

Abstract
Posttranscriptional mechanisms have a critical role in the overall outcome of gene expression. These mechanisms are especially relevant in protozoa from the genus Trypanosoma, which is composed by death threatening parasites affecting people in Sub-saharan Africa or in the Americas. In these parasites the classic view of regulation of transcription initiation to modulate the products of a given gene cannot be applied. This is due to the presence of transcription start sites that give rise to long polycistronic units that need to be processed costranscriptionally by trans-splicing and polyadenylation to give mature monocistronic mRNAs. Posttranscriptional mechanisms such as mRNA degradation and translational repression are responsible for the final synthesis of the required protein products. In this context, RNA-binding proteins (RBPs) in trypanosomes have a relevant role as modulators of mRNA abundance and translational repression by associating to the 3’ untranslated regions in mRNA. Many different RBPs have been proposed to modulate cohorts of mRNAs in trypanosomes. However, the current understanding of their functions lacks a dynamic view on the different steps at which these RBPs are regulated. Here, we discuss different evidences to propose regulatory events for different RBPs in these parasites. These events vary from regulated developmental expression, to biogenesis of cytoplasmic ribonucleoprotein complexes in the nucleus, and condensation of RBPs and mRNA into large cytoplasmic granules. Finally, we discuss how newly identified posttranslational modifications of RBPs and mRNA metabolism-related proteins could have an enormous impact on the modulation of mRNA abundance. To understand these modifications is especially relevant in these parasites due to the fact that the enzymes involved could be interesting targets for drug therapy.
Keywords: Trypanosoma, Posttranscriptional gene expression, Ribonucleoprotein complexes, RNA-binding protein, Developmental regulation, Sleeping sickness, Posttranslational modification, Phosphorylation, Chagas disease
Core tip: We discuss several ways to regulate the function of RNA-binding proteins in trypanosomes. We highlight the propensity of these proteins to engage in interactions with other proteins and RNA, resulting in the formation of large reversible aggregates induced by environmental stress. Finally, the possible role of posttranslational modifications on the function of these proteins is discussed in the context of recent high-throughput proteomic evidences.
INTRODUCTION
The synthesis of the required protein products, and their levels, demands coordinated mechanisms to regulate gene expression in all organisms. These mechanisms operate in eukaryotic cells at the level of transcription initiation and/or mRNA processing, localization, stability or translational efficiency. As an exception to the rule, a group of protozoan parasites belonging to the genus Trypanosoma have evolved towards regulating gene expression without classical transcription initiation control. Within these parasites are T. brucei, the causative agent of Sleeping Sickness in Sub-Saharan Africa[1], and T. cruzi, causing Chagas Disease in the Americas[2]. These parasites have complex life cycles, which involve an hematophagous insect vector and a vertebrate host, each one hosting different life forms with unique characteristics and metabolisms[3]. Unfortunately, there are no effective vaccines against these parasites, and current drug treatments are highly toxic, present low tolerance and require long patient compliance[4]. Although the current drug treatments can be effective during the acute stage of the infection, there is a need for newer, safer and more effective treatments against these neglected diseases.
One peculiarity of these parasites is that their genomes have protein-coding genes organized into large directional gene clusters[5,6], which lack canonical RNA polymerase II promoters. Instead, histone variants and epigenetic regulation seem to mark the boundaries of yet undefined transcription start sites for these long transcriptional units[7,8]. The resultant polycistronic immature mRNAs are then processed by coordinated trans-splicing and polyadenylation events, giving rise to mature monocistronic mRNAs[9]. However, protein-coding sequences in each trypanosomatid transcriptional unit are not functionally related as they are in bacterial operons[5]. From this point onwards, posttranscriptional regulation of gene expression heavily contributes to the levels of the protein products that are synthesized, depending on the cell’s requirements. The expression of functionally related mRNAs seems to be controlled in a coordinated fashion as posttranscriptional regulons[10]. Within this view, structural motifs enriched in 3’ untranslated regions seem to govern the fate of the mRNA molecule inside the cell. Different transcripts carrying the same signature motifs are likely to be regulated in a similar way[11]. These structural motifs serve as ligands for different RNA-binding proteins (RBPs) that associate with the mRNA, forming a ribonucleoprotein (RNP) complex, which is also composed by many other proteins that interact with different parts of the transcript. Consequently, the presence or absence of different RBPs, which in turn can recruit other factors to the RNP complex, likely dictates the fate of the mRNA inside the cell. RBPs interact with mRNA motifs using dedicated functional domains such as the RNA-Recognition Motif (RRM), the Zinc Finger domains, the Pumilio domain and the ALBA domain, to name the most relevant domains found in trypanosomatid RBPs[12].
Trypanosomes have been the muse for the identification of novel biological mechanisms such as RNA trans-splicing, mitochondrial RNA editing, and antigenic variation[13], allowing us to understand their similarities and differences with higher eukaryotic organisms. Identification and characterization of RBPs in trypanosomes started almost 20 years ago. However, it was the concerted sequencing and annotation of their genomes 10 years ago that handled the field with the tools to characterize factors and deeply describe these mechanisms in trypanosomes[14,15]. While T. cruzi lacks some the components required for a functional RNA interference (RNAi) machinery, in T. brucei mRNA levels can be easily downregulated by this mechanism[16], making this organism the workhorse in the field. The usefulness of omics-approaches in trypanosomes has also allowed the identification of the mRNA molecules affected when a certain RBP is down or upregulated[17]. This, added to the current ability to identify the associated mRNA molecules of a given RBP by RNA-crosslinking and sequencing technologies[18], can give a deeper understanding of the role of the RBP under study. However, our current understanding of RBP function in trypanosomes is rather static, and is far from reflecting the dramatic changes in posttranscriptional gene expression these protozoa suffer during their life cycles. In their mammalian and insect hosts, these parasites adopt different life forms that are adapted to environmental changing conditions. Thus, any single factor modulating gene expression in trypanosomes needs to be regulated in order to cope with the changing developmental and environmental conditions they face.
Here, we will expose the yet poorly explored regulation of the function of trypanosomatid RBPs, emphasizing on those involved in the modulation of cytoplasmic mRNA levels and translation. Regulation is probably exerted at different levels: RBP expression for a given developmental stage, localization-mediated function including nucleocytoplasmic shuttling and nuclear RNA-binding, condensation and sequestering into RNP complexes, and posttranslational modifications (PTMs). We aim to analyze the possible additional layers of complexity that RBPs bring to the gene expression game in trypanosomes.
Developmental regulation of RBP expression
The simplest way to regulate the function of a protein in a time manner is to prevent or promote its synthesis. This is especially relevant for trypanosomes, which alternate between different life forms in their insect and mammalian hosts during which gene expression seems to be regulated essentially at the posttranscriptional level. In these parasites, certain developmental programs were confirmed to be orchestrated by RBPs[12]. Originally, the developmentally regulated expression of each RBP was detected using specific antibodies (Table 1), although current high-throughput approaches rely on the analysis of the transcriptome and the proteome[19-22]. While the latter approaches can give an extensive amount of information on the regulated abundance of mRNAs and proteins, including RBPs, biochemical or genetic validation for each RBP is required to confirm developmental regulation. The first reported case of developmentally regulated RBPs in T. brucei was from P34/P37[23]. These are highly similar RRM-containing RBPs that are required for the assembly of the 60S ribosomal subunit[24], being P34 expressed in procyclics and P37 expressed in bloodstream forms[23]. In T. cruzi, the RRM-containing RBP UBP2 was the first to show developmental regulation, being expressed in replicative epimastigotes and amastigotes, while it is almost undetectable in cell-derived trypomastigotes[25]. It remains to be determined if overexpression or the lack of expression of P34/P37, UBP2 or other proteins listed in Table 1, can compromise the developmental program of a trypanosome.
Table 1.
Developmentally expressed RNA-binding proteins in trypanosomes confirmed by experimental data
| RBP | Stage | Ref. |
| T. brucei | ||
| P34 | Procyclics | [23] |
| P37 | Bloodstream | [23] |
| ZFP1 | Procyclics | [26] |
| ZC3H20 | Procyclics | [78] |
| Alba 3 | Tsetse fly except transition stages in proventriculus | [79] |
| Alba 4 | Tsetse fly except transition stages in proventriculus | [79] |
| RBP6 | Metacyclics, epimastigotes | [29] |
| RBP10 | Bloodstream | [28] |
| hnRNPF/H | Mainly bloodstream | [80] |
| T. cruzi | ||
| UBP2 | Mostly epimastigotes, low in amastigotes | [25] |
| RBP3 | Epimastigotes | [81] |
| RBP4 | Epimastigotes | [81] |
| RBP19 | Amastigotes | [82] |
| ZFP2 | Downregulated in metacyclic trypomastigotes | [83] |
RBP: RNA-binding protein.
