

Abstract
The essential oil of Daucus carota subsp. carota from Portugal, with high amounts of geranyl acetate (29.0%), α-pinene (27.2%), and 11αH-himachal-4-en-1β-ol (9.2%), was assessed for its biological potential. The antimicrobial activity was evaluated against several Gram-positive and Gram-negative bacteria, yeasts, dermatophytes, and Aspergillus strains. The minimal inhibitory concentration (MIC) and minimal lethal concentration (MLC) were evaluated showing a significant activity towards Gram-positive bacteria (MIC = 0.32–0.64 μL/mL), Cryptococcus neoformans (0.16 μL/mL), and dermatophytes (0.32–0.64 μL/mL). The inhibition of the germ tube formation and the effect of the oil on Candida albicans biofilms were also unveiled. The oil inhibited more than 50% of filamentation at concentrations as low as 0.04 μL/mL (MIC/128) and decreased both biofilm mass and cell viability. The antioxidant capacity of the oil, as assessed by two in chemico methods, was not relevant. Still, it seems to exhibit some anti-inflammatory potential by decreasing nitric oxide production around 20% in LPS-stimulated macrophages, without decreasing macrophages viability. Moreover, the oils safety profile was assessed on keratinocytes, alveolar epithelial cells, macrophages, and hepatocytes. Overall, the oil demonstrated a safety profile at concentrations below 0.64 μL/mL. The present work highlights the bioactive potential of D. carota subsp. carota suggesting its industrial exploitation.
1. Introduction
Aromatic and medicinal plants, such as those found in Lamiaceae and Apiaceae families, have been widely used in folk medicine to treat several ailments. Their effects are particularly associated with the essential oils, which are widely described as having several bioactive properties such as antioxidant, anti-inflammatory, antifungal, and antibacterial ones [1–3].
Plants of the genus Daucus L. (Apiaceae) grow mostly in temperate regions of Europe, West Asia, and Africa. Nevertheless, some species have been found to grow in North America and Australia [4, 5]. The species Daucus carota L., commonly known as carrot, is recognized worldwide due to its roots widely used for both food and medicinal purposes [6]. In addition, the seed essential oil has also been described as antihelmintic, antimicrobial, hypotensive, and diuretic, amongst other biological properties [4].
This taxon includes eleven highly polymorphic, interrelated, and interhybridized taxa [7–9], among which some have been widely studied with regard to their bioactive properties. Nevertheless, only a few studies identify the subspecies used, a very important aspect to consider bearing in mind the high variability mentioned. For example, D. carota subsp. halophilus essential oil has been reported for its antifungal properties against several human pathogenic fungi [7]. In turn, besides the antifungal activities, D. carota subsp. gummifer essential oil has also been described as an anti-inflammatory agent [10] while that of D. carota subsp. maritimus has been pointed out as exhibiting a potential antibacterial effect [11].
Regarding the subspecies D. carota subsp. carota, the antifungal effects of its essential oil were previously reported [12] and although a significant antifungal effect was claimed, the mechanism of action underlying such effects was not assessed. Therefore, in the present study, besides the antifungal effect of the oil against several yeasts (Candida strains, Cryptococcus neoformans), dermatophytes (Trichophyton spp., Epidermophyton, and Microsporum spp.), and Aspergillus strains, we also aim to elucidate a possible mode of action particularly on Candida albicans. For that, the effect of the oil on the inhibition of the germ tube formation, an important virulence factor, as well as the effect of the oil on preformed biofilms, was considered. Additionally, other biological properties of the essential oil were also evaluated, namely, the antibacterial, antioxidant, and anti-inflammatory properties, in order to identify a broader bioactive potential of the oil for its industrial exploitation. Moreover, considering the lack of cytotoxic studies on the essential oil of this subspecies and the putative interest to develop a plant-based product to be used on humans and/or animals, the safety profile of the essential oil against macrophages (Raw 264.7), keratinocytes (HaCaT), epithelial alveolar cells (A549), and hepatocytes (HepG2) was also evaluated.
