

Summary
Trichinellosis is a helminthic infection where different species of T richinella nematodes are the causative agents. Several molecular assays have been designed to aid diagnostics of trichinellosis. These assays are mostly complex and expensive. The genomes of T richinella species contain certain parasite‐specific genes, which can be detected by polymerase chain reaction (PCR) methods. We selected β‐carbonic anhydrase (β‐ CA) gene as a target, because it is present in many parasites genomes but absent in vertebrates. We developed a novel β‐CA gene‐based method for detection of T richinella larvae in biological samples. We first identified a β‐CA protein sequence from T richinella spiralis by bioinformatic tools using β‐CAs from C aenorhabditis elegans and D rosophila melanogaster. Thereafter, 16 sets of designed primers were tested to detect β‐CA genomic sequences from three species of T richinella, including T . spiralis, T richinella pseudospiralis and T richinella nativa. Among all 16 sets of designed primers, the primer set No. 2 efficiently amplified β‐CA genomic sequences from T . spiralis, T . pseudospiralis and T . nativa without any false‐positive amplicons from other parasite samples including T oxoplasma gondii, T oxocara cati and P arascaris equorum. This robust and straightforward method could be useful for meat inspection in slaughterhouses, quality control by food authorities and medical laboratories.
Introduction
Trichinella spp. are the causative agents of a zoonotic helminthic disease, which is called trichinellosis (Gottstein et al., 2009). Within the main sources of Trichinella spp. [including pigs, horses, wild boars, dogs, walruses, foxes, bears and birds (Alban et al., 2011)], the most important source of human infection worldwide is the domestic pig, e.g. in Europe, whereas horse and wild boar meat have played a significant role during outbreaks within the past three decades (Gottstein et al., 2009). Infection in humans occurs with the ingestion of insufficiently cooked meat contaminated with Trichinella larvae, which are encysted in muscle tissues of the host animals (Clausen et al., 1996; Pozio, 2007; Gottstein et al., 2009). Due to the re‐emerging problem also in Europe, the European Union and some associated non‐European Union member countries still run Trichinella monitoring programmes (Gottstein et al., 2009). Following delivery by the gravid female worm, which lives within the intestinal mucosa of the host, newborn larvae (NBL) predominantly migrate into lymphatic and blood vessels of the host and thereafter to the ultimate target organ, i.e. highly oxygenated muscle. NBL enter the striated muscle cells by the aid of their stylet (Gottstein et al., 2009). NBL survive in muscle nurse cells for years (up to 40 years in humans and over 20 years, e.g. in polar bears) (Froscher et al., 1988; Kumar et al., 1990). Among 12 different genetically detected Trichinella spp., Trichinella spiralis is the most common species in domestic and wild swine and is also the most important aetiological agent that causes wide and global distribution of trichinellosis in humans (Pozio and Darwin Murrell, 2006). Other Trichinella spp. include Trichinella nativa (La Rosa et al., 2001; Nelson et al., 2003), Trichinella pseudospiralis (La Rosa et al., 2001; Pozio and Darwin Murrell, 2006), Trichinella britovi (Gottstein et al., 2009), Trichinella murelli (Ancelle, 1998), Trichinella nelsoni (Marucci et al., 2009), Trichinella papuae (Pozio et al., 2005), Trichinella zimbabwensis (Pozio et al., 2002), Trichinella genotype T8 (Marucci et al., 2009), Trichinella genotype T9 (Zarlenga et al., 2006) and Trichinella genotype T12 (Krivokapich et al., 2012). Seven percent of infected animals carry mixed infections (Airas et al., 2010).
Diagnosis of trichinellosis can be made by several methods. The classical method includes meat inspection and sampling for direct microscopic detection of larvae (Nockler et al., 2000). The other choices are detection of anti‐Trichinella immunoglobulin G (IgG) in animal serum or meat juice by serological methods including enzyme‐linked immunosorbent assay (Gamble et al., 2000; Korinkova et al., 2008), proteomics (Wang et al., 2014), and molecular methods including conventional polymerase chain reaction (PCR), real‐time PCR, loop‐mediated isothermal amplification (Tantrawatpan et al., 2012; Lin et al., 2013) and reverse line blot hybridization (Rombout et al., 2001). Many studies have been focused on detection of novel molecular markers, because the classical methods have limitations, such as labour‐intensiveness, necessity of high level expertise for direct microscopic diagnosis, and high costs due to the required manpower. The potential molecular markers have included Internal transcribed spacer (ITS1 and ITS2) (Zarlenga et al., 1999; Lin et al., 2013), mitochondrial cytochrome C oxidase subunit III (cox3) gene (Van De et al., 2012), mitochondrial large subunit ribosomal RNA (rrnL) (Borsuk et al., 2003; Guenther et al., 2008; Blaga et al., 2009; Li et al., 2011), mitochondrial small subunit ribosomal RNA (rrnS) (Blaga et al., 2009), DNA sequence of the 5S rRNA intergenic spacer regions (Rombout et al., 2001), migratory DNA from Trichinella larvae (Li et al., 2010) and aminopeptidase (TsAP) gene (Zhang et al., 2013).
