

Abstract
Thermus amyloliquefaciens type strain YIM 77409T is a thermophilic, Gram-negative, non-motile and rod-shaped bacterium isolated from Niujie Hot Spring in Eryuan County, Yunnan Province, southwest China. In the present study we describe the features of strain YIM 77409T together with its genome sequence and annotation. The genome is 2,160,855 bp long and consists of 6 scaffolds with 67.4 % average GC content. A total of 2,313 genes were predicted, comprising 2,257 protein-coding and 56 RNA genes. The genome is predicted to encode a complete glycolysis, pentose phosphate pathway, and tricarboxylic acid cycle. Additionally, a large number of transporters and enzymes for heterotrophy highlight the broad heterotrophic lifestyle of this organism. A denitrification gene cluster included genes predicted to encode enzymes for the sequential reduction of nitrate to nitrous oxide, consistent with the incomplete denitrification phenotype of this strain.
Keywords: Thermus, Thermus amyloliquefaciens, Thermophiles, Hot springs, Denitrification
Introduction
Thermus species have been isolated from both natural and man-made thermal environments such as hot springs, hot domestic water, deep mines, composting systems, and sewage sludge [1–5]. The genus has attracted considerable attention as a source of thermostable enzymes, which have important biotechnological applications [6], and as a model organism to study the mechanisms involved in bacterial adaptation to extreme environments [7]. Members of the genus Thermus were formerly considered to be strictly aerobic, based on the characteristics of the type species Thermus aquaticus [2]. However, many studies have shown that Thermus strains also can grow as facultative anaerobes using nitrogen oxides, sulfur, or metals as terminal electron acceptors under oxygen-deprived conditions [8–10]. Cava et al. [11] demonstrated that different T. thermophilus strains can grow anaerobically by reducing nitrate to nitrite or by reducing nitrite to a gaseous nitrogen product.
The nitrogen biogeochemical cycle has been investigated in a few geothermal systems [12], including Great Boiling Spring, a ~80 °C hot spring in the U.S. Great Basin [13–15]. Studies in GBS revealed a high flux of nitrous oxide, particularly in the ~80 °C source pool, suggesting the importance of incomplete denitrifiers in high-temperature environments. A subsequent cultivation and physiological study of heterotrophic denitrifiers suggested a significant role of T. oshimai and T. thermophilus in denitrification in this hot spring [16]. A following study of the whole genomes of one strain from each species, T. oshimai JL-2 and T. thermophilus JL-18, revealed that they have genes encoding the sequential reduction of nitrate to nitrous oxide but lack genes encoding the nitrous oxide reductase, and explains their incomplete denitrification phenotype [17].
Thermus amyloliquefaciens strain YIM 77409T was isolated in the course of an investigation of the culturable thermophiles that inhabit geothermal springs in Yunnan Province, southwest China [18]. Strain YIM 77409T was cultured from a sediment sample collected from Niujie Hot Spring using the serial dilution technique on T5 agar. This organism was able to grow anaerobically using nitrate as a terminal electron acceptor, and may potentially impact the nitrogen biogeochemical cycle. Here we describe a summary classification and a set of the features of Thermus amyloliquefaciens type strain YIM 77409T, together with the genome sequence description and annotation. This work may help to better understand the physiological characters as well as the ecological role of this organism in hot spring ecosystems.
Organism information
Classification and features
A taxonomic study using a polyphasic approach placed strain YIM 77409T in the genus Thermus within the family Thermaceae of the phylum Deinococcus-Thermus and resulted in the description of a novel species, Thermus amyloliquefaciens, according to its ability to digest starch [18]. The highest 16S rRNA gene sequence pairwise similarities for strain YIM 77409T were found with the type strain of T. scotoductus SE-1T (97.6 %), T. antranikianii HN3-7T (96.6 %), T. caliditerraeYIM 77925T (96.5 %), and T. tengchongensisYIM 77924T (96.1 %) using EzTaxon-e [19]. The sequence similarities were less than 96.0 % with all other species. Phylogenetic analyses based on the 16S rRNA gene sequences show that YIM 77409T together with T. caliditerrae, T. scotoductus, T. antranikianii, and T. tengchongensis constitute a distinct monophyletic group within the genus Thermus (Fig. 1). The DNA-DNA hybridization value between strains YIM 77409T and T. scotoductus SE-1T was 30.6 ± 1.6 % [18], which was lower than the threshold value (70 %) for the recognition of microbial species [20]. Similarly, the average nucleotide identity (ANI) score between the two strains based on genome-wide comparisons was 86.6 %, according to the algorithm proposed by Goris et al. [21], which is lower than the ANI threshold range (95–96 %) for species demarcation [22]. Those results indicate that strain YIM 77409T represents a distinct genospecies in the genus Thermus [18].
Fig. 1.
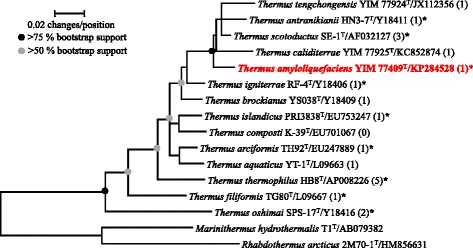
Maximum-likelihood phylogenetic tree of the genus Thermus to highlight the position of Thermus amyloliquefaciens strain YIM 77409T. The tree was reconstructed based on 1374 aligned positions that remained after the application of the Lane mask to the 16S rRNA gene sequences using MEGA 5.0 [54]. Complete deletion of gaps and missing data and Kimura’s two-parameter model was applied. Bootstrap analysis was based on 1000 resamplings. Nodes supported in >75 % (black circles) or >50 % (grey circles) of bootstrap pseudoreplicates (1000 resamplings) for both maximum-likelihood and neighbor-joining methods are indicated. Bar, 0.02 changes per nucleotide. The number of genomes available for each species is included in parentheses (see Table 5) and the asterisk indicates that the genome of the type strain is available. The 16S rRNA gene sequences from Marinithermus hydrothermalis T1T/AB079382 and Rhabdothermus arcticus 2 M70-1T/HM856631 were used as outgroups
Strain YIM 77409T is Gram-negative, facultatively anaerobic, non-motile, and rod shaped (Fig. 2). Cells are 0.4–0.6 μm wide and 1.5–4.5 μm long. Colonies grown on an R2A, T5, and Thermus agar plates for 2 days are yellow and circular. The strain degrades starch and is positive for nitrate reduction. The predominant menaquinone is MK-8. Major fatty acids (>10 %) are iso-C15:0 and iso-C17:0. The polar lipids consist of aminophospholipid, one unidentified phospholipid, and two unidentified glycolipids. Minimum Information about the Genome Sequence [23] of type strain YIM 77409T is provided in Table 1.
