

Abstract
Long-term exposures to reactive oxygen species (ROS) have been linked to scores of chronic diseases. This has motivated interest in oxidation products of the only free cysteine residue (Cys34) of human serum albumin (HSA) as possible biomarkers of ROS exposure. However, Cys34 oxidation products have not been detected in human serum or plasma. Using liquid chromatography-high resolution tandem mass spectrometry, we report accurate masses and molecular compositions of Cys34 oxidation products in the 2432 Da peptide resulting from tryptic digestion of HSA. Peptides containing the expected sulfinic (Cys-SO2H) and sulfonic (Cys-SO3H) acids, as well as an adduct representing addition of one oxygen atom and loss of two hydrogen atoms, were detected in four archived samples of human plasma and one fresh sample of human serum. We speculate that this latter adduct is a sulfinamide formed by intramolecular reaction between either the Cys34 sulfenic acid (Cys-SOH) or sulfinic acid (Cys-SO2H) and the adjacent glutamine residue (Gln33). All three Cys34 adducts were measured in the five human samples with levels decreasing in the order sulfinic acid > (proposed) sulfinamide > sulfonic acid. Parallel measurements of a negative control detected only small amounts of the Cys34 sulfonic acid and the (proposed) sulfinamide and did not detect the sulfinic acid.
Keywords: reactive oxygen species, HSA, Cys34, high resolution mass spectrometry, sulfinic acid, sulfonic acid, sulfinamide
Graphical Abstract
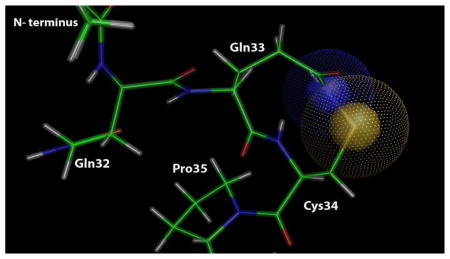
Grigoryan et al. used high-resolution mass spectrometry to detect oxidation products of HSA-Cys34 in human plasma samples. In addition to the sulfinic acid (Cys-SO2H) and the sulfonic acid (Cys-SO3H), the authors detected an added mass of (+O,-H2), which they suspect to be a sulfinamide resulting from intramolecular reaction between the Cys34 sulfenic acid (Cys-SOH) or sulfinic acid (Cys-SO2H) and the amide group of Gln33.
INTRODUCTION
Because they damage DNA and functional proteins, reactive oxygen species (ROS) have long been implicated as causes of chronic diseases. (1, 2) In fact, a recent query of the National Library of Medicine database with search terms “reactive oxygen species”, “disease” and “review” returned more than 3,000 citations. The class of ROS includes 7 free radicals (superoxide, hydroxyl, hydroperoxyl, peroxyl, alkoxyl, carbonate and carbon dioxide radicals) and 7 nonradical species (hydrogen peroxide, hypobromous acid, hypochlorous acid, ozone, singlet oxygen, organic peroxides and peroxynitrous acid), most of which are produced in the body from endogenous processes, including oxidative metabolism, Fenton chemistry (H2O2 + Fe2+), redox cycling, inflammation and infection. Production of ROS also promotes peroxidation of membrane lipids, which alters membrane function and generates reactive aldehydes that contribute to cell damage. (1, 2)
Because ROS are strong electrophiles, they react with nucleophilic loci in the blood to form adducts. Of the many nucleophilic loci in the blood, the only free cysteine residue of human serum albumin (HSA), i.e. Cys34, has received particular attention because of its strong nucleophilicity and because HSA is abundant in human blood (44 mg/mL) and has a relatively long residence time (about one month). (3) Indeed, Cys34 is the most important scavenger of ROS in the cytosol, accounting for as much as 80 percent of the antioxidant capacity (4). Some of the reactivity of Cys34 towards ROS can be traced to the unusually low pKa value of 6.55 for Cys34, compared to about 8.0–8.5 for thiols in most other proteins and peptides. (3, 5) At a physiological pH of 7.4 about 80% of Cys34 should be present in the strongly nucleophilic thiolate form.
Reactions of Cys34 with ROS have been extensively reviewed. (6, 7) The formation of Cys34 sulfenic acid (Cys-SOH) has been proposed as a transient intermediate in the further oxidation to the stable Cys34 sulfinic (Cys-SO2H) and sulfonic (Cys-SO3H) acids and in formation of mixed disulfides with low molecular weight thiols such as cysteine or glutathione. However, the unstable sulfenic acid of Cys34 has been detected only by indirect trapping methods. (7) In contrast, the Cys34 sulfinic and sulfonic acids have been measured in tryptic digests of HSA that had been modified by ROS in vitro (8, 9) but apparently not in samples of HSA modified in vivo.
Using a novel application of triple-quadrupole mass spectrometry (TQ-MS), referred to as fixed-step selected reaction monitoring (SRM), we reported profiles of unknown adducts of HSA-Cys34 in human samples. (10) After digestion, all Cys34 adducts were found in the 3rd largest tryptic peptide of HSA, designated as T3 (with 21 amino acids of sequence ALVLIAFAQYLQQC34PFEDHVK and a mass of 2432 Da). Adduct profiles detected an average of 66 putative Cys34 adducts in tryptic digests of 6 archived specimens of pooled HSA. (10) The more abundant adducts observed in these samples represented added masses below 100 Da, with one major analyte in the mass range of 31.6 ± 2.3 Da suggestive of the Cys34 sulfinic acid (Cys-SO2H, added mass = 32 Da).
Whereas protein sulfinic acids were initially thought to represent artifacts of purification, it is becoming increasingly clear that they are produced by numerous proteins in vivo in response to ROS. (11) Indeed, Jacob et al. (11) noted that “… sulfinic acids can be seen as an almost ideal sensor for OS [oxidative stress], since they are formed in the presence of a range of reactive oxygen and nitrogen species.” Thus, if confirmed as the major adduct detected in our earlier work, the Cys34 sulfinic acid could be useful for evaluating integrated exposures to ROS in human subjects.