2. Material and Methods
2.1. Essential Oil Isolation and Analysis
Ripe umbels with seeds of D. carota subsp. carota were collected at Serra da Lousã, Coimbra (Portugal), on the 1st of July 2013. A voucher specimen (Ligia Salgueiro 78) was deposited at the Herbarium of the Faculty of Pharmacy of the University of Coimbra. The essential oil was obtained by hydrodistillation from air dried umbels in a Clevenger-type apparatus according to the European Pharmacopoeia [13]. Oil analyses were carried out by gas chromatography (GC) and gas chromatography/mass spectrometry (GC/MS). GC was carried out on a Hewlett Packard 6890 gas chromatograph (Agilent Technologies, Palo Alto, California, USA) with HP GC ChemStation Rev. A.05.04 data handling system, equipped with a single injector and two flame ionization detectors (FID). A Graphpak divider (Agilent Technologies, part number 5021-7148) was used for simultaneous sampling in two Supelco (Supelco Inc., Bellefonte, PA, USA) fused silica capillary columns with different stationary phases: SPB-1 (polydimethylsiloxane; 30 m × 0.20 mm i.d., film thickness 0.20 μm) and SupelcoWax-10 (polyethylene glycol; 30 m × 0.20 mm i.d., film thickness 0.20 μm). Conditions were as follows: oven temperature program: 70–220°C (3°C/min), 220°C (15 min); injector temperature: 250°C; carrier gas: helium, adjusted to a linear velocity of 30 cm/s; splitting ratio 1 : 40; detectors temperature: 250°C. GC/MS analyses were performed on a Hewlett Packard 6890 gas chromatograph fitted with HP1 fused silica column (polydimethylsiloxane; 30 m × 0.25 mm i.d., film thickness 0.25 μm), interfaced with Hewlett Packard Mass Selective Detector 5973 (Agilent Technologies, Palo Alto, CA, USA) operated by HP Enhanced ChemStation software, version A.03.00. GC parameters were as above; interface temperature was 250°C; MS source temperature was 230°C; MS quadrupole temperature was 150°C; ionization energy was 70 eV; ionization current was 60 μA; scan range was 35–350 μ, with 4.51 scans/s [14]. The volatile compounds were identified by both their retention indices and mass spectra. Retention indices, calculated by linear interpolation relative to retention times of a series of n-alkanes, were compared with those of authenticated samples from the database of the Laboratory of Pharmacognosy of the Faculty of Pharmacy of the University of Coimbra. Mass spectra were compared with reference spectra from a home-made library or from literature data [15, 16]. Relative amounts of individual components were calculated based on GC peak areas without FID response factor correction.
2.2. Antibacterial Assays
The antibacterial activity of the oil was evaluated against Gram-positive strains (Bacillus subtilis ATCC 6633, Listeria monocytogenes CBISA 3183, and Staphylococcus aureus ATCC 6538) and Gram-negative ones (Escherichia coli ATCC 25922 and Salmonella typhimurium ATCC 14028). The minimal inhibitory concentrations (MICs) and the minimum lethal concentrations (MLCs) were assessed according to the Clinical and Laboratory Standards Institute (CLSI) reference protocol M07-A9 [17]. Briefly, serial doubling dilutions of the oil were prepared in dimethyl sulfoxide (DMSO, Sigma Life Science, Sigma-Aldrich, MO, USA) with concentrations ranging from 0.08 to 20 μL/mL. Recent cultures of each strain were used to prepare the cell suspensions (1-2 × 105 CFU/mL) and cell concentration was confirmed by viable count on Mueller Hinton Agar (Oxoid, Hampshire, England). All tests were performed using Mueller Hinton Broth medium and the test tubes were incubated aerobically at 37°C for 24 h and then MICs were registered. To evaluate MLCs, 20 μL of broth was taken from each negative tube after MIC reading, cultured in Mueller Hinton Agar plates, and incubated as mentioned above. The sensitivity of tested strains was controlled by the use of a reference compound, ampicillin (Fluka BioChemika, Buchs, Switzerland). All tests were performed in duplicate. The MIC and MLC values were considered when three independent assays had the same value.
2.3. Antifungal Activity and Mechanism of Action Assays
The antifungal properties of the essential oil were tested against three Candida reference strains (C. albicans ATCC 10231, C. tropicalis ATCC 13803, and C. parapsilosis ATCC 90018) and two clinical strains (C. krusei H9 and C. guilliermondii MAT23); one Cryptococcus neoformans reference strain (C. neoformans CECT 1078); four dermatophyte strains (Trichophyton rubrum CECT 2794, T. mentagrophytes var. interdigitale CECT 2958, T. verrucosum CECT 2992, and Microsporum gypseum CECT 2908); the remaining dermatophytes were clinically isolated (T. mentagrophytes FF7, M. canis FF1, and Epidermophyton floccosum FF9); two reference Aspergillus strains (A. niger ATCC 16404 and A. fumigatus ATCC 46645); and one Aspergillus strain was from a clinical origin (A. flavus F44). The MICs and MLCs were assessed according to the CLSI reference protocols M27-A3 [18] and M38-A2 [19] for yeasts and filamentous fungi, respectively, as previously described by Zuzarte et al. [20].