Hendriks et al[26] demonstrated that genetic ablation of ZFP1 in T. brucei prevented the correct repositioning of the kinetoplast, a specific configuration for the mitochondrial DNA present in trypanosomes and leishmanias[27]. This phenotype was associated with the incomplete morphological differentiation from bloodstream to procyclic form T. brucei parasites[26], suggesting a role for ZFP1 in T. brucei bloodstream form differentiation. RBP10, which is another RRM-containing protein, showed to be enriched in bloodstream-form T. brucei cells[28]. Knock-down of RBP10 by RNAi in bloodstream forms resulted in the downregulation of transcripts with high abundance in this stage, while overexpression of RBP10 in procyclics revealed an increased abundance of bloodstream specific transcripts[28]. Hence, manipulation of RBP10 protein levels seems to modulate the abundance of specific developmentally regulated transcripts, although there was no commitment to a differentiation event. RBP6-mediated developmental differentiation in T. brucei is probably the most relevant finding regarding developmental expression[29] (Figure 1A). A genome wide high-throughput RNA-seq analysis revealed RBP6 transcripts in T. brucei parasites from the proventriculous were enriched 13-times relatively to procyclics from the midgut. When Kolev et al[29] overexpressed RBP6 in cultured procyclic cells, they observed developmental stages that are normally observed in the insect vector and not in culture, namely long and short epimastigotes and metacyclic trypomastigotes (Figure 1A). RBP6-overexpressing parasites in culture showed the hallmarks of undergoing metacyclogenesis, and could be used to infect mice, showing the full potential of an infective form. All these examples reveal the role of developmentally regulated RBPs in the modulation of the abundance of life form-specific transcripts, pointing to these and other RBPs as master factors regulating developmental programs.
Figure 1.
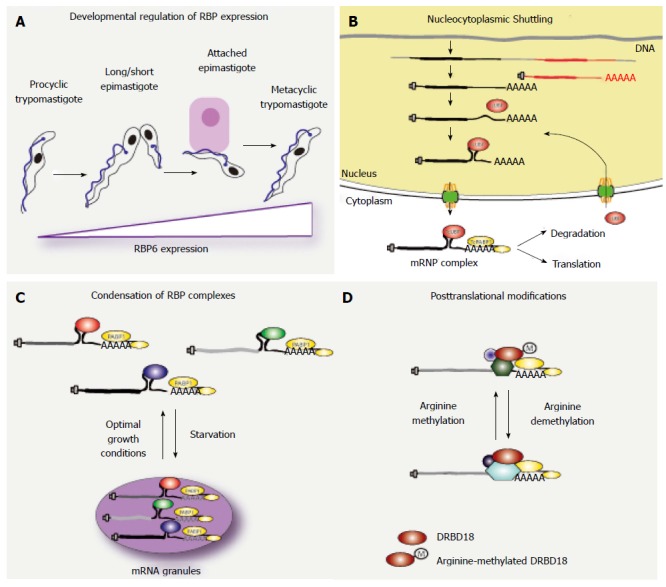
Regulation of RNA-binding proteins function in trypanosomes. Each panel summarizes the proposed mode of regulation of the function of RNA-binding proteins in trypanosomes.
Regulation of RBP localization
A potential way to regulate the function of an RBP is to limit where in the cell it can associate to its RNA target. Although we are beginning to understand the complexity of RBP-mediated posttranscriptional regulation of gene expression, current models often describe static events, where the RBP is functional once it is associated with the RNA. For many RBPs directly involved in nuclear RNA metabolism, or for those showing high sequence identity to characterized factors involved in conserved nuclear mechanisms, a nuclear localization would make sense for the expected function. This has been the case for the LA protein[30], the exoribonuclease XRNE involved in pre-rRNA processing[31], the nuclear cap binding protein CBP20[32], the Splice-Leader RNP complex associated Sm proteins[33], or Lsm proteins[34], to name a few. Notwithstanding this, nuclear localization for other functionally uncharacterized RBPs like ZFP8 in T. cruzi requires further understanding of the function of the protein to assign it to a nuclear function[35]. This was the case of the nuclear protein RBP33 in T. Brucei, which was suggested to be involved in the regulation of nuclear gene silencing[36]. Nuclear localization signals in trypanosomes are difficult to detect because they are not usually predicted by the algorithms developed for mammalian proteins[30,37], adding another layer of complexity to the characterization of RBPs targeted to the nucleus.
For most models of cytoplasmic RBP function there is no information on where and when the RNP complex is formed. This is especially relevant in posttranscriptional regulation, which is believed to happen mostly in the cytoplasm, but has a main actor (the mRNA) with a nuclear history. In T. brucei bloodstream forms, most mRNA half-lives are under 20 min, with few long lived mRNAs with half lives of more than 2 h[17]. In Saccharomyces cerevisiae, expression dynamics of RBPs showed an increased protein abundance and higher half-life as compared to non-RBP proteins[38]. If this is also the case for trypanosomes, it could suggest that a single RBP molecule might complete more than one cycle of RNP complex biogenesis and degradation. In mammalian cells, RNP complex remodeling and recycling of RBPs has been studied from a nuclear perspective, with many factors associating to mRNA in the nucleus, travelling together through the nuclear pore complex and returning to the nucleus for another event of RNP biogenesis[39]. It is clear that analyzing a snapshot for the localization of an RBP will tell where the molecule spends most of its time, but will fail to determine whether it spends time in other cellular locations albeit transiently. The evidence of RBP nucleocytoplasmic shuttling in trypanosomes points to a possible nuclear step for proper cytoplasmic function[40]. This hypothesis arose from the behavior of six single RRM-containing RBPs (UBP1-2, RBP3-6) in T. cruzi[37]. In this case, these cytoplasmic proteins reversibly accumulated in the nucleus when cells were stressed with sodium arsenite. Although this is not a physiological stressor, its use allowed the nuclear accumulation of these RBPs, events that would have been undetected under normal conditions[40]. Active transcription and an intact RRM were required for UBP1 nuclear accumulation, suggesting dependence on nuclear availability of newly synthesized mRNAs for this to happen[37]. Colocalization of UBP1 with target transcripts and poly (A) mRNA in the nucleus suggests that arsenite could be inhibiting nuclear export of mRNA. In the same line of evidence, the cytoplasmic DRBD3 in T. brucei showed a similar response to arsenite stress[41], showing that similar events of nucleocytoplasmic shuttling could be affected by arsenite in both species. Also in T. brucei, PABP2 could accumulate in the nucleus under a combined stress of heat shock and sinefungin addition[42], a drug that prevents trans-splicing of mRNA by inhibiting Cap4 formation of Splice leader molecules[43]. All these evidences point towards a nuclear mRNA-binding event taking place during mRNP biogenesis (Figure 1B). It remains to be determined if this nuclear event is required for the function of these RBPs in the cytoplasm.
Regulation of RBP interactions and condensation in RNP complexes
In the cytoplasm of trypanosomatid cells mRNA molecules are associated with general and specific RBPs. It is believed that these specific RBPs will dictate the fate of the mRNA[44]. Thus, mRNA molecules and RBPs are distributed in RNP complexes throughout the cytoplasm. However, these RNP complexes can be very different in protein composition from each other, with proteins colocalizing in some foci but not in every one of them[45].
Under starvation conditions in T. cruzi and T. brucei, large cytoplasmic granules are formed as a result of the condensation of different RNP complexes[46]. Several evidences suggest that these are not genuine Stress Granules, and consequently were named mRNA granules, since these were characterized based on the dynamics of the mRNA[46]. The list of proteins colocalizing with mRNA granules is in expansion (Table 2), since these structures have been a valuable tool to involve a putative RBP in mRNA metabolism in trypanosomes. No ribosomal proteins from the 40S or 60S subunits were detected in mRNA granules in T. cruzi[46], suggesting that the transcripts accumulating in these structures have already exited translation. In Saccharomyces cerevisiae, metabolic enzymes also reorganize from a homogenous distribution in the cytoplasm to reversible granule-like assemblies under starvation[47]. In this work it was proposed that the recruitment of enzymes to cytoplasmic foci during the nutrient deprivation period could function as a reservoir of critical cellular components for cell-cycle reentry once nutrients are readily available. This would empty the cytoplasm of these proteins, and would prevent their function due to condensation in granules. This hypothesis is especially relevant in trypanosomes for proteins with enzymatic activity like the exoribonuclease XRNA, which is involved in 5’ to 3’ enzymatic digestion of highly unstable and developmentally regulated mRNA[48] (Figure 1C). XRNA is in fact a component of starvation mRNA granules, where it does not seem to be promoting mRNA degradation. In fact, the transcripts stored in mRNA granules in trypanosomes are intact, since these do possess a poly(A) tail and the characteristic mini-exon at the 5’ end[46,49]. This, added to the fact that transcripts in granules can reenter translation if a new source of nutrients is available[50], suggests that these structures can play a role as reservoirs of transcripts during a physiological stress like starvation. This setup would provide fast templates for synthesis of cellular components that would allow differentiation or cell-cycle re-entry.