Here we describe experiments using liquid-chromatography high-resolution tandem-mass spectrometry (LC-HR-MS/MS) to obtain accurate masses and elemental compositions of putative oxidation products of Cys34. With a mass accuracy of less than 2 ppm we observed Cys34 adducts representing added masses of (+O, -H2), (+O2) and (+O3). While the latter two adducts are the familiar Cys34 sulfinic and sulfonic acids, respectively, the first is consistent with a novel sulfinamide produced by an intramolecular reaction involving oxidized Cys34 and the adjacent Gln33. Thus, in addition to confirming the presence of oxidized forms of Cys34 in human plasma, this work also highlights the power of LC-HR-MS/MS for pointing to hitherto unknown protein modifications produced in vivo.
EXPERIMENTAL SECTION
Chemicals and Reagents
Acetic acid, acetonitrile, ammonium sulfate, calcium chloride, formic acid, iodoacetamide (IAA), methanol, hydrochloric acid, sodium chloride, phosphate buffered saline, dimethylsulfoxide (DMSO), tris-HCl buffer, and tris(2-carboxyethyl)phosphine (TCEP) were from Fisher Scientific (Pittsburgh, PA). Trifluroacetic acid (TFA), hydrogen peroxide and porcine trypsin were from Sigma-Aldrich (St. Louis, MO). Water was purified to a resistivity of 18.2 M3cm (at 25 °C) using a Milli-Q Gradient ultrapure water purification system (Millipore, Billerica, MA). Synthetic T3 peptide was custom synthesized by Biomatik (Wilmington, DE). Isotopically-modified T3 (iT3) with sequence AL-[15N, 13C-Val]-LIAFAQYLQQCPFEDH-[15N, 13C-Val]-K was custom synthesized by BioMer Technology (Pleasanton, CA). Reagents for the bicinchoninic acid assay (BCA assay) were from the micro BCA Assay Kit (Thermo-Fisher Scientific, Rockford, IL).
Synthesis of the Internal Standard (IAA-iT3)
A stock solution was prepared by dissolving synthetic iT3 in DMSO at 5 mM, followed by dilution with water to a final concentration of 0.1 mM. A carboxyamidomethylated Cys34 adduct of iT3 (IAA-iT3) was prepared by adding IAA to 0.5 mL of 0.1 mM iT3 DMSO/water stock solution to a final IAA concentration of 100 mM. The reaction mixture was incubated at 25 °C in the dark for 3 h with continuous agitation. The yield of IAA-iT3 was determined by SRM using TQ-MS (TSQ Quantum Ultra, Thermo Fisher Scientific, Waltham, MA) to detect ions corresponding to T3 (average m/z = 812.3, +3) and IAA-iT3 (average m/z = 835.3, +3). Following HPLC purification, a stock solution of IAA-iT3 was obtained at 20 μM in 35% acetonitrile, 0.1% formic acid, 0.01% TFA.
Synthesis of Reference T3 Adducts
Adducts representing reactions between Cys34 of the T3 peptide and several quinones (1,4-benzoquinone, 1,4-naphthoquinone, 1,2-phenanthrone, and an isomeric mixture of estrogen quinones) were prepared and purified as previously described. (10) These adducts were used to optimize chromatographic and mass spectrometric conditions. A mixture containing oxidation products of Cys34 was prepared by adding 3 μL of aqueous 30% H2O2 to 0.5 mL of 0.1 mM aqueous T3 in a sealed vial and incubating at room temperature for 10 h with gentle mixing.
Human Plasma and Serum Samples
Blood was obtained (in heparin) from healthy adult subjects with informed consent in a previous study. (12) Immediately after collection, red blood cells were removed by centrifugation and washed with an equal volume of phosphate-buffered saline, which was combined with the plasma. Samples of diluted plasma were stored at −80 °C for 13 y prior to the current investigation. After thawing, specimens were anonomyzed by pooling aliquots from 4 or 5 subjects based on race, gender and smoking status. (Pooling was performed to ensure the anonymity of subjects. The four pooled samples used in the current investigation represent African-American male and female subjects who were either smokers or nonsmokers.) Aliquoted pooled samples were stored at −80 °C for approximately three months prior to processing and thus underwent a total of two freeze-thaw cycles.
For comparison, a fresh blood sample was obtained by venipuncture from a volunteer subject who was an Asian male nonsmoker. Following centrifugation of this sample to remove red blood cells, the serum was immediately frozen and stored at −80 °C for one month prior to analysis (one freeze-thaw cycle).
The total protein concentration of each plasma or serum sample was determined by the BCA assay.
Purification of HSA and Enrichment of Cys34 Adducts
After thawing, 200 μL of plasma or serum was transferred to a 1.6-mL micro-centrifuge tube. Three hundred μL of saturated ammonium sulfate was added to precipitate the immunoglobulins over 30 min at room temperature with gentle agitation. The tube was centrifuged at 10,000 × g for 20 min to remove precipitates and the supernatant was transferred to a Vivaspin 15R centrifugal filter with a molecular weight cutoff of 10,000 Da (Sartorius, Goettingen, Germany) for buffer exchange. The protein contents, as determined by the BCA assay, werebetween 1.5 and 2.3 mg (mean = 2.0 mg, SD = 0.34 mg) for the archived samples and 2.4 mg for the fresh sample; these quantities were regarded as those of HSA for subsequent calculations. Sufficient enrichment buffer (100 mM tris-HCl, 0.5 M NaCl, pH 7.5) was added to bring the volume to 15 mL and the spin filter was centrifuged at 3,000 × g for 20 min at 20°C. After discarding the flow-through solution, the procedure was repeated two times. The protein solution in the upper compartment (0.3 – 0.5 mL) was transferred to a centrifuge filter tube with 3-mL capacity and 0.45-μm pore size (VectaSpin 3, Whatman) which had been pre-rinsed with enrichment buffer. Samples were enriched for HSA-Cys34 adducts by mixing with a thiol affinity resin to remove unadducted HSA as described by Funk et al. (13) After incubation overnight with continuous rotation, the mixture was centrifuged and the flow-through fraction that contained HSA adducts was removed and buffer-exchanged to digestion buffer (50 mM tris-HCl, pH 8.0) as described above. The enriched sample from the upper compartment (0.3 – 0.5 mL) was transferred to a 1.6-mL low-retention micro-centrifuge tube. The upper compartment was washed with 0.2 – 0.3 mL of digestion buffer which was added to the recovered sample.