Fig. 2.

Scanning electron microscopy image of Thermus amyloliquefaciens strain YIM 77409T grown in Thermus medium broth at 65 °C for 24 h
Table 1.
Classification and general features of Thermus amyloliquefaciens strain YIM 77409T [23]
| MIGS ID | Property | Term | Evidence codea |
|---|---|---|---|
| Classification | Domain Bacteria | TAS [45] | |
| Phylum Deinococcus-Thermus | TAS [46] | ||
| Class Deinococci | TAS [47, 48] | ||
| Order Thermales | TAS [48, 49] | ||
| Family Thermaceae | TAS [48, 50] | ||
| Genus Thermus | TAS [2, 51, 52] | ||
| Species Thermus amyloliquefaciens | TAS [18] | ||
| Type strain: YIM 77409T | TAS [18] | ||
| Gram stain | Negative | TAS [18] | |
| Cell shape | Rod | TAS [18] | |
| Motility | Non-motile | TAS [18] | |
| Sporulation | Nonsporulating | TAS [18] | |
| Temperature range | 50–70 °C | TAS [18] | |
| Optimum temperature | 60–65 °C | TAS [18] | |
| pH range; Optimum | 6.0–8.0; 7.0 | TAS [18] | |
| Carbon source | Glucose, sucrose, glycerol, maltose, raffinose, trehalose, rhamnose, inositol, xylitol, mannitol, sodium malate, mannose and L-arabinose | TAS [18] | |
| MIGS-6 | Habitat | Terrestrial hot springs | TAS [18] |
| MIGS-6.3 | Salinity | Not reported | |
| MIGS-22 | Oxygen requirement | Facultatively anaerobic | TAS [18] |
| MIGS-15 | Biotic relationship | Free-living | TAS [18] |
| MIGS-14 | Pathogenicity | Non-pathogen | NAS |
| MIGS-4 | Geographic location | Niujie hot spring in Eryuan County, Yunnan Province, southwest China | TAS [18] |
| MIGS-5 | Sample collection | 2010 | NAS |
| MIGS-4.1 | Latitude | N 26°15'01. 4" | NAS |
| MIGS-4.2 | Longitude | E 99°59'22. 3" | NAS |
| MIGS-4.4 | Altitude | 2060 m | NAS |
IDA Inferred from Direct Assay, TAS Traceable Author Statement (i.e., a direct report exists in the literature), NAS Non-traceable Author Statement (i.e., not directly observed for the living, isolated sample, but based on a generally accepted property for the species, or anecdotal evidence). These evidence codes are from the Gene Ontology project [53]
a Evidence codes
Genome sequencing information
Genome project history
T. amyloliquefaciens strain YIM 77409T was selected for whole genome sequencing based on its phylogenetic position, denitrifying phenotype, and also for its biotechnological potential. Comparison of the genome of this organism to that of other sequenced Thermus species may provide insights into the molecular basis of the denitrification process in this genus. The genome project for strain YIM 77409T was deposited in the Genomes OnLine Database [24] and the complete sequences were deposited in GenBank. Sequencing, finishing, and annotation were performed by the Department of Energy Joint Genome Institute (Walnut Creek, CA, USA) using state of the art sequencing technology [25]. A summary of the project information associated with MIGS version 2.0 compliance [23] is shown in Table 2.
Table 2.
Project information
| MIGS ID | Property | Term |
|---|---|---|
| MIGS 31 | Finishing quality | Permanent Draft |
| MIGS-28 | Libraries used | PacBio 10 kb |
| MIGS 29 | Sequencing platforms | PacBio RS |
| MIGS 31.2 | Fold coverage | 384.9X PacBio |
| MIGS 30 | Assemblers | HGAP version 2.1.1 |
| MIGS 32 | Gene calling method | Prodigal 2.5; GenePRIMP |
| Locus Tag | BS74 | |
| Genbank ID | JQMV00000000 | |
| GenBank Date of Release | August 28, 2014 | |
| Database: IMG | 2579778517 | |
| GOLD ID | Gp0050852 | |
| BIOPROJECT | PRJNA234787 | |
| MIGS 13 | Source Material Identifier | YIM 77409T |
| Project relevance | Biotechnological |
Growth conditions and genomic DNA preparation
T. amyloliquefaciens type strain YIM 77409T was grown aerobically in Thermus medium at 65 °C for 2 days [18] and DNA was isolated from 0.5–1.0 g of cell pellet using the Joint Genome Institute CTAB bacterial genomic DNA isolation protocol [26].
Genome sequencing and assembly
The draft genome of T. amyloliquefaciens type strain YIM 77409T was generated at the DOE JGI using Pacific Biosciences sequencing technology [27]. A PacBio SMRTbell™ library was constructed and sequenced on the PacBio RS platform using three SMRT cells, which generated 264,235 filtered subreads totaling 751.5 Mbp with an N50 contig length of 2,065,958 bp. All general aspects of library construction and sequencing can be found at the JGI website. All raw reads were assembled using HGAP version 2.1.1 [28]. The final draft assembly produced 6 contigs in 6 scaffolds, totaling 2.16 Mbp in size. The input read coverage was 384.9 × .
