

Abstract
Alcohol drinking is a major etiological factor of oro-esophageal squamous cell carcinoma (OESCC). Both local and systemic effects of ethanol may promote carcinogenesis, especially among chronic alcoholics. However, molecular mechanisms of ethanol-associated OESCC are still not well understood. In this review, we summarize current understandings and propose three mechanisms of ethanol-associated OESCC: (1) Disturbance of systemic metabolism of nutrients: during ethanol metabolism in the liver, systemic metabolism of retinoids, zinc, iron and methyl groups is altered. These nutrients are known to be associated with the development of OESCC. (2) Disturbance of redox metabolism in squamous epithelial cells: when ethanol is metabolized in oro-esophageal squamous epithelial cells, reactive oxygen species are generated and produce oxidative damage. Meanwhile, ethanol may also disturb fatty-acid metabolism in these cells. (3) Disturbance of signaling pathways in squamous epithelial cells: due to its physico-chemical properties, ethanol changes cell membrane fluidity and shape, and may thus impact multiple signaling pathways. Advanced molecular techniques in genomics, epigenomics, metabolomics and microbiomics will help us elucidate how ethanol promotes OESCC.
Keywords: ethanol, oro-esophageal squamous cell carcinoma, molecular mechanisms
1. Overview
Cancers of the upper aerodigestive tract (oral cavity, pharynx, larynx, and esophagus) represent a major public health problem worldwide, making up nearly 4.4% of all malignancies in the United States alone. Approximately 73,240 new cases were diagnosed in 2014, with an estimated mortality of 27,450 [1]. Among these cancers, the incidence rates of oro-esophageal cancers have been increasing in the developed countries, especially among young males [2,3]. The most common malignancy in the oro-esophagus, oro-esophageal squamous cell carcinoma (OESCC) develops from precancerous lesions, and histopathologically follows a step-wise pattern of hyperplasia, dysplasia and squamous cell carcinoma (SCC) [4]. In the United States, the five-year survival rate has not improved significantly despite advances in radiotherapy and chemotherapy [1]. Survivors are usually left with severe functional compromise [5]. Moreover, 2% of esophageal cancer and 11% of head and neck cancer patients develop a second cancer due to field cancerization [6,7].
Many epidemiological studies have consistently shown that alcohol drinking is an etiological factor of human OESCC. OESCC has a stronger association with alcohol drinking than do cancers of other organ sites. According to a meta-analysis, strong direct trends in risk were observed for cancers of the oral cavity and pharynx (Relative Risk (RR) =6.0 for 100 g/day of ethanol), the esophagus (RR=4.2) and the larynx (RR=3.9) [8]. Risk and mortality of OESCC were associated with alcohol drinking in a dose-dependent manner [8–10]. Meanwhile, the odds ratio of oral cancer patients with dysplasia increased with alcohol consumption [11]. Genetic polymorphisms of ethanol-metabolizing genes, such as acetaldehyde dehydrogenase (ALDH) and alcohol dehydrogenases (ADH), are associated with OESCC [12–14]. Several studies have identified ALDH2*1/2 heterozygotes as a high-risk group for OESCC [15–19], and ADH genotypes not including ADH3 were associated with OESCC as well [13,20,21]. Tobacco use and alcohol drinking have synergistic effects on carcinogenesis; combined use explained more than 61% of OESCC [22,23].
After ingestion, ethanol is rapidly absorbed through the stomach and small intestine into the blood stream, and metabolized mainly in the liver before elimination. Pulmonary and urinary elimination is minimal. In the liver, ethanol is oxidized to acetaldehyde by ADH. A small amount is oxidized by cytochrome P450 2E1 (CYP2E1) and catalase. Acetaldehyde is released from the liver and metabolized into acetate by ALDH. Finally, acetate is oxidized to produce carbon dioxide, fatty acids and water. Previous studies have shown that CYP2E1 also catalyzes conversion of ethanol to acetaldehyde, and acetaldehyde to acetate [24,25].
Both ethanol and acetaldehyde may enter oro-esophageal epithelial cells through local permeation or systemic circulation. Ethanol concentration in the saliva is equal to concentration in the blood. In the saliva, ethanol is oxidized by microbes to acetaldehyde. Since further metabolism of acetaldehyde to acetate by oral bacteria is limited, acetaldehyde tends to accumulate in the saliva [26–28]. Antiseptic can significantly reduce salivary acetaldehyde concentration after alcohol drinking for this reason [27]. Tobacco changes the oral bacterial flora rapidly from Gram-negative to Gram-positive bacteria, and leads to a high concentration of acetaldehyde in saliva [29]. In agreement with these observations, alcoholics with oropharyngeal cancer had a high concentration of salivary acetaldehyde as a result of alcohol drinking, tobacco smoking and poor oral hygiene [30]. However, due to the short duration of contact and limited permeation into the epithelium, topical effects of ethanol and acetaldehyde on squamous epithelial cells are potentially weak. Presumably only superficial cells in the epithelium may be impacted by this mechanism in vivo [31].
Local and systemic effects of ethanol may influence carcinogenesis, especially among chronic alcoholics. However, the molecular mechanisms for ethanol-associated OESCC are still not well understood. Certain mechanisms of ethanol-associated cancer are supported by experimental studies of OESCC, but the majority of hypotheses are purely speculative or extrapolated from studies on cancers of other organ sites. Proposed mechanisms include: (1) Enhanced cell proliferation and altered expression of cytokeratin suggesting inhibition of squamous cell differentiation [32]; (2) Enhanced penetration of carcinogens into the squamous epithelium [31]; (3) Impaired antioxidant defense and enhanced production of reactive oxygen species (ROS) in the squamous epithelium [26]; (4) Interference with DNA repair machinery and DNA synthesis [33]; (5) Disturbed systemic metabolism of nutrients [26]; (6) Impaired immune function [34]; (7) Induced chronic inflammation and enhanced angiogenesis [35].
On one hand, further experimental studies are needed to examine these mechanisms in ethanol-associated OESCC. On the other hand, these mechanisms have not been systematized to provide an overview of how ethanol promotes OESCC. In this review, we summarize current data and propose three major mechanisms of ethanol-associated OESCC: (1) Disturbance of systemic metabolism of nutrients; (2) Disturbance of metabolism in squamous epithelial cells; (3) Disturbance of signaling pathways in squamous epithelial cells.
2. Disturbance of systemic metabolism of nutrients
2.1. Inhibition of retinol metabolism
When metabolized, ethanol impairs retinoid metabolism by inhibiting retinol metabolism to retinoic acid via competing with retinol for ADH and ALDH active sites, and by accelerating catabolism of vitamin A through induction of CYP2E1 [36]. A recent study showed that acetaldehyde inhibited formation of retinoic acid from retinal in rat esophagus ex vivo [37]. Lecithin:retinol acyltransferase which regulates retinol metabolism by esterifying retinol, is down-regulated in human head and neck SCC cells. In a study using knockout mice, partial retinol deficiency during carcinogen treatment promoted cell proliferation and carcinogenesis in tongue epithelium [38]. Retinoic acid is known to exert profound effects on cellular growth, differentiation, and cancer development in the oro-esophageal epithelium through its interaction with receptors [36,39]. These studies suggest that ethanol promotes OESCC through inhibition of retinoic acid signaling. In fact, a retinoid X receptor agonist and a retinoic acid receptor γ selective agonist inhibited 4-nitroquinoline 1-oxide (4NQO)-induced oral carcinogenesis in mice [40].
2.2. Zinc deficiency
Alcohol abuse has long been associated with zinc deficiency [41]. Ethanol treatment down-regulates the expression of zinc transporters 1 and 4, as well as the zinc storage protein metallothionein 1 in alveolar macrophages, disrupting zinc bioavailability [42]. ADH is a zinc metalloenzyme, and removal of zinc from ADH leads to a complete loss of its catalytic activity [43]. While zinc supplementation prevents alcoholic liver injury through attenuation of oxidative stress [43], zinc depletion is known to enhance oro-esophageal carcinogenesis in rats and mice [44,45]. Mechanistically, zinc deficiency causes extensive alterations in gene expression in mouse and rat esophageal epithelium [46–48]. In particular, a group of cancer-related pro-inflammatory genes were up-regulated (CXC and CC chemokines, chemokine receptors, cytokines and cyclooxygenase 2, S100A8/A9, and nuclear factor κB (NFκB)), suggesting that multiple inflammatory pathways participate in zinc deficiency-related OESCC. Consistent with this observation, zinc supplementation caused a shift to a less proliferative cancer phenotype by normalizing the inflammatory gene signature, inhibiting cell proliferation, and stimulating apoptosis [49,50].
2.3. Iron overload
Alcohol drinking has been shown to cause iron overload in the liver [51]. Ethanol increases total iron content via overexpression of genes involved in iron transport (divalent metal transporter 1, transferrin receptor 1, ferroportin, ceruloplasmin) and iron storage (L-ferritin) [52]. We have shown that iron accumulation in the esophagus promoted inflammation-associated carcinogenesis [53]. Mechanistically, iron overload may initiate and promote carcinogenesis through oxidative damage [53] and modification of the immune reaction [54]. It is expected that oxidative stress affects carcinogenesis through redox signaling pathways inside cells [55,56].
2.4. Increased requirements for methyl groups
Chronic alcoholism increases the requirements for methyl groups and causes dietary methyl group deficiency [26]. Deficiency of S-adenosylmethionine, folate and betaine, primarily due to low intake and destruction by acetaldehyde, is common in alcoholics. Inhibition of methyl group transfer regulates expression of genes involved in carcinogenesis [57,58]. DNA hypomethylation of oncogenes (e.g., c-Ha-ras, c-Ki-ras and c-fos) is associated with an increased incidence of liver cancer in rats [59,60]. These data suggest that ethanol may contribute to OESCC through aberrant gene methylation [61,62]. In addition, aberrant gene methylation may impact signaling pathways through critical pathway genes, such as Notch4 of the Notch signaling pathway [63], PTEN of the phospoinositide 3-kinase (PI3K)/Akt pathway [64], and Wnt inhibitory factor 1 (WIF1) of the Wnt signaling pathway [65]. Recent studies have also showed that aberrant methylation of histones and non-histone proteins also modulates multiple signaling pathways [66].
3. Disturbance of redox metabolism in squamous epithelial cells
After ethanol and acetaldehyde get into epithelial cells through systemic circulation, they undergo intracellular metabolism [67]. It should be noted that ethanol metabolism in the oro-esophageal epithelial cells is different from that in the liver due to different enzyme profiles. Oral epithelium expresses χ and σ type ADH with high Km values (ADH3 and ADH4) [68], and a negligible amount of ALDH. CYP2E1 is not normally expressed in oral epithelium, but is inducible by chronic ethanol treatment [69–71]. As a result, oro-esophageal epithelial cells tend to accumulate acetaldehyde. This may partially explain why the oro-esophagus is more susceptible to ethanol-associated carcinogenesis when compared to other organ sites.
When ethanol is metabolized, ROS are produced in cells. ROS produced by the mitochondrial electron transport chain, CYP2E1, and cytosolic enzymes [72], result in oxidative damage of DNA, proteins and lipids. Chronic alcohol drinking promotes esophageal tumorigenesis which is associated with ROS production; administration of an antioxidant (α-tocopherol) attenuates this effect [73].
It has been well established that ethanol and its toxic metabolite, acetaldehyde, form DNA adducts and thus cause DNA mutations [74]. Acetaldehyde reacts directly with the exocyclic amino group of deoxyguanosine to form stable DNA adducts [75]. As the most abundant DNA adduct, N2-ethylidene-2′-deoxyguanosine (N2-EtidG), impairs the DNA repair system and apoptosis [26]. The level of N2-EtidG is increased in oro-esophageal epithelial cells after alcohol drinking in mice [76] and humans [77]. Moreover, the N2-EtidG level is dependent on ALDH2 and ADH genotypes. In animals, lack of ALDH2 leads to an increased level of N2-EtidG in the mouse upper aerodigestive tract after ethanol treatment [76]. In humans, the blood level of N2-EtidG was significantly higher in patients with a combination of the ADH1B*2 and ALDH2*2 alleles [78].
Excessive ethanol may also lead to generation of protein adducts with the aldehydic final products of lipid peroxidation, malondialdehde (MDA) and 4-hydroxynonenal (HNE) [79,80]. As the most abundant adduct species, HNE levels in mouse liver are dependent on both the dose and the duration of ethanol feeding [81]. Similarly, the amount of MDA is found to be significantly increased in the heart of rats treated with ethanol [82]. In oral leukoplakia and cancer, more positive staining for MDA and HNE adducts was observed in the dysplastic or malignant epithelial cells than in normal cells, and this staining was significantly correlated with acetaldehyde exposure [80]. HNE reacts with DNA bases such as deoxyadenosine and deoxycytidine to form highly mutagenic DNA adducts [83]. Furthermore, ethanol can induce hyperacetylation of proteins such as histone H3, p53 and SREBP-1c, perturbing hepatic function [84].