To elucidate a possible mechanism of action underlying the antifungal effects, two assays were considered: the inhibition of C. albicans germ tube formation and the disruption of its preformed biofilms, in the presence of the essential oil. The first assay was tested as previously reported by Pinto et al. [21]. The percentage of germ tubes was determined as the number of cells showing hyphae at least as long as the diameter of the blastospore. Cells showing a constriction at the point of connection of the hyphae to the mother cell, typical for pseudohyphae, were excluded. Results are shown as mean ± standard deviation of three independent determinations. The effect of the essential oil on preformed C. albicans biofilm was evaluated using the method described by Taweechaisupapong et al. [22] with some modifications. Briefly, a loop of SDA culture of C. albicans grown for 24 h at 37°C was suspended in Yeast Peptone Dextrose (YPD) broth (1% yeast extract, 2% peptone, and 2% dextrose) and incubated for 24 h at 37°C. Then, cells were thoroughly washed twice with sterile PBS (pH 7.4) (0.8% NaCl, 0.02% KH2PO4, 0.31% Na2HPO4·12H2O, and 0.02% KCl). Between each washing step, the suspension was submitted to 10 min centrifugation at 3000 g. Cell density was adjusted to approximately 1 × 106 CFU/mL, using a haemocytometer, and then 100 μL of the final suspension was added to 96-well microtiter plates and incubated for 24 h at 37°C, to form the biofilms. Following three washing steps with PBS, the essential oils (1.25–10 μL/mL, prepared in RPMI) were added and incubated for 24 h, at 37°C. Both negative and positive controls were considered using sterile RPMI broth and inoculated RPMI broth, respectively. Biofilm mass was quantified using crystal violet according to Raut et al. [23]. Biofilm viability was evaluated using the XTT assay, as described by Saharkhiz et al. [24] with some modifications. Briefly, after biofilm formation and treatment with essential oils, the medium was removed and biofilms were thoroughly washed with PBS. To a solution of XTT (1 mg/mL), menadione (10 mM in acetone) was added to a final concentration of 4 μM. 100 μL of this solution was added following incubation for 2 h at 37°C in the dark. The absorbance was observed at 490 nm and biofilm viability was determined by comparing the absorbance of treated samples with those of untreated ones. Results are shown as mean ± standard deviation of three independent determinations performed in duplicate.
2.4. Antioxidant Assays
The antioxidant properties of the essential oil were determined using two different antioxidant assays, namely, the 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS•+) scavenging and oxygen radical absorbance capacity (ORAC) assays. The ABTS•+ scavenging assay was performed according to the procedure described by Re et al. [25], with some modifications. Briefly, the ABTS•+ stock solution was prepared by the reaction of ABTS-NH4 aqueous solution (7 mM) with 2.45 mM dipotassium persulfate in the dark at room temperature for 12–16 h. This solution was then diluted until absorbance of 0.700 ± 0.03 at 734 nm. To determine the scavenging activity, 1 mL of ABTS•+ was added to 100 μL of 0.64–20 mg/mL essential oil solution made in DMSO. After 20 min, the absorbance was read at 734 nm in a spectrophotometer against a blank (absolute ethanol). The antioxidant power of the samples was expressed as IC50 (μg/mL) and compared to that of the standard compound, Trolox (0.75–12 μg/mL). Data are shown as mean values ± standard deviation of three independent assays.
The ORAC assay was carried out using the method described by Garrett et al. [26] slightly modified. Briefly, 150 μL of fluorescein (10 nM) was pipetted to a 96-well plate and 25 μL of Trolox (25–200 μM) or sample (0.32–10 mg/mL in phosphate buffer) was added. This mixture was incubated at 37°C for 10 min. After that, 25 μL of 2,2′-azobis(2-amidino-propane) dihydrochloride (153 mM) was added to each well except that of negative control that contained 25 μL of phosphate buffer. The fluorescence was immediately read on a plate reader every 1 min, in a total of 60 min. The emission wavelength was set at 530/20 nm and excitation wavelength at 485/20 nm. The area under the curve (AUC) was determined as described elsewhere [27]. The results, expressed as Trolox Equivalent (TE)/mg oil, are shown as mean ± standard deviation of at least three independent determinations.
2.5. Anti-Inflammatory Assay
The anti-inflammatory effect of the essential oil was determined through in chemico and in vitro assays using S-nitroso-N-acetyl-D,L-penicillamine (SNAP) as nitric oxide (NO) donor and through evaluation of NO release from lipopolysaccharide- (LPS-) stimulated macrophages, respectively.
For the in chemico assay, several concentrations of the oil (0.08–1.25 μL/mL) were incubated with 0.9 μL of the SNAP solution (100 mM) in endotoxin-free Dulbecco's Modified Eagle Medium (DMEM), in a final volume of 300 μL, for 3 h. The NO scavenging activity was evaluated by quantifying nitrite levels in the medium using the Griess reaction, as previously mentioned [10]. For the in vitro assay, Raw 264.7, a mouse leukaemic macrophage cell line ATCC (TIB-71), was cultured in DMEM supplemented with 10% (v/v) non-inactivated foetal bovine serum, 3.02 g/L sodium bicarbonate, 100 μg/mL streptomycin, and 100 U/mL penicillin at 37°C, in a humidified atmosphere of 95% air and 5% CO2. To evaluate the anti-inflammatory potential of the oil, macrophages (0.3 × 106 cells/well) were cultured in 48-well microplates and allowed to stabilize for 12 h. Following this period, cells were either maintained in culture medium (control) or preincubated with different concentrations of the essential oil for 1 h and later activated with LPS (1 μg/mL) for 24 h. Nitric oxide was quantified by measuring the accumulation of nitrites using the colorimetric Griess assay [28].