One effective strategy for specific detection of Trichinella infection is identification of a genomic or proteomic determinant, which is absent in host genomes or proteomes (Duplessis and Moineau, 2001). Beta carbonic anhydrase (β‐CA) could be one of such host‐specific markers (Fasseas et al., 2010; Syrjanen et al., 2010; Zolfaghari Emameh et al., 2014; 2015). Six evolutionary divergent gene families encode for CAs (α, β, γ, δ, ζ and η) (Lane et al., 2005; Xu et al., 2008; Ferry, 2010; Del Prete et al., 2014; Zolfaghari Emameh et al., 2014). Different CA isozymes are involved in many biological processes, such as respiration involving transport of CO2 and bicarbonate between metabolizing tissues, pH homeostasis, electrolyte transfer, bone resorption, and calcification. Moreover, they participate in some biosynthetic reactions, such as gluconeogenesis, lipogenesis and ureagenesis in animals (Vullo et al., 2004; 2005; Alterio et al., 2006; Nishimori et al., 2007; Supuran, 2008), growth of some bacterial species (Merlin et al., 2003; Mitsuhashi et al., 2004) and photosynthesis in plants and algae (Fawcett et al., 1990; Funke et al., 1997). The previous investigations have confirmed that β‐CA gene sequences are completely absent in vertebrate genomes, whereas they can be found in plants, algae, yeasts, bacteria, archaea, protozoa and invertebrate metazoans (Fawcett et al., 1990; Funke et al., 1997; Smith et al., 2000; Merlin et al., 2003; Mitsuhashi et al., 2004; Aguilera et al., 2005; Fasseas et al., 2010; Syrjanen et al., 2010; Zolfaghari Emameh et al., 2014). By this background, we hypothesized that genomic β‐CA sequences from prokaryotic pathogens and eukaryotic parasites would be an ideal host‐specific determinant for molecular diagnostics, with minimal interfering effects from mammalian genomes.
In this study, we designed and piloted a PCR method for rapid detection of three species of Trichinella larvae, including T. spiralis, T. pseudospiralis and T. nativa in meat samples.
Results
Multiple sequence alignment (MSA)
MSA of the β‐CA protein sequence from T. spiralis (UniProt ID: E5SH53) against the sequences from Drosophila melanogaster (UniProt ID: Q9VHJ5) and C aenorhabditis elegans (UniProt ID: Q22460) revealed that they all contain the first (CxDxR; C: cystein, D: aspartic acid, R: arginine, and x: any residue) and second (HxxC; H: histidine, C: cystein, and x: any residue) highly conserved residues (Fig. 1). Hence, the β‐CA protein sequence from T. spiralis (UniProt ID: E5SH53) extracted correctly through the Basic Local Alignment Search Tool (BLAST) homology analysis and contained identical highly conserved residues to the predefined β‐CA protein sequences.
Figure 1.

Multiple sequence alignment (MSA) of β‐CA protein sequences from D . melanogaster (UniProt ID: Q9VHJ5), C . elegans (UniProt ID: Q22460) and T . spiralis (UniProt ID: E5SH53). The first (CxDxR) and second (HxxC) highly conserved residues are shown by arrows and marked with green colour.
Primer design and PCR
Among the four different pairs of primers (Nos. 1–4) only the primer set No. 2 (PCR product size 191 bp) showed a clear positive DNA band on agarose gel electrophoresis for detection of T. spiralis β‐CA genomic sequence in larvae and meat samples (data not shown). It also produced positive bands on larvae samples from T. pseudospiralis and T. nativa (Fig. 2). The next rounds of PCR analyses were performed to find out whether it was possible to design primers specific for T. pseudospiralis and T. nativa. We used the same reverse primer (primer set No. 2) which worked well for the first PCR round. The PCR products representing partial sequences of T. pseudospiralis and T. nativa β‐CA gene were sequenced, and the forward primers were designed based on the obtained sequences. These primers were included in the sets Nos. 5–7 for T. pseudospiralis and 8–11 for T. nativa, and the PCR results are shown in Fig. 3. Most primers produced multiple bands. When the most promising primers (No. 6 for T. pseudospiralis and No. 10 for T. nativa) were tested against all three species of Trichinella, they completely cross‐amplified because of the high sequence similarity (data not shown). Due to the interspecies cross‐amplification we further designed four new reverse primers which were used together with the previous forward primers resulting in primer set Nos. 12–16.
Figure 2.
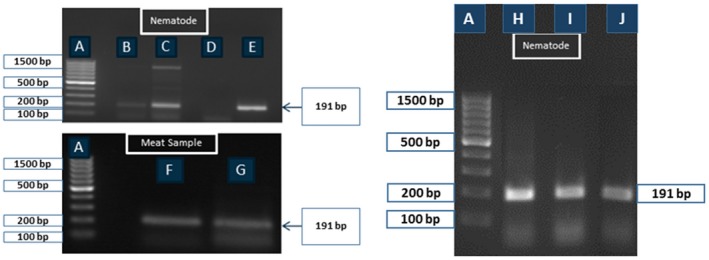
PCR‐based detection of genomic β‐CA sequence from T richinella larvae and infected meat samples. The primer set No. 2 showed a clear PCR product (size = 191 kb) in all T . spiralis‐positive samples. (A) 100 bp ladder; (B) one larva; (C) five larvae; (D) negative control (PCR reaction mixture without digested sample); (E) more than 10 larvae; (F and G) 5 mg of T . spiralis infected mouse meat samples; (H) T . nativa (10 larvae); (I) T . pseudospiralis (10 larvae); (J) T . spiralis (10 larvae).
Figure 3.
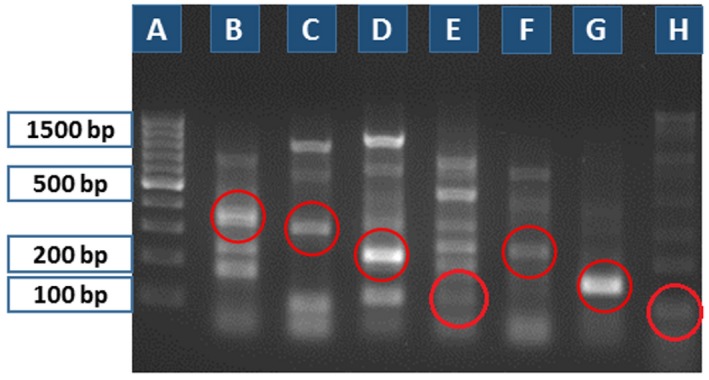
Second round of PCR‐based diagnosis tested on T . nativa and T . pseudospiralis samples (10 larvae/each lane). Primer set Nos. 5–7 for T . pseudospiralis and primer set Nos. 8–11 for T . nativa. (A) 100 bp ladder; (B) primer set No. 8 (372 bp), (C) primer set No. 9 (317 bp), (D) primer set No. 10 (226 bp), (E) primer set No. 11 (116 bp) for T . nativa; and (F) primer set No. 5 (251 bp), (G) primer set No. 6 (158 bp), (H) primer set No. 7 (103 bp) for T . pseudospiralis. The relevant PCR products are shown in red circles.