Digestion of Enriched HSA
Each adduct-enriched HSA sample was diluted with 50 mM tris-HCl buffer (pH 8.0) containing 10 % methanol to a volume of 510 μL. To reduce disulfide bonds, 200 mM TCEP was added to give a final concentration of 2 mM TCEP. The mixture was incubated at 37 °C for 15 min with gentle shaking. A volume of 13.8 μL of 200 mM CaCl2 was added to stimulate trypsin activity. Trypsin was added at a 1:10 ratio (trypsin:protein, w/w). The final volume of the digestion mixture was 550 μL. Samples were transferred to disposable polypropylene PULSE tubes and digested at 37 °C in a pressurized system (Barocycler NEP2320, Pressure Biosciences Inc., South Easton, MA), with pressure cycled between ambient (15 s) and 20,000 psi (45 s) for 30 min. Following digestion, samples were transferred to 1.5-mL silanized polypropylene Eppendorf tubes (Fisher, Pittsburg, PA) and spiked with 3.2 – 3.5 μL of IAA-iT3 solution (60 – 70 pmol of internal standard).
Off-line HPLC Fractionation of T3 Adducts
Immediately following digestion, T3 and its Cys34 adducts were fractionated by HPLC (1100 Series, Agilent Technologies, Santa Clara, CA) using a UV-Vis detector at 280 nm. Injections were carried out with an autosampler (1200 Series, Agilent Technologies, Santa Clara, CA), equipped with a 1-mL stainless steel sample loop, and fractions containing T3 peptides were collected using an analytical fraction collector (1200 Series, Agilent Technologies, Santa Clara, CA) that had been calibrated with Delay Sensor Calibrant (Agilent Technologies, Santa Clara, CA). The HSA tryptic digest was separated on a C8 analytical column (300SB Zorbax, 50 × 4.6 mm, 3.5-μm particle size, 300-Å pore size, Agilent Technologies) equipped with a C8 guard cartridge (12.5×4.6 mm, 3.5-μm particle size, 300-Å pore size, Agilent Technologies). Buffer A was H2O/0.1% formic acid/0.01% TFA (v/v/v) and buffer B was acetonitrile/0.1% formic acid/0.01% TFA (v/v/v). About 500 μL of the tryptic digest, containing 70 pmol of internal standard, was injected into the system at a flow rate of 500 μL/min. The elution gradient was as follows: 15 % B initial condition; linear gradient from 15 % B to 37 % B in 5 min; linear gradient from 30 % B to 75 % B in 0.1 min; linear gradient from 75 % B to 95 % B in 1.9 min; and hold at 95 % B for 3 min. Prior injections of adducted T3 peptides (Cys34 modified with 1,4-benzoquinone, 1,4-naphthoquinone, 1,2-phenanthrone, and an isomeric mixture of estrogen quinones) were used to identify the elution times and collection windows for the T3-adduct fractions (11.8 to 13.2 min, total volume 0.7 mL). The final concentration of the internal standard in the collected fractions was 100 nM. Eluates were stored in liquid nitrogen prior to mass spectrometry.
Use of Synthetic T3 as a Negative Control
A stock solution of T3 was prepared by dissolving synthetic T3 in DMSO at 5 mM, followed by dilution with methanol to a final concentration of 0.1 mM. One hundred μL of T3 stock solution (equivalent to 0.66 mg HSA) was diluted with 0.5 mL of freshly degassed enrichment buffer (0.1 M Tris-HCl, 0.5 mM NaCl) and the solution was incubated overnight at room temperature (without thiol affinity resin). After incubation, the solution was carried through the trypsin-digestion protocol as described above (without the buffer exchange step). Three μL of 20 μM IAA-iT3 internal standard (60 pmol) was added to the sample after the digestion step. The sample was fractionated by HPLC and 600-μL of the T3 fraction containing IAA-iT3 at 100 nM was collected. This fraction was stored at −80 °C prior to mass spectrometry.
Liquid Chromatography-Tandem Mass Spectrometry of T3 Adducts
HPLC fractions of T3 and its adducts from the plasma and serum samples were analyzed using an Agilent 1200 liquid chromatograph (LC; Santa Clara, CA) that was connected in-line with an LTQ Orbitrap XL hybrid mass spectrometer equipped with an Ion Max electrospray ionization source (ESI; Thermo Fisher Scientific, Waltham, MA).
The LC employed a reverse-phase, C8 analytical column (300SB Zorbax, 1.0 × 150 mm, 3.5-μm particles, 300-Å pore size, Agilent) and a 100-μL sample loop. Solvent A was H2O/0.1% formic acid/0.01% TFA (v/v/v) and solvent B was acetonitrile/0.1% formic acid/0.01% TFA (v/v/v). Sample solutions, contained in autosampler vials sealed with septa caps, were loaded into the autosampler compartment prior to analysis. An injection volume of 20 μL (partial loop) was used for each sample. The elution program consisted of a linear gradient from 20 % to 55 % B over 45 min, a linear gradient to 95 % B over 0.1 min, isocratic conditions at 95 % B for 14.9 min, a linear gradient to 15 % B over 0.1 min, and isocratic conditions at 15 % B for 19.9 min, at a flow rate of 80 μL/min. The column and sample compartments were maintained at 25 °C and 4 °C, respectively. Solvent (H2O) blanks were run between samples, and the injection needle was rinsed with methanol after each injection to avoid cross-contamination.