Table 2.
List of components identified in starvation mRNA granules in trypanosomes
| Component | Organism |
| poly(A) | T. brucei, T. cruzi |
| UBP1 | T. brucei, T. cruzi |
| UBP2 | T. cruzi |
| RBP3 | T. cruzi |
| RBP4 | T. cruzi |
| RBP5 | T. cruzi |
| RBP6b | T. cruzi |
| PABP1 | T. brucei, T. cruzi |
| PABP2 | T. brucei, T. cruzi |
| DRBD3 | T. brucei |
| DRBD4 | T. brucei |
| ALBA1 | T. brucei |
| ALBA2 | T. brucei |
| ALBA3 | T. brucei |
| ALBA4 | T. brucei |
| ZFP3 | T. brucei |
| DHH1 | T. brucei, T. cruzi |
| XRNA | T. brucei, T. cruzi |
| SCD6 | T. brucei |
| PBP1 | T. brucei |
| MKT1 | T. brucei |
| CAF1 | T. brucei |
| NOT1 | T. brucei |
| NOT5 | T. brucei |
| UPF1 | T. brucei |
| DBP1 | T. brucei |
| AGO1 | T. brucei |
| eIF4G1 | T. brucei |
| eIF4G4 | T. brucei |
| Serine-threonine kinase receptor-associated protein | T. brucei |
| mRNA cap guanine-N7 methyltransferase | T. brucei |
| Phosphatase | T. brucei |
| Methyltransferase | T. brucei |
Current evidence failed to identify a single RBP required for the formation of mRNA granules, although the overexpression of SCD6 is sufficient to induce the formation of aggregates similar to starvation granules, which contain mRNA, DHH1 and eIF4E[49]. However, depletion of SCD6 by RNAi in T. brucei did not affect the formation of starvation mRNA granules[51], suggesting that redundant factors might be involved in the formation of these structures. Of the experimentally validated components of these structures, many contain RNA-binding domains (RBD), such as the RRM or ALBA domain, or contain domains involved in interactions with other mRNA metabolism-related proteins (Table 2). Very recently, Fritz et al[52] developed a very ingenious approach to purify starvation induced mRNA granules that is based on the release of these structures from the subpellicular microtubule cage after cytoskeleton depolymerization, followed by mass spectrometry (MS)[52]. By this approach, they could detect many proteins already identified in mRNA granules, and many new components, 17 of which could be efficiently validated by eYFP-fusion protein colocalization to mRNA granules markers. These included RNA metabolism related proteins, as well as one phosphatase, one methyltransferase, and four hypothetical proteins (Table 2).
Other domains accompanying RBDs in RBPs can provide a scaffold for protein interactions and additional regulation[53]. This is the case of low complexity (LC) sequences, defined by low information content due to the repetition of a few amino acids. LC sequences enriched in RBPs and mRNA metabolism related proteins resemble prion-like domains, allowing these proteins to self-assemble. For instance, the yeast Lsm4 protein, a core P-body component, appears to be involved in granule formation by self-aggregation of its Q/N rich domain[54]. Reijns et al[55] identified 20 yeast P-body proteins with above average Q/N content, and demonstrated a reduced association to these structures when these sequences were removed from Ccr4p, Pop2p and Dhh1p. In trypanosomes, proteins like UBP1, UBP2, RBP3 and RBP6, which contain Q-rich sequences, are found in starvation mRNA granules[46]. In spite of this, the presence of a functional RRM is necessary and sufficient for the localization of UBP1 to these structures, showing a direct association to transcripts in starvation mRNA granules for this RBP[46]. Q or G-rich sequences were also identified in many other RRM-type RBPs[56], and a poly-Q sequence has been proposed to be a motif involved in the interaction of MKT1 with several mRNA-related proteins[57]. In mammalian cells, the LC sequence of Fused in Sarcoma (FUS) is able to form reversible hydrogel droplets in vitro, which can trap other LC sequence-containing molecules[58]. These sequences are predicted to form intrinsically unstructured domains, and were shown to orchestrate the dynamic assembly of RNP granules in yeasts and mammalian cells through homotypic and heterotypic associations[59]. Thus, the polymeric association of different RBPs in RNP complexes seems to control the dynamic assembly of RNA granules. It is therefore the presence of LC sequences, or a combination of these with an RBD, that could be regulating the function of an RBP, and probably of the bound mRNA, through their recruitment to RNP granules.
To add a layer of complexity, Han et al[60] demonstrated that phosphorylation of FUS LC sequence inhibits its association to homotypic hydrogel droplets. The hypothesis arising from these results is that regulatory phosphorylations can prevent LC sequence-containing RBPs to transition into the RNA granule, thus regulating RNP formation and composition[61]. This hypothesis guides our review to probably the most important, yet the less explored, way to modulate the function of an RBP in trypanosomes.
Regulation by PTMs
PTM of proteins has shown to increase the functional diversity of the proteome by the covalent addition of functional groups, peptides or proteins[62]. Amongst the many PTM a protein can suffer, we will discuss here those that were shown to modify cytoplasmic or nuclear RBPs in trypanosomes, namely phosphorylation, SUMOylation, and arginine-methylation. To understand the role these PTMs on RBPs, and many other proteins, will have a huge impact in trypanosomes, since these modifications involve one or more enzymes that could be used as targets for pharmacological intervention for these parasitic diseases.
It was more than 20 years ago when the first RBP with PTMs was identified in T. brucei. This was the case of Nopp44/46, which is an RBP belonging to a family composed by several members with different number of RGG repeats[63]. Nop44/46 is phosphorylated on serine and tyrosine residues, and has been proposed to be required for formation of the large ribosomal subunit in T. brucei[64]. Numerous proteomic approaches have recently been used to identify phosphorylated proteins in T. brucei and T. cruzi (Table 3), since phosphorylation events are especially relevant due to their impact in a broad spectrum of processes and regulatory functions[65]. In T. brucei, the phosphoproteome of the bloodstream form was the first to be explored based on phosphopeptide enrichment coupled to MS[66]. In this study, serine, threonine and tyrosine phosphorylated residues were detected, showing conserved phosphorylation sequence motifs in regulatory kinases, suggesting that phosphorylation-based signaling mechanisms are conserved in trypanosomes[66]. However, a previous genome wide survey for protein kinases in Leishmania, T. brucei and T. cruzi showed these parasites lack receptor-linked tyrosine and tyrosine kinase-like kinases, but instead posses dual specificity kinases[67]. Urbaniak and collaborators[68] reported a quantitative phosphorylation study of the T. brucei bloodstream and procyclic forms by SILAC labeling. The results from this work revealed a significant number of phosphorylated ZFPs, RRM-type RBPs, and mRNA metabolism-related proteins (Table 3), some of which showed lifecycle specific regulation of phosphorylation status. In this work it was proposed that the dynamic phosphorylation of RBPs could have the potential to modulate gene expression, as these proteins could be acting as ultimate effector molecules of some of the trypanosomatid signaling cascades[68]. T. cruzi has also been explored extensively at the phosphoproteomic level[69-71], revealing many new and already T. brucei identified phosphorylated RBP and related proteins (Table 3). In a very interesting study, Queiroz et al[71] performed a comparative proteomic and phosphoproteomic analysis during amastigogenesis, the transition from non-replicating and infective trypomastigotes to replicating non-infective amastigotes. From these analyses several RBPs and mRNA-related proteins were identified as being developmentally regulated between the final morphological stages, or during the transformation process[71]. To note was that several RBPs also showed developmental phosphorylation, suggesting that these modifications could be regulating the function of these proteins. It has been suggested that phosphorylation may induce global conformational changes in an RBP, either promoting or inhibiting protein-protein or protein-RNA interactions allosterically[72]. Thus, regulatory phosphorylation events in RBPs could have an enormous impact on gene expression regulation in trypanosomes due to the prevalence of posttranscriptional mechanisms in these parasites.