Genome annotation
Genes were identified using Prodigal [29] as part of the JGI microbial annotation pipeline [30], followed by a round of manual curation using the JGI GenePRIMP pipeline [31]. The predicted coding sequences were translated and used to search against the Integrated Microbial Genomes non-redundant database, UniProt, TIGRfam, Pfam, PRIAM, KEGG, COG, and InterPro databases. These data sources were combined to assert a product description for each predicted protein. The rRNA genes are predicted using hmmsearch tool from the package HMMER 3.0 [32] and a set of in-house curated HMMs derived from an alignment of full-length rRNA genes selected from IMG isolate genomes; tRNA genes were found using tRNAscan-SE 1.3.1 [33]; other non-coding RNAs and regulatory RNA features were found by searching the genome for the corresponding Rfam profiles using INFERNAL 1.0.2 package [34]. Additional gene prediction analysis and manual functional annotation was performed using the Integrated Microbial Genomes Expert Review platform developed by the JGI [35]. The analysis of the genome presented here and the annotations are for the version available through IMG (2579778517).
Genome properties
The T. amyloliquefaciensYIM 77409T high quality draft genome is 2,160,855 bp long with a 67.4 % G + C content. The genomes comprise 2,257 protein-coding genes and 56 RNA genes. The coding regions accounted for 94 % of the whole genome and 1,839 genes were assigned to a putative function with the remaining annotated as hypothetical proteins. A total of 1,558 genes (67.4 %) were assigned to COGs. The properties and the statistics of the genome are presented in Table 3. The distribution of genes into COG functional categories is presented in Table 4.
Table 3.
Genome statistics
| Attribute | Value | % of Totala |
|---|---|---|
| Genome size (bp) | 2,160,855 | 100.0 |
| DNA coding (bp) | 2,031,100 | 94.0 |
| DNA G + C (bp) | 1,457,281 | 67.4 |
| DNA scaffolds | 6 | 100.0 |
| Total genes | 2,313 | 100.0 |
| Protein coding genes | 2,257 | 97.6 |
| RNA genes | 56 | 2.4 |
| Pseudo genesb | 74 | 3.2 |
| Genes in internal clusters | 1,932 | 83.5 |
| Genes with function prediction | 1,839 | 79.5 |
| Genes assigned to COGs | 1,558 | 67.4 |
| Genes with Pfam domains | 1,842 | 79.6 |
| Genes with signal peptides | 110 | 4.8 |
| Genes with transmembrane helices | 439 | 19.0 |
| CRISPR repeats | 5 |
aThe total is based on either the size of the genome in base pairs or the total number of protein coding genes in the annotated genome
bPseudogenes may also be counted as protein coding or RNA genes, so is not additive under total gene count
Table 4.
Number of genes associated with general COG functional categories
| Code | Value | %age | Description |
|---|---|---|---|
| J | 179 | 10.4 | Translation, ribosomal structure and biogenesis |
| A | 4 | 0.2 | RNA processing and modification |
| K | 76 | 4.4 | Transcription |
| L | 63 | 3.7 | Replication, recombination and repair |
| B | 2 | 0.1 | Chromatin structure and dynamics |
| D | 22 | 1.3 | Cell cycle control, Cell division, chromosome partitioning |
| V | 35 | 2.0 | Defense mechanisms |
| T | 66 | 3.8 | Signal transduction mechanisms |
| M | 77 | 4.5 | Cell wall/membrane biogenesis |
| N | 16 | 0.9 | Cell motility |
| U | 14 | 0.8 | Intracellular trafficking and secretion |
| O | 90 | 5.2 | Posttranslational modification, protein turnover, chaperones |
| C | 131 | 7.6 | Energy production and conversion |
| G | 105 | 6.1 | Carbohydrate transport and metabolism |
| E | 183 | 10.6 | Amino acid transport and metabolism |
| F | 75 | 4.4 | Nucleotide transport and metabolism |
| H | 121 | 7.0 | Coenzyme transport and metabolism |
| I | 91 | 5.3 | Lipid transport and metabolism |
| P | 82 | 4.8 | Inorganic ion transport and metabolism |
| Q | 40 | 2.3 | Secondary metabolites biosynthesis, transport and catabolism |
| R | 170 | 9.9 | General function prediction only |
| S | 68 | 4.0 | Function unknown |
| - | 755 | 32.6 | Not in COGs |
The total is based on the total number of protein coding genes in the genome
Insights from the genome sequence
Comparisons with other Thermus spp. genomes
Twenty-two Thermus genomes from 12 different species have been sequenced, including T. amyloliquefaciens type strain YIM 77409T, and 7 of them have finished genome sequences. The phylogenetic coverage of these genomes is shown in Fig. 1 and their basic properties are summarized in Table 5. The Thermus genomes range in size from 2.04 Mb (Thermus sp. RLM) to 2.56 Mb (T. tengchongensisYIM 77401); GC contents vary from 64.8 % (T. scotoductusDSM 8553T) to 69.5 % (T. thermophilus HB8T), predicted gene number range from 2,043 (T. sp. RLM) to 2,789 (T. brockianus). The genome size (2.16 Mb) and GC contents (67.4 %) of strain YIM 77409T are around the average value, but the gene number of this strain is lower than the average, possibly indicating gene loss through genomic streamlining in this species. In addition, the percentage of protein-coding genes with functional prediction (79.5 %) is higher than the average, whereas the percentage of protein-coding genes with COGs (67.4 %) is similar to the average of the genus Thermus.
Table 5.