Cell membranes are sensitive to oxidative damage. The methylene groups between two double bonds of polyunsaturated fatty acid in cell membranes make these lipids highly sensitive to oxidation [79]. ROS-induced lipid peroxidation of cell membranes has been implicated in malignant transformation [79]. For the sake of self-protection or adaptation, cells respond to ethanol and ROS through at least two mechanisms. In response to oxidative stress, nuclear factor erythroid 2-like factor (Nrf2) released from Kelch-like ECH associating protein 1 (Keap1) translocates into the nucleus and induces transcription of target genes [85,86]. Nrf2 signaling plays an essential role in the protection of cells against oxidative damage and carcinogenesis [87,88]. Ethanol suppresses Nrf2 expression in alveolar epithelial cells and an Nrf2 activator (sulforaphane) blocks the effects of ethanol [89]. Activation of Nrf2 prevents ethanol-induced oxidative stress, lipid accumulation and accelerated acetaldehyde metabolism [85,90].
Cells may also respond to ethanol through modification of intracellular fatty acid metabolism. In the liver, heavy doses ethanol increase intracellular concentration of free fatty acids and block fatty acid oxidation through inhibition of peroxisome proliferator-activated receptor α [91–94]. It is interesting that in yeast cells, endosperm effective 1 and elongation enzyme 1 are known to be associated with ethanol resistance [95,96]. Functional mammalian homologues of these genes, EH-domain containing genes (Ehd1, Ehd2, Ehd3, Ehd4) and very long chain fatty acid elongase (Elovl3, Elovl6) are associated with lipid metabolism and membrane functions. Knockdown of Ehd1 reduced cellular cholesterol and triglyceride, and disrupted free cholesterol distribution in the cell [97,98]. Elovl6 deficient mice became obese and developed hepatosteatosis when fed a high-fat diet [99,100]. Inhibition of Elovl3 reduced intracellular triglycerides significantly [101]. Both Elovl6 and Elovl3 were activated by peroxisome proliferator-activated receptor γ during adipogenesis [102].
4. Disturbance of signaling pathways in squamous epithelial cells
As a short-chain alcohol, ethanol diffuses in squamous epithelial cells. In this process, ethanol molecules are expected to accumulate near the interface region between lipids and the surrounding water. Ethanol is able to form hydrogen bonds with lipids in the bilayer, significantly change membrane fluidity and deformability, and leave the cell membrane unstable. Interestingly, ethanol has a biphasic effect on bilayer deformability as it first increases and then decreases deformability [103]. Ethanol reduces the membrane’s surface tension and lateral pressure, which affects the conformations of membrane proteins [104], and disrupts protein-lipid interactions [105]. On the other hand, ethanol can activate a G-protein-gated inwardly-rectifying potassium channel via a defined alcohol pocket and changes in affinity for membrane phospholipid signaling [106]. These small changes in cell membranes can strongly affect the function of intrinsic membrane proteins. Therefore, it is conceivable that ethanol may act on multiple signaling pathways as shown below.
4.1. Toll-like receptor 4 (TLR4) signaling
Treatment with lipopolysaccharide, a TLR4 ligand, results in recruitment of CD14 and TLR4 into the lipid rafts, and thus triggers downstream TLR signaling events in cells. When cells are co-treated with ethanol, reorganization of the actin cytoskeleton is disturbed, impacting TLR4 and CD14 clustering and co-localization, and TLR4 signaling is suppressed [107]. However, TLR4 was overexpressed in OESCC in vivo and in vitro [108–110]. Lipopolysaccharide activation of TLR4 on cancer cells enhanced cell proliferation and inflammation through the PI3K/Akt and NFkB pathways [111]. In an animal study using mice lacking the receptor for advanced glycation end product, TLR4 was found to mediate S100A8/A9-induced pro-inflammatory reaction [112]. TLR4 may also contribute to immune escape and chemoresistance [108]. TLR4 in host cells was involved in the regulation of tumor-related cachexia and tumor control [113]. Interestingly, a functional study showed defects in TLR4-mediated signaling in peripheral blood lymphocytes from oral cancer patients [114]. These data suggest that TLR4 may play different roles in epithelial cells and immune cells. It is still unknown how ethanol may promote OESCC through TLR4 signaling.
4.2. Notch signaling
Notch signaling is mediated through ligands binding to Notch receptors. The receptors are then cleaved to allow Notch intracellular domain (NICD) release from the membrane and entry into the nucleus to form a transcriptional complex with recombining binding protein for immunoglobulin kappa j (RBPJ). NICD displaces the repressive cofactors bound to RBPJ and recruits a transcriptional activator complex, which initiates transcription of Notch downstream target genes [115]. Notch is known to regulate squamous differentiation in the skin [116] and esophagus [117,118], particularly during commitment of keratinocytes to terminal differentiation via a Hes1-dependent mechanism [119,120]. In the esophagus, loss of Notch signaling perturbs esophageal squamous differentiation [117]. In the oral cavity, Notch1 is highly activated in oral epithelium differentiation, and disrupted Notch1 interferes with normal palate development [121]. Exome sequencing has shown in multiple studies that loss-of-function Notch mutations are frequently seen in OESCC [122–127].
The effects of ethanol on Notch signaling vary in different organs. Notch signaling is inhibited by ethanol in the pancreas and smooth muscle cells [128,129], but activated in endothelial cells and fetal cerebral cortex [130,131]. In the oro-esophagus, we found that alcohol drinking promoted epithelial cell proliferation and suppressed squamous differentiation in mouse forestomach, particularly after long-term treatment. Meanwhile, Notch signaling was down-regulated by ethanol treatment (unpublished data). These data suggest Notch inhibition as a potential mechanism of suppression of squamous differentiation due to alcohol drinking. Using genetically modified mice and chemical modulators of Notch signaling, we are testing Notch inhibition as a novel molecular mechanism of ethanol-associated OESCC.
4.3. Sonic hedgehog (Shh) signaling
Shh signaling is one of the most well-known developmental signaling pathways. In the ligand-producing cells, Shh-N is packaged into lipid raft domains and then shipped extracellularly to the receiving cells. Shh signaling is activated when Shh-N binds to Patched which releases its inhibition on Smoothened, leading to activation of Gli transcription factors to regulate gene expression [132,133]. In fetal alcohol syndrome models, Shh signaling is suppressed by ethanol. Shh deficiency makes animals more susceptible to fetal alcohol phenotypes. Supplementation of Shh mRNA [134–136] or cholesterol rescues these phenotypes [137]. In vitro studies find that ethanol perturbs proper association of active Shh ligand with lipid rafts destined for the membrane [132,133]. Furthermore, ethanol exposure leads to a decrease in the amount of membrane-associated cholesterol, which is vital for proper Shh signaling [137]. Contrary to this observation, ethanol activates Shh signaling, and thus promotes carcinogenesis in the liver [138,139]. In OESCC, Shh signaling is aberrantly activated [140–144], suggesting that ethanol may activate Shh signaling in the oro-esophagus.
4.4. Wnt signaling
Wnt signaling plays a crucial role in the regulation of cell proliferation, survival and migration [145]. Binding of Wnt to its receptor (frizzled) and co-receptors (low-density lipoprotein receptor-related protein 5 or 6) transduces a signal across the plasma membrane and activates Wnt signaling [145]. β-catenin plays an important role in the epithelial-mesenchymal transition that characterizes tissue regeneration and cancer proliferation [146]. It is interesting that ethanol suppresses Wnt signaling in human neural stem cells [147], but activates it in the fetal cerebral cortex of humans and mice [130]. In diethylnitrosamine-induced hepatocarcinogenesis, ethanol promotes Wnt signaling in mouse liver [148]. In OESCC, higher expression of nuclear β-catenin is significantly associated with poor histological differentiation and clinical outcome [149]. Total β-catenin level in the tongue increases after treatment with a carcinogen and ethanol [150]. In esophageal SCC, overexpression of Wnt2 is found in tumor-associated fibroblasts and is correlated with poor survival [145]. These data suggest that ethanol may activate Wnt signaling in the oro-esophagus.
4.5. Transforming growth factor β (TGFβ) signaling
TGFβ signaling is a key regulator of epithelial cell proliferation, immune function and angiogenesis [151]. In the canonical pathway, TGFβ ligands bind to the transmembrane receptor (type 2 TGFβ receptor) that recruits type 1 TGFβ receptor, resulting in Smad-mediated signaling. TGFβ also signals through non-Smad pathways [152,153]. TGFβ signaling is inhibited by ethanol in rat cerebral cortex [154]. However, TGFβ is activated by ethanol in mouse liver and rat alveolar macrophage [155,156]. Meanwhile, exposure to ethanol increases fibroblast contraction, which is reduced by TGFβ inhibition [157]. In head and neck SCC, loss of type 2 TGFβ receptor or Smad4 not only abrogates TGFβ-mediated tumor suppression, but also causes a compensatory increase in TGFβ ligand expression that promotes inflammation and angiogenesis [151].
4.6. NFκB signaling
NFκB signaling is involved in many cellular processes, including inflammation and tumorigenesis. Once activated, the regulatory subunit phosphorylates specific Ser residues, leading to protein ubiquitination, subsequent proteasome processing and NFκB releasing. Transcription factors accumulate in the nucleus to activate the target genes [86]. Ethanol activates NFκB signaling and up-regulates expression of inflammatory mediators, and thus induces hepatotoxicity [158–160]. In the oro-esophagus, NFκB alleles are associated with oral carcinogenesis [161]. It is still not known whether ethanol may regulate NFκB signaling in oro-esophageal epithelial cells.
4.7. Other signaling
Mitogen-activated protein kinase (MAPK) signaling plays a key role in cell growth, proliferation, migration and apoptosis. Modulation of MAPK signaling by ethanol is distinctive, depending on the context (cell type, acute or chronic exposure, normal or transformed cell phenotype) [162]. Chronic ethanol treatment inhibits p42/44 MAPK and growth factor-stimulated p42/44 MAPK activation. In contrast, chronic ethanol potentiates endotoxin-stimulated p42/44 MAPK and p38 MAPK signaling in Kupffer cells. In mouse tongue epithelial cells, ethanol alone increased the expression of p38 MAPK, but not Erk1/2 MAPK [150].
PI3K/Akt signaling is a critical regulatory axis for cell growth, motility and survival [163]. The effects of ethanol on PI3K/Akt signaling vary depending on the cell type and ethanol concentration. PI3K/Akt activation was involved in acute ethanol-induced fatty liver in mice [164]. Similarly, activation of Akt was observed in the nucleus accumbens of rodents after acute systemic administration of alcohol [165]. On the other hand, PI3K/Akt signaling was inhibited by ethanol in primary cortical neurons [166]. Interestingly, low-concentration alcohol reduced, but high-concentration enhanced, Akt activity in rat cardiac tissue and cardiocytes [167].
G-protein coupled receptors (GPCRs) are involved in many biological functions, including inflammation, carcinogenesis and chemotaxis, and may cross-talk with other signaling pathways in mediating these cellular processes [168]. GPR 55 was overexpressed in monocyte-derived dendritic cells from alcohol users or those treated with ethanol [169]. The ability of ethanol to increase gamma-aminobutyric acid release can be regulated by different GPCRs, such as cannabinoid-1 receptor, corticotropin-releasing factor 1 receptor and 5-hydroxytryptamine 2C receptor [170].
5. Overall effects of ethanol on the oro-esophageal epithelium
Ethanol has been shown to cause several changes in cellular behaviors of oro-esophageal squamous epithelial cells. Treatment of human OESCC cells with ethanol enhances cell proliferation and alters expression of cytokeratins suggesting inhibition of squamous cell differentiation [32]. In vivo, chronic drinking of acetaldehyde causes hyperproliferation and suppresses differentiation of oro-esophageal epithelial cells [171]. Meanwhile, chronic exposure to ethanol causes epithelial atrophy and reduces basal cell size of rat esophageal mucosa [172]. These changes in cell proliferation and differentiation may be explained by a recent study showing that ethanol promotes symmetric division of stem cells in oral epithelium [150].
Chronic inflammation induced by alcohol drinking promotes carcinogenesis. In the intestine, ethanol-induced mast cell-mediated inflammation promotes tumorigenesis in a mouse model [173]. Ethanol supports macrophage recruitment and reinforces invasion and migration of Lewis lung carcinoma in mice [174]. Similarly, we observed that ethanol promoted oral carcinogenesis through activation of the 5-lipoxygenase pathway, a pro-inflammatory pathway of arachidonic acid metabolism. Angiogenesis was also promoted in 4NQO-induced mouse tongue SCC by ethanol [35].