Table 3.
RNA-binding proteins and mRNA metabolism-related proteins with identified posttranslational modifications in trypanosomes
| RBP | T. brucei | T. cruzi | Posttranslational modification |
| PUF domain | |||
| PUF1 | Tb927.10.4430 | TcCLB.397937.10/TcCLB.508625.160 | Phosphorylated in T. brucei[68] Phosphorylated in T. cruzi[71] |
| PUF2 | Tb927.10.12660 | TcCLB.507831.110 | Phosphorylated in T. brucei[66,68] Phosphorylated in T. cruzi[69] |
| PUF3 | Tb927.10.310 | TcCLB.508787.30/TcCLB.468005.9 | Phosphorylated in T. brucei[66,68] |
| PUF4 | Tb927.6.820 | TcCLB.509399.190/TcCLB.510073.30 | Phosphorylated in T. brucei[66,68] |
| PUF6 | Tb927.10.15870 | TcCLB.510125.10 | Phosphorylated in T. brucei[66,68] Phosphorylated in T. cruzi[71] Sumoylated in T. cruzi[73] |
| PUF7 | Tb927.11.14960 | TcCLB.508445.99/TcCLB.511715.100 | Sumoylated in T. cruzi[73] |
| PUF9 | Tb927.1.2600 | TcCLB.506563.10/TcCLB.503869.40 | Phosphorylated in T. brucei[68] Arginine-methylated in T. brucei[75] |
| PUF10 | Tb927.11.6740 | TcCLB.506773.130/TcCLB.508799.70 | Arginine-methylated in T. brucei[75] |
| PUF11 | Tb927.11.10810 | TcCLB.503757.30/TcCLB.503719.39 | Phosphorylated in T. brucei[68] |
| Arginine-methylated in T. brucei[75] | |||
| RRM domain | |||
| PABP1 | Tb927.9.9290 | TcCLB.506885.70 | Phosphorylated in T. brucei[68] |
| Phosphorylated in T. cruzi[70,71] Sumoylated in T. cruzi[73] | |||
| PABP2 | Tb927.9.10770 | TcCLB.508461.140 | Phosphorylated in T. brucei[68] |
| UBP1 | Tb927.11.500 | TcCLB.507093.220 | Phosphorylated in T. cruzi[84] |
| RBP3 | Tb927.11.530 | TcCLB.507093.250 | Phosphorylated in T. brucei[68] |
| RBP8 | Tb927.7.320 | TcCLB.504165.20/TcCLB.508981.20 | Phosphorylated in T. brucei[68] |
| RBP9 | Tb927.11.12120 | TcCLB.511127.10/TcCLB.511481.70 | Phosphorylated in T. brucei[68] |
| RBP10 | Tb927.8.2780 | TcCLB.508989.30/TcCLB.509569.120 | Phosphorylated in T. brucei[66,68] |
| RBP12 | Tb927.10.12740 | TcCLB.511277.200 | Phosphorylated in T. brucei[68] Phosphorylated in T. cruzi[71] |
| U1 snRNP (RBP24) | Tb927.8.4830 | TcCLB.508837.140/TcCLB.511383.30 | Phosphorylated in T. brucei[68] |
| RBP28 | Tb927.3.1030 | TcCLB.511871.110/TcCLB.511863.20 | Phosphorylated in T. brucei[68] Arginine-methylated in T. brucei[75] |
| RBP29 | Tb927.10.13720 | TcCLB.511277.580 | Phosphorylated in T. cruzi[69,71] |
| RBP30 | Tb927.5.1750 | TcCLB.505229.20/TcCLB.510823.20 | Phosphorylated in T. brucei[68] |
| RBP33 | Tb927.8.990 | TcCLB.503733.50/TcCLB.508569.90 | Phosphorylated in T. brucei[68] Arginine-methylated in T. brucei[75] |
| RBP35 | Tb927.9.12360 | TcCLB.510741.40 | Phosphorylated in T. brucei[68] Phosphorylated in T. cruzi[70] |
| RBP38 | Tb927.11.5850 | TcCLB.508641.180/TcCLB.508515.30 | Phosphorylated in T. brucei[68] |
| RBP42 | Tb927.6.4440 | TcCLB.509167.140 | Phosphorylated in T. cruzi[71] |
| RBSR1 | Tb927.9.6870 | TcCLB.506625.70 | Phosphorylated in T. brucei[68] Phosphorylated in T. cruzi[69] |
| RBSR4 | Tb927.10.3500 | TcCLB.510265.40/TcCLB.510311.50 | Phosphorylated in T. brucei[68] Arginine-methylated in T. brucei[75] |
| DRBD2 | Tb927.9.13990 | TcCLB.510755.120/TcCLB.508413.50 | Phosphorylated in T. cruzi[69] |
| DRBD5 | Tb927.6.3480 | TcCLB.469785.40/TcCLB.507025.50 | Phosphorylated in T. brucei[68] |
| DRBD7 | Tb927.4.400 | TcCLB.507873.30/TcCLB.510689.60 | Phosphorylated in T. brucei[68] |
| DRBD10 | Tb927.11.16020 | TcCLB.507037.20/TcCLB.508707.80 | Phosphorylated in T. brucei[68] |
| DRBD11 | Tb927.3.3940 | TcCLB.503683.30/TcCLB.509999.120 | Phosphorylated in T. brucei[68] |
| DRBD13 | Tb927.8.6650 | TcCLB.506399.40/TcCLB.509243.20 | Phosphorylated in T. brucei[68] |
| DRBD17 | Tb927.8.710 | TcCLB.507649.100/TcCLB.508567.100 | Phosphorylated in T. brucei[68] Arginine-methylated in T. brucei[75] |
| DRBD18 | Tb927.11.14090 | TcCLB.511727.190 | Arginine-methylated in T. brucei[75] |
| HNRNPH | Tb927.2.3880 | TcCLB.511109.130/TcCLB.504157.10 | Phosphorylated in T. brucei[68] Phosphorylated in T. cruzi[70,71] Sumoylated in T. cruzi[73] |
| TRRM1 | Tb927.2.4710 | TcCLB.509317.60/TcCLB.511621.50 | Phosphorylated in T. brucei[66,68] Phosphorylated in T. cruzi[69] Sumoylated in T. cruzi[73] |
| TRRM3 | Tb927.3.3670 | TcCLB.506989.100/TcCLB.510149.140 | Arginine-methylated in T. brucei[75] |
| PPCTI | Tb927.5.3750 | TcCLB.503619.20/TcCLB.511647.40 | Phosphorylated in T. brucei[68] |
| MRD1 | Tb927.8.4170 | TcCLB.509561.110/TcCLB.503897.90 | Phosphorylated in T. brucei[68] |
| TSR1 | Tb927.8.900 | TcCLB.509607.30/TcCLB.503715.10 | Phosphorylated in T. brucei[68] |
| NRBD1/2 (P34/P37) | Tb927.11.14000/Tb927.11.14020 | TcCLB.511727.270/TcCLB.511727.290 | Arginine methylated in T. brucei[75] |
| Conserved | Tb927.10.7030 | TcCLB.506779.100/TcCLB.511153.100 | Arginine-methylated in T. brucei[75] |
| Conserved | Tb927.11.7310 | TcCLB.506779.100/TcCLB.511153.100 | Arginine-methylated in T. brucei[75] |
| Conserved | Tb927.11.14090 | TcCLB.511727.190 | Arginine-methylated in T. brucei[75] |
| Conserved | Tb927.11.6240 | TcCLB.506297.230/TcCLB.510101.80 | Arginine-methylated in T. brucei[75] |
| Zinc finger domains | |||
| ZFP1 | Tb927.6.3490 | TcCLB.511511.6 | Phosphorylated in T. brucei[66,68] |
| ZFP3 | Tb927.3.720 | TcCLB.509719.69/TcCLB.509231.39 | Phosphorylated in T. brucei[68] |
| ZC3H5 | Tb927.3.740 | TcCLB.511867.10/TcCLB.507775.10 | Phosphorylated in T. brucei[68] |
| ZC3H7 | Tb927.3.1340 | TcCLB.509233.210 | Phosphorylated in T. brucei[68] |
| ZC3H8 | Tb927.3.5250 | TcCLB.510143.120/TcCLB.508409.310 | Phosphorylated in T. brucei[68] |
| Arginine-methylated in T. brucei[75] | |||
| ZC3H11 | Tb927.5.810 | TcCLB.507305.40 | Phosphorylated in T. brucei[68,85] |
| ZC3H12 | Tb927.5.1570 | TcCLB.510819.119/TcCLB.506739.99 | Phosphorylated in T. brucei[86] |
| ZC3H13 | Tb927.5.1580 | No homologue | Phosphorylated in T. brucei[66,68] |
| ZC3H17 | Tb927.7.930 | TcCLB.508879.10/TcCLB.508215.10 | Phosphorylated in T. brucei[68] |
| ZC3H18 | Tb927.7.2140 | TcCLB.511807.160 | Phosphorylated in T. brucei[68] |
| ZC3H20 | Tb927.7.2660 | TcCLB.503567.9/TcCLB.506859.204 | Phosphorylated in T. brucei[68] |
| ZC3H21 | Tb927.7.2670 | TcCLB.506859.230/TcCLB.511817.10 | Phosphorylated in T. brucei[68] |
| ZC3H22 | Tb927.7.2680 | TcCLB.506859.240/TcCLB.511817.20 | Phosphorylated in T. brucei[68] |
| ZC3H23 | Tb927.7.4980 | TcCLB.509149.20/TcCLB.508175.350 | Phosphorylated in T. brucei[68] |