Comparison of basic genome features of Thermus strains
| Genome Name | Status | Genome Size (Mb) | GC Content (%) | Gene Count | No. of protein coding genes with function prediction | Percentage (%) | No. of protein coding genes with COGs | Percentage (%) | IMG Genome ID |
|---|---|---|---|---|---|---|---|---|---|
| T. amyloliquefaciens YIM 77409T | Draft | 2.16 | 67.4 | 2313 | 1839 | 79.5 | 1558 | 67.4 | 2579778517 |
| T. scotoductus SA-01 | Finished | 2.36 | 64.9 | 2514 | 1878 | 74.7 | 1704 | 67.8 | 649633105 |
| T. scotoductus KI2 | Draft | 2.48 | 65.5 | 2643 | 2159 | 81.7 | 1808 | 68.4 | 2574179778 |
| T. scotoductus DSM 8553T | Draft | 2.07 | 64.8 | 2305 | 1816 | 78.8 | 1484 | 64.4 | 2518645614 |
| T. antranikianii DSM 12462T | Draft | 2.17 | 64.8 | 2321 | 1939 | 83.5 | 1654 | 71.3 | 2522572193 |
| T. caliditerrae YIM 77777T | Draft | 2.22 | 67.2 | 2327 | 1901 | 81.7 | 1646 | 70.7 | 2582581225 |
| T. tengchongensis YIM 77401 | Draft | 2.56 | 66.4 | 2750 | 2158 | 78.5 | 1818 | 66.1 | 2574179781 |
| T. arciformis CGMCC 1.6992T | Draft | 2.44 | 68.7 | 2672 | 2052 | 76.8 | 1704 | 63.8 | 2617270932 |
| T. thermophilus HB8T | Finished | 2.12 | 69.5 | 2302 | 1498 | 65.1 | 1550 | 67.3 | 637000323 |
| T. thermophilus JL-18 | Finished | 2.31 | 69.0 | 2508 | 1984 | 79.1 | 1717 | 68.5 | 2508501108 |
| T. thermophilus SG0.5JP17-16 | Finished | 2.30 | 68.7 | 2488 | 2024 | 81.4 | 1700 | 68.3 | 2505679077 |
| T. thermophilus HB27 | Finished | 2.13 | 69.4 | 2273 | 1517 | 66.7 | 1562 | 68.7 | 637000322 |
| T. thermophilus ATCC 33923 | Draft | 2.15 | 69.4 | 2366 | 1928 | 81.5 | 1603 | 67.8 | 2554235155 |
| T. islandicus DSM 21543T | Draft | 2.26 | 68.4 | 2470 | 1965 | 79.6 | 1654 | 67.0 | 2524614852 |
| T. oshimai JL-2 | Finished | 2.40 | 68.6 | 2548 | 2018 | 79.2 | 1735 | 68.1 | 2508501045 |
| T. oshimai DSM 12092T | Draft | 2.26 | 68.7 | 2409 | 1960 | 81.4 | 1700 | 70.6 | 2515154080 |
| T. igniterrae ATCC 700962T | Draft | 2.23 | 68.8 | 2379 | 1962 | 82.5 | 1661 | 69.8 | 2515154172 |
| T. aquaticus Y51MC23 | Draft | 2.34 | 68.1 | 2595 | 1740 | 67.1 | 1530 | 59.0 | 645058872 |
| T. brockianus | Draft | 2.48 | 66.8 | 2789 | 2004 | 71.9 | 1709 | 61.3 | 2502171156 |
| T. sp. CCB_US3_UF1 | Finished | 2.26 | 68.6 | 2333 | 1935 | 82.9 | 1655 | 70.9 | 2511231187 |
| T. sp. RLM | Draft | 2.04 | 68.3 | 2043 | 1636 | 80.1 | 1326 | 64.9 | 2513237279 |
| T. sp. NMX2.A1 | Draft | 2.29 | 65.3 | 2522 | 1954 | 77.5 | 1666 | 66.1 | 2514885041 |
Profiles of metabolic network and pathway
The T. amyloliquefaciensYIM 77409T genome encodes genes for complete glycolysis, gluconeogenesis, tricarboxylic acid cycle, pyruvate dehydrogenase, and pentose phosphate pathway. Twenty ABC transporters were identified in the YIM 77409T genome, including amino acid, oligopeptide/dipeptide, N-acetyl-D-glucosamine, maltose, nucleoside, sugar, phosphonate, phosphate, thiamin, cation, and ammonium transporters as well as other permeases. The genome also encodes glucosidases, glycosidases, proteases, and peptidases. The finding of three genes probably coding for esterase (BS74_RS04020, BS74_RS04625, BS74_RS10315) and one gene probably coding for amylopullulanase (BS74_RS00620) are consistent with the observed lipase and amylase activities observed in strain YIM 77409T. A number of genes assigned to a classical electron transport chain have been identified in the strain YIM 77409T genome. Respiratory complex I NADH quinone oxidoreductases consists of NADH quinone oxidoreductase chains A-N (BS74_RS03070-BS74_RS03135), NADH quinone oxidoreductase subunit 15 (BS74_RS02790), and two quinone oxidoreductases (BS74_RS00610, BS74_RS06600). Complex II consists of succinate dehydrogenase (cytochrome b556 subunit SdhC (BS74_RS07950), SdhA (BS74_RS07940), SdhB (BS74_RS07935), and SdhD (BS74_RS07945). A four-subunit cytochrome bc1 complex found in T. thermophilus was also identified in strain YIM 77409T (BS74_RS10415-BS74_RS10430) [36, 37]. The terminal cytochrome oxidase is encoded by four cytochrome c oxidase genes ctaC1 (BS74_RS00820), caaA (BS74_RS00825), ctaD2 (BS74_RS04775), and ctaC2 (BS74_RS04780). Other cytochrome c oxidase genes observed in T. scotoductus SA-01, ctaH, ctaE1, ctaE2, ctaD1, and coxM (TSC_C00960-TSC_C01000), were not found in the YIM 77409T genome.