6. Summary
Ethanol consumption clearly contributes to OESCC as both a co-carcinogen and a tumor promoter potentially via multiple mechanisms (Figure 1). Further studies are warranted to understand how these signaling pathways contribute to ethanol-associated OESCC. One challenge comes from known interactions between pathways. For example, Wnt signaling is associated with Notch signaling. Knockdown of Notch1 enhances the activation of Wnt target genes, whereas chemical inhibition of γ-secretase or expression of a Notch1 mutant inhibits Wnt signaling [175,176]. Notch signaling also interacts with Nrf2 signaling. In mouse embryonic fibroblasts, Nrf2 regulates Notch1 expression through a functional antioxidant response element in the promoter. Nrf2 disruption impedes liver regeneration which can be rescued by reestablishment of Notch1 signaling [177].
Figure 1.
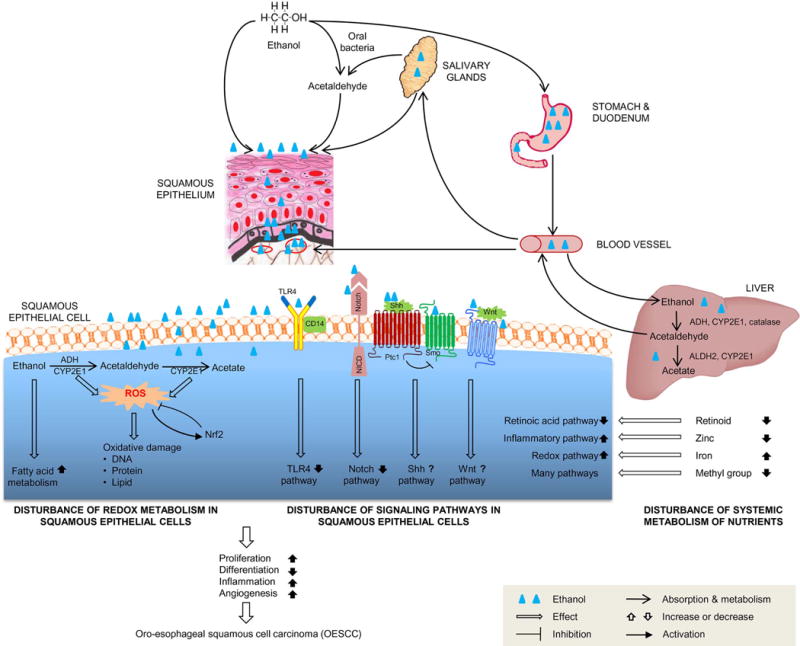
Molecular mechanisms of ethanol-associated OESCC. After ingestion, ethanol is quickly absorbed through the stomach and duodenum into blood stream, and metabolized mainly in the liver. Ethanol is oxidized into acetaldehyde by ADH, CYP2E1 and catalase, and then into acetate by ALDH. Systemic metabolism of ethanol causes reduction of retinoid, zinc and methyl group and accumulation of iron. Through the blood stream, ethanol is circulated to the oro-esophageal epithelium and salivary glands. In the saliva, ethanol is oxidized by microbes to acetaldehyde. When ethanol and acetaldehyde contact oro-esophageal epithelial cells, ethanol perturbs the lipid bilayer of cell membrane and interferes the function of intrinsic membrane proteins, such as TLR4, Notch, Shh and Wnt pathway receptors, and thus activates or inhibits downstream signaling. Inside the epithelial cells, ethanol metabolism causes oxidative damages to DNA, proteins and lipids, and modulates fatty acid metabolism as well. These systemic and local effects of ethanol and acetaldehyde stimulate cell proliferation, inflammation and angiogenesis, and suppress squamous cell differentiation, and therefore promote OESCC.
Another challenge is lack of proper in vivo models. Studies on animal models have failed to demonstrate a strong promoting effect of ethanol on OESCC. In one study, life-long exposure to 10% ethanol in drink significantly increased the incidence of oral cancer in Sprague-Dawley rats [178]. Several previous studies have found that co-administration of ethanol enhanced chemically induced OESCC [73,179–181]. In the oral cavity, studies have suggested, but failed to confirm from a statistical point of view, the cancer-promoting activity of ethanol feeding or painting on 7,12-dimethylbenz[a]anthracene (DMBA)-induced hamster cheek pouch carcinogenesis [179,182,183]. In the esophagus, ethanol was reported to have only weak or no promoting activity on carcinogenesis [184–188]. Most of these studies could not reach solid conclusions with enough statistical power due to small sample sizes. Although concentrated ethanol is believed to be more effective in promoting cancer than diluted ethanol, one major limitation in animal studies is poor tolerance of lab animals to concentrated ethanol. ~20% (w/v, or 25% v/v) or ~32% (w/v, or 40% v/v) is the upper limit of ethanol concentration in drinking water for mice or rats, respectively [189]. In addition, ethanol passes the oral cavity and esophagus quickly, thus its effects on the oro-esophageal squamous epithelial cells in vivo tend to be weak and transient. Several approaches have been used to enhance voluntary alcohol drinking in animals. Sweeteners are commonly used in ethanol self-administration paradigms, either to initiate ethanol drinking or to increase ethanol consumption [190]. A variety of studies performed in at least seven strains of rats and most inbred strains of mice indicated a close association between consumption of sweet solution (i.e. 0.2% saccharin, 5% sucrose) and ethanol intake [191–195]. Supersaccharin (0.125% saccharin and 3% sucrose) is a commonly used sweetener for rats [196]. Gradual increase of ethanol concentration in drink and using a prandial model (postmeal drinking) can also help rodents acclimate to relatively concentrated ethanol [197]. We use forestomach as the surrogate target organ site in a carcinogen-induced OESCC model based on the histological similarities between the esophagus and the forestomach [49,198].
Recently, advanced molecular techniques have been widely used in cancer research. Genomics techniques such as NextGen sequencing, CGH array and SNP array are used for detection of DNA alterations, RNA-Seq and microarray for detection of mRNA differential expression, and methylation array for detection of alterations in gene methylation [199,200]. NextGen sequencing data of ESCC show that non-alcohol users show a relatively distinct mutation pattern and have better prognosis than alcohol users [201]. Metabolomics has been used to identify biomarkers of ethanol-induced liver disease [202,203]. Microbiomics may help us understand how ethanol modifies oral bacteria and whether certain bacteria may be more carcinogenic than others [204,205]. We expect that many new discoveries will be made in our understanding of the molecular mechanisms of ethanol-associated OESCC.
Highlights.
Alcohol drinking is a major etiological factor of oro-esophageal squamous cell carcinoma.
Mechanisms of ethanol-associated oro-esophageal squamous cell carcinoma are reviewed.
Disturbance of systemic metabolism of nutrients, of redox metabolism and of signaling pathways in squamous epithelial cells are proposed mechanisms.
Acknowledgments
This work is supported by research grants from the NIH/NIAAA (U54 AA019765) and the National Natural Science Foundation of China (81372897).
Abbreviations
- 4NQO
4-nitroquinoline 1-oxide
- ADH
alcohol dehydrogenase
- ALDH
acetaldehyde dehydrogenase
- CYP2E1
cytochrome P450 2E1
- Edh
EH-domain containing gene
- ELOVL
very long chain fatty acid elongase
- GPCRs
G-Protein coupled receptors
- HNE
4-hydroxynonenal
- MAPK
mitogen-activated protein kinase
- MDA
malondialdehde
- N2-EtidG
N2-ethylidene-2′-deoxyguanosine
- NFκB
nuclear factor κB
- NICD
Notch intracellular domain
- Nrf2
nuclear factor erythroid 2-like factor
- OESCC
oro-esophageal squamous cell carcinoma
- PI3K
phosphoinositide 3-kinase
- RBPJ
recombining binding protein for immunoglobulin kappa j
- ROS
reactive oxygen species
- RR
relative risk
- SCC
squamous cell carcinoma
- Shh
sonic hedgehog
- TGFβ
transforming growth factor beta
- TLR4
Toll-like receptor 4
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Conflict of interest statement
The authors declare there is no conflict of interest regarding the publication of this paper.
References
- 1.Siegel R, Ma J, Zou Z, Jemal A. Cancer statistics, 2014. CA Cancer J Clin. 2014;64:9–29. doi: 10.3322/caac.21208. [DOI] [PubMed] [Google Scholar]
- 2.Mackenzie J, Ah-See K, Thakker N, Sloan P, Maran AG, Birch J, Macfarlane GJ. Increasing incidence of oral cancer amongst young persons: what is the aetiology? Oral Oncol. 2000;36:387–389. doi: 10.1016/s1368-8375(00)00009-9. [DOI] [PubMed] [Google Scholar]
- 3.Davis S, Severson RK. Increasing incidence of cancer of the tongue in the United States among young adults. Lancet. 1987;2:910–911. doi: 10.1016/s0140-6736(87)91393-6. [DOI] [PubMed] [Google Scholar]
- 4.Stelow EB, Mills SE. Squamous cell carcinoma variants of the upper aerodigestive tract. Am J Clin Pathol. 2005;124(Suppl):S96–109. doi: 10.1309/CR5JXUY3J2YGTC1D. [DOI] [PubMed] [Google Scholar]
- 5.Franceschi S, Levi F, Vecchia C. Decline in 5-year survival rates for cancer of head and neck. Lancet. 1992;340:47. doi: 10.1016/0140-6736(92)92456-p. [DOI] [PubMed] [Google Scholar]
- 6.Chuang SC, Scelo G, Tonita JM, Tamaro S, Jonasson JG, Kliewer EV, Hemminki K, Weiderpass E, Pukkala E, Tracey E, Friis S, Pompe-Kirn V, Brewster DH, Martos C, Chia KS, Boffetta P, Brennan P, Hashibe M. Risk of second primary cancer among patients with head and neck cancers: A pooled analysis of 13 cancer registries. Int J Cancer. 2008;123:2390–2396. doi: 10.1002/ijc.23798. [DOI] [PubMed] [Google Scholar]
- 7.Chuang SC, Hashibe M, Scelo G, Brewster DH, Pukkala E, Friis S, Tracey E, Weiderpass E, Hemminki K, Tamaro S, Chia KS, Pompe-Kirn V, Kliewer EV, Tonita JM, Martos C, Jonasson JG, Dresler CM, Boffetta P, Brennan P. Risk of second primary cancer among esophageal cancer patients: a pooled analysis of 13 cancer registries. Cancer Epidemiol Biomarkers Prev. 2008;17:1543–1549. doi: 10.1158/1055-9965.EPI-07-2876. [DOI] [PubMed] [Google Scholar]
- 8.Bagnardi V, Blangiardo M, La Vecchia C, Corrao G. A meta-analysis of alcohol drinking and cancer risk. Br J Cancer. 2001;85:1700–1705. doi: 10.1054/bjoc.2001.2140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Li Y, Mao Y, Zhang Y, Cai S, Chen G, Ding Y, Guo J, Chen K, Jin M. Alcohol drinking and upper aerodigestive tract cancer mortality: A systematic review and meta-analysis. Oral Oncol. 2014 doi: 10.1016/j.oraloncology.2013.12.015. [DOI] [PubMed] [Google Scholar]
- 10.Marron M, Boffetta P, Moller H, Ahrens W, Pohlabeln H, Benhamou S, Bouchardy C, Lagiou P, Lagiou A, Slamova A, Schejbalova M, Merletti F, Richiardi L, Kjaerheim K, Agudo A, Castellsague X, Macfarlane TV, Macfarlane GJ, Talamini R, Barzan L, Canova C, Simonato L, Biggs AM, Thomson P, Conway DI, McKinney PA, Znaor A, Healy CM, McCartan BE, Brennan P, Hashibe M. Risk of upper aerodigestive tract cancer and type of alcoholic beverage: a European multicenter case-control study. Eur J Epidemiol. 2012;27:499–517. doi: 10.1007/s10654-012-9699-1. [DOI] [PubMed] [Google Scholar]
- 11.Morse DE, Psoter WJ, Cleveland D, Cohen D, Mohit-Tabatabai M, Kosis DL, Eisenberg E. Smoking and drinking in relation to oral cancer and oral epithelial dysplasia. Cancer Causes Control. 2007;18:919–929. doi: 10.1007/s10552-007-9026-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Tanaka F, Yamamoto K, Suzuki S, Inoue H, Tsurumaru M, Kajiyama Y, Kato H, Igaki H, Furuta K, Fujita H, Tanaka T, Tanaka Y, Kawashima Y, Natsugoe S, Setoyama T, Tokudome S, Mimori K, Haraguchi N, Ishii H, Mori M. Strong interaction between the effects of alcohol consumption and smoking on oesophageal squamous cell carcinoma among individuals with ADH1B and/or ALDH2 risk alleles. Gut. 2010;59:1457–1464. doi: 10.1136/gut.2009.205724. [DOI] [PubMed] [Google Scholar]
- 13.Wu C, Kraft P, Zhai K, Chang J, Wang Z, Li Y, Hu Z, He Z, Jia W, Abnet CC, Liang L, Hu N, Miao X, Zhou Y, Liu Z, Zhan Q, Liu Y, Qiao Y, Zhou Y, Jin G, Guo C, Lu C, Yang H, Fu J, Yu D, Freedman ND, Ding T, Tan W, Goldstein AM, Wu T, Shen H, Ke Y, Zeng Y, Chanock SJ, Taylor PR, Lin D. Genome-wide association analyses of esophageal squamous cell carcinoma in Chinese identify multiple susceptibility loci and gene-environment interactions. Nat Genet. 2012;44:1090–1097. doi: 10.1038/ng.2411. [DOI] [PubMed] [Google Scholar]