The results revealed that the primer set No. 13 from T. nativa and primer set No. 12 from T. pseudospiralis produced the strongest DNA bands on the agarose gel electrophoresis (Fig. 4). For evaluation of cross‐amplification these primers were tested on all three Trichinella spp. Figure 5 shows that there was again a complete cross‐amplification between different Trichinella spp. by these primers. This finding led us to conclude that finding of species‐specific primers for Trichinella β‐CA genes is probably impossible due to the high sequence similarity. Based on our results the primer set No. 2 was considered the most potential tool for diagnostic purposes, even though it was not able to discriminate between different Trichinella spp. Therefore, its specificity was further tested on prepared samples from Toxoplasma gondii, T oxocara cati, and P arascaris equorum. No false‐positive reactions were detectable (Fig. 6).
Figure 4.
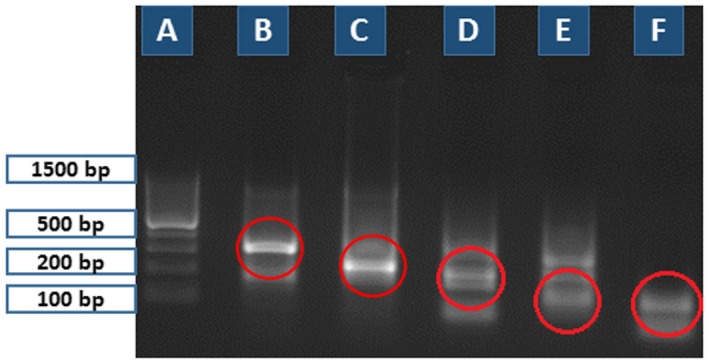
Third round of PCR tested on T . nativa and T . pseudospiralis samples (10 larvae/each lane). Primer set No. 12 for T . pseudospiralis and primer set Nos. 13–16 for T . nativa. (A) 100 bp ladder; (B) primer set No. 13 (319 bp), (C) primer set No. 14 (171 bp), (D) primer set No. 15 (168 bp), (E) primer set No. 16 (129 bp) for T . nativa, and (F) primer set No. 12 (116 bp) for T . pseudospiralis. The relevant PCR products are shown in red circles.
Figure 5.
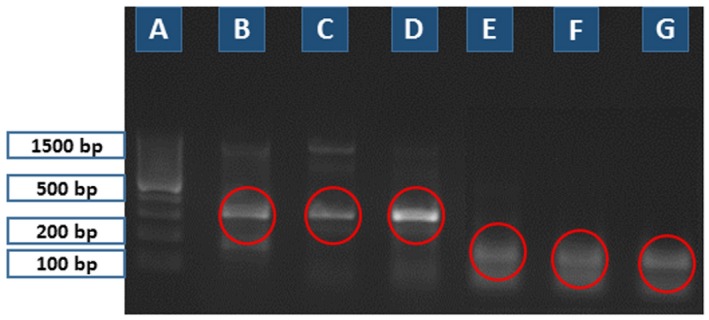
Evaluation of possible cross‐amplification between the primers designed for T . nativa and T . pseudospiralis β‐CA genes and templates from T. spiralis, T. nativa, and T . pseudospiralis (10 larvae/each lane). (A) 100 bp ladder; tested primer set No. 13 (product size 319 bp) on T . nativa (B), T . pseudospiralis (C), and T . spiralis (D); and primer set No. 12 (product size 116 bp) on T . nativa (E), T . pseudospiralis (F), and T . spiralis (G).
Figure 6.
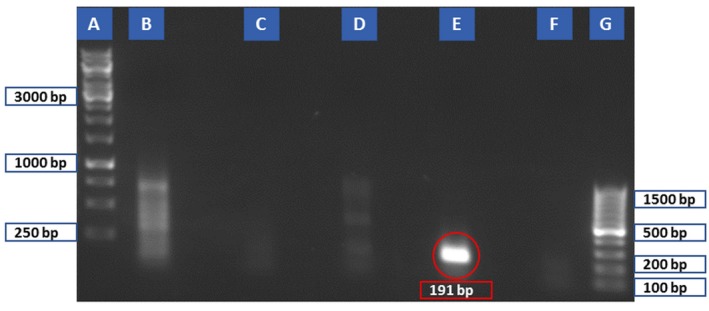
Evaluation of primer set No. 2 on different lysed parasites. (A) 1 kb ladder, (B) T . gondii, (C) T . cati, (D) P . equorum, (E) T . spiralis as the positive control, (F) negative control, and (G) 100 bp ladder. Red colour circle shows 191 bp PCR product of primer set No. 2 on T . spiralis.
Discussion
Many protozoan, arthropod, and nematode species (invertebrate metazoans) contain β‐CA gene and protein sequences in their genomes and proteomes respectively. BLAST homology analysis of predefined β‐CA protein sequences from D. melanogaster (Uniprot ID: Q9VHJ5) and C. elegans (Uniprot ID: Q22460) resulted in identification of more than 75 β‐CA protein sequences from protozoans and invertebrate metazoans (Zolfaghari Emameh et al., 2014). One of them was β‐CA protein sequence from T. spiralis (UniProt ID: E5SH53). Our previous results also revealed that β‐CA proteins show high similarity among various Drosophila species and other species of dipteran flies (Zolfaghari Emameh et al., 2014). This led us to hypothesize that the high similarity is extensible to other species of one defined genus, such as Trichinella spp. including the clinically most relevant species: T. spiralis, T. pseudospiralis, and T. nativa. In principle, the high sequence homology at gene level could provide a straightforward avenue to develop a genus‐specific PCR assay for diagnostics of Trichinella infection (trichinellosis). On the other hand, the high sequence similarity could be a difficult challenge for development of a species‐specific assay based on the β‐CA gene.