Genes involved in denitrification
Denitrification is a respiratory process to reduce nitrate or nitrite stepwise to nitrogen gas (NO3− → NO2− → NO → N2O → N2), and plays a major role in converting bioavailable nitrogen to recalcitrant dinitrogen gas [38]. Denitrification normally occurs under oxygen-limiting conditions, and is catalyzed by four types of nitrogen oxide reductases in sequence: nitrate reductase (Nar or Nap), nitrite reductase (Nir), nitric oxide reductase (Nor), and nitrous oxide reductase (Nos) [39, 40]. Previous studies have demonstrated that some Thermus species have incomplete denitrification phenotypes terminating with the production of nitrite or nitrous oxide [16, 41]. This incomplete denitrification is partly encoded by a conjugative element (nitrate conjugative element, NCE) that can be transferred among strains [42]. The NCE is composed of two main operons, nar and nrc, and the transcription factors DnrS and DnrT, which are required for their expression under anaerobic conditions when nitrate is present [43, 44]. The periplasmic nitrate reductase subunits NapB and NapC were not found in the genome of T. amyloliquefaciensYIM 77409T, consistent with the use of the Nar system in the Thermales. Figure 3 shows the organization of the nar operon and neighboring genes involved in denitrification in T. amyloliquefaciensYIM 77409T, T. tengchongensisYIM 77401, and T. scotoductus SA-01. They are located on the chromosome in strains YIM 77409T and YIM 77401, as in T. scotoductus SA-01. However, these gene clusters are located on megaplasmids in T. thermophilus and T. oshimai strains [17]. The nar operons show a high degree of synteny and consist of narCGHJIKT encoding the associated periplasmic cytochrome NarC, the membrane-bound nitrate reductase (NarGHI), the dedicated chaperone NarJ, the nitrate/proton symporter (NarK1), which might also function in nitrite extrusion in T. thermophilus HB8T, and the nitrate/nitrite antiporter (NarK2). Regulatory protein A and a denitrification regulator gene operon dnrST are adjacent to the nar operons. Strain YIM 77409T contains a putative nirS, which encodes the isofunctional tetraheme cytochrome cd1-containing nitrite reductase. The nirK, encoding a Cu-containing nitrite reductase in T. scotoductus SA-01, is absent in strain YIM 77409T and YIM 77401. Genes encoding conserved hypothetical proteins, coenzyme PQQ synthesis protein (PqqE), and nitric oxide reductase subunit b (NorB) and c (NorC) were also presented in the YIM 77409T genome. Genes encoding the periplasmic multicopper enzyme nitrous oxide reductase (Nos), which catalyzes the last step of the denitrification (N2O → N2), were not observed in the YIM 77409T genome or in any Thermus spp. genomes. Physiological experiments with nitrate as the sole terminal electron acceptor also confirm that strain YIM 77409T can convert nitrate to nitrous oxide under anaerobic conditions, but not to nitrogen gas.
Fig. 3.
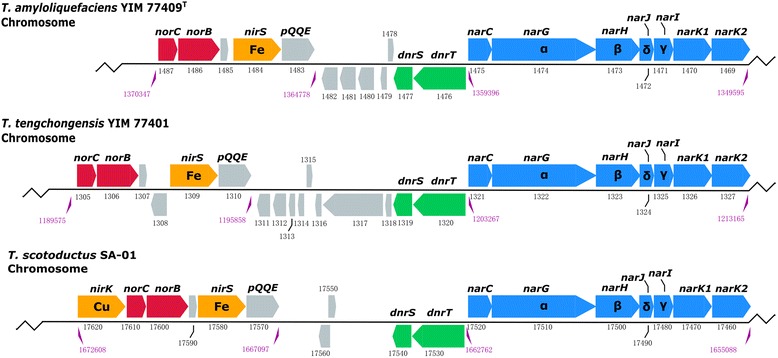
Molecular organization of identified nar operon and neighboring genes involved in denitrification located on the chromosome of T. amyloliquefaciens YIM 77409T, T. tengchongensis YIM 77401, and T. scotoductus SA-01. Fe: heme protein-containing nitrite reductase, Cu: copper-containing nitrite reductase. Numbers below the genes indicate the provisional ORF numbers in T. amyloliquefaciens YIM 77409T and T. tengchongensis YIM 77401, the locations in the chromosome are indicated below. nar: nitrate reductase gene; nir: nitrite reductase gene; nor: nitric oxide reductase gene; dnr: denitrification regulator gene [43, 55, 56]. This figure is modified from Murugapiran et al. [17]
Conclusions
The genus Thermus is the archetypal thermophilic bacterium and has been isolated from both natural and man-made thermal environments. Members of this genus are of significance as a source of thermophilic enzymes of great biotechnological interest and as an excellent laboratory models to study the molecular basis of thermal stability. Here, we report the annotation of a high quality draft genome sequence of Thermus amyloliquefaciensYIM 77409T. Analysis of the genome revealed that strain YIM 77409T encodes enzymes involved in complete glycolysis, pentose phosphate pathway, tricarboxylic acid cycle, pyruvate dehydrogenase, and pentose phosphate pathway. The genome sequence of strain YIM 77409T provides insights to better understand the molecular mechanisms of the incomplete denitrification phenotype and the ecological roles that Thermus species play in nitrogen cycling. Combined analysis of this genome and other Thermus genomes also provides important insights into the evolution and ecology of this group and the role it may play in the high-temperature nitrogen biogeochemical cycle.
Acknowledgements
The work conducted by the U.S. Department of Energy Joint Genome Institute, a DOE Office of Science User Facility, is supported by the Office of Science of the U.S. Department of Energy under Contract No. DE-AC02-05CH11231. Additional support was supported by the Key Project of International Cooperation of Ministry of Science & Technology (MOST) (No. 2013DFA31980), Natural Science Foundation of China (No. 31470139), National Science Foundation grant (OISE-0968421). E-M Zhou received the Scholarship Award for Excellent Doctoral Student granted by Yunnan Province, and China Scholarship Council (CSC, File No. 201307030004). W-J Li was also supported by the Guangdong Province Higher Vocational Colleges & Schools Pearl River Scholar Funded Scheme (2014). B.P. Hedlund was also funded by a gift from Greg Fullmer through the UNLV Foundation.