| ZC3H28 | Tb927.9.9450 | TcCLB.506885.200/TcCLB.510729.220 | Phosphorylated in T. brucei[68] Phosphorylated in T. cruzi[70] |
| ZC3H29 | Tb927.9.9520 | TcCLB.510729.210/TcCLB.506885.204 | Phosphorylated in T. brucei[68] |
| ZC3H30 | Tb927.10.1540 | TcCLB.506977.110 | Phosphorylated in T. brucei[68] |
| ZC3H31 | Tb927.10.5150 | TcCLB.506009.10/TcCLB.510295.59 | Sumoylated in T. cruzi[73] |
| ZC3H32 | Tb927.10.5250 | TcCLB.503795.10 /TcCLB.506679.10 | Phosphorylated in T. brucei[68] |
| ZC3H34 | Tb927.10.12330 | TcCLB.507787.140/TcCLB.507625.70 | Phosphorylated in T. brucei[68] Arginine-methylated in T. brucei[75] |
| ZC3H35 | Tb927.10.12740 | TcCLB.511263.30/TcCLB.507831.40 | Phosphorylated in T. brucei[68] |
| ZC3H37/38 | Tb927.10.12780 | TcCLB.507831.20/TcCLB.511267.20/ TcCLB.511263.50 | Phosphorylated in T. brucei[68] |
| ZC3H39 | Tb927.10.14930 | TcCLB.508895.50 | Phosphorylated in T. brucei[68] |
| ZC3H40 | Tb927.10.14950 | TcCLB.508895.60/TcCLB.506211.60 | Phosphorylated in T. brucei[66,68] Phosphorylated in T. cruzi[69,71] |
| ZC3H41 | Tb927.11.1980 | TcCLB.508355.330/TcCLB.508357.9 | Phosphorylated in T. brucei[68] |
| ZC3H43 | Tb927.11.7450 | TcCLB.511151.20/TcCLB.508241.90 | Phosphorylated in T. brucei[68] |
| ZC3H44 | Tb927.11.7890 | TcCLB.506933.50 | Phosphorylated in T. brucei[68] |
| ZC3H46 | Tb927.11.16550 | TcCLB.507089.30/TcCLB.504085.70 | Phosphorylated in T. brucei[68] |
| ZC3H47 | Tb927.6.4960 | TcCLB.506945.210 | Phosphorylated in T. brucei[68] Phosphorylated in T. cruzi[69] |
| Conserved | Tb927.11.3970 | TcCLB.509229.90/TcCLB.506733.140 | Phosphorylated in T. brucei[68] |
| Conserved | Tb927.9.14120 | TcCLB.510759.100/TcCLB.506999.120 | Arginine-methylated in T. brucei[75] |
| Other | |||
| NOP44/46 | Tb927.8.760 | TcCLB.510859.17/TcCLB.510859.10 | Phosphorylated in T. brucei[68] Arginine-methylated in T. brucei[75] |
| SMD2 | Tb927.2.5850 | TcCLB.508667.49/TcCLB.511189.80 | Phosphorylated in T. brucei[68] |
| DCL2 | Tb927.3.1230 | No homologue | Phosphorylated in T. brucei[68] Arginine-methylated in T. brucei[75] |
| CAF1 | Tb927.6.600 | TcCLB.511827.60/TcCLB.510535.60 | Phosphorylated in T. brucei[68] |
| Not1 | Tb927.10.1510 | TcCLB.509247.30 | Phosphorylated in T. cruzi[70] |
| XRNA | Tb927.7.4900 | TcCLB.507817.80 | Phosphorylated in T. cruzi[69] |
| ALBA3 | Tb927.4.2040 | TcCLB.510877.40 | Phosphorylated in T. brucei[68] Arginine-methylated in T. brucei[75] |
| ALBA4 | Tb927.4.2030 | TcCLB.510877.30 | Phosphorylated in T. brucei[68] Arginine-methylated in T. brucei[75] |
| eIF4E | Tb927.11.11770 | TcCLB.508827.30 | Phosphorylated in T. brucei[68] Phosphorylated in T. cruzi[70] |
| Tb927.6.1870 | TcCLB.509037.40/TcCLB.421959.10 | Phosphorylated inT. brucei[68] | |
| eIF4G2 | Tb927.9.5460 | TcCLB.508277.340/TcCLB.506445.20 | Phosphorylated in T. brucei[68] |
| Tb927.11.10560 | TcCLB.510285.100/TcCLB.504827.130 | Phosphorylated in T. brucei[68] | |
| eIF4G5 | Tb927.8.4500 | TcCLB.508989.90 | Phosphorylated in T. brucei[68] |
RBP: RNA-binding protein; RRM: RNA-recognition motif.
In another study Bayona et al[73] reported the functionality of the small ubiquitin-like modifier (SUMO) pathway in T. cruzi. The addition of SUMO is a covalent and reversible modification that can usually affect a protein’s normal function. Initially SUMOylation was believed to be only involved in nuclear events such as nucleocytoplasmic transport, DNA replication, gene transcription and DNA damage response, but with the identification of numerous other targets it now seems that SUMOylation can affect numerous processes at both sides of the nuclear envelope[74]. Within the many proteins modified by SUMO addition in T. cruzi, some belong to the RBP category (Table 3)[73]. Given that this type of modification is usually very unstable and it is lost during typical experimental manipulations, adequate experimental approaches will be required in the future to address the effect of SUMOylation on RBPs in trypanosomes.
Last but not least, the potential of arginine-methylation was explored by a global proteomic analysis in T. brucei[75]. In this work, methylated arginine residues were identified mainly in glycine-rich contexts, as described in other organisms. The functional classification of proteins derived from this work revealed an interesting amount of proteins involved in mRNA metabolism (Table 3)[75], and other related proteins. The relevance of this finding in T. brucei is highlighted by the impact that arginine-methylation can have on protein-protein and protein-nucleic acid interactions and subcellular localization of proteins[76]. DRBD18, a double RRM-containing RBP, was initially identified as having three methyl-arginines[75]. In a posterior characterization of DRBD18 in T. brucei, methylmimetic or hypomethylated mutants were expressed in parasites downregulated for endogenous DRBD18. Both mutants exhibited a differential modulation of the T. brucei transcriptome, clearly showing opposing effects of due to methylarginine content[77]. Surprisingly, these differences did not seem to be regulated at the level of subcellular localization or affinity for RNA molecules, since both mutants showed similar association to RNA molecules in the cytoplasm. Instead, the methylation state of DRBD18 arginine residues seemed to modulate the interactions of these DRBD18 variants with other protein factors involved in mRNA metabolism, since they showed to associate to different RNP complexes (Figure 1D). Thus, differential arginine-methylation was proposed to shape the composition of DRBD18 RNP complexes, and thus to modulate the fate of the associated mRNA[77].
CONCLUSION
Although the above-described events might seem yet unrelated, future work might provide evidences to connect different mechanisms to regulate the function of an RBP in trypanosomes. This is especially relevant for localization, aggregation and interaction events that might be regulated by posttranslational modifications. Given that RBPs do not have enzymatic activities, and hence cannot be tested as druggable targets, it is tempting to speculate that the enzymes (kinases, phosphatases, SUMO E3 ligases and methyltransferases) responsible for these posttranslational modifications might be indeed good candidates for drug therapy. Approaches targeting any of these protein modifications are likely to have a very high impact for these neglected diseases, either by discovery of new drugs or by drug repurposing.