A number of PCR‐based diagnostic methods have been developed for detection of infectious diseases, and some of them have been applied for parasitic diseases, such as trichinellosis (Li et al., 2010). In our study, we designed and tested 16 different pairs of primers for detection of β‐CA genomic sequences of Trichinella spp. At the beginning of assay development, it is necessary to test the efficiency of designed primers. Our method using the primer set No. 2 seemed to detect minute amounts of DNA (a single larva/reaction). Thereafter, the set No. 2 was considered the most promising primer pair for the development of genus‐specific tool for Trichinella diagnostics. The results also revealed that interspecies cross‐amplification is a recurrent challenge in this assay system when the DNA sequences are highly similar. None of the tested primers were able to discriminate between different Trichinella spp. As already pointed out, the described method was no able to discriminate between various species of Trichinella. This same problem hampers most detection methods reported to date. All species and genotypes of the genus Trichinella are morphologically indistinguishable at all developmental stages. Consequently, only biochemical or molecular methods can be used to identify the genotype of the parasite (Gajadhar et al., 2009).
Our results showed that the developed assay is robust enough for screening of Trichinella infection in routine meat samples. The positive signals were genus‐specific since no false‐positive bands were detectable when samples from other parasites, a protozoan T. gondii and two nematodes T. cati and P. equorum, were used as templates. Importantly, no β‐CA genes are present in mammalian genomes (Syrjanen et al., 2010; Zolfaghari Emameh et al., 2014). Therefore, host‐derived false‐positive reactions would be unlikely. Other advantages of the present method include both ease of use and rapidity of detection. The meat samples are lysed overnight, and thereafter, the sample is ready for PCR amplification. No separate DNA isolation procedure is needed for the preparation of the template. The PCR reaction and gel electrophoresis take about 3 h after which the positive bands are clearly visible for documentation.
As an overall conclusion, even though there are many diagnostic methods available for trichinellosis, most of them are either expensive, time‐consuming or require special expertise. Therefore, new simple tools are urgently needed for screening of biological samples in livestock slaughterhouses and quality controlling laboratories of meat processing companies. Our developed method is based on detection of β‐CA genomic sequences from Trichinella spp. The greatest advantage of this method is that mammalian hosts do not contain any β‐CA gene sequences in their genomes. The results indicated that the PCR method is Trichinella genus‐specific and robust enough for routine screening purposes.
Experimental procedures
Identification of β‐CA protein sequence from T . spiralis
β‐CA protein sequence identification number (ID: E5SH53) of Trichinella spiralis in UniProt database (http://www.uniprot.org/) was retrieved by BLAST homology analysis using the previously defined β‐CA protein sequences from D. melanogaster (Uniprot ID: Q9VHJ5) and C. elegans (Uniprot ID: Q22460) (Pearson, 2014; Zolfaghari Emameh et al., 2014). The corresponding genomic sequence ID of T. spiralis (EFV55868) was obtained from Ensembl Metazoa (http://metazoa.ensembl.org/index.html). Moreover, MSA by Clustal Omega algorithm (Sievers et al., 2011) within EMBL‐EBI database (http://www.ebi.ac.uk/Tools/msa/clustalo/) was applied to approve the blast homology result. MSA revealed highly conserved sequences within β‐CA protein sequences from T. spiralis, D. melanogaster, and C. elegans.
Preparation of parasites and infected meat samples
Parasite samples and Trichinella spp.‐infected mouse muscle specimens were prepared at the Department of Veterinary Biosciences, University of Helsinki, Finland. The Animal Experiment Board in Finland had approved the study protocol. The tested Trichinella spp. involved T. spiralis, T. pseudospiralis and T. nativa. The blinded samples, including both infected and uninfected meat, were packed and shipped to Tissue Biology group, School of Medicine, University of Tampere, Finland, for PCR analysis. After arriving in the laboratory, the samples were transferred to −20°C freezer. To test the specificity of different primers, three more parasite samples were analysed including T. gondii (in culture medium), T. cati and P. equorum (in 70% ethanol).
Lysis of the parasites and meat samples
The meat and larvae samples were handled under the laminar hood. Toxoplasma gondii parasites were separated from the culture medium by centrifugation (Heraeus Biofuge Fresco, Thermo Scientific, Waltham, MA) at 13 000 RPM for 10 min (Leroux et al., 2015). Then the following lysis protocol was performed on all the samples: 300 μl of lysis buffer (DirectPCR Tail Lysis reagent, Viagen Biotech, LA) was added to 5 mg of samples. Then 6 μl of proteinase K (Thermo Scientific) was added to the mixture. The lysis tubes were incubated in rotating incubator (HYBAID, Thermo Scientific) at 55°C overnight.
Primer design
Three sets of primers for genomic sequence of β‐CA (exonic and intronic) from T. spiralis (Ensembl Metazoa ID: EFV55868) were designed automatically by NCBI Primer‐BLAST tool (http://www.ncbi.nlm.nih.gov/tools/primer‐blast/) (Ye et al., 2012), and one additional primer set was designed manually (Table 1). The expected sizes of the PCR products were calculated by Oligo Calc (http://www.basic.northwestern.edu/biotools/oligocalc.html) (Kibbe, 2007). To obtain genomic sequences for β‐CA of T. pseudospiralis and T. nativa we first performed PCR amplification using the designed T. spiralis primers, and T. pseudospiralis and T. nativa larvae were used as templates. The corresponding bands for T. pseudospiralis and T. nativa on the agarose gel were cut and sequenced as described in the sequencing section. The obtained β‐CA genomic sequences from T. pseudospiralis and T. nativa were aligned by Clustal Omega algorithm (http://www.ebi.ac.uk/Tools/msa/clustalo/) (Sievers et al., 2011) within EMBL‐EBI database (http://www.ebi.ac.uk/Tools/msa/clustalo/) against the genomic sequence of β‐CA from T. spiralis, and the most non‐identical regions were selected for the second round of primer design. We designed 3 and 4 different forward primers manually for T. pseudospiralis (primer set Nos. 5–7) and T. nativa (primer set Nos. 8–11), respectively, and used with the same reverse primer No. 2 which was described for the first round. In the third round of primer design, one new reverse primer was designed for T. pseudospiralis (primer set No. 12) and used with its own forward primer No. 7. Also, four new reverse primers were designed for T. nativa and tested with its own forward primers Nos. 8 and 10 (primer set Nos. 13–16). Three other species, T. gondii, T. cati, and P. equorum were tested for any false‐positive cross‐amplifications with other parasites.