Connections between the column exit and the ESI probe of the mass spectrometer were made using PEEK tubing (0.005-in. inner diameter × 1/16-in outer diameter, Agilent). External mass calibration was performed prior to analysis using the standard Thermo LTQ calibration mixture containing caffeine, the peptide MRFA, and Ultramark 1621 dissolved in 51% acetonitrile/25% methanol/23% water/1% acetic acid (v/v). The ESI source parameters were as follows: ion transfer tube temperature 275 °C, normalized sheath gas (nitrogen) flow rate 15%, ESI voltage 2.5 kV, ion transfer tube voltage 35 V, and tube lens voltage 110 V. The ESI emitter tip was positioned approximately 7 mm from the entrance to the ion transfer tube. Mass spectra were acquired in the positive ion mode over the range m/z = 500 to 1500 using the Orbitrap mass analyzer, in profile format, with a resolution setting of 6×104 (at m/z = 400, measured at full width at half maximum peak height). In the data-dependent mode, the three most intense ions exceeding an intensity threshold of 10,000 counts were selected from each mass spectrum for MS/MS analysis using collision-induced dissociation (CID). MS/MS spectra were acquired in the positive ion mode using the linear ion trap, in profile format, with the following parameters: precursor ion mass range for data-dependent MS/MS m/z = 600 to 1000, isolation width 3 m/z units, normalized collision energy 25 %, activation time 30 ms, activation Q 0.25, and default charge state +3. Singly charged ions were excluded from data-dependent MS/MS analysis. To avoid the occurrence of redundant MS/MS measurements, real-time dynamic exclusion was enabled to preclude re-selection of previously analyzed precursor ions, using the following parameters: repeat count 1, repeat duration 30 s, exclusion list size 100, exclusion duration 180 s, and exclusion width 3 m/z units. Mass spectra and MS/MS spectra were processed using Xcalibur software (version 2.0.7, Thermo Fisher Scientific).
The presence of unadducted T3 and of IAA-iT3 (internal standard) was confirmed by manual analysis of the full MS and MS/MS spectra, with the assistance of the MS-Product algorithm for assignment of monoisotopic m/z values and the internal fragments of corresponding peptides (University of California, San Francisco, http://prospector2.ucsf.edu). Mass accuracies of T3 and IAA-iT3 standards were less than 3 ppm. Elemental compositions of the unknown adducts were inferred from the measured accurate masses and searches of the UNIMOD protein modifications database for mass spectrometry (www.unimod.org) and the Molecular Weight Calculator algorithm (v. 6.48, created by Matthew Monroe in 2010). Isotope distributions were simulated using Xcalibur software (version 2.0.7, Thermo Fisher Scientific). Retention times of each peptide from different LC-HRMS runs were adjusted to the internal standard (IAA-iT3).
Mass Spectrometry of Cys34 Oxidation Products Produced by In Vitro Modification
High resolution MS/MS spectra of synthetic T3 oxidation products, prepared in vitro, were acquired continuously by direct infusion to improve mass-spectral signal-to-noise ratios of fragment ions through signal averaging over 6 – 10 min. The sample was infused into the ESI probe of the LTQ Orbitrap XL mass spectrometer via a 250-μL syringe at a flow rate of 7 μL/min using the built-in LTQ syringe pump. The connection between the syringe needle and the ESI probe was made using PEEK tubing (0.005-in. inner diameter × 1/16-in. outer diameter, Agilent). The CID MS/MS spectra of isolated precursor ions were acquired in both the linear ion trap and the Orbitrap mass analyzer. The mass spectrometer parameters were as described above, except that the normalized collision energy for CID was optimized for T3 sequence coverage at 18%.
Molecular Modeling of Possible Intramolecular Reactions of Oxidized Cys34
The 3D structures of the T3 peptide and its oxidation products were produced in .pdb format with Chem3D Ultra 9.0. MM2 and MOPAC programs were used for minimization of intramolecular bond energies of the 3D peptide structure. Optimized 3D structures were visualized and measurements of the distances between probable interacting functional groups were calculated with PyMOL version 0.98 (http://www.pymol.sourceforge.net).
RESULTS
Detection of T3 Adducts Based on MS/MS Spectra
The fragmentation of unadducted T3 and a set of T3 adducts upon collision induced dissociation in a TQ-MS was reported previously. (10) Similar MS/MS fragmentation patterns of the T3 peptide and its synthetic adducts (Cys34 modified with 1,4-benzoquinone, 1,4-naphthoquinone, 1,2-phenanthrone, and an isomeric mixture of estrogen quinones) were obtained with the high resolution LTQ Orbitrap instrument (results not shown). Singly charged b-series ions (b3+ – 284.19, b4+ – 397.28, b5+ – 510.36, b6+ – 581.40, b11+ – 1203.42, b12+ – 1331.77 and b13+ – 1459.83 m/z) were detected in all cases, confirming that no other amino acid residue had been modified at positions prior to Cys34. The most abundant peaks in MS/MS spectra for the y-series ions were doubly charged and in the higher m/z range. In each case, y142+ y152+, y162+, y172+, y182+ and y192+ were detected and each contained chemically modified Cys34. Figure 1 shows MS/MS spectra of unadducted T3 and IAA-iT3 (internal standard) peptides. Note that b-series and y-series fragment ions of IAA-iT3 in the MS/MS spectrum are shifted by 6 Da, due to the addition of one 15N atom and five 13C atoms contributed by a labeled valine residue.
Figure 1.

HR-MS/MS spectra of T3 (A) and IAA-iT3 (internal standard) (B) measured using a LTQ Orbitrap mass spectrometer. Singly charged b-series fragment ions (b3+ - b7+ and b11+ – b14+) and doubly charged y-series fragment ions (y142+ – y192+) are shown for both peptides. Note that these y-series ions for IAA-iT3 contain the adducted Cys34 residue. Both b-series and y-series fragment ions for the IAA-iT3 peptide are shifted by 6 Da to reflect a labeled valine residue containing one 15N atom and 5 13C atoms.
To detect unknown modifications of T3 peptides, each set of MS/MS spectra was manually screened for spectra containing more than five singly charged b-series ions for the T3 peptide. The selected ion chromatogram (SIC) and corresponding mass spectra of precursor ions were then extracted from the total ion chromatogram (TIC) and used to identify the monoisotopic masses of modified T3 peptides and to calculate molecular masses of the putative adducts.
Detection of Cys34 Adducts in Human Plasma and Serum
The four purified digests of HSA from pooled plasma samples representing male and female smokers and nonsmokers, as well as the digest from fresh serum obtained from a male nonsmoker, were analyzed by LTQ Orbitrap HRMS. Table 1 shows the three putative T3 adducts with observed and theoretical monoisotopic m/z values as well as retention times, calculated added masses to the thiol form of Cys34 and the elemental composition of each adduct. These adducts were unambiguously identified (within <2 ppm) as oxidation products of Cys34, representing (+O, -H2) (possibly a sulfinamide formed by an intramolecular reaction, see discussion), the sulfinic acid (+O2) and the sulfonic acid (+O3) and were detected in all five human samples. For each of the three adducts, Figures 2 – 4 show the SIC (A), full MS (B), isotopic simulation for the expected elemental composition (C), and MS/MS spectra (D) obtained from the indicated plasma sample. Each of the three MS/MS spectra clearly shows characteristic b-series and y-series ions, the latter of which point to Cys34 as the site of adduction of the T3 peptide.
