ncomms4753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Micci L, Cervasi B, Ende ZS, Iriele RI, Reyes-Aviles E, Vinton C, et al. Paucity of IL-21-producing CD4(+) T cells is associated with Th17 cell depletion in SIV infection of rhesus macaques. Blood. 2012;120(19):3925–3935. doi: 10.1182/blood-2012-04-420240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Pallikkuth S, Micci L, Ende ZS, Iriele RI, Cervasi B, Lawson B, et al. Maintenance of intestinal Th17 cells and reduced microbial translocation in SIV-infected rhesus macaques treated with interleukin (IL)-21. PLoS Pathog. 2013;9(7):e1003471. doi: 10.1371/journal.ppat.1003471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ma HL, Whitters MJ, Konz RF, Senices M, Young DA, Grusby MJ, et al. IL-21 activates both innate and adaptive immunity to generate potent antitumor responses that require perforin but are independent of IFN-gamma. Journal of immunology. 2003;171(2):608–615. doi: 10.4049/jimmunol.171.2.608. [DOI] [PubMed] [Google Scholar]
- 25.Pallikkuth S, Rogers K, Villinger F, Dosterii M, Vaccari M, Franchini G, et al. Interleukin-21 administration to rhesus macaques chronically infected with simian immunodeficiency virus increases cytotoxic effector molecules in T cells and NK cells and enhances B cell function without increasing immune activation or viral replication. Vaccine. 2011;29(49):9229–9238. doi: 10.1016/j.vaccine.2011.09.118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Byakwaga H, Boum Y, 2nd, Huang Y, Muzoora C, Kembabazi A, Weiser SD, et al. The kynurenine pathway of tryptophan catabolism, CD4+ T-cell recovery, and mortality among HIV-infected Ugandans initiating antiretroviral therapy. The Journal of infectious diseases. 2014;210(3):383–391. doi: 10.1093/infdis/jiu115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Gaston MA. Enterobacter: an emerging nosocomial pathogen. The Journal of hospital infection. 1988;11(3):197–208. doi: 10.1016/0195-6701(88)90098-9. [DOI] [PubMed] [Google Scholar]
- 28.Perazella MA. Acute renal failure in HIV-infected patients: a brief review of common causes. The American journal of the medical sciences. 2000;319(6):385–391. doi: 10.1097/00000441-200006000-00008. [DOI] [PubMed] [Google Scholar]
- 29.Alcantara S, Reece J, Amarasena T, Rose RD, Manitta J, Amin J, et al. Thrombocytopenia is strongly associated with simian AIDS in pigtail macaques. J Acquir Immune Defic Syndr. 2009;51(4):374–379. doi: 10.1097/QAI.0b013e3181a9cbcf. [DOI] [PubMed] [Google Scholar]
- 30.Klatt NR, Estes JD, Sun X, Ortiz AM, Barber JS, Harris LD, et al. Loss of mucosal CD103+ DCs and IL-17+ and IL-22+ lymphocytes is associated with mucosal damage in SIV infection. Mucosal immunology. 2012;5(6):646–657. doi: 10.1038/mi.2012.38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Almeida JR, Price DA, Papagno L, Arkoub ZA, Sauce D, Bornstein E, et al. Superior control of HIV-1 replication by CD8+ T cells is reflected by their avidity, polyfunctionality, and clonal turnover. J Exp Med. 2007;204(10):2473–2485. doi: 10.1084/jem.20070784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Betts MR, Nason MC, West SM, De Rosa SC, Migueles SA, Abraham J, et al. HIV nonprogressors preferentially maintain highly functional HIV-specific CD8+ T cells. Blood. 2006;107(12):4781–4789. doi: 10.1182/blood-2005-12-4818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Brenchley JM, Knox KS, Asher AI, Price DA, Kohli LM, Gostick E, et al. High frequencies of polyfunctional HIV-specific T cells are associated with preservation of mucosal CD4 T cells in bronchoalveolar lavage. Mucosal immunology. 