Abbreviations
- GBS
Great Boiling Spring
- CTAB
cetyl trimethyl ammonium bromide
- PacBio
Pacific Biosciences
- ANI
average nucleotide identity
- NCE
nitrate conjugative element
Footnotes
Competing interests
None of the authors have any competing interests in the manuscript.
Authors’ contributions
WJL and HM supplied the strain. EMZ, CRC, LL, YRY, HM, TTY, and WDX performed the laboratory experiments. MH, AC, MP, KP, NV, NM, DS, TBKR, CYN, CD, NS, VM, NI, AS, NK, and TW were involved in aspects of genome production including sequencing, assembling, annotation and GenBank submission. EMZ, SKM, WJL, and BPH analyzed the genomic data and drafted the manuscript. All authors read and approved the final manuscript.
Contributor Information
Wen-Jun Li, Email: liwenjun3@mail.sysu.edu.cn.
Brian P. Hedlund, Email: brian.hedlund@unlv.edu
References
- 1.Yabe S, Kato A, Hazaka M, Yokota A. Thermaerobacter composti sp. nov., a novel extremely thermophilic bacterium isolated from compost. J Gen Appl Microbiol. 2009;55(5):323–8. doi: 10.2323/jgam.55.323. [DOI] [PubMed] [Google Scholar]
- 2.Brock TD, Freeze H. Thermus aquaticus gen. n. and sp. n., a nonsporulating extreme thermophile. J Bacteriol. 1969;98(1):289–97. doi: 10.1128/jb.98.1.289-297.1969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Kristjánsson JK, Hjörleifsdóttir S, Marteinsson VT, Alfredsson GA. Thermus scotoductus, sp. nov., a pigment-producing thermophilic bacterium from hot tap water in Iceland and including Thermus sp. X-1. Syst Appl Microbiol. 1994;17(1):44–50. doi: 10.1016/S0723-2020(11)80030-5. [DOI] [Google Scholar]
- 4.Yu T-T, Yao J-C, Ming H, Yin Y-R, Zhou E-M, Liu M-J, et al. Thermus tengchongensis sp. nov., isolated from a geothermally heated soil sample in Tengchong, Yunnan, south-west China. Antonie Van Leeuwenhoek. 2013;103(3):513–8. doi: 10.1007/s10482-012-9833-9. [DOI] [PubMed] [Google Scholar]
- 5.Kieft T, Fredrickson J, Onstott T, Gorby Y, Kostandarithes H, Bailey T, et al. Dissimilatory reduction of Fe (III) and other electron acceptors by a Thermus isolate. Appl Environ Microbiol. 1999;65(3):1214–21. doi: 10.1128/aem.65.3.1214-1221.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bruins ME, Janssen AE, Boom RM. Thermozymes and their applications. Appl Biochem Biotechnol. 2001;90(2):155–86. doi: 10.1385/ABAB:90:2:155. [DOI] [PubMed] [Google Scholar]
- 7.Rothschild LJ, Mancinelli RL. Life in extreme environments. Nature. 2001;409(6823):1092–101. doi: 10.1038/35059215. [DOI] [PubMed] [Google Scholar]
- 8.Gihring TM, Banfield JF. Arsenite oxidation and arsenate respiration by a new Thermus isolate. FEMS Microbiol Lett. 2001;204(2):335–40. doi: 10.1111/j.1574-6968.2001.tb10907.x. [DOI] [PubMed] [Google Scholar]
- 9.Balkwill DL, Kieft T, Tsukuda T, Kostandarithes HM, Onstott T, Macnaughton S, et al. Identification of iron-reducing Thermus strains as Thermus scotoductus. Extremophiles. 2004;8(1):37–44. doi: 10.1007/s00792-003-0357-0. [DOI] [PubMed] [Google Scholar]
- 10.Skirnisdottir S, Hreggvidsson GO, Holst O, Kristjansson JK. Isolation and characterization of a mixotrophic sulfur-oxidizing Thermus scotoductus. Extremophiles. 2001;5(1):45–51. doi: 10.1007/s007920000172. [DOI] [PubMed] [Google Scholar]
- 11.Cava F, Zafra O, Da Costa MS, Berenguer J. The role of the nitrate respiration element of Thermus thermophilus in the control and activity of the denitrification apparatus. Environ Microbiol. 2008;10(2):522–33. doi: 10.1111/j.1462-2920.2007.01472.x. [DOI] [PubMed] [Google Scholar]
- 12.Hamilton TL, Koonce E, Howells A, Havig JR, Jewell T, José R, et al. Competition for ammonia influences the structure of chemotrophic communities in geothermal springs. Appl Environ Microbiol. 2014;80(2):653–61. doi: 10.1128/AEM.02577-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Dodsworth JA, Hungate BA, Hedlund BP. Ammonia oxidation, denitrification and dissimilatory nitrate reduction to ammonium in two US Great Basin hot springs with abundant ammonia-oxidizing archaea. Environ Microbiol. 2011;13(8):2371–86. doi: 10.1111/j.1462-2920.2011.02508.x. [DOI] [PubMed] [Google Scholar]
- 14.Dodsworth JA, Hungate B, de la Torre JR, Jiang H, Hedlund BP. Measuring nitrification, denitrification, and related biomarkers in terrestrial geothermal ecosystems. Methods Enzymol. 2011;486:171–203. doi: 10.1016/B978-0-12-381294-0.00008-0. [DOI] [PubMed] [Google Scholar]
- 15.Edwards TA, Calica NA, Huang DA, Manoharan N, Hou W, Huang L, et al. Cultivation and characterization of thermophilic Nitrospira species from geothermal springs in the US Great Basin, China, and Armenia. FEMS Microbiol Ecol. 2013;85(2):283–92. doi: 10.1111/1574-6941.12117. [DOI] [PubMed] [Google Scholar]
- 16.Hedlund BP, McDonald A, Lam J, Dodsworth JA, Brown J, Hungate B. Potential role of Thermus thermophilus and T. oshimai in high rates of nitrous oxide (N2O) production in ~80 °C hot springs in the US Great Basin. Geobiology. 2011;9(6):471–80. doi: 10.1111/j.1472-4669.2011.00295.x. [DOI] [PubMed] [Google Scholar]