Table 1.
Results of LC-high resolution mass spectrometry of tryptic HSA digests from four pooled plasma samples representing male and female subjects (S, smoker; NS, nonsmoker) and one fresh serum sample from a male nonsmoker.
| Theoretical monoisotopic mass (+3, m/z) | Observed monoisotopic mass (+3, m/z) (mass accuracy, ppm) | Added mass, Da | Retention time*, min | Elemental Composition | Putative adduct | ||||
|---|---|---|---|---|---|---|---|---|---|
| Male NS | Male S | Female NS | Female S | Fresh serum | |||||
| 811.7594 | 811.7576 | 811.7587 | 811.7591 | 811.7563 | 811.7584 | N/A | 34.83 (0.09) | N/A | T3 (unadducted peptide) |
| (2.22) | (0.86) | (0.37) | (3.82) | (1.23) | |||||
| 816.4191 | 816.4199 | 816.4192 | 816.4201 | 816.4184 | 816.4192 | 13.9845 | 32.17 (0.05) | O | Sulfinamide (with loss of H2) |
| (0.98) | (0.12) | (1.22) | (0.86) | (0.12) | |||||
| 822.4226 | 822.4230 | 822.4239 | 822.4239 | 822.4226 | 822.4226 | 31.9963 | 32.26 (0.13) | O2 | Sulfinic acid |
| (0.49) | (1.58) | (1.58) | (0.00) | (0.00) | |||||
| 827.7543 | 827.7541 | 827.7555 | 827.7559 | 827.7545 | 827.7546 | 47.9913 | 32.48 (0.08) | O3 | Sulfonic acid |
| (0.24) | (1.45) | (1.93) | (0.24) | (0.36) | |||||
| 834.7757 | 834.7761 | 834.7771 | 834.7761 | 834.7762 | 834.7759 | 57.0234 | 33.24 (0.12) | C2H4NO | IAA-iT3 (internal standard) |
| (0.48) | (1.68) | (0.48) | (0.59) | (0.24) | |||||
Mean (SD) retention time from the four archived samples, after adjustment for the internal standard (IAA-iT3).
Figure 2.

LC-HRMS/MS data acquired for the putative sulfinamide adduct (+O, -H2) in a pooled plasma sample from male nonsmokers. (A) Selected ion chromatogram (SIC) for m/z = 816.42 (+3). (B) Isotope distribution for the SIC peak eluting at 32.18 min. (C) Simulated isotope distribution for a triply protonated ion with an empirical formula C114H171N27O31S1. (D) MS/MS spectrum of the precursor ion at m/z = 816.75 (+3) acquired in the linear ion trap.
Figure 4.

LC-HRMS/MS data acquired for the sulfonic acid adduct (+O3) in a pooled plasma sample from male smokers. (A) Selected ion chromatogram (SIC) for m/z = 827.75 (+3). (B) Isotope distribution for the SIC peak eluting at 32.38 min. (C) Simulated isotope distribution for a triply protonated ion with an empirical formula C114H173N27O33S1. (D) MS/MS spectrum of the precursor ion at m/z = 828.09 (+3) acquired in the linear ion trap.
Based upon the abundances of these peptides relative to that of the IAA-iT3 peptide used as an internal standard, the concentrations of these adducts in the five human samples are shown in Figure 5. Adducts corresponding to all three oxidation products were readily detected in all samples with relative abundances in the order: sulfinic acid (+O2) > (possible) sulfinamide (+O, -H2) > sulfonic acid (+O3). For the least abundant adduct (+O3), the mean signal/noise was 15 (SD = 3.8) for the four plasma samples.
Figure 5.

Estimated levels of three adducts representing Cys34 oxidation products in pooled samples of archived human plasma (4 or 5 subjects per pooled sample) (NS = nonsmokers, S = smokers). The sample labeled as ‘Fresh’ was obtained from the serum of a non-smoking male subject shortly before processing. Quantitation was based upon the internal standard (IAA-iT3).
Detection of Cys34 Adducts in the Negative Control
A portion of the synthetic T3 peptide was carried through the full procedure to which the plasma and serum samples had been subjected, namely, adduct enrichment (without addition of thiol affinity resin), digestion and HPLC purification. LTQ Orbitrap HPLC-MS/MS detected small amounts of adducted peptides representing the oxidation products (+O, -H2) (mass accuracy = 1.3 ppm; signal = 3.8 pmol/mg HSA) and (+O3) (mass accuracy = 1.2 ppm; signal = 4.8 pmol/mg HSA) but not the product representing (+O2).
Detection of Oxidized Cys34 Adducts Produced by In Vitro Modification
High-resolution MS of the in vitro reaction mixture containing oxidized Cys34 of the T3 peptide revealed the presence of two Cys34 oxidation products, namely a major product (87 % yield) at m/z = 827.7544 (+3) representing (+O3) and a minor product (8 % yield) at m/z = 816.4178 (+3) representing addition of oxygen and loss of two hydrogen atoms (+O, -H2). The Cys34 oxidation product representing (+O2) (m/z = 822.4226, +3) was not detected.