2008;1(1):49–58. doi: 10.1038/mi.2007.5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Favre D, Mold J, Hunt PW, Kanwar B, Loke P, Seu L, et al. Tryptophan catabolism by indoleamine 2,3-dioxygenase 1 alters the balance of TH17 to regulatory T cells in HIV disease. Science translational medicine. 2010;2(32):32ra36. doi: 10.1126/scitranslmed.3000632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Kanwar B, Favre D, McCune JM. Th17 and regulatory T cells: implications for AIDS pathogenesis. Current opinion in HIV and AIDS. 2010;5(2):151–157. doi: 10.1097/COH.0b013e328335c0c1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Spolski R, Leonard WJ. Interleukin-21: a double-edged sword with therapeutic potential. Nature reviews Drug discovery. 2014;13(5):379–395. doi: 10.1038/nrd4296. [DOI] [PubMed] [Google Scholar]
- 37.Bettelli E, Carrier Y, Gao W, Korn T, Strom TB, Oukka M, et al. Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature. 2006;441(7090):235–238. doi: 10.1038/nature04753. [DOI] [PubMed] [Google Scholar]
- 38.Zhou L, Lopes JE, Chong MM, Ivanov II, Min R, Victora GD, et al. TGF-beta-induced Foxp3 inhibits T(H)17 cell differentiation by antagonizing RORgammat function. Nature. 2008;453(7192):236–240. doi: 10.1038/nature06878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Quintana FJ, Basso AS, Iglesias AH, Korn T, Farez MF, Bettelli E, et al. Control of T(reg) and T(H)17 cell differentiation by the aryl hydrocarbon receptor. Nature. 2008;453(7191):65–71. doi: 10.1038/nature06880. [DOI] [PubMed] [Google Scholar]
- 40.Kristoff J, Haret-Richter G, Ma D, Ribeiro RM, Xu C, Cornell E, et al. Early microbial translocation blockade reduces SIV-mediated inflammation and viral replication. J Clin Invest. 2014;124(6):2802–2806. doi: 10.1172/JCI75090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Lichterfeld M, Kaufmann DE, Yu XG, Mui SK, Addo MM, Johnston MN, et al. Loss of HIV-1-specific CD8+ T cell proliferation after acute HIV-1 infection and restoration by vaccine-induced HIV-1-specific CD4+ T cells. J Exp Med. 2004;200(6):701–712. doi: 10.1084/jem.20041270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Appay V, Nixon DF, Donahoe SM, Gillespie GM, Dong T, King A, et al. HIV-specific CD8(+) T cells produce antiviral cytokines but are impaired in cytolytic function. J Exp Med. 2000;192(1):63–75. doi: 10.1084/jem.192.1.63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Klatt NR, Harris LD, Vinton CL, Sung H, Briant JA, Tabb B, et al. Compromised gastrointestinal integrity in pigtail macaques is associated with increased microbial translocation, immune activation, and IL-17 production in the absence of SIV infection. Mucosal immunology. 2010;3(4):387–398. doi: 10.1038/mi.2010.14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Viaud S, Saccheri F, Mignot G, Yamazaki T, Daillere R, Hannani D, et al. The intestinal microbiota modulates the anticancer immune effects of cyclophosphamide. Science. 2013;342(6161):971–976. doi: 10.1126/science.1240537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Iida N, Dzutsev A, Stewart CA, Smith L, Bouladoux N, Weingarten RA, et al. Commensal bacteria control cancer response to therapy by modulating the tumor microenvironment. Science. 2013;342(6161):967–970. doi: 10.1126/science.1240527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Brenchley JM, Vinton C, Tabb B, Hao XP, Connick E, Paiardini M, et al. Differential infection patterns of CD4+ T cells and lymphoid tissue viral burden distinguish progressive and nonprogressive lentiviral infections. Blood. 2012;120(20):4172–4181. doi: 10.1182/blood-2012-06-437608. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.