- 14.Oze I, Matsuo K, Suzuki T, Kawase T, Watanabe M, Hiraki A, Ito H, Hosono S, Ozawa T, Hatooka S, Yatabe Y, Hasegawa Y, Shinoda M, Kiura K, Tajima K, Tanimoto M, Tanaka H. Impact of multiple alcohol dehydrogenase gene polymorphisms on risk of upper aerodigestive tract cancers in a Japanese population. Cancer Epidemiol Biomarkers Prev. 2009;18:3097–3102. doi: 10.1158/1055-9965.EPI-09-0499. [DOI] [PubMed] [Google Scholar]
- 15.Yokoyama A, Watanabe H, Fukuda H, Haneda T, Kato H, Yokoyama T, Muramatsu T, Igaki H, Tachimori Y. Multiple cancers associated with esophageal and oropharyngolaryngeal squamous cell carcinoma and the aldehyde dehydrogenase-2 genotype in male Japanese drinkers. Cancer Epidemiol Biomarkers Prev. 2002;11:895–900. [PubMed] [Google Scholar]
- 16.Yokoyama A, Omori T, Yokoyama T, Sato Y, Mizukami T, Matsushita S, Higuchi S, Maruyama K, Ishii H, Hibi T. Risk of squamous cell carcinoma of the upper aerodigestive tract in cancer-free alcoholic Japanese men: an endoscopic follow-up study. Cancer Epidemiol Biomarkers Prev. 2006;15:2209–2215. doi: 10.1158/1055-9965.EPI-06-0435. [DOI] [PubMed] [Google Scholar]
- 17.Yokoyama A, Omori T. Genetic polymorphisms of alcohol and aldehyde dehydrogenases and risk for esophageal and head and neck cancers. Jpn J Clin Oncol. 2003;33:111–121. doi: 10.1093/jjco/hyg026. [DOI] [PubMed] [Google Scholar]
- 18.Matsuo K, Hamajima N, Shinoda M, Hatooka S, Inoue M, Takezaki T, Tajima K. Gene-environment interaction between an aldehyde dehydrogenase-2 (ALDH2) polymorphism and alcohol consumption for the risk of esophageal cancer. Carcinogenesis. 2001;22:913–916. doi: 10.1093/carcin/22.6.913. [DOI] [PubMed] [Google Scholar]
- 19.Hashibe M, Boffetta P, Zaridze D, Shangina O, Szeszenia-Dabrowska N, Mates D, Janout V, Fabianova E, Bencko V, Moullan N, Chabrier A, Hung R, Hall J, Canzian F, Brennan P. Evidence for an important role of alcohol- and aldehyde-metabolizing genes in cancers of the upper aerodigestive tract. Cancer Epidemiol Biomarkers Prev. 2006;15:696–703. doi: 10.1158/1055-9965.EPI-05-0710. [DOI] [PubMed] [Google Scholar]
- 20.Bouchardy C, Hirvonen A, Coutelle C, Ward PJ, Dayer P, Benhamou S. Role of alcohol dehydrogenase 3 and cytochrome P-4502E1 genotypes in susceptibility to cancers of the upper aerodigestive tract. Int J Cancer. 2000;87:734–740. [PubMed] [Google Scholar]
- 21.Harty LC, Caporaso NE, Hayes RB, Winn DM, Bravo-Otero E, Blot WJ, Kleinman DV, Brown LM, Armenian HK, Fraumeni JF, Jr, Shields PG. Alcohol dehydrogenase 3 genotype and risk of oral cavity and pharyngeal cancers. J Natl Cancer Inst. 1997;89:1698–1705. doi: 10.1093/jnci/89.22.1698. [DOI] [PubMed] [Google Scholar]
- 22.Pelucchi C, Gallus S, Garavello W, Bosetti C, La Vecchia C. Cancer risk associated with alcohol and tobacco use: focus on upper aero-digestive tract and liver. Alcohol Res Health. 2006;29:193–198. [PMC free article] [PubMed] [Google Scholar]
- 23.Anantharaman D, Marron M, Lagiou P, Samoli E, Ahrens W, Pohlabeln H, Slamova A, Schejbalova M, Merletti F, Richiardi L, Kjaerheim K, Castellsague X, Agudo A, Talamini R, Barzan L, Macfarlane TV, Tickle M, Simonato L, Canova C, Conway DI, McKinney PA, Thomson P, Znaor A, Healy CM, McCartan BE, Hashibe M, Brennan P, Macfarlane GJ. Population attributable risk of tobacco and alcohol for upper aerodigestive tract cancer. Oral Oncol. 2011;47:725–731. doi: 10.1016/j.oraloncology.2011.05.004. [DOI] [PubMed] [Google Scholar]
- 24.Bell-Parikh LC, Guengerich FP. Kinetics of cytochrome P450 2E1-catalyzed oxidation of ethanol to acetic acid via acetaldehyde. The Journal of biological chemistry. 1999;274:23833–23840. doi: 10.1074/jbc.274.34.23833. [DOI] [PubMed] [Google Scholar]
- 25.Kunitoh S, Imaoka S, Hiroi T, Yabusaki Y, Monna T, Funae Y. Acetaldehyde as well as ethanol is metabolized by human CYP2E1. J Pharmacol Exp Ther. 1997;280:527–532. [PubMed] [Google Scholar]
- 26.Seitz HK, Stickel F. Molecular mechanisms of alcohol-mediated carcinogenesis. Nat Rev Cancer. 2007;7:599–612. doi: 10.1038/nrc2191. [DOI] [PubMed] [Google Scholar]
- 27.Homann N, Jousimies-Somer H, Jokelainen K, Heine R, Salaspuro M. High acetaldehyde levels in saliva after ethanol consumption: methodological aspects and pathogenetic implications. Carcinogenesis. 1997;18:1739–1743. doi: 10.1093/carcin/18.9.1739. [DOI] [PubMed] [Google Scholar]
- 28.Homann N, Tillonen J, Meurman JH, Rintamaki H, Lindqvist C, Rautio M, Jousimies-Somer H, Salaspuro M. Increased salivary acetaldehyde levels in heavy drinkers and smokers: a microbiological approach to oral cavity cancer. Carcinogenesis. 2000;21:663–668. doi: 10.1093/carcin/21.4.663. [DOI] [PubMed] [Google Scholar]
- 29.Salaspuro MP. Acetaldehyde, microbes, and cancer of the digestive tract. Crit Rev Clin Lab Sci. 2003;40:183–208. doi: 10.1080/713609333. [DOI] [PubMed] [Google Scholar]
- 30.Poschl G, Seitz HK. Alcohol and cancer. Alcohol Alcohol. 2004;39:155–165. doi: 10.1093/alcalc/agh057. [DOI] [PubMed] [Google Scholar]
- 31.Du X, Squier CA, Kremer MJ, Wertz PW. Penetration of N-nitrosonornicotine (NNN) across oral mucosa in the presence of ethanol and nicotine. J Oral Pathol Med. 2000;29:80–85. doi: 10.1034/j.1600-0714.2000.290205.x. [DOI] [PubMed] [Google Scholar]
- 32.Kornfehl J, Temmel A, Formanek M, Knerer B. Effects of ethanol treatment of proliferation and differentiation in a head and neck squamous cell carcinoma cell line. Alcohol Clin Exp Res. 1999;23:1102–1107. [PubMed] [Google Scholar]
- 33.McKillop IH, Schrum LW. Alcohol and liver cancer. Alcohol. 2005;35:195–203. doi: 10.1016/j.alcohol.2005.04.004. [DOI] [PubMed] [Google Scholar]
- 34.Purohit V, Rapaka R, Kwon OS, Song BJ. Roles of alcohol and tobacco exposure in the development of hepatocellular carcinoma. Life Sci. 2013;92:3–9. doi: 10.1016/j.lfs.2012.10.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Guo Y, Wang X, Zhang X, Sun Z, Chen X. Ethanol promotes chemically induced oral cancer in mice through activation of the 5-lipoxygenase pathway of arachidonic acid metabolism. Cancer Prev Res (Phila) 2011;4:1863–1872. doi: 10.1158/1940-6207.CAPR-11-0206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Wang XD. Alcohol, vitamin A, and cancer. Alcohol. 2005;35:251–258. doi: 10.1016/j.alcohol.2005.04.006. [DOI] [PubMed] [Google Scholar]
- 37.Shiraishi-Yokoyama H, Yokoyama H, Matsumoto M, Imaeda H, Hibi T. Acetaldehyde inhibits the formation of retinoic acid from retinal in the rat esophagus. Scand J Gastroenterol. 2006;41:80–86. doi: 10.1080/00365520510023936. [DOI] [PubMed] [Google Scholar]
- 38.Liu L, Tang XH, Scognamiglio T, Gudas LJ. Oral carcinogenesis induced by 4-nitroquinoline 1-oxide in lecithin:retinol acyltransferase gene knockout mice. J Nutr Biochem. 2010;21:975–982. doi: 10.1016/j.jnutbio.2009.07.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Toma S, Bonelli L, Sartoris A, Mira E, Antonelli A, Beatrice F, Giordano C, Benazzo M, Caroggio A, Cavalot AL, Gandolfo S, Garozzo A, Margarino G, Schenone G, Spadini N, Sirotova Z, Zibordi F, Balzarini F, Serafini I, Miani P, Cortesina G. 13-cis retinoic acid in head and neck cancer chemoprevention: results of a randomized trial from the Italian Head and Neck Chemoprevention Study Group. Oncol Rep. 2004;11:1297–1305. [PubMed] [Google Scholar]
- 40.Tang XH, Osei-Sarfo K, Urvalek AM, Zhang T, Scognamiglio T, Gudas LJ. Combination of bexarotene and the retinoid CD1530 reduces murine oral-cavity carcinogenesis induced by the carcinogen 4-nitroquinoline 1-oxide. Proceedings of the National Academy of Sciences of the United States of America. 2014;111:8907–8912. doi: 10.1073/pnas.1404828111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Joshi PC, Mehta A, Jabber WS, Fan X, Guidot DM. Zinc deficiency mediates alcohol-induced alveolar epithelial and macrophage dysfunction in rats. American journal of respiratory cell and molecular biology. 2009;41:207–216. doi: 10.1165/rcmb.2008-0209OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Curry-McCoy TV, Guidot DM, Joshi PC. Chronic alcohol ingestion in rats decreases Kruppel-like factor 4 expression and intracellular zinc in the lung. Alcohol Clin Exp Res. 2013;37:361–371. doi: 10.1111/j.1530-0277.2012.01946.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Zhou Z, Wang L, Song Z, Saari JT, McClain CJ, Kang YJ. Zinc supplementation prevents alcoholic liver injury in mice through attenuation of oxidative stress. The American journal of pathology. 2005;166:1681–1690. doi: 10.1016/S0002-9440(10)62478-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Fong LY, Zhang L, Jiang Y, Farber JL. Dietary zinc modulation of COX-2 expression and lingual and esophageal carcinogenesis in rats. J Natl Cancer Inst. 2005;97:40–50. doi: 10.1093/jnci/dji006. [DOI] [PubMed] [Google Scholar]
- 45.Mathur A, Wallenius K, Abdulla M. Influence of zinc on onset and progression of oral carcinogenesis in rats. Acta odontologica Scandinavica. 1979;37:277–284. doi: 10.3109/00016357909004697. [DOI] [PubMed] [Google Scholar]
- 46.Liu CG, Zhang L, Jiang Y, Chatterjee D, Croce CM, Huebner K, Fong LY. Modulation of gene expression in precancerous rat esophagus by dietary zinc deficit and replenishment. Cancer research. 2005;65:7790–7799. doi: 10.1158/0008-5472.CAN-05-1345. [DOI] [PubMed] [Google Scholar]
- 47.Taccioli C, Wan SG, Liu CG, Alder H, Volinia S, Farber JL, Croce CM, Fong LY. Zinc replenishment reverses overexpression of the proinflammatory mediator S100A8 and esophageal preneoplasia in the rat. Gastroenterology. 2009;136:953–966. doi: 10.1053/j.gastro.2008.11.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Taccioli C, Chen H, Jiang Y, Liu XP, Huang K, Smalley KJ, Farber JL, Croce CM, Fong LY. Dietary zinc deficiency fuels esophageal cancer development by inducing a distinct inflammatory signature. Oncogene. 2012;31:4550–4558. doi: 10.1038/onc.2011.592. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Sun J, Liu J, Pan X, Quimby D, Zanesi N, Druck T, Pfeifer GP, Croce CM, Fong LY, Huebner K. Effect of zinc supplementation on N-nitrosomethylbenzylamine-induced forestomach tumor development and progression in tumor suppressor-deficient mouse strains. Carcinogenesis. 2011;32:351–358. doi: 10.1093/carcin/bgq251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Fong LY, Jiang Y, Rawahneh ML, Smalley KJ, Croce CM, Farber JL, Huebner K. Zinc supplementation suppresses 4-nitroquinoline 1-oxide-induced rat oral carcinogenesis. Carcinogenesis. 2011;32:554–560. doi: 10.1093/carcin/bgr004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Harrison-Findik DD. Role of alcohol in the regulation of iron metabolism. World J Gastroenterol. 2007;13:4925–4930. doi: 10.3748/wjg.v13.i37.4925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Do TH, Gaboriau F, Cannie I, Batusanski F, Ropert M, Moirand R, Brissot P, Loreal O, Lescoat G. Iron-mediated effect of alcohol on hepatocyte differentiation in HepaRG cells. Chem Biol Interact. 2013;206:117–125. doi: 10.1016/j.cbi.2013.08.016. [DOI] [PubMed] [Google Scholar]