Precursor ions corresponding to the (+O, -H2) and (+O3) adducts were studied in detail by direct infusion of the reaction mixture into the LTQ Orbitrap mass spectrometer. Singly charged b-series ions and doubly charged y-series ions were detected and clearly assigned over the major portion of the peptide sequence, including the site of modification (Cys34). As shown in Figure 6A, fragment ions at m/z = 436.22 - representing the doubly charged y72+ fragment for the PFEDHVK segment of the T3 peptide which immediately precedes Cys34 (from the C terminus) - were observed in MS/MS spectra for the putative (+O, -H2) adduct (upper panel) and (+O3) adduct (lower panel). Importantly, the y82+ fragment ion representing CPFEDHVK (m/z = 487.72) is absent from both MS/MS spectra. Rather, the observed y82+ fragment ions show mass increases of 14 Da (Figure 6A, upper panel) and 48 Da (Figure 6B, lower panel), indicating that Cys34 was the site of modification for both the (+O, -H2) and (+O3) adducts. In addition, the doubly charged fragment ions at m/z = 470.73 and m/z = 462.22 in the MS/MS spectrum of the putative (+O, -H2) adduct (Figure 6A, upper panel) correspond to losses of SO and (SO+NH3) from the y82+ fragment ion (m/z = 494.71), respectively, neither of which was observed in the MS/MS spectrum for the putative (O3) adduct (Figure 6A, lower panel). Rather, the most prominent fragment ion in the MS/MS spectrum of the (+O3) product corresponds to (y8-H2SO3)2+ (m/z = 511.72). Release of (y8-SO)2+ and (y8-SO-NH3)2+ fragment ions from the putative (+O, -H2) adduct is consistent with formation of an intramolecular cyclic sulfinamide as described in discussion. Similar patterns of mass losses were reported by Shetty and Nubert for the sulfinamide product and sulfonic acid of a synthetic peptide containing a cysteine residue. (14, 15)
Figure 6.

HR-MS/MS spectra of synthetic oxidation products of the T3 peptide. The spectra were acquired over the range m/z = 220 to 2000 using the Orbitrap mass analyzer and recorded for 6–10 minutes. CID fragment ions of the (+O, -H2) product (m/z = 816.4176, +3, upper panels) and sulfonic acid (+O3) of Cys34 (m/z = 827.7544, +3, lower panels) are shown at the informative m/z ranges of 435 to 580 (A) and 780 to 818 (B). The arrows in (A) show the absence of a fragment ion at m/z = 487.72 (+3), corresponding to unmodified Cys34. Fragment ions containing modified Cys34 are marked by asterisks.
After obtaining specific characteristics of fragment ions from HR-MS/MS of synthetic T3 peptides (Figure 6), more detailed analysis of MS/MS spectra representing the m/z = 816.42 precursor ion (+O, -H2) was performed for the five human plasma samples. Although the complexity and low signal intensities precluded assignment of specific mass losses from precursor ions, the doubly charged y-series ions from y72+ to y192+ were clearly assigned in each sample, thereby indicating that Cys34 was the site of modification. Furthermore, the accurate masses of the (+O, -H2) adduct in the four samples differed by less than 1.6 ppm from the corresponding adduct in the synthetic mixture of oxidized T3 (m/z = 816.4178, +3), strongly suggesting that they represent the same Cys34 adduct.
DISCUSSION
Formation of the Cys34 sulfenic acid (Cys-SOH) has been proposed to be a transient intermediate in the further oxidation to the Cys34 sulfinic (Cys-SO2H) and sulfonic (Cys-SO3H) acids. (6, 7) However, none of these Cys34 oxidation products has been reported in human samples following formation in vivo. Thus, our accurate-mass measurements of T3 adducts representing both the Cys34 sulfinic acid (m/z = 822.4226, +3) and Cys34 sulfonic acid (m/z = 827.7543, +3) (see Table 1) in human plasma and serum represent the first such observations. Interestingly, all five human samples contained higher concentrations of the sulfinic acid than of the sulfonic acid (Figure 5). This result is similar to that of Aldini et al. (8) who reported the sulfinic acid as being the sole Cys34 oxidation product following incubation of human plasma with the radical inducer 2,2′-azobis(2-amidinopropane). On the other hand Finch et al. detected only the Cys34 sulfonic acid in HSA incubated with H2O2 (9), and we detected the sulfonic acid in high yield (87 %) in a solution of the T3 peptide which had been incubated with H2O2. In any case, we detected both Cys34 sulfinic and sulfonic acids in pooled plasma samples from smoking and nonsmoking subjects and in a fresh sample of serum from a nonsmoker. Although estimated adduct levels varied across the samples (Figure 5), the small sample size precludes conclusions as to possible differences.
The sulfenic acid of HSA Cys34 (+O) has been difficult to detect due to its high reactivity. (7) Extraction of the m/z value corresponding to the Cys34 sulfenic acid (817.091, +3) did not reveal a peak in the TICs of HSA digests from the five human samples in our investigation. However, manual screening of data acquired from all five samples identified MS/MS spectra of a triply charged precursor ion with m/z = 816.75 that had characteristic sequence tags for the T3 peptide (singly charged b-series ions at low m/z and prominent doubly charged y-series ions at high m/z) (Figure 2). The SIC at the same m/z range showed a peak with a mean retention time of 32.17 (SD = 0.05) min. The full MS at that retention time revealed a +3 charged peptide with a mean monoisotopic m/z value of 816.41949 (SD = 0.0008) in the five samples (Table 1). A search in the UNIMOD protein-modifications database pointed to a T3 adduct resulting from addition of one oxygen atom (+O) and subtraction of two hydrogen atoms (-H2). The theoretical monoisotopic m/z value for the triply protonated ion of this hypothetical species, with the empirical formula C140H172N27O31S1, is 816.4191, which agrees with the m/z value measured in the human samples to within 2 ppm (Table 1) and which also matched the corresponding adduct in the synthetic mixture of oxidized T3 (within 1.6 ppm).
The molecular composition of an adduct representing (+O, -H2) would be consistent with a sulfinamide product (m/z = 816.4191) incorporating an intramolecular covalent bond between the oxidized Cys34 residue and the amide group of the adjacent glutamine residue (Gln33). Such a structure is motivated by chemistry showing that cysteine sulfenic acids can react with amide, amine or imide groups of neighboring amino acids to eliminate water and produce intramolecular cyclic sulfenamides that can be subsequently oxidized to sulfinamides and sulfonamides, (14, 16) and is consistent with the (y8-SO)2+ and (y8-SO-NH3)2+ fragment ions observed in the MS/MS spectrum of the putative (+O, -H2) adduct (Figure 6A, upper panel). More careful analysis of the T3 structure ruled out the possibility of formation of a pentacyclic sulfenamide product [as suggested for other cysteine containing peptides by (15, 17)] because the next amino acid in the T3 sequence is Pro35, which is an imino acid.