Footnotes
Supported by The Agencia Nacional de Promoción Científica y Tecnológica (ANPCyT) to Alejandro Cassola.
Conflict-of-interest statement: The authors declare no conflicts of interest regarding this manuscript.
Open-Access: This article is an open-access article that was selected by an in-house editor and fully peer-reviewed by external reviewers. It is distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Peer-review started: June 1, 2015
First decision: August 8, 2015
Article in press: January 29, 2016
P- Reviewer: Lee HC, Rholam M S- Editor: Qiu S L- Editor: A E- Editor: Jiao XK
References
- 1.Brun R, Blum J, Chappuis F, Burri C. Human African trypanosomiasis. Lancet. 2010;375:148–159. doi: 10.1016/S0140-6736(09)60829-1. [DOI] [PubMed] [Google Scholar]
- 2.Rassi A, Rassi A, Marin-Neto JA. Chagas disease. Lancet. 2010;375:1388–1402. doi: 10.1016/S0140-6736(10)60061-X. [DOI] [PubMed] [Google Scholar]
- 3.Stuart K, Brun R, Croft S, Fairlamb A, Gürtler RE, McKerrow J, Reed S, Tarleton R. Kinetoplastids: related protozoan pathogens, different diseases. J Clin Invest. 2008;118:1301–1310. doi: 10.1172/JCI33945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Patterson S, Wyllie S. Nitro drugs for the treatment of trypanosomatid diseases: past, present, and future prospects. Trends Parasitol. 2014;30:289–298. doi: 10.1016/j.pt.2014.04.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Daniels JP, Gull K, Wickstead B. Cell biology of the trypanosome genome. Microbiol Mol Biol Rev. 2010;74:552–569. doi: 10.1128/MMBR.00024-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.El-Sayed NM, Myler PJ, Blandin G, Berriman M, Crabtree J, Aggarwal G, Caler E, Renauld H, Worthey EA, Hertz-Fowler C, et al. Comparative genomics of trypanosomatid parasitic protozoa. Science. 2005;309:404–409. doi: 10.1126/science.1112181. [DOI] [PubMed] [Google Scholar]
- 7.Ekanayake DK, Minning T, Weatherly B, Gunasekera K, Nilsson D, Tarleton R, Ochsenreiter T, Sabatini R. Epigenetic regulation of transcription and virulence in Trypanosoma cruzi by O-linked thymine glucosylation of DNA. Mol Cell Biol. 2011;31:1690–1700. doi: 10.1128/MCB.01277-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Siegel TN, Hekstra DR, Kemp LE, Figueiredo LM, Lowell JE, Fenyo D, Wang X, Dewell S, Cross GA. Four histone variants mark the boundaries of polycistronic transcription units in Trypanosoma brucei. Genes Dev. 2009;23:1063–1076. doi: 10.1101/gad.1790409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Michaeli S. Trans-splicing in trypanosomes: machinery and its impact on the parasite transcriptome. Future Microbiol. 2011;6:459–474. doi: 10.2217/fmb.11.20. [DOI] [PubMed] [Google Scholar]
- 10.Ouellette M, Papadopoulou B. Coordinated gene expression by post-transcriptional regulons in African trypanosomes. J Biol. 2009;8:100. doi: 10.1186/jbiol203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Najafabadi HS, Lu Z, MacPherson C, Mehta V, Adoue V, Pastinen T, Salavati R. Global identification of conserved post-transcriptional regulatory programs in trypanosomatids. Nucleic Acids Res. 2013;41:8591–8600. doi: 10.1093/nar/gkt647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kolev NG, Ullu E, Tschudi C. The emerging role of RNA-binding proteins in the life cycle of Trypanosoma brucei. Cell Microbiol. 2014;16:482–489. doi: 10.1111/cmi.12268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Gull K. The biology of kinetoplastid parasites: insights and challenges from genomics and post-genomics. Int J Parasitol. 2001;31:443–452. doi: 10.1016/s0020-7519(01)00154-0. [DOI] [PubMed] [Google Scholar]
- 14.Berriman M, Ghedin E, Hertz-Fowler C, Blandin G, Renauld H, Bartholomeu DC, Lennard NJ, Caler E, Hamlin NE, Haas B, et al. The genome of the African trypanosome Trypanosoma brucei. Science. 2005;309:416–422. doi: 10.1126/science.1112642. [DOI] [PubMed] [Google Scholar]
- 15.El-Sayed NM, Myler PJ, Bartholomeu DC, Nilsson D, Aggarwal G, Tran AN, Ghedin E, Worthey EA, Delcher AL, Blandin G, et al. The genome sequence of Trypanosoma cruzi, etiologic agent of Chagas disease. Science. 2005;309:409–415. doi: 10.1126/science.1112631. [DOI] [PubMed] [Google Scholar]
- 16.Ullu E, Tschudi C, Chakraborty T. RNA interference in protozoan parasites. Cell Microbiol. 2004;6:509–519. doi: 10.1111/j.1462-5822.2004.00399.x. [DOI] [PubMed] [Google Scholar]
- 17.Clayton C. The regulation of trypanosome gene expression by RNA-binding proteins. PLoS Pathog. 2013;9:e1003680. doi: 10.1371/journal.ppat.1003680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Jensen KB, Darnell RB. CLIP: crosslinking and immunoprecipitation of in vivo RNA targets of RNA-binding proteins. Methods Mol Biol. 2008;488:85–98. doi: 10.1007/978-1-60327-475-3_6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Atwood JA, Weatherly DB, Minning TA, Bundy B, Cavola C, Opperdoes FR, Orlando R, Tarleton RL. The Trypanosoma cruzi proteome. Science. 2005;309:473–476. doi: 10.1126/science.1110289. [DOI] [PubMed] [Google Scholar]
- 20.Cirovic O, Ochsenreiter T. Whole proteome analysis of the protozoan parasite Trypanosoma brucei using stable isotope labeling by amino acids in cell culture and mass spectrometry. Methods Mol Biol. 2014;1188:47–55. doi: 10.1007/978-1-4939-1142-4_4. [DOI] [PubMed] [Google Scholar]
- 21.Kolev NG, Franklin JB, Carmi S, Shi H, Michaeli S, Tschudi C. The transcriptome of the human pathogen Trypanosoma brucei at single-nucleotide resolution. PLoS Pathog. 2010;6:e1001090. doi: 10.1371/journal.ppat.1001090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Minning TA, Weatherly DB, Atwood J, Orlando R, Tarleton RL. The steady-state transcriptome of the four major life-cycle stages of Trypanosoma cruzi. BMC Genomics. 2009;10:370. doi: 10.1186/1471-2164-10-370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Zhang J, Ruyechan W, Williams N. Developmental regulation of two nuclear RNA binding proteins, p34 and p37, from Trypanosoma brucei. Mol Biochem Parasitol. 1998;92:79–88. doi: 10.1016/s0166-6851(97)00228-4. [DOI] [PubMed] [Google Scholar]
- 24.Prohaska K, Williams N. Assembly of the Trypanosoma brucei 60S ribosomal subunit nuclear export complex requires trypanosome-specific proteins P34 and P37. Eukaryot Cell. 2009;8:77–87. doi: 10.1128/EC.00234-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.D’Orso I, Frasch AC. TcUBP-1, an mRNA destabilizing factor from trypanosomes, homodimerizes and interacts with novel AU-rich element- and Poly(A)-binding proteins forming a ribonucleoprotein complex. J Biol Chem. 2002;277:50520–50528. doi: 10.1074/jbc.M209092200. [DOI] [PubMed] [Google Scholar]
- 26.Hendriks EF, Matthews KR. Disruption of the developmental programme of Trypanosoma brucei by genetic ablation of TbZFP1, a differentiation-enriched CCCH protein. Mol Microbiol. 2005;57:706–716. doi: 10.1111/j.1365-2958.2005.04679.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Simpson L. The mitochondrial genome of kinetoplastid protozoa: genomic organization, transcription, replication, and evolution. Annu Rev Microbiol. 1987;41:363–382. doi: 10.1146/annurev.mi.41.100187.002051. [DOI] [PubMed] [Google Scholar]
- 28.Wurst M, Seliger B, Jha BA, Klein C, Queiroz R, Clayton C. Expression of the RNA recognition motif protein RBP10 promotes a bloodstream-form transcript pattern in Trypanosoma brucei. Mol Microbiol. 2012;83:1048–1063. doi: 10.1111/j.1365-2958.2012.07988.x. [DOI] [PubMed] [Google Scholar]