- 53.Chen X, Yang CS. Esophageal adenocarcinoma: a review and perspectives on the mechanism of carcinogenesis and chemoprevention. Carcinogenesis. 2001;22:1119–1129. doi: 10.1093/carcin/22.8.1119. [DOI] [PubMed] [Google Scholar]
- 54.Deugnier Y. Iron and liver cancer. Alcohol. 2003;30:145–150. doi: 10.1016/s0741-8329(03)00129-0. [DOI] [PubMed] [Google Scholar]
- 55.Schieber M, Chandel NS. ROS function in redox signaling and oxidative stress. Curr Biol. 2014;24:R453–462. doi: 10.1016/j.cub.2014.03.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Nogueira V, Hay N. Molecular pathways: reactive oxygen species homeostasis in cancer cells and implications for cancer therapy. Clinical cancer research: an official journal of the American Association for Cancer Research. 2013;19:4309–4314. doi: 10.1158/1078-0432.CCR-12-1424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Lu SC, Mato JM. Role of methionine adenosyltransferase and S-adenosylmethionine in alcohol-associated liver cancer. Alcohol. 2005;35:227–234. doi: 10.1016/j.alcohol.2005.03.011. [DOI] [PubMed] [Google Scholar]
- 58.Mason JB, Choi SW. Effects of alcohol on folate metabolism: implications for carcinogenesis. Alcohol. 2005;35:235–241. doi: 10.1016/j.alcohol.2005.03.012. [DOI] [PubMed] [Google Scholar]
- 59.Zapisek WF, Cronin GM, Lyn-Cook BD, Poirier LA. The onset of oncogene hypomethylation in the livers of rats fed methyl-deficient, amino acid-defined diets. Carcinogenesis. 1992;13:1869–1872. doi: 10.1093/carcin/13.10.1869. [DOI] [PubMed] [Google Scholar]
- 60.Pogribny IP, Ross SA, Wise C, Pogribna M, Jones EA, Tryndyak VP, James SJ, Dragan YP, Poirier LA. Irreversible global DNA hypomethylation as a key step in hepatocarcinogenesis induced by dietary methyl deficiency. Mutat Res. 2006;593:80–87. doi: 10.1016/j.mrfmmm.2005.06.028. [DOI] [PubMed] [Google Scholar]
- 61.Gasche JA, Goel A. Epigenetic mechanisms in oral carcinogenesis. Future Oncol. 2012;8:1407–1425. doi: 10.2217/fon.12.138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Kuo IY, Chang JM, Jiang SS, Chen CH, Chang IS, Sheu BS, Lu PJ, Chang WL, Lai WW, Wang YC. Prognostic CpG methylation biomarkers identified by methylation array in esophageal squamous cell carcinoma patients. Int J Med Sci. 2014;11:779–787. doi: 10.7150/ijms.7405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Jithesh PV, Risk JM, Schache AG, Dhanda J, Lane B, Liloglou T, Shaw RJ. The epigenetic landscape of oral squamous cell carcinoma. Br J Cancer. 2013;108:370–379. doi: 10.1038/bjc.2012.568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Pan QF, Li WT, Dong HC, Chen YZ, Yin L, Liu W, Wang WW, Liu D, Li SG, Gu WY, Chen JZ, Yang L, Zhang WJ, Li F. PTEN hypermethylation profiles of Chinese Kazakh patients with esophageal squamous cell carcinoma. Dis Esophagus. 2014;27:396–402. doi: 10.1111/dote.12106. [DOI] [PubMed] [Google Scholar]
- 65.Chan SL, Cui Y, van Hasselt A, Li H, Srivastava G, Jin H, Ng KM, Wang Y, Lee KY, Tsao GS, Zhong S, Robertson KD, Rha SY, Chan AT, Tao Q. The tumor suppressor Wnt inhibitory factor 1 is frequently methylated in nasopharyngeal and esophageal carcinomas. Lab Invest. 2007;87:644–650. doi: 10.1038/labinvest.3700547. [DOI] [PubMed] [Google Scholar]
- 66.Biggar KK, Li SS. Non-histone protein methylation as a regulator of cellular signalling and function. Nat Rev Mol Cell Biol. 2015;16:5–17. doi: 10.1038/nrm3915. [DOI] [PubMed] [Google Scholar]
- 67.Ogden GR. Alcohol and oral cancer. Alcohol. 2005;35:169–173. doi: 10.1016/j.alcohol.2005.04.002. [DOI] [PubMed] [Google Scholar]
- 68.Dong YJ, Peng TK, Yin SJ. Expression and activities of class IV alcohol dehydrogenase and class III aldehyde dehydrogenase in human mouth. Alcohol. 1996;13:257–262. doi: 10.1016/0741-8329(95)02052-7. [DOI] [PubMed] [Google Scholar]
- 69.Shimizu M, Lasker JM, Tsutsumi M, Lieber CS. Immunohistochemical localization of ethanol-inducible P450IIE1 in the rat alimentary tract. Gastroenterology. 1990;99:1044–1053. doi: 10.1016/0016-5085(90)90625-b. [DOI] [PubMed] [Google Scholar]
- 70.Lechevrel M, Wild CP. Absence of a differential induction of cytochrome P450 2E1 by different alcoholic beverages in rats: implications for the aetiology of human oesophageal cancer. Archives of toxicology. 1997;71:690–695. doi: 10.1007/s002040050446. [DOI] [PubMed] [Google Scholar]
- 71.Yang SP, Medling T, Raner GM. Cytochrome P450 expression and activities in rat, rabbit and bovine tongue, Comparative biochemistry and physiology. Toxicology & pharmacology: CBP. 2003;136:297–308. doi: 10.1016/j.cca.2003.10.003. [DOI] [PubMed] [Google Scholar]
- 72.Albano E. Alcohol, oxidative stress and free radical damage. Proc Nutr Soc. 2006;65:278–290. doi: 10.1079/pns2006496. [DOI] [PubMed] [Google Scholar]
- 73.Eskelson CD, Odeleye OE, Watson RR, Earnest DL, Mufti SI. Modulation of cancer growth by vitamin E and alcohol. Alcohol Alcohol. 1993;28:117–125. [PubMed] [Google Scholar]
- 74.Yang SF, Chang CW, Wei RJ, Shiue YL, Wang SN, Yeh YT. Involvement of DNA damage response pathways in hepatocellular carcinoma. BioMed research international. 2014;2014:153867. doi: 10.1155/2014/153867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Brooks PJ, Zakhari S. Acetaldehyde and the genome: beyond nuclear DNA adducts and carcinogenesis. Environ Mol Mutagen. 2014;55:77–91. doi: 10.1002/em.21824. [DOI] [PubMed] [Google Scholar]
- 76.Yu HS, Oyama T, Matsuda T, Isse T, Yamaguchi T, Tanaka M, Tsuji M, Kawamoto T. The effect of ethanol on the formation of N2-ethylidene-dG adducts in mice: implications for alcohol-related carcinogenicity of the oral cavity and esophagus. Biomarkers. 2012;17:269–274. doi: 10.3109/1354750X.2012.666675. [DOI] [PubMed] [Google Scholar]
- 77.Balbo S, Meng L, Bliss RL, Jensen JA, Hatsukami DK, Hecht SS. Kinetics of DNA adduct formation in the oral cavity after drinking alcohol. Cancer Epidemiol Biomarkers Prev. 2012;21:601–608. doi: 10.1158/1055-9965.EPI-11-1175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Yukawa Y, Muto M, Hori K, Nagayoshi H, Yokoyama A, Chiba T, Matsuda T. Combination of ADH1B*2/ALDH2*2 polymorphisms alters acetaldehyde-derived DNA damage in the blood of Japanese alcoholics. Cancer Sci. 2012;103:1651–1655. doi: 10.1111/j.1349-7006.2012.02360.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Choudhari SK, Chaudhary M, Gadbail AR, Sharma A, Tekade S. Oxidative and antioxidative mechanisms in oral cancer and precancer: a review. Oral Oncol. 2013;50:10–18. doi: 10.1016/j.oraloncology.2013.09.011. [DOI] [PubMed] [Google Scholar]
- 80.Warnakulasuriya S, Parkkila S, Nagao T, Preedy VR, Pasanen M, Koivisto H, Niemela O. Demonstration of ethanol-induced protein adducts in oral leukoplakia (pre-cancer) and cancer. J Oral Pathol Med. 2008;37:157–165. doi: 10.1111/j.1600-0714.2007.00605.x. [DOI] [PubMed] [Google Scholar]
- 81.Roychowdhury S, McMullen MR, Pritchard MT, Li W, Salomon RG, Nagy LE. Formation of gamma-ketoaldehyde-protein adducts during ethanol-induced liver injury in mice. Free radical biology & medicine. 2009;47:1526–1538. doi: 10.1016/j.freeradbiomed.2009.07.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Niemela O, Parkkila S, Worrall S, Emery PW, Preedy VR. Generation of aldehyde-derived protein modifications in ethanol-exposed heart. Alcohol Clin Exp Res. 2003;27:1987–1992. doi: 10.1097/01.ALC.0000099260.58926.F2. [DOI] [PubMed] [Google Scholar]
- 83.Aleynik SI, Leo MA, Aleynik MK, Lieber CS. Increased circulating products of lipid peroxidation in patients with alcoholic liver disease. Alcohol Clin Exp Res. 1998;22:192–196. [PubMed] [Google Scholar]
- 84.Shepard BD, Tuma PL. Alcohol-induced protein hyperacetylation: mechanisms and consequences. World J Gastroenterol. 2009;15:1219–1230. doi: 10.3748/wjg.15.1219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Wu KC, Liu J, Klaassen CD. Role of Nrf2 in preventing ethanol-induced oxidative stress and lipid accumulation. Toxicol Appl Pharmacol. 2012;262:321–329. doi: 10.1016/j.taap.2012.05.010. [DOI] [PubMed] [Google Scholar]
- 86.Buelna-Chontal M, Zazueta C. Redox activation of Nrf2 & NF-kappaB: a double end sword? Cell Signal. 2013;25:2548–2557. doi: 10.1016/j.cellsig.2013.08.007. [DOI] [PubMed] [Google Scholar]
- 87.Ganan-Gomez I, Wei Y, Yang H, Boyano-Adanez MC, Garcia-Manero G. Oncogenic functions of the transcription factor Nrf2. Free radical biology & medicine. 2013;65:750–764. doi: 10.1016/j.freeradbiomed.2013.06.041. [DOI] [PubMed] [Google Scholar]
- 88.Ohkoshi A, Suzuki T, Ono M, Kobayashi T, Yamamoto M. Roles of Keap1-Nrf2 system in upper aerodigestive tract carcinogenesis. Cancer Prev Res (Phila) 2013;6:149–159. doi: 10.1158/1940-6207.CAPR-12-0401-T. [DOI] [PubMed] [Google Scholar]
- 89.Jensen JS, Fan X, Guidot DM. Alcohol causes alveolar epithelial oxidative stress by inhibiting the nuclear factor (erythroid-derived 2)-like 2-antioxidant response element signaling pathway. American journal of respiratory cell and molecular biology. 2013;48:511–517. doi: 10.1165/rcmb.2012-0334OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Ushida Y, Talalay P. Sulforaphane accelerates acetaldehyde metabolism by inducing aldehyde dehydrogenases: relevance to ethanol intolerance. Alcohol Alcohol. 2013;48:526–534. doi: 10.1093/alcalc/agt063. [DOI] [PubMed] [Google Scholar]
- 91.Crabb DW, Galli A, Fischer M, You M. Molecular mechanisms of alcoholic fatty liver: role of peroxisome proliferator-activated receptor alpha. Alcohol. 2004;34:35–38. doi: 10.1016/j.alcohol.2004.07.005. [DOI] [PubMed] [Google Scholar]
- 92.Sozio M, Crabb DW. Alcohol and lipid metabolism. Am J Physiol Endocrinol Metab. 2008;295:E10–16. doi: 10.1152/ajpendo.00011.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Fernando H, Bhopale KK, Kondraganti S, Kaphalia BS, Shakeel Ansari GA. Lipidomic changes in rat liver after long-term exposure to ethanol. Toxicol Appl Pharmacol. 2011;255:127–137. doi: 10.1016/j.taap.2011.05.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Lieber CS. Alcoholic fatty liver: its pathogenesis and mechanism of progression to inflammation and fibrosis. Alcohol. 2004;34:9–19. doi: 10.1016/j.alcohol.2004.07.008. [DOI] [PubMed] [Google Scholar]