Simultaneously, cell viability was also determined using the resazurin method described by Riss et al. [29]. Metabolic active cells reduce resazurin (blue) into resorufin (pink) and therefore the magnitude of dye reduction is correlated with the number of viable cells. After the treatment described above for macrophages, resazurin solution (0.125 mg/mL) was added (1 : 10) and cells were further incubated at 37°C for 30 min in a humidified atmosphere of 95% air and 5% CO2. Quantification was performed using an ELISA microplate reader (SLT, Austria) at 570 nm, with a reference wavelength of 620 nm. A cell-free control was performed in order to exclude nonspecific effects of the oils on resazurin (data not shown).
2.6. Toxicological Profile
Cytotoxicity was evaluated in several mammalian cell lines, namely, human hepatocellular carcinoma cell line HepG2, ATCC number 77400; human keratinocyte cell line HaCaT, obtained from DKFZ (Heidelberg); human alveolar epithelial cell line A549, ATCC number CCL-185; and the mouse leukaemic monocyte macrophage cell line, previously mentioned.
Briefly, Raw 264.7 (0.6 × 106 cells/mL), HepG2 (0.5 × 106 cells/mL), HaCaT (0.2 × 106 cells/mL), and A549 (0.2 × 106 cells/mL) cell suspensions were prepared. Then, cells were cultured in 48-well microplates in a final volume of 600 μL for 12 h and were further cultured with different concentrations (0.08 to 1.25 μL/mL) of the essential oil, for 24 h. At the end, 60 μL of resazurin (0.125 mg/mL) was added and the plates were then incubated for 30 min (Raw 264.7), 60 min (HepG2 and A549), and 120 min (HaCaT) at 37°C, in a humidified atmosphere of 95% air and 5% CO2. Cell viability was determined by reading the absorbance at 570 nm with a reference filter at 620 nm against a negative control (cells cultured in the absence of the oil) in an ELISA microplate reader (SLT, Austria). A cell-free control was performed in order to exclude unspecific effects of the oil on resazurin (data not shown).
2.7. Statistical Analysis
Data are expressed as mean ± standard error of the mean (SEM). Statistical significance was determined using one-way analysis of variance (ANOVA), followed by Dunnett's post hoc test. The statistical analysis was performed using Prism 5.0 Software (GraphPad Software). Differences were considered significant for p < 0.05.
3. Results and Discussion
3.1. Chemical Composition
The essential oil of D. carota subsp. carota was obtained from the umbels with a yield of 0.9% (v/w). Constituents of the oil are listed in Table 1, according to their elution order on a polydimethylsiloxane column. The oil is predominantly composed of hydrocarbon monoterpenes (46.6%) and oxygenated monoterpenes (29.5%), with geranyl acetate (29.0%) and α-pinene (27.2%) being the main components. Notably, these compounds were also identified as the main constituents of the essential oils obtained from flowering umbels of the same species grown in another region of Portugal (Cantanhede) [12], despite quantitative differences (37.9% for α-pinene and 15.0% for geranyl acetate). In turn, in opposition to that study, the D. carota subsp. carota oil herein obtained had a significant amount of oxygen containing sesquiterpenes (15.6% versus 2.5–3.1%), with 11αH-himachal-4-en-1β-ol being the main compound. This constituent was also identified as one of the main compounds in D. carota subsp. carota oil from plants of Italian origin [12].
Table 1.
Composition of the essential oil of Daucus carota subsp. carota.
| RIa | RIp | Compounds∗ | % |
|---|---|---|---|
| 922 | 1030 | α-Thujene | t |
| 930 | 1030 | α-Pinene | 27.2 |
| 943 | 1073 | Camphene | 0.9 |
| 964 | 1128 | Sabinene | 0.1 |
| 970 | 1118 | β-Pinene | 4.5 |
| 980 | 1161 | Myrcene | 2.5 |
| 1006 | 1185 | α-Terpinene | t |
| 1013 | 1272 | p-Cymene | 0.1 |
| 1020 | 1206 | Limonene | 9.0 |
| 1025 | 1235 | Z-β-Ocimene | 0.4 |
| 1035 | 1250 | E-β-Ocimene | 0.4 |
| 1047 | 1250 | γ-Terpinene | 1.4 |
| 1081 | 1543 | Linalool | t |
| 1158 | 1595 | Terpinen-4-ol | 0.1 |
| 1176 | 1699 | Verbenone | 0.1 |
| 1233 | 1838 | Geraniol | 0.1 |
| 1266 | 1574 | Bornyl acetate | 0.1 |
| 1345 | 1466 | α-Longipene | 1.0 |
| 1362 | 1753 | Geranyl acetate | 29.0 |
| 1411 | 1590 | E-β-Caryophyllene | 0.4 |
| 1443 | 1660 | α-Humulene | 0.4 |
| 1459 | 2172 | (E)-Methyl isoeugenol | 1.4 |
| 1466 | 1699 | Germacrene D | 0.1 |
| 1488 | 1699 | β-Himachalene | 1.3 |
| 1498 | 1720 | β-Bisabolene | 0.3 |
| 1557 | 1968 | Caryophyllene oxide | 0.2 |
| 1581 | 2001 | Carotol | 6.2 |
| 1623 | 2089 | 11αH-Himachal-4-en-1β-ol | 9.2 |
|
| |||
| Monoterpene hydrocarbons | 46.6 | ||
| Oxygen containing monoterpenes | 29.5 | ||
| Sesquiterpene hydrocarbons | 3.5 | ||
| Oxygen containing sesquiterpenes | 15.6 | ||
| Others | 1.4 | ||
|
| |||
| Total | 96.6 | ||
∗Compounds listed in order to their elution on the SPB-1 column.