- 29.Kolev NG, Ramey-Butler K, Cross GA, Ullu E, Tschudi C. Developmental progression to infectivity in Trypanosoma brucei triggered by an RNA-binding protein. Science. 2012;338:1352–1353. doi: 10.1126/science.1229641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Marchetti MA, Tschudi C, Kwon H, Wolin SL, Ullu E. Import of proteins into the trypanosome nucleus and their distribution at karyokinesis. J Cell Sci. 2000;113(Pt5):899–906. doi: 10.1242/jcs.113.5.899. [DOI] [PubMed] [Google Scholar]
- 31.Sakyiama J, Zimmer SL, Ciganda M, Williams N, Read LK. Ribosome biogenesis requires a highly diverged XRN family 5’-& gt; 3’ exoribonuclease for rRNA processing in Trypanosoma brucei. RNA. 2013;19:1419–1431. doi: 10.1261/rna.038547.113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Li H, Tschudi C. Novel and essential subunits in the 300-kilodalton nuclear cap binding complex of Trypanosoma brucei. Mol Cell Biol. 2005;25:2216–2226. doi: 10.1128/MCB.25.6.2216-2226.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Biton M, Mandelboim M, Arvatz G, Michaeli S. RNAi interference of XPO1 and Sm genes and their effect on the spliced leader RNA in Trypanosoma brucei. Mol Biochem Parasitol. 2006;150:132–143. doi: 10.1016/j.molbiopara.2006.07.004. [DOI] [PubMed] [Google Scholar]
- 34.Tkacz ID, Cohen S, Salmon-Divon M, Michaeli S. Identification of the heptameric Lsm complex that binds U6 snRNA in Trypanosoma brucei. Mol Biochem Parasitol. 2008;160:22–31. doi: 10.1016/j.molbiopara.2008.03.003. [DOI] [PubMed] [Google Scholar]
- 35.Ericsson AO, Faria LO, Cruz WB, Martins de Sá C, Lima BD. TcZFP8, a novel member of the Trypanosoma cruzi CCHC zinc finger protein family with nuclear localization. Genet Mol Res. 2006;5:553–563. [PubMed] [Google Scholar]
- 36.Fernández-Moya SM, Carrington M, Estévez AM. Depletion of the RNA-binding protein RBP33 results in increased expression of silenced RNA polymerase II transcripts in Trypanosoma brucei. PLoS One. 2014;9:e107608. doi: 10.1371/journal.pone.0107608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Cassola A, Frasch AC. An RNA recognition motif mediates the nucleocytoplasmic transport of a trypanosome RNA-binding protein. J Biol Chem. 2009;284:35015–35028. doi: 10.1074/jbc.M109.031633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Mittal N, Roy N, Babu MM, Janga SC. Dissecting the expression dynamics of RNA-binding proteins in posttranscriptional regulatory networks. Proc Natl Acad Sci USA. 2009;106:20300–20305. doi: 10.1073/pnas.0906940106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Chen CY, Shyu AB. Emerging mechanisms of mRNP remodeling regulation. Wiley Interdiscip Rev RNA. 2014;5:713–722. doi: 10.1002/wrna.1241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Cassola A, Noé G, Frasch AC. RNA recognition motifs involved in nuclear import of RNA-binding proteins. RNA Biol. 2010;7:339–344. doi: 10.4161/rna.7.3.12087. [DOI] [PubMed] [Google Scholar]
- 41.Fernández-Moya SM, García-Pérez A, Kramer S, Carrington M, Estévez AM. Alterations in DRBD3 ribonucleoprotein complexes in response to stress in Trypanosoma brucei. PLoS One. 2012;7:e48870. doi: 10.1371/journal.pone.0048870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Kramer S, Bannerman-Chukualim B, Ellis L, Boulden EA, Kelly S, Field MC, Carrington M. Differential localization of the two T. brucei poly(A) binding proteins to the nucleus and RNP granules suggests binding to distinct mRNA pools. PLoS One. 2013;8:e54004. doi: 10.1371/journal.pone.0054004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.McNally KP, Agabian N. Trypanosoma brucei spliced-leader RNA methylations are required for trans splicing in vivo. Mol Cell Biol. 1992;12:4844–4851. doi: 10.1128/mcb.12.11.4844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Kramer S, Carrington M. Trans-acting proteins regulating mRNA maturation, stability and translation in trypanosomatids. Trends Parasitol. 2011;27:23–30. doi: 10.1016/j.pt.2010.06.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Cassola A. RNA Granules Living a Post-transcriptional Life: the Trypanosomes’ Case. Curr Chem Biol. 2011;5:108–117. doi: 10.2174/2212796811105020108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Cassola A, De Gaudenzi JG, Frasch AC. Recruitment of mRNAs to cytoplasmic ribonucleoprotein granules in trypanosomes. Mol Microbiol. 2007;65:655–670. doi: 10.1111/j.1365-2958.2007.05833.x. [DOI] [PubMed] [Google Scholar]
- 47.Narayanaswamy R, Levy M, Tsechansky M, Stovall GM, O’Connell JD, Mirrielees J, Ellington AD, Marcotte EM. Widespread reorganization of metabolic enzymes into reversible assemblies upon nutrient starvation. Proc Natl Acad Sci USA. 2009;106:10147–10152. doi: 10.1073/pnas.0812771106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Li CH, Irmer H, Gudjonsdottir-Planck D, Freese S, Salm H, Haile S, Estévez AM, Clayton C. Roles of a Trypanosoma brucei 5’-& gt; 3’ exoribonuclease homolog in mRNA degradation. RNA. 2006;12:2171–2186. doi: 10.1261/rna.291506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Krüger T, Hofweber M, Kramer S. SCD6 induces ribonucleoprotein granule formation in trypanosomes in a translation-independent manner, regulated by its Lsm and RGG domains. Mol Biol Cell. 2013;24:2098–2111. doi: 10.1091/mbc.E13-01-0068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.De Gaudenzi JG, Noé G, Campo VA, Frasch AC, Cassola A. Gene expression regulation in trypanosomatids. Essays Biochem. 2011;51:31–46. doi: 10.1042/bse0510031. [DOI] [PubMed] [Google Scholar]
- 51.Cristodero M, Schimanski B, Heller M, Roditi I. Functional characterization of the trypanosome translational repressor SCD6. Biochem J. 2014;457:57–67. doi: 10.1042/BJ20130747. [DOI] [PubMed] [Google Scholar]
- 52.Fritz M, Vanselow J, Sauer N, Lamer S, Goos C, Siegel TN, Subota I, Schlosser A, Carrington M, Kramer S. Novel insights into RNP granules by employing the trypanosome’s microtubule skeleton as a molecular sieve. Nucleic Acids Res. 2015;43:8013–8032. doi: 10.1093/nar/gkv731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Lunde BM, Moore C, Varani G. RNA-binding proteins: modular design for efficient function. Nat Rev Mol Cell Biol. 2007;8:479–490. doi: 10.1038/nrm2178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Decker CJ, Teixeira D, Parker R. Edc3p and a glutamine/asparagine-rich domain of Lsm4p function in processing body assembly in Saccharomyces cerevisiae. J Cell Biol. 2007;179:437–449. doi: 10.1083/jcb.200704147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Reijns MA, Alexander RD, Spiller MP, Beggs JD. A role for Q/N-rich aggregation-prone regions in P-body localization. J Cell Sci. 2008;121:2463–2472. doi: 10.1242/jcs.024976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.De Gaudenzi J, Frasch AC, Clayton C. RNA-binding domain proteins in Kinetoplastids: a comparative analysis. Eukaryot Cell. 2005;4:2106–2114. doi: 10.1128/EC.4.12.2106-2114.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Singh A, Minia I, Droll D, Fadda A, Clayton C, Erben E. Trypanosome MKT1 and the RNA-binding protein ZC3H11: interactions and potential roles in post-transcriptional regulatory networks. Nucleic Acids Res. 2014;42:4652–4668. doi: 10.1093/nar/gkt1416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Kato M, Han TW, Xie S, Shi K, Du X, Wu LC, Mirzaei H, Goldsmith EJ, Longgood J, Pei J, et al. Cell-free formation of RNA granules: low complexity sequence domains form dynamic fibers within hydrogels. Cell. 2012;149:753–767. doi: 10.1016/j.cell.2012.04.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Jonas S, Izaurralde E. The role of disordered protein regions in the assembly of decapping complexes and RNP granules. Genes Dev. 2013;27:2628–2641. doi: 10.1101/gad.227843.113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Han TW, Kato M, Xie S, Wu LC, Mirzaei H, Pei J, Chen M, Xie Y, Allen J, Xiao G, et al. Cell-free formation of RNA granules: bound RNAs identify features and components of cellular assemblies. Cell. 2012;149:768–779. doi: 10.1016/j.cell.2012.04.016. [DOI] [PubMed] [Google Scholar]