- 95.Lewis JA, Elkon IM, McGee MA, Higbee AJ, Gasch AP. Exploiting natural variation in Saccharomyces cerevisiae to identify genes for increased ethanol resistance. Genetics. 2010;186:1197–1205. doi: 10.1534/genetics.110.121871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Stanley D, Bandara A, Fraser S, Chambers PJ, Stanley GA. The ethanol stress response and ethanol tolerance of Saccharomyces cerevisiae. J Appl Microbiol. 2010;109:13–24. doi: 10.1111/j.1365-2672.2009.04657.x. [DOI] [PubMed] [Google Scholar]
- 97.Blume JJ, Halbach A, Behrendt D, Paulsson M, Plomann M. EHD proteins are associated with tubular and vesicular compartments and interact with specific phospholipids. Exp Cell Res. 2007;313:219–231. doi: 10.1016/j.yexcr.2006.10.006. [DOI] [PubMed] [Google Scholar]
- 98.Naslavsky N, Rahajeng J, Rapaport D, Horowitz M, Caplan S. EHD1 regulates cholesterol homeostasis and lipid droplet storage. Biochem Biophys Res Commun. 2007;357:792–799. doi: 10.1016/j.bbrc.2007.04.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Shimano H. Novel qualitative aspects of tissue fatty acids related to metabolic regulation: lessons from Elovl6 knockout. Prog Lipid Res. 2012;51:267–271. doi: 10.1016/j.plipres.2011.12.004. [DOI] [PubMed] [Google Scholar]
- 100.Matsuzaka T, Shimano H, Yahagi N, Kato T, Atsumi A, Yamamoto T, Inoue N, Ishikawa M, Okada S, Ishigaki N, Iwasaki H, Iwasaki Y, Karasawa T, Kumadaki S, Matsui T, Sekiya M, Ohashi K, Hasty AH, Nakagawa Y, Takahashi A, Suzuki H, Yatoh S, Sone H, Toyoshima H, Osuga J, Yamada N. Crucial role of a long-chain fatty acid elongase, Elovl6, in obesity-induced insulin resistance. Nat Med. 2007;13:1193–1202. doi: 10.1038/nm1662. [DOI] [PubMed] [Google Scholar]
- 101.Zadravec D, Brolinson A, Fisher RM, Carneheim C, Csikasz RI, Bertrand-Michel J, Boren J, Guillou H, Rudling M, Jacobsson A. Ablation of the very-long-chain fatty acid elongase ELOVL3 in mice leads to constrained lipid storage and resistance to diet-induced obesity. FASEB J. 2010;24:4366–4377. doi: 10.1096/fj.09-152298. [DOI] [PubMed] [Google Scholar]
- 102.Kobayashi T, Fujimori K. Very long-chain-fatty acids enhance adipogenesis through coregulation of Elovl3 and PPARgamma in 3T3-L1 cells. Am J Physiol Endocrinol Metab. 2012;302:E1461–1471. doi: 10.1152/ajpendo.00623.2011. [DOI] [PubMed] [Google Scholar]
- 103.Sonmez M, Ince HY, Yalcin O, Ajdzanovic V, Spasojevic I, Meiselman HJ, Baskurt OK. The effect of alcohols on red blood cell mechanical properties and membrane fluidity depends on their molecular size. PLoS One. 2013;8:e76579. doi: 10.1371/journal.pone.0076579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Frischknecht AL, Frink LJ. Alcohols reduce lateral membrane pressures: predictions from molecular theory. Biophysical journal. 2006;91:4081–4090. doi: 10.1529/biophysj.106.091918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Chiou JS, Ma SM, Kamaya H, Ueda I. Anesthesia cutoff phenomenon: interfacial hydrogen bonding. Science. 1990;248:583–585. doi: 10.1126/science.2159183. [DOI] [PubMed] [Google Scholar]
- 106.Bodhinathan K, Slesinger PA. Molecular mechanism underlying ethanol activation of G-protein-gated inwardly rectifying potassium channels. Proceedings of the National Academy of Sciences of the United States of America. 2013;110:18309–18314. doi: 10.1073/pnas.1311406110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Szabo G, Dolganiuc A, Dai Q, Pruett SB. TLR4, ethanol, and lipid rafts: a new mechanism of ethanol action with implications for other receptor-mediated effects. Journal of immunology. 2007;178:1243–1249. doi: 10.4049/jimmunol.178.3.1243. [DOI] [PubMed] [Google Scholar]
- 108.Sun Z, Luo Q, Ye D, Chen W, Chen F. Role of toll-like receptor 4 on the immune escape of human oral squamous cell carcinoma and resistance of cisplatin-induced apoptosis. Mol Cancer. 2012;11:33. doi: 10.1186/1476-4598-11-33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Ren WH, Zhang LM, Liu HQ, Gao L, Chen C, Qiang C, Wang XL, Liu CY, Li SM, Huang C, Qi H, Zhi KQ. Protein overexpression of CIRP and TLR4 in oral squamous cell carcinoma: an immunohistochemical and clinical correlation analysis. Med Oncol. 2014;31:120. doi: 10.1007/s12032-014-0120-7. [DOI] [PubMed] [Google Scholar]
- 110.Sheyhidin I, Nabi G, Hasim A, Zhang RP, Ainiwaer J, Ma H, Wang H. Overexpression of TLR3, TLR4, TLR7 and TLR9 in esophageal squamous cell carcinoma. World J Gastroenterol. 2011;17:3745–3751. doi: 10.3748/wjg.v17.i32.3745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Szczepanski MJ, Czystowska M, Szajnik M, Harasymczuk M, Boyiadzis M, Kruk-Zagajewska A, Szyfter W, Zeromski J, Whiteside TL. Triggering of Toll-like receptor 4 expressed on human head and neck squamous cell carcinoma promotes tumor development and protects the tumor from immune attack. Cancer research. 2009;69:3105–3113. doi: 10.1158/0008-5472.CAN-08-3838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Mark R, Bermejo JL, Bierhaus A, Plinkert PK, Angel P, Hess J. The receptor for advanced glycation end products is dispensable in a mouse model of oral and esophageal carcinogenesis. Histol Histopathol. 2013;28:1585–1594. doi: 10.14670/HH-28.1585. [DOI] [PubMed] [Google Scholar]
- 113.Vahle AK, Kerem A, Ozturk E, Bankfalvi A, Lang S, Brandau S. Optimization of an orthotopic murine model of head and neck squamous cell carcinoma in fully immunocompetent mice–role of toll-like-receptor 4 expressed on host cells. Cancer Lett. 2012;317:199–206. doi: 10.1016/j.canlet.2011.11.027. [DOI] [PubMed] [Google Scholar]
- 114.Paleja B, Anand A, Chaukar D, D’Cruz A, Chiplunkar S. Decreased functional response to Toll like receptor ligands in patients with oral cancer. Hum Immunol. 2013;74:927–936. doi: 10.1016/j.humimm.2013.04.018. [DOI] [PubMed] [Google Scholar]
- 115.Borggrefe T, Oswald F. The Notch signaling pathway: transcriptional regulation at Notch target genes. Cell Mol Life Sci. 2009;66:1631–1646. doi: 10.1007/s00018-009-8668-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Okuyama R, Tagami H, Aiba S. Notch signaling: its role in epidermal homeostasis and in the pathogenesis of skin diseases. Journal of dermatological science. 2008;49:187–194. doi: 10.1016/j.jdermsci.2007.05.017. [DOI] [PubMed] [Google Scholar]
- 117.Ohashi S, Natsuizaka M, Yashiro-Ohtani Y, Kalman RA, Nakagawa M, Wu L, Klein-Szanto AJ, Herlyn M, Diehl JA, Katz JP, Pear WS, Seykora JT, Nakagawa H. NOTCH1 and NOTCH3 coordinate esophageal squamous differentiation through a CSL-dependent transcriptional network. Gastroenterology. 2010;139:2113–2123. doi: 10.1053/j.gastro.2010.08.040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Ohashi S, Natsuizaka M, Naganuma S, Kagawa S, Kimura S, Itoh H, Kalman RA, Nakagawa M, Darling DS, Basu D, Gimotty PA, Klein-Szanto AJ, Diehl JA, Herlyn M, Nakagawa H. A NOTCH3-mediated squamous cell differentiation program limits expansion of EMT-competent cells that express the ZEB transcription factors. Cancer research. 2011;71:6836–6847. doi: 10.1158/0008-5472.CAN-11-0846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Blanpain C, Lowry WE, Pasolli HA, Fuchs E. Canonical notch signaling functions as a commitment switch in the epidermal lineage. Genes & development. 2006;20:3022–3035. doi: 10.1101/gad.1477606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Wang X, Pasolli HA, Williams T, Fuchs E. AP-2 factors act in concert with Notch to orchestrate terminal differentiation in skin epidermis. The Journal of cell biology. 2008;183:37–48. doi: 10.1083/jcb.200804030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Casey LM, Lan Y, Cho ES, Maltby KM, Gridley T, Jiang R. Jag2-Notch1 signaling regulates oral epithelial differentiation and palate development. Dev Dyn. 2006;235:1830–1844. doi: 10.1002/dvdy.20821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Agrawal N, Frederick MJ, Pickering CR, Bettegowda C, Chang K, Li RJ, Fakhry C, Xie TX, Zhang J, Wang J, Zhang N, El-Naggar AK, Jasser SA, Weinstein JN, Trevino L, Drummond JA, Muzny DM, Wu Y, Wood LD, Hruban RH, Westra WH, Koch WM, Califano JA, Gibbs RA, Sidransky D, Vogelstein B, Velculescu VE, Papadopoulos N, Wheeler DA, Kinzler KW, Myers JN. Exome sequencing of head and neck squamous cell carcinoma reveals inactivating mutations in NOTCH1. Science. 2011;333:1154–1157. doi: 10.1126/science.1206923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Stransky N, Egloff AM, Tward AD, Kostic AD, Cibulskis K, Sivachenko A, Kryukov GV, Lawrence MS, Sougnez C, McKenna A, Shefler E, Ramos AH, Stojanov P, Carter SL, Voet D, Cortes ML, Auclair D, Berger MF, Saksena G, Guiducci C, Onofrio RC, Parkin M, Romkes M, Weissfeld JL, Seethala RR, Wang L, Rangel-Escareno C, Fernandez-Lopez JC, Hidalgo-Miranda A, Melendez-Zajgla J, Winckler W, Ardlie K, Gabriel SB, Meyerson M, Lander ES, Getz G, Golub TR, Garraway LA, Grandis JR. The mutational landscape of head and neck squamous cell carcinoma. Science. 2011;333:1157–1160. doi: 10.1126/science.1208130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Agrawal N, Jiao Y, Bettegowda C, Hutfless SM, Wang Y, David S, Cheng Y, Twaddell WS, Latt NL, Shin EJ, Wang LD, Wang L, Yang W, Velculescu VE, Vogelstein B, Papadopoulos N, Kinzler KW, Meltzer SJ. Comparative genomic analysis of esophageal adenocarcinoma and squamous cell carcinoma. Cancer discovery. 2012;2:899–905. doi: 10.1158/2159-8290.CD-12-0189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.C. India Project Team of the International Cancer Genome. Mutational landscape of gingivo-buccal oral squamous cell carcinoma reveals new recurrently-mutated genes and molecular subgroups. Nature communications. 2013;4:2873. doi: 10.1038/ncomms3873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Song X, Xia R, Li J, Long Z, Ren H, Chen W, Mao L. Common and complex notch1 mutations in chinese oral squamous cell carcinoma. Clinical cancer research: an official journal of the American Association for Cancer Research. 2014;20:701–710. doi: 10.1158/1078-0432.CCR-13-1050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Pickering CR, Zhang J, Yoo SY, Bengtsson L, Moorthy S, Neskey DM, Zhao M, Ortega Alves MV, Chang K, Drummond J, Cortez E, Xie TX, Zhang D, Chung W, Issa JP, Zweidler-McKay PA, Wu X, El-Naggar AK, Weinstein JN, Wang J, Muzny DM, Gibbs RA, Wheeler DA, Myers JN, Frederick MJ. Integrative genomic characterization of oral squamous cell carcinoma identifies frequent somatic drivers. Cancer discovery. 2013;3:770–781. doi: 10.1158/2159-8290.CD-12-0537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Schneider KJ, Scheer M, Suhr M, Clemens DL. Ethanol administration impairs pancreatic repair after injury. Pancreas. 2012;41:1272–1279. doi: 10.1097/MPA.0b013e31824bde37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Morrow D, Cullen JP, Liu W, Cahill PA, Redmond EM. Alcohol inhibits smooth muscle cell proliferation via regulation of the Notch signaling pathway. Arteriosclerosis, thrombosis, and vascular biology. 