- 17.Murugapiran SK, Huntemann M, Wei C-L, Han J, Detter JC, Han C, et al. Thermus oshimai JL-2 and T. thermophilus JL-18 genome analysis illuminates pathways for carbon, nitrogen, and sulfur cycling. Stand Genomic Sci. 2013;7(3):449–68. doi: 10.4056/sigs.3667269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Yu T-T, Ming H, Yao J-C, Zhou E-M, Park D-J, Hozzein WN, et al. Thermus amyloliquefaciens sp. nov., isolated from a hot spring sediment sample. Int J Syst Evol Microbiol. 2015;65(8):2491–5. doi: 10.1099/ijs.0.000289. [DOI] [PubMed] [Google Scholar]
- 19.Kim OS, Cho YJ, Lee K, Yoon SH, Kim M, Na H, et al. Introducing EzTaxon-e: a prokaryotic 16S rRNA gene sequence database with phylotypes that represent uncultured species. Int J Syst Evol Microbiol. 2012;62(Pt 3):716–21. doi: 10.1099/ijs.0.038075-0. [DOI] [PubMed] [Google Scholar]
- 20.Wayne LG, Brenner DJ, Colwell RR, Grimont PAD, Kandler O, Krichevsky MI, et al. Report of the ad hoc Committee on reconciliation of approaches to bacterial systematics. Int J Syst Evol Microbiol. 1987;37(4):463–4. doi: 10.1099/00207713-37-4-463. [DOI] [Google Scholar]
- 21.Goris J, Konstantinidis KT, Klappenbach JA, Coenye T, Vandamme P, Tiedje JM. DNA-DNA hybridization values and their relationship to whole-genome sequence similarities. Int J Syst Evol Microbiol. 2007;57(1):81–91. doi: 10.1099/ijs.0.64483-0. [DOI] [PubMed] [Google Scholar]
- 22.Kim M, Oh H-S, Park S-C, Chun J. Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcation of prokaryotes. Int J Syst Evol Microbiol. 2014;64(Pt 2):346–51. doi: 10.1099/ijs.0.059774-0. [DOI] [PubMed] [Google Scholar]
- 23.Field D, Garrity G, Gray T, Morrison N, Selengut J, Sterk P, et al. The minimum information about a genome sequence (MIGS) specification. Nat Biotechnol. 2008;26(5):541–7. doi: 10.1038/nbt1360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Reddy T, Thomas AD, Stamatis D, Bertsch J, Isbandi M, Jansson J, et al. The Genomes OnLine Database (GOLD) v. 5: a metadata management system based on a four level (meta) genome project classification. Nucleic Acids Res. 2014:gku950. [DOI] [PMC free article] [PubMed]
- 25.Mavromatis K, Land ML, Brettin TS, Quest DJ, Copeland A, Clum A, et al. The fast changing landscape of sequencing technologies and their impact on microbial genome assemblies and annotation. 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.DOE Joint Genome Institute http://my.jgi.doe.gov/general.
- 27.Eid J, Fehr A, Gray J, Luong K, Lyle J, Otto G, et al. Real-time DNA sequencing from single polymerase molecules. Science. 2009;323(5910):133–8. doi: 10.1126/science.1162986. [DOI] [PubMed] [Google Scholar]
- 28.Chin C-S, Alexander DH, Marks P, Klammer AA, Drake J, Heiner C, et al. Nonhybrid, finished microbial genome assemblies from long-read SMRT sequencing data. Nat Methods. 2013;10(6):563–9. doi: 10.1038/nmeth.2474. [DOI] [PubMed] [Google Scholar]
- 29.Hyatt D, Chen G-L, LoCascio PF, Land ML, Larimer FW, Hauser LJ. Prodigal: prokaryotic gene recognition and translation initiation site identification. BMC Bioinformatics. 2010;11(1):119. doi: 10.1186/1471-2105-11-119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Mavromatis K, Ivanova NN, Chen I-MA, Szeto E, Markowitz VM, Kyrpides NC. The DOE-JGI Standard operating procedure for the annotations of microbial genomes. Stand Genomic Sci. 2009;1(1):63. doi: 10.4056/sigs.632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Pati A, Ivanova NN, Mikhailova N, Ovchinnikova G, Hooper SD, Lykidis A, et al. GenePRIMP: a gene prediction improvement pipeline for prokaryotic genomes. Nat Meth. 2010;7(6):455–7. doi: 10.1038/nmeth.1457. [DOI] [PubMed] [Google Scholar]
- 32.Eddy SR. Accelerated profile HMM searches. PLoS Comput Biol. 2011;7(10):e1002195. doi: 10.1371/journal.pcbi.1002195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lowe TM, Eddy SR. tRNAscan-SE: a program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res. 1997;25(5):0955–964. doi: 10.1093/nar/25.5.0955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Nawrocki EP, Kolbe DL, Eddy SR. Infernal 1.0: inference of RNA alignments. Bioinformatics. 2009;25(10):1335–7. doi: 10.1093/bioinformatics/btp157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Markowitz VM, Mavromatis K, Ivanova NN, Chen I-MA, Chu K, Kyrpides NC. IMG ER: a system for microbial genome annotation expert review and curation. Bioinformatics. 2009;25(17):2271–8. doi: 10.1093/bioinformatics/btp393. [DOI] [PubMed] [Google Scholar]
- 36.Mooser D, Maneg O, Corvey C, Steiner T, Malatesta F, Karas M, et al. A four-subunit cytochrome bc1 complex complements the respiratory chain of Thermus thermophilus. Biochimica et Biophysica Acta (BBA)-Bioenergetics. 2005;1708(2):262–74. doi: 10.1016/j.bbabio.2005.03.008. [DOI] [PubMed] [Google Scholar]