t: traces (≤0.05%).
RIa: retention indices on the SPB-1 column relative to C8 to C24 n-alkanes.
RIp: retention indices on the SupelcoWax-10 column relative to C8 to C24 n-alkanes.
3.2. Antibacterial Activity
The antibacterial potential of the oil against both Gram-positive strains (Bacillus subtilis, Listeria monocytogenes, and Staphylococcus aureus) and Gram-negative ones (Escherichia coli and Salmonella typhimurium) is summarized in Table 2. The results show that the oil was significantly more effective against Gram-positive bacteria, with MIC values in the range of 0.32–0.64 μL/mL. Differences observed between Gram-positive and Gram-negative bacteria are mainly due to their distinct cell wall structure, as the cell wall of Gram-negative bacteria is much more complex comprising an outer membrane composed of hydrophilic polysaccharides chains that act as a barrier for hydrophobic essential oils [30].
Table 2.
Antibacterial activity (MIC and MLC) of D. carota subsp. carota essential oil.
| Strains | Essential oil | Ampicillin | ||
|---|---|---|---|---|
| MICa | MLCa | MICb | MLCb | |
| Gram-positive | ||||
| Bacillus subtilis ATCC 6633 | 0.32 | 0.64 | 0.06 | 0.025 |
| Listeria monocytogenes CBISA 3183 | 0.64 | >10 | 2 | 16 |
| Staphylococcus aureus ATCC 6538 | 0.32 | 0.64 | 0.25 | 0.5 |
| Gram-negative | ||||
| Escherichia coli ATCC 25922 | >10 | >10 | 8 | 16 |
| Salmonella typhimurium ATCC 14028 | >10 | >10 | 4 | 8 |
MIC and MLC were determined by a macrodilution method and expressed in a μL/mL and in b μg/mL.
Results were obtained from three independent experiments performed in duplicate.
Previously, the antibacterial activity of essential oils from the herb, flowering, and mature umbels of wild carrot growing in Poland was also tested [9]. Although direct comparisons between that study and the present one cannot be considered since a different antibacterial test was used (agar dilution method versus macrodilution broth method), the oils obtained in the previous work were much less effective against Gram-positive bacteria (MIC = 3–5 μL/mL). These differences might be explained by distinct chemical compositions (α-pinene and sabinene versus α-pinene and geranyl acetate), as it is known that sabinene is devoid of antibacterial activity [31]. Instead, the essential oil herein used was primarily rich in geranyl acetate and α-pinene. These compounds have been tested for their antibacterial potential and several studies have pointed out the high antibacterial activity of α-pinene [32, 33] and weak activity of geranyl acetate [30], which may justify the activity of the oil. Nevertheless, minor compounds may also interfere with the antibacterial activity, and their potential effect should not be discarded.
3.3. Antifungal Activity and Mechanisms of Action
The antifungal activity of the essential oil against human and animal pathogens is presented in Table 3. In general, the oil was more effective against Cryptococcus neoformans (MIC = 0.16 μL/mL) and dermatophyte strains, with MICs ranging from 0.32 to 0.64 μL/mL. Regardless of the oil being much less effective against Candida spp. and Aspergillus spp., it showed a very low MIC for C. guilliermondii, similar to that found for dermatophytes (0.32 μL/mL), thus suggesting some specificity of the oil for this strain. Overall, the oil showed both fungistatic and fungicidal effects against most of the strains tested since the MIC values were similar to MLC ones. Of note is the fact that the main isolated compounds identified in the oil herein studied, namely, geranyl acetate, α-pinene, and limonene, have also been previously assessed for their antifungal potential. Geranyl acetate demonstrated good antifungal effects against dermatophytes and Cryptococcus neoformans; however, it had a weak performance in inhibiting the growth of Candida strains and Aspergillus spp. [2, 21]. Similarly, α-pinene showed inhibitory effects against C. albicans and Cryptococcus neoformans [34, 35] as well as a potent effect against dermatophyte strains [36]. Moreover, Pinto et al. [21] also demonstrated that this compound exhibits a strong fungistatic and fungicidal activity, with this effect being preeminent for Candida and Aspergillus spp. Several authors have also described the antifungal activity of limonene against several fungi strains [36–39]. Therefore, the activity of these major compounds of D. carota subsp. carota essential oil may be responsible for the higher antifungal effects of this oil.