2010;30:2597–2603. doi: 10.1161/ATVBAHA.110.215681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Hashimoto-Torii K, Kawasawa YI, Kuhn A, Rakic P. Combined transcriptome analysis of fetal human and mouse cerebral cortex exposed to alcohol. Proceedings of the National Academy of Sciences of the United States of America. 2011;108:4212–4217. doi: 10.1073/pnas.1100903108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Morrow D, Cullen JP, Cahill PA, Redmond EM. Ethanol stimulates endothelial cell angiogenic activity via a Notch- and angiopoietin-1-dependent pathway. Cardiovascular research. 2008;79:313–321. doi: 10.1093/cvr/cvn108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.McCarthy N, Eberhart JK. Gene-ethanol interactions underlying fetal alcohol spectrum disorders. Cell Mol Life Sci. 2014 doi: 10.1007/s00018-014-1578-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Mao H, Diehl AM, Li YX. Sonic hedgehog ligand partners with caveolin-1 for intracellular transport. Lab Invest. 2009;89:290–300. doi: 10.1038/labinvest.2008.163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Zhang C, Ojiaku P, Cole GJ. Forebrain and hindbrain development in zebrafish is sensitive to ethanol exposure involving agrin, Fgf, and sonic hedgehog function. Birth Defects Res A Clin Mol Teratol. 2013;97:8–27. doi: 10.1002/bdra.23099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Zhang C, Turton QM, Mackinnon S, Sulik KK, Cole GJ. Agrin function associated with ocular development is a target of ethanol exposure in embryonic zebrafish. Birth Defects Res A Clin Mol Teratol. 2011;91:129–141. doi: 10.1002/bdra.20766. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Loucks EJ, Ahlgren SC. Deciphering the role of Shh signaling in axial defects produced by ethanol exposure. Birth Defects Res A Clin Mol Teratol. 2009;85:556–567. doi: 10.1002/bdra.20564. [DOI] [PubMed] [Google Scholar]
- 137.Li YX, Yang HT, Zdanowicz M, Sicklick JK, Qi Y, Camp TJ, Diehl AM. Fetal alcohol exposure impairs Hedgehog cholesterol modification and signaling. Lab Invest. 2007;87:231–240. doi: 10.1038/labinvest.3700516. [DOI] [PubMed] [Google Scholar]
- 138.Chan IS, Guy CD, Machado MV, Wank A, Kadiyala V, Michelotti G, Choi S, Swiderska-Syn M, Karaca G, Pereira TA, Yip-Schneider MT, Max Schmidt C, Diehl AM. Alcohol Activates the Hedgehog Pathway and Induces Related Procarcinogenic Processes in the Alcohol-Preferring Rat Model of Hepatocarcinogenesis. Alcohol Clin Exp Res. 2013 doi: 10.1111/acer.12279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Jung Y, Brown KD, Witek RP, Omenetti A, Yang L, Vandongen M, Milton RJ, Hines IN, Rippe RA, Spahr L, Rubbia-Brandt L, Diehl AM. Accumulation of hedgehog-responsive progenitors parallels alcoholic liver disease severity in mice and humans. Gastroenterology. 2008;134:1532–1543. doi: 10.1053/j.gastro.2008.02.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Yang L, Wang LS, Chen XL, Gatalica Z, Qiu S, Liu Z, Stoner G, Zhang H, Weiss H, Xie J. Hedgehog signaling activation in the development of squamous cell carcinoma and adenocarcinoma of esophagus. Int J Biochem Mol Biol. 2012;3:46–57. [PMC free article] [PubMed] [Google Scholar]
- 141.Wei L, Xu Z. Cross-signaling among phosphinositide-3 kinase, mitogen-activated protein kinase and sonic hedgehog pathways exists in esophageal cancer. Int J Cancer. 2011;129:275–284. doi: 10.1002/ijc.25673. [DOI] [PubMed] [Google Scholar]
- 142.Wang Y, Ding Q, Yen CJ, Xia W, Izzo JG, Lang JY, Li CW, Hsu JL, Miller SA, Wang X, Lee DF, Hsu JM, Huo L, Labaff AM, Liu D, Huang TH, Lai CC, Tsai FJ, Chang WC, Chen CH, Wu TT, Buttar NS, Wang KK, Wu Y, Wang H, Ajani J, Hung MC. The crosstalk of mTOR/S6K1 and Hedgehog pathways. Cancer Cell. 2012;21:374–387. doi: 10.1016/j.ccr.2011.12.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Wang YF, Chang CJ, Lin CP, Chang SY, Chu PY, Tai SK, Li WY, Chao KS, Chen YJ. Expression of hedgehog signaling molecules as a prognostic indicator of oral squamous cell carcinoma. Head Neck. 2012;34:1556–1561. doi: 10.1002/hed.21958. [DOI] [PubMed] [Google Scholar]
- 144.Cavicchioli Buim ME, Gurgel CA, Goncalves Ramos EA, Lourenco SV, Soares FA. Activation of sonic hedgehog signaling in oral squamous cell carcinomas: a preliminary study. Human pathology. 2011;42:1484–1490. doi: 10.1016/j.humpath.2010.12.015. [DOI] [PubMed] [Google Scholar]
- 145.Anastas JN, Moon RT. WNT signalling pathways as therapeutic targets in cancer. Nat Rev Cancer. 2013;13:11–26. doi: 10.1038/nrc3419. [DOI] [PubMed] [Google Scholar]
- 146.Lecarpentier Y, Claes V, Duthoit G, Hebert JL. Circadian rhythms, Wnt/beta-catenin pathway and PPAR alpha/gamma profiles in diseases with primary or secondary cardiac dysfunction. Front Physiol. 2014;5:429. doi: 10.3389/fphys.2014.00429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Vangipuram SD, Lyman WD. Ethanol affects differentiation-related pathways and suppresses Wnt signaling protein expression in human neural stem cells. Alcohol Clin Exp Res. 2012;36:788–797. doi: 10.1111/j.1530-0277.2011.01682.x. [DOI] [PubMed] [Google Scholar]
- 148.Mercer KE, Hennings L, Sharma N, Lai K, Cleves MA, Wynne RA, Badger TM, Ronis MJ. Alcohol consumption promotes diethylnitrosamine-induced hepatocarcinogenesis in male mice through activation of the Wnt/beta-catenin signaling pathway. Cancer Prev Res (Phila) 2014;7:675–685. doi: 10.1158/1940-6207.CAPR-13-0444-T. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Santoro A, Pannone G, Papagerakis S, McGuff HS, Cafarelli B, Lepore S, De Maria S, Rubini C, Mattoni M, Staibano S, Mezza E, De Rosa G, Aquino G, Losito S, Loreto C, Crimi S, Bufo P, Lo Muzio L. Beta-catenin and epithelial tumors: a study based on 374 oropharyngeal cancers. BioMed research international. 2014;2014:948264. doi: 10.1155/2014/948264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Osei-Sarfo K, Tang XH, Urvalek AM, Scognamiglio T, Gudas LJ. The molecular features of tongue epithelium treated with the carcinogen 4-nitroquinoline-1-oxide and alcohol as a model for HNSCC. Carcinogenesis. 2013;34:2673–2681. doi: 10.1093/carcin/bgt223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.White RA, Malkoski SP, Wang XJ. TGFbeta signaling in head and neck squamous cell carcinoma. Oncogene. 2010;29:5437–5446. doi: 10.1038/onc.2010.306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Principe DR, Doll JA, Bauer J, Jung B, Munshi HG, Bartholin L, Pasche B, Lee C, Grippo PJ. TGF-beta: Duality of function between tumor prevention and carcinogenesis. J Natl Cancer Inst. 2014;106 doi: 10.1093/jnci/djt369. djt369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Pickup M, Novitskiy S, Moses HL. The roles of TGFbeta in the tumour microenvironment. Nat Rev Cancer. 2013;13:788–799. doi: 10.1038/nrc3603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Miller MW. Expression of transforming growth factor-beta in developing rat cerebral cortex: effects of prenatal exposure to ethanol. The Journal of comparative neurology. 2003;460:410–424. doi: 10.1002/cne.10658. [DOI] [PubMed] [Google Scholar]
- 155.Gerjevic LN, Liu N, Lu S, Harrison-Findik DD. Alcohol Activates TGF-Beta but Inhibits BMP Receptor-Mediated Smad Signaling and Smad4 Binding to Hepcidin Promoter in the Liver. International journal of hepatology. 2012;2012:459278. doi: 10.1155/2012/459278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Brown SD, Brown LA. Ethanol (EtOH)-induced TGF-beta1 and reactive oxygen species production are necessary for EtOH-induced alveolar macrophage dysfunction and induction of alternative activation. Alcohol Clin Exp Res. 2012;36:1952–1962. doi: 10.1111/j.1530-0277.2012.01825.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Law BA, Carver WE. Activation of cardiac fibroblasts by ethanol is blocked by TGF-beta inhibition. Alcohol Clin Exp Res. 2013;37:1286–1294. doi: 10.1111/acer.12111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Tahir M, Rehman MU, Lateef A, Khan R, Khan AQ, Qamar W, Ali F, O’Hamiza O, Sultana S. Diosmin protects against ethanol-induced hepatic injury via alleviation of inflammation and regulation of TNF-alpha and NF-kappaB activation. Alcohol. 2013;47:131–139. doi: 10.1016/j.alcohol.2012.12.010. [DOI] [PubMed] [Google Scholar]
- 159.Iimuro Y, Gallucci RM, Luster MI, Kono H, Thurman RG. Antibodies to tumor necrosis factor alfa attenuate hepatic necrosis and inflammation caused by chronic exposure to ethanol in the rat. Hepatology. 1997;26:1530–1537. doi: 10.1002/hep.510260621. [DOI] [PubMed] [Google Scholar]
- 160.Ye Q, Wang X, Wang Q, Xia M, Zhu Y, Lian F, Ling W. Cytochrome P4502E1 inhibitor, chlormethiazole, decreases lipopolysaccharide-induced inflammation in rat Kupffer cells with ethanol treatment. Hepatol Res. 2013;43:1115–1123. doi: 10.1111/hepr.12063. [DOI] [PubMed] [Google Scholar]
- 161.Lin CW, Hsieh YS, Hsin CH, Su CW, Lin CH, Wei LH, Yang SF, Chien MH. Effects of NFKB1 and NFKBIA gene polymorphisms on susceptibility to environmental factors and the clinicopathologic development of oral cancer. PLoS One. 2012;7:e35078. doi: 10.1371/journal.pone.0035078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Aroor AR, Shukla SD. MAP kinase signaling in diverse effects of ethanol. Life Sci. 2004;74:2339–2364. doi: 10.1016/j.lfs.2003.11.001. [DOI] [PubMed] [Google Scholar]
- 163.Broek RV, Mohan S, Eytan DF, Chen Z, Van Waes C. The PI3K/Akt/mTOR axis in head and neck cancer: functions, aberrations, crosstalk, and therapies. Oral Dis. 2013 doi: 10.1111/odi.12206. [DOI] [PubMed] [Google Scholar]
- 164.Zeng T, Zhang CL, Song FY, Zhao XL, Yu LH, Zhu ZP, Xie KQ. PI3K/Akt pathway activation was involved in acute ethanol-induced fatty liver in mice. Toxicology. 2012;296:56–66. doi: 10.1016/j.tox.2012.03.005. [DOI] [PubMed] [Google Scholar]
- 165.Neasta J, Ben Hamida S, Yowell QV, Carnicella S, Ron D. AKT signaling pathway in the nucleus accumbens mediates excessive alcohol drinking behaviors. Biol Psychiatry. 2011;70:575–582. doi: 10.1016/j.biopsych.2011.03.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Liu L, Cao JX, Sun B, Li HL, Xia Y, Wu Z, Tang CL, Hu J. Mesenchymal stem cells inhibition of chronic ethanol-induced oxidative damage via upregulation of phosphatidylinositol-3-kinase/Akt and modulation of extracellular signal-regulated kinase 1/2 activation in PC12 cells and neurons. Neuroscience. 2010;167:1115–1124. doi: 10.1016/j.neuroscience.2010.01.057. [DOI] [PubMed] [Google Scholar]