Table 1.
Designed primers for β‐ CA genomic sequences from T . spiralis, T . pseudospiralis and T . nativa.
| No. | Designed primers | Species name | Product length (bp) |
|---|---|---|---|
| 1 |
Forward: 5′‐AGAGACTGCCCGTTCACAAG‐3′ Reverse: 3′‐CTGGGAGAGTTTGTCAGCGT‐5′ |
T. spiralis | 882 |
| 2 |
Forward: 5′‐TTTGAGCGCACTAGCATCCA‐3′ Reverse: 3′‐TCCATTCTGCATCACGCTGT‐5′ |
T. spiralis | 191 |
| 3 |
Forward: 5′‐AGACTGCCCGTTCACAAGTT‐3′ Reverse: 3′‐TGGATGCTAGTGCGCTCAAA‐5′ |
T. spiralis | 530 |
| 4 |
Forward: 5′‐ATTAATAAGTGAAAAGCACA‐3′ Reverse: 3′‐GTTTTAGAACTGGACACTGT‐5′ |
T. spiralis | 83 |
| 5 |
Forward: 5′‐CTCACCCATCACCCCGGCTT‐3′ Reverse: 3′‐TCCATTCTGCATCACGCTGT‐5′ |
T. pseudospiralis | 251 |
| 6 |
Forward: 5′‐GGCCAGGCCGTTCATCTGGT‐3′ Reverse: 3′‐TCCATTCTGCATCACGCTGT‐5′ |
T. pseudospiralis | 158 |
| 7 |
Forward: 5′‐ATCGCGCTCTCGCGATTGGG‐3′ Reverse: 3′‐TCCATTCTGCATCACGCTGT‐5′ |
T. pseudospiralis | 103 |
| 8 |
Forward: 5′‐TCCCAGACCAGCGGNAGCAC‐3′ Reverse: 3′‐TCCATTCTGCATCACGCTGT‐5′ |
T. nativa | 372 |
| 9 |
Forward: 5′‐GACCCAGCGCGCTTTCGTTG‐3′ Reverse: 3′‐TCCATTCTGCATCACGCTGT‐5′ |
T. nativa | 317 |
| 10 |
Forward: 5′‐CGGATACCACGGGCCGATGT‐3′ Reverse: 3′‐TCCATTCTGCATCACGCTGT‐5′ |
T. nativa | 226 |
| 11 |
Forward: 5′‐AGTCGCCCAGCTTGATCGCG‐3′ Reverse: 3′‐TCCATTCTGCATCACGCTGT‐5′ |
T. nativa | 116 |
| 12 |
Forward: 5′‐ATCGCGCTCTCGCGATTGGG‐3′ Reverse: 3′‐CAACCGATACCGAACGGACC‐5′ |
T. pseudospiralis | 116 |
| 13 |
Forward: 5′‐TCCCAGACCAGCGGNAGCAC‐3′ Reverse: 3′‐CGGACCGAACTCTGGTACAG‐5′ |
T. nativa | 319 |
| 14 |
Forward: 5′‐CGGATACCACGGGCCGATGT‐3′ Reverse: 3′‐CGGACCGAACTCTGGTACAG‐5′ |
T. nativa | 171 |
| 15 |
Forward: 5′‐TCCCAGACCAGCGGNAGCAC‐3′ Reverse: 3′‐ACATCGGCCCGTGGTATCCG‐5′ |
T. nativa | 168 |
| 16 |
Forward: 5′‐TCCCAGACCAGCGGNAGCAC‐3′ Reverse: 3′‐GTGAGTCCAGCAGCAACCCG‐5′ |
T. nativa | 129 |
PCR
PCR was performed based on the following reaction mixture: 12.5 μl of 2X KAPA ReadyMix (KAPA 2G Robust HotStart ReadyMix PCR Kit, Kapa Biosystems, Wilmington, MA), 1.25 μl of forward primer (Oligomer, Helsinki, Finland), 1.25 μl of reverse primer (Oligomer), 9 μl of dH2O and 1 μl of lysed samples. Lysed T. spiralis larvae served as a positive control and the negative control was the PCR master mix without any lysed sample. The PCR assay was run on the thermocycler (PTC100, MJ Research Inc, Waltham, MA) according to the following details: 95°C (3 min), [95°C (10 s), 53°C (10 s), 72°C (10 s)] × 36 cycles, 72°C (5 min).
Sequencing of β‐CA genomic sequences from T . pseudospiralis and T . nativa
We cut the PCR products of T. pseudospiralis and T. nativa templates from an agarose gel. Illustra‐GFX PCR DNA and Gel Band Purification Kit (GE Healthcare, Buckinghamshire, UK) was applied for extraction and purification of the corresponding DNA bands. At the final step, the overnight‐dried samples were suspended in HiDi (Life Technologies Europe, Applied Biosystems, Finland), which was followed by vortexing and spinning. The samples were heated at 95°C for 2 min and transferred to ice for 2 min. The samples were sent to Core Facilities and Research Services, BioMediTech, Tampere, Finland for DNA sequencing. The sequencing was carried out with HITACHI 3130x/Genetic Analyzer (Life Technologies Europe, Applied Biosystems).