Table 3.
Antifungal activity (MIC and MLC) of Daucus carota subsp. carota essential oil for Candida spp., Cryptococcus neoformans, dermatophyte, and Aspergillus strains.
| Strains | Essential oil | Fluconazole | Amphotericin | |||
|---|---|---|---|---|---|---|
| MICa | MLCa | MICb | MLCb | MICb | MLCb | |
| Candida albicans ATCC 10231 | 5 | 5 | 1 | >128 | NT | NT |
| Candida guilliermondii MAT23 | 0.32 | 0.32 | 8 | 8 | NT | NT |
| Candida krusei H9 | 5 | 5 | 64 | 64–128 | NT | NT |
| Candida parapsilosis ATCC 90018 | 10 | >10 | <1 | <1 | NT | NT |
| Candida tropicalis ATCC 13803 | 5 | >10 | 4 | >128 | NT | NT |
| Cryptococcus neoformans CECT 1078 | 0.16 | 0.16 | 16 | 128 | NT | NT |
| Epidermophyton floccosum FF9 | 0.32 | 0.32 | 16 | 16 | NT | NT |
| Microsporum canis FF1 | 0.64 | 0.64 | 128 | 128 | NT | NT |
| Microsporum gypseum CECT 2908 | 0.64 | 0.64 | 128 | >128 | NT | NT |
| Trichophyton mentagrophytes FF7 | 0.64 | 0.64 | 16–32 | 32–64 | NT | NT |
| Trichophyton mentagrophytes var. interdigitale CECT 2958 | 0.64 | 1.25 | 128 | ≥128 | NT | NT |
| Trichophyton rubrum CECT 2794 | 0.32 | 0.32 | 16 | 64 | NT | NT |
| Trichophyton verrucosum CECT 2992 | 0.64 | 0.64 | >128 | >128 | NT | NT |
| Aspergillus flavus F44 | >10 | >10 | NT | NT | 2 | 8 |
| Aspergillus fumigatus ATCC 46645 | 2.5 | >10 | NT | NT | 2 | 4 |
| Aspergillus niger ATCC 16404 | 1.25 | >10 | NT | NT | 1-2 | 4 |
MIC and MLC were determined by a macrodilution method and expressed in a µL/mL and in b µg/mL.
Results were obtained from three independent determinations performed in duplicate.
Although studies on the antifungal activity of D. carota subsp. carota oil were previously carried out, the mechanism of action underlying this effect remains unknown. Therefore, in the present study, we attempt to elucidate possible modes of action on C. albicans. For that, two assays were selected, namely, the inhibition of germ tube formation and the disruption of preformed biofilms.
The effects of subinhibitory concentrations of the essential oil on the inhibition of C. albicans germ tube formation are presented in Table 4. The oil was able to achieve more than 50% of filamentation inhibition at concentrations as low as 0.04 μL/mL (MIC/128). This is quite interesting, since filamentation (dimorphic transition from yeast to filamentous form) in C. albicans is essential for virulence [40] and it seems that filamentation inhibition per se is sufficient to treat disseminated candidosis [41]. The striking difference between MICs and filamentation-inhibiting concentrations seems to suggest that different mechanisms of action may be responsible for these two biological effects. Geranyl acetate, the major compound of D. carota subsp. carota oil, may be responsible for this activity as assessed by Zore et al. [42]. This compound was highly effective against serum-induced morphogenesis (yeast to hyphal form transition in C. albicans ATCC 10231) with only 73 μg/mL causing 63% inhibition of germ tube induction [42].
Table 4.
Influence of subinhibitory concentrations of the essential oil of Daucus carota subsp. carota on germ tube formation of C. albicans ATCC 10231.
| Essential oil concentration | Candida albicans ATCC 10231 |
|---|---|
| (µL/mL) | (% of filamentous cells) |
| 0.00 (control)a | 100.00 ± 0.00 |
| 5.00 (MIC) | 0.00 ± 0.00 |
| 2.50 (MIC/2) | 0.59 ± 1.0 |
| 1.25 (MIC/4) | 0.88 ± 1.54 |
| 0.64 (MIC/8) | 1.63 ± 2.82 |
| 0.32 (MIC/16) | 2.52 ± 4.36 |
| 0.16 (MIC/32) | 2.90 ± 1.25 |
| 0.08 (MIC/64) | 21.49 ± 10.89 |
| 0.04 (MIC/128) | 44.44 ± 8.60 |
| 0.02 (MIC/256) | 68.54 ± 5.09 |
aSamples with 1% (v/v) DMSO.