- 167.Umoh NA, Walker RK, Al-Rubaiee M, Jeffress MA, Haddad GE. Acute alcohol modulates cardiac function as PI3K/Akt regulates oxidative stress. Alcohol Clin Exp Res. 2014;38:1847–1864. doi: 10.1111/acer.12459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Thomas SM, Bhola NE, Zhang Q, Contrucci SC, Wentzel AL, Freilino ML, Gooding WE, Siegfried JM, Chan DC, Grandis JR. Cross-talk between G protein-coupled receptor and epidermal growth factor receptor signaling pathways contributes to growth and invasion of head and neck squamous cell carcinoma. Cancer research. 2006;66:11831–11839. doi: 10.1158/0008-5472.CAN-06-2876. [DOI] [PubMed] [Google Scholar]
- 169.Agudelo M, Yndart A, Morrison M, Figueroa G, Munoz K, Samikkannu T, Nair MP. Differential expression and functional role of cannabinoid genes in alcohol users. Drug Alcohol Depend. 2013;133:789–793. doi: 10.1016/j.drugalcdep.2013.08.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Kelm MK, Criswell HE, Breese GR. Ethanol-enhanced GABA release: a focus on G protein-coupled receptors. Brain Res Rev. 2011;65:113–123. doi: 10.1016/j.brainresrev.2010.09.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Homann N, Karkkainen P, Koivisto T, Nosova T, Jokelainen K, Salaspuro M. Effects of acetaldehyde on cell regeneration and differentiation of the upper gastrointestinal tract mucosa. J Natl Cancer Inst. 1997;89:1692–1697. doi: 10.1093/jnci/89.22.1692. [DOI] [PubMed] [Google Scholar]
- 172.Mascres C, Ming-Wen F, Joly JG. Morphologic changes of the esophageal mucosa in the rat after chronic alcohol ingestion. Exp Pathol. 1984;25:147–153. doi: 10.1016/s0232-1513(84)80033-x. [DOI] [PubMed] [Google Scholar]
- 173.Wimberly AL, Forsyth CB, Khan MW, Pemberton A, Khazaie K, Keshavarzian A. Ethanol-induced mast cell-mediated inflammation leads to increased susceptibility of intestinal tumorigenesis in the APC Delta468 min mouse model of colon cancer. Alcohol Clin Exp Res. 2013;37(Suppl 1):E199–208. doi: 10.1111/j.1530-0277.2012.01894.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Yu K, Yang J, Wang F, Chen L, Lu Y, Luo J, Wang S. Ethanol supports macrophage recruitment and reinforces invasion and migration of lewis lung carcinoma. Alcohol Clin Exp Res. 2014;38:2597–2606. doi: 10.1111/acer.12512. [DOI] [PubMed] [Google Scholar]
- 175.Polakis P. Wnt signaling in cancer. Cold Spring Harb Perspect Biol. 2012;4 doi: 10.1101/cshperspect.a008052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Kwon C, Cheng P, King IN, Andersen P, Shenje L, Nigam V, Srivastava D. Notch post-translationally regulates beta-catenin protein in stem and progenitor cells. Nat Cell Biol. 2011;13:1244–1251. doi: 10.1038/ncb2313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Wakabayashi N, Shin S, Slocum SL, Agoston ES, Wakabayashi J, Kwak MK, Misra V, Biswal S, Yamamoto M, Kensler TW. Regulation of notch1 signaling by nrf2: implications for tissue regeneration. Science signaling. 2010;3:ra52. doi: 10.1126/scisignal.2000762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Soffritti M, Belpoggi F, Cevolani D, Guarino M, Padovani M, Maltoni C. Results of long-term experimental studies on the carcinogenicity of methyl alcohol and ethyl alcohol in rats. Ann N Y Acad Sci. 2002;982:46–69. doi: 10.1111/j.1749-6632.2002.tb04924.x. [DOI] [PubMed] [Google Scholar]
- 179.Nachiappan V, Mufti SI, Eskelson CD. Ethanol-mediated promotion of oral carcinogenesis in hamsters: association with lipid peroxidation. Nutr Cancer. 1993;20:293–302. doi: 10.1080/01635589309514297. [DOI] [PubMed] [Google Scholar]
- 180.Tsutsumi M, George J, Ishizawa K, Fukumura A, Takase S. Effect of chronic dietary ethanol in the promotion of N-nitrosomethylbenzylamine-induced esophageal carcinogenesis in rats. J Gastroenterol Hepatol. 2006;21:805–813. doi: 10.1111/j.1440-1746.2005.04040.x. [DOI] [PubMed] [Google Scholar]
- 181.Aze Y, Toyoda K, Furukawa F, Mitsumori K, Takahashi M. Enhancing effect of ethanol on esophageal tumor development in rats by initiation of diethylnitrosamine. Carcinogenesis. 1993;14:37–40. doi: 10.1093/carcin/14.1.37. [DOI] [PubMed] [Google Scholar]
- 182.Elzay RP. Effect of alcohol and cigarette smoke as promoting agents in hamster pouch carcinogenesis. J Dent Res. 1969;48:1200–1205. doi: 10.1177/00220345690480061801. [DOI] [PubMed] [Google Scholar]
- 183.Elzay RP. Local effect of alcohol in combination with DMBA on hamster cheek pouch. J Dent Res. 1966;45:1788–1795. doi: 10.1177/00220345660450063401. [DOI] [PubMed] [Google Scholar]
- 184.Mufti SI, Becker G, Sipes IG. Effect of chronic dietary ethanol consumption on the initiation and promotion of chemically-induced esophageal carcinogenesis in experimental rats. Carcinogenesis. 1989;10:303–309. doi: 10.1093/carcin/10.2.303. [DOI] [PubMed] [Google Scholar]
- 185.Shikata N, Singh Y, Senzaki H, Shirai K, Watanabe T, Tsubura A. Effect of ethanol on esophageal cell proliferation and the development of N-methyl-N’-nitro-N-nitrosoguanidine induced-esophageal carcinoma in shrews. J Cancer Res Clin Oncol. 1996;122:613–618. doi: 10.1007/BF01221193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Kaneko M, Morimura K, Nishikawa T, Wanibuchi H, Osugi H, Kinoshita H, Koide A, Mori Y, Fukishima S. Weak enhancing effects of simultaneous ethanol administration on chemically induced rat esophageal tumorigenesis. Oncol Rep. 2002;9:1069–1073. [PubMed] [Google Scholar]
- 187.Morimura K, Hori T, Kaneko M, Nishikawa T, Nishikawa A, Wanibuchi H, Takada N, Osugi H, Fukushima S. Promotion of chemically induced rat esophageal tumorigenesis with post-initiation ethanol modification. Teratog Carcinog Mutagen. 2001;21:295–301. doi: 10.1002/tcm.1017. [DOI] [PubMed] [Google Scholar]
- 188.Gurski RR, Schirmer CC, Kruel CR, Komlos F, Kruel CD, Edelweiss MI. Induction of esophageal carcinogenesis by diethylnitrosamine and assessment of the promoting effect of ethanol and N-nitrosonornicotine: experimental model in mice. Dis Esophagus. 1999;12:99–105. doi: 10.1046/j.1442-2050.1999.00010.x. [DOI] [PubMed] [Google Scholar]
- 189.D’Souza El-Guindy NB, Kovacs EJ, De Witte P, Spies C, Littleton JM, de Villiers WJ, Lott AJ, Plackett TP, Lanzke N, Meadows GG. Laboratory models available to study alcohol-induced organ damage and immune variations: choosing the appropriate model. Alcohol Clin Exp Res. 2010;34:1489–1511. doi: 10.1111/j.1530-0277.2010.01234.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Roberts AJ, Heyser CJ, Koob GF. Operant self-administration of sweetened versus unsweetened ethanol: effects on blood alcohol levels. Alcohol Clin Exp Res. 1999;23:1151–1157. [PubMed] [Google Scholar]
- 191.Overstreet DH, Kampov-Polevoy AB, Rezvani AH, Murrelle L, Halikas JA, Janowsky DS. Saccharin intake predicts ethanol intake in genetically heterogeneous rats as well as different rat strains. Alcohol Clin Exp Res. 1993;17:366–369. doi: 10.1111/j.1530-0277.1993.tb00777.x. [DOI] [PubMed] [Google Scholar]
- 192.Yoneyama N, Crabbe JC, Ford MM, Murillo A, Finn DA. Voluntary ethanol consumption in 22 inbred mouse strains. Alcohol. 2008;42:149–160. doi: 10.1016/j.alcohol.2007.12.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Belknap JK, Crabbe JC, Young ER. Voluntary consumption of ethanol in 15 inbred mouse strains. Psychopharmacology. 1993;112:503–510. doi: 10.1007/BF02244901. [DOI] [PubMed] [Google Scholar]
- 194.Kampov-Polevoy AB, Garbutt JC, Janowsky DS. Association between preference for sweets and excessive alcohol intake: a review of animal and human studies. Alcohol Alcohol. 1999;34:386–395. doi: 10.1093/alcalc/34.3.386. [DOI] [PubMed] [Google Scholar]
- 195.Hubbell CL, Marglin SH, Spitalnic SJ, Abelson ML, Wild KD, Reid LD. Opioidergic, serotonergic, and dopaminergic manipulations and rats’ intake of a sweetened alcoholic beverage. Alcohol. 1991;8:355–367. doi: 10.1016/0741-8329(91)90573-f. [DOI] [PubMed] [Google Scholar]
- 196.Gilpin NW, Misra K, Koob GF. Neuropeptide Y in the central nucleus of the amygdala suppresses dependence-induced increases in alcohol drinking. Pharmacol Biochem Behav. 2008;90:475–480. doi: 10.1016/j.pbb.2008.04.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Tabakoff B, Hoffman PL. Animal models in alcohol research. Alcohol Res Health. 2000;24:77–84. [PMC free article] [PubMed] [Google Scholar]
- 198.Feith DJ, Pegg AE, Fong LY. Targeted expression of ornithine decarboxylase antizyme prevents upper aerodigestive tract carcinogenesis in p53-deficient mice. Carcinogenesis. 2013;34:570–576. doi: 10.1093/carcin/bgs377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Tan M, Myers JN, Agrawal N. Oral cavity and oropharyngeal squamous cell carcinoma genomics. Otolaryngol Clin North Am. 2013;46:545–566. doi: 10.1016/j.otc.2013.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Jeck WR, Parker J, Carson CC, Shields JM, Sambade MJ, Peters EC, Burd CE, Thomas NE, Chiang DY, Liu W, Eberhard DA, Ollila D, Grilley-Olson J, Moschos S, Neil Hayes D, Sharpless NE. Targeted next generation sequencing identifies clinically actionable mutations in patients with melanoma. Pigment Cell Melanoma Res. 2014;27:653–663. doi: 10.1111/pcmr.12238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Song Y, Li L, Ou Y, Gao Z, Li E, Li X, Zhang W, Wang J, Xu L, Zhou Y, Ma X, Liu L, Zhao Z, Huang X, Fan J, Dong L, Chen G, Ma L, Yang J, Chen L, He M, Li M, Zhuang X, Huang K, Qiu K, Yin G, Guo G, Feng Q, Chen P, Wu Z, Wu J, Zhao J, Luo L, Fu M, Xu B, Chen B, Li Y, Tong T, Wang M, Liu Z, Lin D, Zhang X, Yang H, Zhan Q. Identification of genomic alterations in oesophageal squamous cell cancer. Nature. 2014;509:91–95. doi: 10.1038/nature13176. [DOI] [PubMed] [Google Scholar]
- 202.Manna SK, Thompson MD, Gonzalez FJ. Application of mass spectrometry-based metabolomics in identification of early noninvasive biomarkers of alcohol-induced liver disease using mouse model. Adv Exp Med Biol. 2015;815:217–238. doi: 10.1007/978-3-319-09614-8_13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203.Shi X, Yao D, Chen C. Identification of N-acetyltaurine as a novel metabolite of ethanol through metabolomics-guided biochemical analysis. The Journal of biological chemistry. 2012;287:6336–6349. doi: 10.1074/jbc.M111.312199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Jabbour Z, do Nascimento C, Kotake BG, El-Hakim M, Henderson JE, de Albuquerque RF., Junior Assessing the oral microbiota of healthy and alcohol-treated rats using whole-genome DNA probes from human bacteria. Arch Oral Biol. 2013;58:317–323. doi: 10.1016/j.archoralbio.2012.07.017. [DOI] [PubMed] [Google Scholar]
- 205.Yu G, Gail MH, Shi J, Klepac-Ceraj V, Paster BJ, Dye BA, Wang GQ, Wei WQ, Fan JH, Qiao YL, Dawsey SM, Freedman ND, Abnet CC. Association between upper digestive tract microbiota and cancer-predisposing states in the esophagus and stomach. Cancer Epidemiol Biomarkers Prev. 2014;23:735–741. doi: 10.1158/1055-9965.EPI-13-0855. [DOI] [PMC free article] [PubMed] [Google Scholar]