Using MM2 and MOPAC programs, we investigated possible intramolecular interactions of the Cys34 sulfenic acid with nearby amino acid residues. This showed that the Cys34-SOH group was in a favorable position to react with the amide group of Gln33 to form a sulfenamide, as illustrated in Figure 7, whereas the amide group of Gln32 is sufficiently remote from the Cys34 sulfenic acid to make reaction implausible. Subsequent oxidation of the sulfur atom in the sulfenamide would yield the putative sulfinamide product as indicated by the proposed reaction scheme in Figure 8. Alternatively, Figure 8 shows that the Cys34 sulfinic acid could eliminate water in an analogous intramolecular reaction to produce the same sulfinamide product. We recognize that our assignment of a potential sulfinamide structure to the putative (+O, -H2) adduct is very preliminary and will require additional confirmation.
Figure 7.
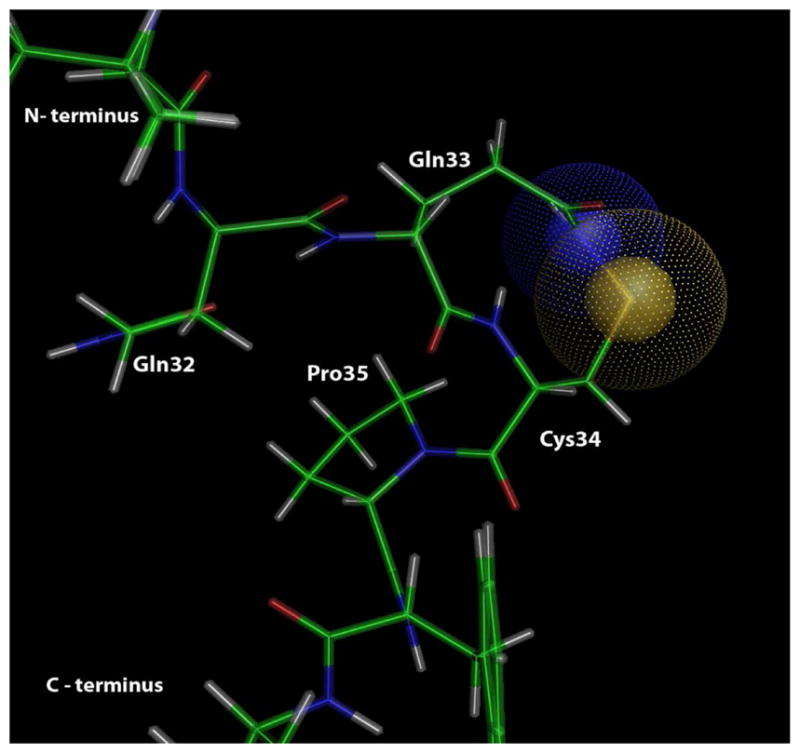
3D structure of proposed sulfenamide adduct resulting from formation of an intramolecular bond between the Cys34 sulfenic acid and the amide group of Gln33 (see Figure 8 for proposed reaction scheme).
Figure 8.

Proposed reaction pathways leading to oxidation products of Cys34 corresponding to accurate masses observed in this investigation. Confirmed masses (within 2 ppm) are indicated in bold face.
Being mindful of the possibility that oxidation of Cys34 might occur during processing of HSA samples, we carried a negative control, consisting of the synthetic T3 peptide, through the laboratory procedures in parallel with the human samples. Small quantities of the Cys34 sulfonic acid and (proposed) sulfinamide were detected in this negative control at levels equivalent to 36% and 6% of the respective average concentrations of the human samples. However, no accurate mass was detected in the negative control for the Cys34 sulfinic acid, which was the most abundant oxidation product observed in human samples. This indicates that although some oxidation of Cys34 may have occurred during laboratory processing, notably to produce the sulfonic acid, such artifacts cannot account for either the qualitative or quantitative results from the in vivo samples (Figure 5). Furthermore, the similarity of adduct profiles observed in the archived plasma samples and the fresh serum sample (Figure 5) offers assurance that the long period of storage of the archived specimens at −80 °C (13 y) did not unduly influence the measured adduct levels.
As noted in the introduction, ROS have been implicated as causative factors in many chronic diseases [reviewed by (1, 2)] and in vitro oxidation of Cys34 has itself been the object of intensive research [reviewed by (6, 18)]. Indeed, the specific roles that Cys34 plays in the antioxidant properties of HSA and associated pathological conditions have been recognized. (4, 19, 20) A critical missing piece of the puzzle has been measurements of Cys34 oxidation products in human blood. Our study offers a first step in establishing this important evidence and we encourage investigators to consider measuring these Cys34 oxidation products in samples of human serum or plasma from subjects with and without conditions that are known to produce oxidative stress. Additional work is also needed to verify the structure of the mono-oxygenated Cys34 adduct (the putative sulfinamide) that was detected in this study.
Figure 3.

LC-HRMS/MS data acquired for the sulfinic acid adduct (+O2) in a pooled plasma sample from female smokers. (A) Selected ion chromatogram (SIC) for m/z = 822.42 (+3). (B) Isotope distribution for the SIC peak eluting at 32.19 min. (C) Simulated isotope distribution for a triply protonated ion with an empirical formula C114H173N27O32S1. (D) MS/MS spectrum of the precursor ion at m/z = 822.76 (+3) acquired in the linear ion trap.
Acknowledgments
Funding Sources
This work was supported by a contract from the American Chemistry Council Long-range Research Initiative and by grant U54ES016115 from the U.S. National Institute for Environmental Health Sciences (NIEHS) through the trans-National Institutes of Health (NIH) Genes, Environment, and Health Initiative.
Abbreviations
- CID
collision-induced dissociation
- DMSO
dimethylsulfoxide
- ESI
electrospray ionization
- HSA
human serum albumin
- IAA
iodoacetamide
- IAA-iT3
carboxyamidomethylated Cys34 adduct of iT3
- iT3
isotopically-modified T3
- LC-HRMS
liquid chromatography high resolution mass spectrometry
- LC-HR-MS/MS
liquid chromatography high resolution tandem mass spectrometry
- MS/MS
tandem mass spectrometry
- ROS
reactive oxygen species
- SIC
selected ion chromatogram
- SRM
selected reaction monitoring
- T3
third largest peptide of human serum albumin digested with trypsin
- TCEP
tris(2-carboxyethyl)phosphine
- TFA
trifluoroacetic acid
- TIC
total ion chromatogram
- TQ-MS
triple-quadrupole mass spectrometry
Footnotes
Conflict of Interest Statement
The authors declare no competing financial interest.