Figures 1 and 2 represent the effect of the essential oil on preformed C. albicans biofilms. The crystal violet method quantifies the biomass of the biofilm by staining it with the dye whereas the XTT assay evaluates cell viability by analysing the formation of a water soluble crystal formed after mitochondrial metabolization. Results show that the oil promoted a decrease of the biofilm biomass even for the lowest concentrations tested (Figure 1). Therefore, the results showed that the oil was able to interfere with preformed biofilms by reducing the amount of the attached biomass. Regarding biofilm cells viability, concentrations higher than 1.25 μL/mL also reduced cell viability (Figure 2), compromising biofilm development. Note that the biofilm formation is a survival mechanism, contributing to microbial virulence and persistence [43, 44] since biofilms are very difficult to eliminate due to their high antifungal resistance in comparison to free-living cells. These results highlight the promising antibiofilm activity paving the way for future translational research on the treatment of disseminative candidiasis.
Figure 1.
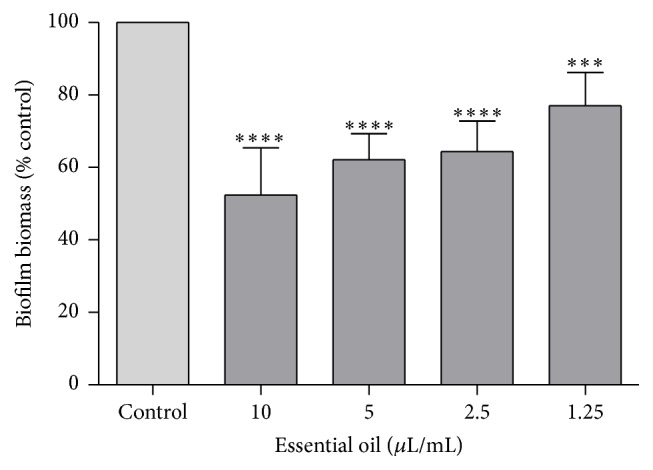
Biofilm biomass after treatment with D. carota subsp. carota essential oil, using the crystal violet assay. Biofilm biomass was determined using the formula (Abs620 sample/Abs620 control) ∗ 100. Results are shown as mean ± standard deviation of at least three independent determinations carried out in duplicate. ∗∗∗ p < 0.001, ∗∗∗∗ p < 0.0001, compared to control using one-way ANOVA followed by Dunnett's multiple comparison test. Control (100%) corresponds to an absorbance mean value of 1.587.
Figure 2.
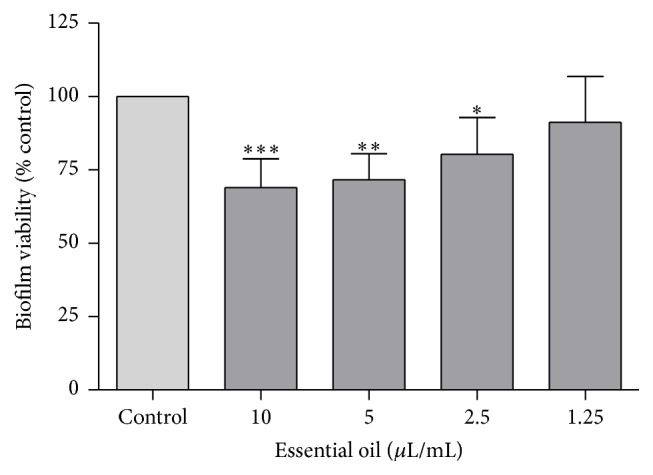
Biofilm viability after treatment with D. carota subsp. carota essential oil using the XTT viability assay. Results are shown as mean ± standard deviation of at least three independent determinations carried out in duplicate. ∗ p < 0.05, ∗∗ p < 0.01, and ∗∗∗ p < 0.001, compared to control using one-way ANOVA followed by Dunnett's multiple comparison test. Control (100%) corresponds to an absorbance mean value of 0.621.
3.4. Antioxidant Analysis
The antioxidant analysis of the essential oil was carried out using the ABTS•+ scavenging and ORAC assays. Table 5 summarizes the results obtained. It was seen that the essential oil is neither a good scavenger of ABTS•+ (IC50 = 1924.25 μg/mL) nor a good peroxyl-induced oxidation inhibitor (ORAC values of 7.13 μmol/TE/mg oil). Comparison of the present results with others for the same plant species is not possible due to the absence of the latter.
Table 5.
Antioxidant analysis of D. carota subsp. carota essential oil.
| Sample | ABTS•+a | ORACb |
|---|---|---|
| Essential oil | 1924.25 | 7.13 |
| Trolox | 5.53 | — |
aValues expressed as IC50 (µg/mL).
bValues expressed as µmol TE/mg.