Authors' contributions
RZE carried out blast homology analysis and MSA on T. spiralis, T. pseudospiralis, T. nativa, C. elegans, and D. melanogaster β‐CA sequences. RZE and MK participated in the primers design and setting up the PCR method. AN and AS prepared the parasite and infected meat samples. All authors participated in the design of the study. RZE and MK drafted the first version of the paper. All authors read and approved the final manuscript.
Conflict of Interest
None declared.
Funding Information To perform these studies RZE received a scholarship support from the Ministry of Science, Research and Technology, and National Institute of Genetic Engineering and Biotechnology of Islamic Republic of Iran. This study was also funded by Academy of Finland, Finnish Cultural Foundation (Pirkanmaa Regional Fund), Sigrid Juselius Foundation, Jane and Aatos Erkko Foundation, Tampere Tuberculosis Foundation, and Competitive Research Funding of the Tampere University Hospital.
References
- Aguilera, J. , Van Dijken, J.P. , De Winde, J.H. , and Pronk, J.T. (2005) Carbonic anhydrase (Nce103p): an essential biosynthetic enzyme for growth of Saccharomyces cerevisiae at atmospheric carbon dioxide pressure. Biochem J 391: 311–316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Airas, N. , Saari, S. , Mikkonen, T. , Virtala, A.M. , Pellikka, J. , Oksanen, A. , et al (2010) Sylvatic Trichinella spp. infection in Finland. J Parasitol 96: 67–76. [DOI] [PubMed] [Google Scholar]
- Alban, L. , Pozio, E. , Boes, J. , Boireau, P. , Boue, F. , Claes, M. , et al (2011) Towards a standardised surveillance for Trichinella in the European Union. Prev Vet Med 99: 148–160. [DOI] [PubMed] [Google Scholar]
- Alterio, V. , Vitale, R.M. , Monti, S.M. , Pedone, C. , Scozzafava, A. , Cecchi, A. , et al (2006) Carbonic anhydrase inhibitors: X‐ray and molecular modeling study for the interaction of a fluorescent antitumor sulfonamide with isozyme II and IX. J Am Chem Soc 128: 8329–8335. [DOI] [PubMed] [Google Scholar]
- Ancelle, T. (1998) History of trichinellosis outbreaks linked to horse meat consumption 1975–1998. Euro Surveill 3: 86–89. [DOI] [PubMed] [Google Scholar]
- Blaga, R. , Fu, B. , Le Rhun, D. , Le Naour, E. , Heckman, A. , Zocevic, A. , et al (2009) Use of mitochondrial RNA genes for the differentiation of four Trichinella species by multiplex PCR amplification. J Helminthol 83: 121–128. [DOI] [PubMed] [Google Scholar]
- Borsuk, P. , Moskwa, B. , Pastusiak, K. , and Cabaj, W. (2003) Molecular identification of Trichinella spiralis and Trichinella britovi by diagnostic multiprimer large mitochondrial rRNA amplification. Parasitol Res 91: 374–377. [DOI] [PubMed] [Google Scholar]
- Clausen, M.R. , Meyer, C.N. , Krantz, T. , Moser, C. , Gomme, G. , Kayser, L. , et al (1996) Trichinella infection and clinical disease. QJM 89: 631–636. [DOI] [PubMed] [Google Scholar]
- Del Prete, S. , Vullo, D. , Fisher, G.M. , Andrews, K.T. , Poulsen, S.A. , Capasso, C. , and Supuran, C.T. (2014) Discovery of a new family of carbonic anhydrases in the malaria pathogen Plasmodium falciparum – the eta‐carbonic anhydrases. Bioorg Med Chem Lett 24: 4389–4396. [DOI] [PubMed] [Google Scholar]
- Duplessis, M. , and Moineau, S. (2001) Identification of a genetic determinant responsible for host specificity in Streptococcus thermophilus bacteriophages. Mol Microbiol 41: 325–336. [DOI] [PubMed] [Google Scholar]
- Fasseas, M.K. , Tsikou, D. , Flemetakis, E. , and Katinakis, P. (2010) Molecular and biochemical analysis of the beta class carbonic anhydrases in Caenorhabditis elegans . Mol Biol Rep 37: 2941–2950. [DOI] [PubMed] [Google Scholar]
- Fawcett, T.W. , Browse, J.A. , Volokita, M. , and Bartlett, S.G. (1990) Spinach carbonic anhydrase primary structure deduced from the sequence of a cDNA clone. J Biol Chem 265: 5414–5417. [PubMed] [Google Scholar]
- Ferry, J.G. (2010) The gamma class of carbonic anhydrases. Biochim Biophys Acta 1804: 374–381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Froscher, W. , Gullotta, F. , Saathoff, M. , and Tackmann, W. (1988) Chronic trichinosis. Clinical, bioptic, serological and electromyographic observations. Eur Neurol 28: 221–226. [DOI] [PubMed] [Google Scholar]
- Funke, R.P. , Kovar, J.L. , and Weeks, D.P. (1997) Intracellular carbonic anhydrase is essential to photosynthesis in Chlamydomonas reinhardtii at atmospheric levels of CO2. Demonstration via genomic complementation of the high‐CO2‐requiring mutant ca‐1. Plant Physiol 114: 237–244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gajadhar, A.A. , Pozio, E. , Gamble, H.R. , Nockler, K. , Maddox‐Hyttel, C. , Forbes, L.B. , et al (2009) Trichinella diagnostics and control: mandatory and best practices for ensuring food safety. Vet Parasitol 159: 197–205. [DOI] [PubMed] [Google Scholar]
- Gamble, H.R. , Bessonov, A.S. , Cuperlovic, K. , Gajadhar, A.A. , van Knapen, F. , Noeckler, K. , et al (2000) International Commission on Trichinellosis: recommendations on methods for the control of Trichinella in domestic and wild animals intended for human consumption. Vet Parasitol 93: 393–408. [DOI] [PubMed] [Google Scholar]