- 61.Weber SC, Brangwynne CP. Getting RNA and protein in phase. Cell. 2012;149:1188–1191. doi: 10.1016/j.cell.2012.05.022. [DOI] [PubMed] [Google Scholar]
- 62.Beck-Sickinger AG, Mörl K. Posttranslational modification of proteins. Expanding nature’s inventory. Angew Chem Int Ed Engl. 2006;45:1020–1020. [Google Scholar]
- 63.Parsons M, Ledbetter JA, Schieven GL, Nel AE, Kanner SB. Developmental regulation of pp44/46, tyrosine-phosphorylated proteins associated with tyrosine/serine kinase activity in Trypanosoma brucei. Mol Biochem Parasitol. 1994;63:69–78. doi: 10.1016/0166-6851(94)90009-4. [DOI] [PubMed] [Google Scholar]
- 64.Jensen BC, Brekken DL, Randall AC, Kifer CT, Parsons M. Species specificity in ribosome biogenesis: a nonconserved phosphoprotein is required for formation of the large ribosomal subunit in Trypanosoma brucei. Eukaryot Cell. 2005;4:30–35. doi: 10.1128/EC.4.1.30-35.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Jensen ON. Modification-specific proteomics: characterization of post-translational modifications by mass spectrometry. Curr Opin Chem Biol. 2004;8:33–41. doi: 10.1016/j.cbpa.2003.12.009. [DOI] [PubMed] [Google Scholar]
- 66.Nett IR, Martin DM, Miranda-Saavedra D, Lamont D, Barber JD, Mehlert A, Ferguson MA. The phosphoproteome of bloodstream form Trypanosoma brucei, causative agent of African sleeping sickness. Mol Cell Proteomics. 2009;8:1527–1538. doi: 10.1074/mcp.M800556-MCP200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Parsons M, Worthey EA, Ward PN, Mottram JC. Comparative analysis of the kinomes of three pathogenic trypanosomatids: Leishmania major, Trypanosoma brucei and Trypanosoma cruzi. BMC Genomics. 2005;6:127. doi: 10.1186/1471-2164-6-127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Urbaniak MD, Martin DM, Ferguson MA. Global quantitative SILAC phosphoproteomics reveals differential phosphorylation is widespread between the procyclic and bloodstream form lifecycle stages of Trypanosoma brucei. J Proteome Res. 2013;12:2233–2244. doi: 10.1021/pr400086y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Marchini FK, de Godoy LM, Rampazzo RC, Pavoni DP, Probst CM, Gnad F, Mann M, Krieger MA. Profiling the Trypanosoma cruzi phosphoproteome. PLoS One. 2011;6:e25381. doi: 10.1371/journal.pone.0025381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Nakayasu ES, Gaynor MR, Sobreira TJ, Ross JA, Almeida IC. Phosphoproteomic analysis of the human pathogen Trypanosoma cruzi at the epimastigote stage. Proteomics. 2009;9:3489–3506. doi: 10.1002/pmic.200800874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Queiroz RM, Charneau S, Mandacaru SC, Schwämmle V, Lima BD, Roepstorff P, Ricart CA. Quantitative proteomic and phosphoproteomic analysis of Trypanosoma cruzi amastigogenesis. Mol Cell Proteomics. 2014;13:3457–3472. doi: 10.1074/mcp.M114.040329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Thapar R. Structural basis for regulation of RNA-binding proteins by phosphorylation. ACS Chem Biol. 2015;10:652–666. doi: 10.1021/cb500860x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Bayona JC, Nakayasu ES, Laverrière M, Aguilar C, Sobreira TJ, Choi H, Nesvizhskii AI, Almeida IC, Cazzulo JJ, Alvarez VE. SUMOylation pathway in Trypanosoma cruzi: functional characterization and proteomic analysis of target proteins. Mol Cell Proteomics. 2011;10:M110.007369. doi: 10.1074/mcp.M110.007369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Meulmeester E, Melchior F. Cell biology: SUMO. Nature. 2008;452:709–711. doi: 10.1038/452709a. [DOI] [PubMed] [Google Scholar]
- 75.Lott K, Li J, Fisk JC, Wang H, Aletta JM, Qu J, Read LK. Global proteomic analysis in trypanosomes reveals unique proteins and conserved cellular processes impacted by arginine methylation. J Proteomics. 2013;91:210–225. doi: 10.1016/j.jprot.2013.07.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Bedford MT, Clarke SG. Protein arginine methylation in mammals: who, what, and why. Mol Cell. 2009;33:1–13. doi: 10.1016/j.molcel.2008.12.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Lott K, Mukhopadhyay S, Li J, Wang J, Yao J, Sun Y, Qu J, Read LK. Arginine methylation of DRBD18 differentially impacts its opposing effects on the trypanosome transcriptome. Nucleic Acids Res. 2015;43:5501–5523. doi: 10.1093/nar/gkv428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Ling AS, Trotter JR, Hendriks EF. A zinc finger protein, TbZC3H20, stabilizes two developmentally regulated mRNAs in trypanosomes. J Biol Chem. 2011;286:20152–20162. doi: 10.1074/jbc.M110.139261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Subota I, Rotureau B, Blisnick T, Ngwabyt S, Durand-Dubief M, Engstler M, Bastin P. ALBA proteins are stage regulated during trypanosome development in the tsetse fly and participate in differentiation. Mol Biol Cell. 2011;22:4205–4219. doi: 10.1091/mbc.E11-06-0511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Gupta SK, Kosti I, Plaut G, Pivko A, Tkacz ID, Cohen-Chalamish S, Biswas DK, Wachtel C, Waldman Ben-Asher H, Carmi S, et al. The hnRNP F/H homologue of Trypanosoma brucei is differentially expressed in the two life cycle stages of the parasite and regulates splicing and mRNA stability. Nucleic Acids Res. 2013;41:6577–6594. doi: 10.1093/nar/gkt369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.De Gaudenzi JG, D’Orso I, Frasch AC. RNA recognition motif-type RNA-binding proteins in Trypanosoma cruzi form a family involved in the interaction with specific transcripts in vivo. J Biol Chem. 2003;278:18884–18894. doi: 10.1074/jbc.M301756200. [DOI] [PubMed] [Google Scholar]
- 82.Pérez-Díaz L, Duhagon MA, Smircich P, Sotelo-Silveira J, Robello C, Krieger MA, Goldenberg S, Williams N, Dallagiovanna B, Garat B. Trypanosoma cruzi: molecular characterization of an RNA binding protein differentially expressed in the parasite life cycle. Exp Parasitol. 2007;117:99–105. doi: 10.1016/j.exppara.2007.03.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Mörking PA, Rampazzo Rde C, Walrad P, Probst CM, Soares MJ, Gradia DF, Pavoni DP, Krieger MA, Matthews K, Goldenberg S, et al. The zinc finger protein TcZFP2 binds target mRNAs enriched during Trypanosoma cruzi metacyclogenesis. Mem Inst Oswaldo Cruz. 2012;107:790–799. doi: 10.1590/s0074-02762012000600014. [DOI] [PubMed] [Google Scholar]
- 84.Cassola A, Romaniuk MA, Primrose D, Cervini G, D’Orso I, Frasch AC. Association of UBP1 to ribonucleoprotein complexes is regulated by interaction with the trypanosome ortholog of the human multifunctional P32 protein. Mol Microbiol. 2015;97:1079–1096. doi: 10.1111/mmi.13090. [DOI] [PubMed] [Google Scholar]
- 85.Droll D, Minia I, Fadda A, Singh A, Stewart M, Queiroz R, Clayton C. Post-transcriptional regulation of the trypanosome heat shock response by a zinc finger protein. PLoS Pathog. 2013;9:e1003286. doi: 10.1371/journal.ppat.1003286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Ouna BA, Stewart M, Helbig C, Clayton C. The Trypanosoma brucei CCCH zinc finger proteins ZC3H12 and ZC3H13. Mol Biochem Parasitol. 2012;183:184–188. doi: 10.1016/j.molbiopara.2012.02.006. [DOI] [PubMed] [Google Scholar]