References
- 1.Dalle-Donne I, Rossi R, Colombo R, Giustarini D, Milzani A. Biomarkers of oxidative damage in human disease. Clin Chem. 2006;52:601–623. doi: 10.1373/clinchem.2005.061408. [DOI] [PubMed] [Google Scholar]
- 2.Halliwell B, Whiteman M. Measuring reactive species and oxidative damage in vivo and in cell culture: how should you do it and what do the results mean? Br J Pharmacol. 2004;142:231–255. doi: 10.1038/sj.bjp.0705776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Aldini G, Vistoli G, Regazzoni L, Gamberoni L, Facino RM, Yamaguchi S, Uchida K, Carini M. Albumin is the main nucleophilic target of human plasma: a protective role against pro-atherogenic electrophilic reactive carbonyl species? Chem Res Toxicol. 2008;21:824–835. doi: 10.1021/tx700349r. [DOI] [PubMed] [Google Scholar]
- 4.Bourdon E, Blache D. The importance of proteins in defense against oxidation. Antioxid Redox Signal. 2001;3:293–311. doi: 10.1089/152308601300185241. [DOI] [PubMed] [Google Scholar]
- 5.Stewart AJ, Blindauer CA, Berezenko S, Sleep D, Tooth D, Sadler PJ. Role of Tyr84 in controlling the reactivity of Cys34 of human albumin. Febs J. 2005;272:353–362. doi: 10.1111/j.1742-4658.2004.04474.x. [DOI] [PubMed] [Google Scholar]
- 6.Carballal S, Alvarez B, Turell L, Botti H, Freeman BA, Radi R. Sulfenic acid in human serum albumin. Amino Acids. 2007;32:543–551. doi: 10.1007/s00726-006-0430-y. [DOI] [PubMed] [Google Scholar]
- 7.Turell L, Botti H, Carballal S, Ferrer-Sueta G, Souza JM, Duran R, Freeman BA, Radi R, Alvarez B. Reactivity of sulfenic acid in human serum albumin. Biochemistry. 2008;47:358–367. doi: 10.1021/bi701520y. [DOI] [PubMed] [Google Scholar]
- 8.Aldini G, Gamberoni L, Orioli M, Beretta G, Regazzoni L, Maffei Facino R, Carini M. Mass spectrometric characterization of covalent modification of human serum albumin by 4-hydroxy-trans-2-nonenal. J Mass Spectrom. 2006;41:1149–1161. doi: 10.1002/jms.1067. [DOI] [PubMed] [Google Scholar]
- 9.Finch JW, Crouch RK, Knapp DR, Schey KL. Mass spectrometric identification of modifications to human serum albumin treated with hydrogen peroxide. Arch Biochem Biophys. 1993;305:595–599. doi: 10.1006/abbi.1993.1466. [DOI] [PubMed] [Google Scholar]
- 10.Li H, Grigoryan H, Funk WE, Lu SS, Rose S, Williams ER, Rappaport SM. Profiling Cys34 adducts of human serum albumin by fixed-step selected reaction monitoring. Mol Cell Proteomics. 2011;10:M110. doi: 10.1074/mcp.M110.004606. 004606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Jacob C, Holme AL, Fry FH. The sulfinic acid switch in proteins. Org Biomol Chem. 2004;2:1953–1956. doi: 10.1039/b406180b. [DOI] [PubMed] [Google Scholar]
- 12.Lin YS, McKelvey W, Waidyanatha S, Rappaport SM. Variability of albumin adducts of 1,4-benzoquinone, a toxic metabolite of benzene, in human volunteers. Biomarkers. 2006;11:14–27. doi: 10.1080/13547500500382975. [DOI] [PubMed] [Google Scholar]
- 13.Funk WE, Li H, Iavarone AT, Williams ER, Riby J, Rappaport SM. Enrichment of cysteinyl adducts of human serum albumin. Anal Biochem. 2010;400:61–68. doi: 10.1016/j.ab.2010.01.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Shetty V, Spellman DS, Neubert TA. Characterization by tandem mass spectrometry of stable cysteine sulfenic acid in a cysteine switch peptide of matrix metalloproteinases. J Am Soc Mass Spectrom. 2007;18:1544–1551. doi: 10.1016/j.jasms.2007.05.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Shetty V, Neubert TA. Characterization of novel oxidation products of cysteine in an active site motif peptide of PTP1B. J Am Soc Mass Spectrom. 2009;20:1540–1548. doi: 10.1016/j.jasms.2009.04.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Fu X, Mueller DM, Heinecke JW. Generation of intramolecular and intermolecular sulfenamides, sulfinamides, and sulfonamides by hypochlorous acid: a potential pathway for oxidative cross-linking of low-density lipoprotein by myeloperoxidase. Biochemistry. 2002;41:1293–1301. doi: 10.1021/bi015777z. [DOI] [PubMed] [Google Scholar]
- 17.Salmeen A, Andersen JN, Myers MP, Meng TC, Hinks JA, Tonks NK, Barford D. Redox regulation of protein tyrosine phosphatase 1B involves a sulphenyl-amide intermediate. Nature. 2003;423:769–773. doi: 10.1038/nature01680. [DOI] [PubMed] [Google Scholar]
- 18.Turell L, Botti H, Carballal S, Radi R, Alvarez B. Sulfenic acid--a key intermediate in albumin thiol oxidation. J Chromatogr B Analyt Technol Biomed Life Sci. 2009;877:3384–3392. doi: 10.1016/j.jchromb.2009.03.035. [DOI] [PubMed] [Google Scholar]
- 19.Roche M, Rondeau P, Singh NR, Tarnus E, Bourdon E. The antioxidant properties of serum albumin. FEBS Lett. 2008;582:1783–1787. doi: 10.1016/j.febslet.2008.04.057. [DOI] [PubMed] [Google Scholar]
- 20.Oettl K, Stauber RE. Physiological and pathological changes in the redox state of human serum albumin critically influence its binding properties. Br J Pharmacol. 2007;151:580–590. doi: 10.1038/sj.bjp.0707251. [DOI] [PMC free article] [PubMed] [Google Scholar]