- Gottstein, B. , Pozio, E. , and Nockler, K. (2009) Epidemiology, diagnosis, treatment, and control of trichinellosis. Clin Microbiol Rev 22: 127–145. Table of Contents. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guenther, S. , Nockler, K. , von Nickisch‐Rosenegk, M. , Landgraf, M. , Ewers, C. , Wieler, L.H. , and Schierack, P. (2008) Detection of Trichinella spiralis, T. britovi and T. pseudospiralis in muscle tissue with real‐time PCR. J Microbiol Methods 75: 287–292. [DOI] [PubMed] [Google Scholar]
- Kibbe, W.A. (2007) OligoCalc: an online oligonucleotide properties calculator. Nucleic Acids Res 35: W43–W46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Korinkova, K. , Kovarcik, K. , Pavlickova, Z. , Svoboda, M. , and Koudela, B. (2008) Serological detection of Trichinella spiralis in swine by ELISA (enzyme‐linked immunosorbent assay) using an excretory‐secretory (E/S) antigen. Parasitol Res 102: 1317–1320. [DOI] [PubMed] [Google Scholar]
- Krivokapich, S.J. , Pozio, E. , Gatti, G.M. , Prous, C.L. , Ribicich, M. , Marucci, G. , et al (2012) Trichinella patagoniensis n. sp. (Nematoda), a new encapsulated species infecting carnivorous mammals in South America. Int J Parasitol 42: 903–910. [DOI] [PubMed] [Google Scholar]
- Kumar, V. , Pozio, E. , de Borchgrave, J. , Mortelmans, J. , and De Meurichy, W. (1990) Characterization of a Trichinella isolate from polar bear. Ann Soc Belg Med Trop 70: 131–135. [PubMed] [Google Scholar]
- La Rosa, G. , Marucci, G. , Zarlenga, D.S. , and Pozio, E. (2001) Trichinella pseudospiralis populations of the Palearctic region and their relationship with populations of the Nearctic and Australian regions. Int J Parasitol 31: 297–305. [DOI] [PubMed] [Google Scholar]
- Lane, T.W. , Saito, M.A. , George, G.N. , Pickering, I.J. , Prince, R.C. , and Morel, F.M. (2005) Biochemistry: a cadmium enzyme from a marine diatom. Nature 435: 42. [DOI] [PubMed] [Google Scholar]
- Leroux, L.P. , Dasanayake, D. , Rommereim, L.M. , Fox, B.A. , Bzik, D.J. , Jardim, A. , and Dzierszinski, F.S. (2015) Secreted Toxoplasma gondii molecules interfere with expression of MHC‐II in interferon gamma‐activated macrophages. Int J Parasitol 45: 319–332. [DOI] [PubMed] [Google Scholar]
- Li, F. , Wang, Z.Q. , and Cui, J. (2010) Early detection by polymerase chain reaction of migratory Trichinella spiralis larvae in blood of experimentally infected mice. Foodborne Pathog Dis 7: 887–892. [DOI] [PubMed] [Google Scholar]
- Li, L.Z. , Wang, Z.Q. , Jiang, P. , Zhang, X. , Ren, H.J. , and Cui, J. (2011) Molecular identification of a Trichinella isolate from a naturally infected pig in Tibet, China. Korean J Parasitol 49: 381–384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin, Z. , Cao, J. , Zhang, H. , Zhou, Y. , Deng, M. , Li, G. , and Zhou, J. (2013) Comparison of three molecular detection methods for detection of Trichinella in infected pigs. Parasitol Res 112: 2087–2093. [DOI] [PubMed] [Google Scholar]
- Marucci, G. , La Grange, L.J. , La Rosa, G. , and Pozio, E. (2009) Trichinella nelsoni and Trichinella T8 mixed infection in a lion (Panthera leo) of the Kruger National Park (South Africa). Vet Parasitol 159: 225–228. [DOI] [PubMed] [Google Scholar]
- Merlin, C. , Masters, M. , McAteer, S. , and Coulson, A. (2003) Why is carbonic anhydrase essential to Escherichia coli? J Bacteriol 185: 6415–6424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mitsuhashi, S. , Ohnishi, J. , Hayashi, M. , and Ikeda, M. (2004) A gene homologous to beta‐type carbonic anhydrase is essential for the growth of Corynebacterium glutamicum under atmospheric conditions. Appl Microbiol Biotechnol 63: 592–601. [DOI] [PubMed] [Google Scholar]
- Nelson, M. , Wright, T.L. , Pierce, A. , and Krogwold, R.A. (2003) A common‐source outbreak of trichinosis from consumption of bear meat. J Environ Health 65: 16–19, 24. [PubMed] [Google Scholar]
- Nishimori, I. , Minakuchi, T. , Onishi, S. , Vullo, D. , Scozzafava, A. , and Supuran, C.T. (2007) Carbonic anhydrase inhibitors. DNA cloning, characterization, and inhibition studies of the human secretory isoform VI, a new target for sulfonamide and sulfamate inhibitors. J Med Chem 50: 381–388. [DOI] [PubMed] [Google Scholar]
- Nockler, K. , Pozio, E. , Voigt, W.P. , and Heidrich, J. (2000) Detection of Trichinella infection in food animals. Vet Parasitol 93: 335–350. [DOI] [PubMed] [Google Scholar]
- Pearson, W.R. (2014) BLAST and FASTA similarity searching for multiple sequence alignment. Methods Mol Biol 1079: 75–101. [DOI] [PubMed] [Google Scholar]
- Pozio, E. (2007) World distribution of Trichinella spp. infections in animals and humans. Vet Parasitol 149: 3–21. [DOI] [PubMed] [Google Scholar]
- Pozio, E. , and Darwin Murrell, K. (2006) Systematics and epidemiology of Trichinella . Adv Parasitol 63: 367–439. [DOI] [PubMed] [Google Scholar]