- 37.Janzon J, Ludwig B, Malatesta F. Electron transfer kinetics of soluble fragments indicate a direct interaction between complex III and the caa3 oxidase in Thermus thermophilus. Iubmb Life. 2007;59(8–9):563–9. doi: 10.1080/15216540701242482. [DOI] [PubMed] [Google Scholar]
- 38.Knowles R. Denitrification. Microbiol Rev. 1982;46(1):43. doi: 10.1128/mr.46.1.43-70.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Kraft B, Strous M, Tegetmeyer HE. Microbial nitrate respiration-genes, enzymes and environmental distribution. J Biotechnol. 2011;155(1):104–17. doi: 10.1016/j.jbiotec.2010.12.025. [DOI] [PubMed] [Google Scholar]
- 40.Zumft WG. Cell biology and molecular basis of denitrification. Microbiol Mol Biol Rev. 1997;61(4):533–616. doi: 10.1128/mmbr.61.4.533-616.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Ramírez-Arcos S, Fernández-Herrero LA, Berenguer J. A thermophilic nitrate reductase is responsible for the strain specific anaerobic growth of Thermus thermophilus HB8. Biochimica et Biophysica Acta (BBA)-Gene Structure and Expression. 1998;1396(2):215–27. doi: 10.1016/S0167-4781(97)00183-8. [DOI] [PubMed] [Google Scholar]
- 42.Ramírez-Arcos S, Fernández-Herrero LA, Marín I, Berenguer J. Anaerobic growth, a property horizontally transferred by an Hfr-like mechanism among extreme thermophiles. J Bacteriol. 1998;180(12):3137–43. doi: 10.1128/jb.180.12.3137-3143.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Cava F, Zafra O, Magalon A, Blasco F, Berenguer J. A new type of NADH dehydrogenase specific for nitrate respiration in the extreme thermophile Thermus thermophilus. J Biol Chem. 2004;279(44):45369–78. doi: 10.1074/jbc.M404785200. [DOI] [PubMed] [Google Scholar]
- 44.Cava F, Berenguer J. Biochemical and regulatory properties of a respiratory island encoded by a conjugative plasmid in the extreme thermophile Thermus thermophilus. Biochem Soc Trans. 2006;34(1):97–100. doi: 10.1042/BST0340097. [DOI] [PubMed] [Google Scholar]
- 45.Woese CR, Kandler O, Wheelis ML. Towards a natural system of organisms: proposal for the domains Archaea, Bacteria and Eucarya. Proc Natl Acad Sci U S A. 1990;87:4576–9. doi: 10.1073/pnas.87.12.4576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Weisburg W, Giovannoni S, Woese C. The Deinococcus-Thermus phylum and the effect of rRNA composition on phylogenetic tree construction. Syst Appl Microbiol. 1989;11(2):128–34. doi: 10.1016/S0723-2020(89)80051-7. [DOI] [PubMed] [Google Scholar]
- 47.Garrity GM, Holt JG, Class I. Deinococci class. nov. In: Garrity GM, Boone DR, Castenholz RW, editors. Bergey’s Manual of Systematic Bacteriology, Second Edition, Volume 1. New York: Springer; 2001. p. 395. [Google Scholar]
- 48.Validation List No. 85: validation of publication of new names and new combinations previously effectively published outside the IJSEM. Int J Syst Evol Microbiol. 2002;52(3):685–90. [DOI] [PubMed]
- 49.Rainey FA, da Costa MS. Order II. Thermales ord. nov. In: Garrity GM, Boone DR, Castenholz RW, editors. Bergey’s manual of systematic bacteriology, second edition, volume 1. New York: Springer; 2001. p. 403. [Google Scholar]
- 50.da Costa MS, Rainey FA. Family I. Thermaceae fam. nov. In: Garrity GM, Boone DR, Castenholz RW, editors. Bergey’s manual of systematic bacteriology, second edition, volume 1. New York: Springer; 2001. pp. 403–404. [Google Scholar]
- 51.Nobre MF, Trüper HG, da Costa MS. Transfer of Thermus ruber (Loginova et al. 1984), Thermus silvanus (Tenreiro et al. 1995), and Thermus chliarophilus (Tenreiro et al. 1995) to Meiothermus gen. nov. as Meiothermus ruber comb, nov., Meiothermus silvanus comb. nov., and Meiothermus chliarophilus comb. nov., respectively, and emendation of the genus Thermus. Int J Syst Evol Microbiol. 1999;49(4):1951. [Google Scholar]
- 52.Skerman V, McGowan V, Sneath P, Moore W, Moore LV. Approved lists. Int J Syst Bacteriol. 1980;30:225–420. doi: 10.1099/00207713-30-1-225. [DOI] [Google Scholar]
- 53.Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, et al. Gene ontology: tool for the unification of biology. Nat Genet. 2000;25(1):25–9. doi: 10.1038/75556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol. 2011;28(10):2731–9. doi: 10.1093/molbev/msr121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Cava F, Laptenko O, Borukhov S, Chahlafi Z, Blas-Galindo E, Gómez-Puertas P, et al. Control of the respiratory metabolism of Thermus thermophilus by the nitrate respiration conjugative element NCE. Mol Microbiol. 2007;64(3):630–46. doi: 10.1111/j.1365-2958.2007.05687.x. [DOI] [PubMed] [Google Scholar]
- 56.Gounder K, Brzuszkiewicz E, Liesegang H, Wollherr A, Daniel R, Gottschalk G, et al. Sequence of the hyperplastic genome of the naturally competent Thermus scotoductus SA-01. BMC Genomics. 2011;12(1):577. doi: 10.1186/1471-2164-12-577. [DOI] [PMC free article] [PubMed] [Google Scholar]