- Pozio, E. , Foggin, C.M. , Marucci, G. , La Rosa, G. , Sacchi, L. , Corona, S. , et al (2002) Trichinella zimbabwensis n.sp. (Nematoda), a new non‐encapsulated species from crocodiles (Crocodylus niloticus) in Zimbabwe also infecting mammals. Int J Parasitol 32: 1787–1799. [DOI] [PubMed] [Google Scholar]
- Pozio, E. , Owen, I.L. , Marucci, G. , and La Rosa, G. (2005) Inappropriate feeding practice favors the transmission of Trichinella papuae from wild pigs to saltwater crocodiles in Papua New Guinea. Vet Parasitol 127: 245–251. [DOI] [PubMed] [Google Scholar]
- Rombout, Y.B. , Bosch, S. , and Van Der Giessen, J.W. (2001) Detection and identification of eight Trichinella genotypes by reverse line blot hybridization. J Clin Microbiol 39: 642–646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sievers, F. , Wilm, A. , Dineen, D. , Gibson, T.J. , Karplus, K. , Li, W. , et al (2011) Fast, scalable generation of high‐quality protein multiple sequence alignments using Clustal Omega. Mol Syst Biol 7: 539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith, K.S. , Cosper, N.J. , Stalhandske, C. , Scott, R.A. , and Ferry, J.G. (2000) Structural and kinetic characterization of an archaeal beta‐class carbonic anhydrase. J Bacteriol 182: 6605–6613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Supuran, C.T. (2008) Carbonic anhydrases: novel therapeutic applications for inhibitors and activators. Nat Rev Drug Discov 7: 168–181. [DOI] [PubMed] [Google Scholar]
- Syrjanen, L. , Tolvanen, M. , Hilvo, M. , Olatubosun, A. , Innocenti, A. , Scozzafava, A. , et al (2010) Characterization of the first beta‐class carbonic anhydrase from an arthropod (Drosophila melanogaster) and phylogenetic analysis of beta‐class carbonic anhydrases in invertebrates. BMC Biochem 11: 28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tantrawatpan, C. , Intapan, P.M. , Thanchomnang, T. , Lulitanond, V. , Boonmars, T. , Wu, Z. , et al (2012) Differential detection of Trichinella papuae, T. spiralis and T. pseudospiralis by real‐time fluorescence resonance energy transfer PCR and melting curve analysis. Vet Parasitol 185: 210–215. [DOI] [PubMed] [Google Scholar]
- Van De, N. , Trung, N.V. , Ha, N.H. , Nga, V.T. , Ha, N.M. , Thuy, P.T. , et al (2012) An outbreak of trichinosis with molecular identification of Trichinella sp. in Vietnam. Korean J Parasitol 50: 339–343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vullo, D. , Franchi, M. , Gallori, E. , Antel, J. , Scozzafava, A. , and Supuran, C.T. (2004) Carbonic anhydrase inhibitors. Inhibition of mitochondrial isozyme V with aromatic and heterocyclic sulfonamides. J Med Chem 47: 1272–1279. [DOI] [PubMed] [Google Scholar]
- Vullo, D. , Innocenti, A. , Nishimori, I. , Pastorek, J. , Scozzafava, A. , Pastorekova, S. , and Supuran, C.T. (2005) Carbonic anhydrase inhibitors. Inhibition of the transmembrane isozyme XII with sulfonamides – a new target for the design of antitumor and antiglaucoma drugs? Bioorg Med Chem Lett 15: 963–969. [DOI] [PubMed] [Google Scholar]
- Wang, L. , Cui, J. , Hu, D.D. , Liu, R.D. , and Wang, Z.Q. (2014) Identification of early diagnostic antigens from major excretory‐secretory proteins of Trichinella spiralis muscle larvae using immunoproteomics. Parasit Vectors 7: 40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu, Y. , Feng, L. , Jeffrey, P.D. , Shi, Y. , and Morel, F.M. (2008) Structure and metal exchange in the cadmium carbonic anhydrase of marine diatoms. Nature 452: 56–61. [DOI] [PubMed] [Google Scholar]
- Ye, J. , Coulouris, G. , Zaretskaya, I. , Cutcutache, I. , Rozen, S. , and Madden, T.L. (2012) Primer‐BLAST: a tool to design target‐specific primers for polymerase chain reaction. BMC Bioinformatics 13: 134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zarlenga, D.S. , Chute, M.B. , Martin, A. , and Kapel, C.M. (1999) A multiplex PCR for unequivocal differentiation of all encapsulated and non‐encapsulated genotypes of Trichinella . Int J Parasitol 29: 1859–1867. [DOI] [PubMed] [Google Scholar]
- Zarlenga, D.S. , Rosenthal, B.M. , La Rosa, G. , Pozio, E. , and Hoberg, E.P. (2006) Post‐Miocene expansion, colonization, and host switching drove speciation among extant nematodes of the archaic genus Trichinella . Proc Natl Acad Sci USA 103: 7354–7359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, Y. , Wang, Z. , Li, L. , and Cui, J. (2013) Molecular characterization of Trichinella spiralis aminopeptidase and its potential as a novel vaccine candidate antigen against trichinellosis in BALB/c mice. Parasit Vectors 6: 246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zolfaghari Emameh, R. , Barker, H. , Tolvanen, M.E. , Ortutay, C. , and Parkkila, S. (2014) Bioinformatic analysis of beta carbonic anhydrase sequences from protozoans and metazoans. Parasit Vectors 7: 38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zolfaghari Emameh, R. , Syrjanen, L. , Barker, H. , Supuran, C.T. , and Parkkila, S. (2015) Drosophila melanogaster: a model organism for controlling Dipteran vectors and pests. J Enzyme Inhib Med Chem 30: 505–513. [DOI] [PubMed] [Google Scholar]