3.5. Anti-Inflammatory Activity
Chemical NO scavenging is a method possessing two valences; that is, it allows to evaluate the antioxidant potential of the essential oil by testing its ability to arrest this radical but also allows preliminary screening of the anti-inflammatory potential, since NO is a crucial mediator in inflammation. Figure 3 summarizes the NO scavenging activity of the essential oil. The results showed that the essential oil had no scavenging activity towards NO for all the tested concentrations (0.08–1.25 μL/mL). In order to deeply explore whether the essential oil modulates NO production, we also used an in vitro model of inflammation consisting of macrophages stimulated with Toll-like receptor 4 agonist LPS. Figures 4(a) and 4(b) summarize the NO release and the cell viability of LPS-stimulated macrophages treated with different concentrations of the essential oil, respectively. As far as we know, this is the first report on the anti-inflammatory activity of D. carota subsp. carota. As shown in Figure 4(a), incubation of macrophages with LPS, for 24 h, resulted in a significant increase in nitrite production. Taking into account the toxicity of the oil presented in Figure 4(b), inhibition of NO production was only considered for nontoxic concentrations of the oil. Indeed, NO production decreased by 19.04%, relatively to LPS (p < 0.05), without affecting cell viability in the presence of 0.64 μL/mL of the oil. These results suggest a potential anti-inflammatory effect of the oil. Nevertheless, further experiments on different proinflammatory mediators and signal transduction pathways should be considered to confirm this activity.
Figure 3.
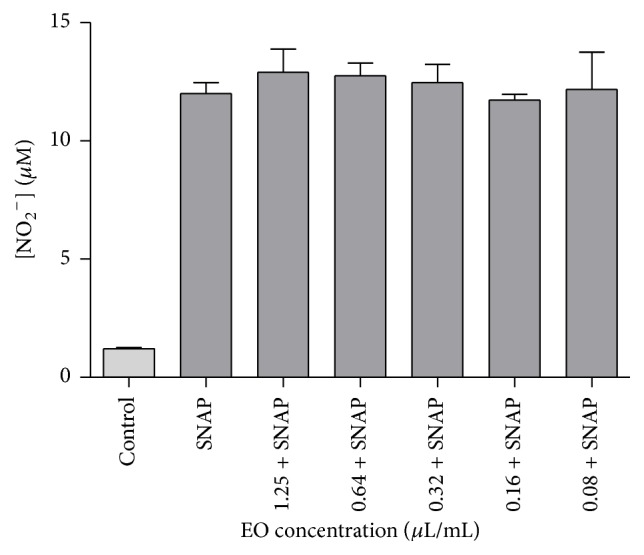
NO scavenging activity of Daucus carota subsp. carota essential oil. Different concentrations of essential oil (1.25–0.08 μL/mL) were incubated with the NO donor, SNAP (100 mM), in culture medium for 3 h. Results are shown as mean ± SEM of three independent assays, done in duplicate.
Figure 4.

Anti-inflammatory effect of Daucus carota subsp. carota in LPS-stimulated Raw 264.7 macrophages: (a) NO production and (b) cell viability. Macrophages were treated with essential oil (1.25–0.08 μL/mL) for 1 h prior to LPS (1 μg/mL) activation and further incubated for 24 h. NO release was determined in the supernatants of the cultures using the Griess reagent (a) and cell viability was assessed on adherent cells using the resazurin reagent and expressed as percentage of cell viability by control cells (b). Results are shown as mean ± SEM of at least three independent assays. (∗ p < 0.05; ∗∗ p < 0.01; ∗∗∗∗ p < 0.0001, compared to LPS). Cell viability control (100%) corresponds to an absorbance mean value of 0.435.
The essential oil's major compounds, namely, geranyl acetate and α-pinene, may account for most of the oil's anti-inflammatory potential since previous studies have pointed out their anti-inflammatory potential (e.g., [45–47]).
3.6. Toxicological Profile
The cytotoxicity of the essential oil was screened in several mammalian cells lines in order to evaluate a potential pharmacological application of D. carota subsp. carota essential oil and the gathered results are summarized in Table 6. It can be inferred that the concentration of 0.64 μL/mL induces different cell viability results among all the cell lines studied, with macrophages being the most resilient (92.83% ± 1.04 cell viability) and hepatocytes the most susceptible (60.73% ± 6.51 cell viability). On the other hand, it is possible to conclude that concentrations below 0.64 μL/mL are devoid of toxicity, presenting a safety profile for most of the cells studied. Lower concentrations of the oil trigger an increase in resazurin reduction, which may suggest augmentation of the metabolic activity of the cells or a rise in cell proliferation. Further studies should be done to further explore these results. It is, however, important to emphasize that no studies have been previously conducted regarding the cytotoxic effect of D. carota subsp. carota essential oil. Nevertheless, our group has previously reported that geranyl acetate has very detrimental cytotoxic effects [2].
Table 6.
Effect of Daucus carota subsp. carota essential oil on cell lines viability.
| Essential oil | Macrophages | Epithelial alveolar | Hepatocytes | Keratinocytes |
|---|---|---|---|---|
| (µL/mL) | Raw 264.7 (%) | cells A549 (%) | HepG2 (%) | HaCaT (%) |
| 0.00 (control) | 100 ± 0.0 | 100 ± 0.0 | 100 ± 0.0 | 100 ± 0.0 |
| 1.25 | 9.01 ± 9.01∗∗∗ | 64.25 ± 4.66∗∗ | 34.54 ± 4.92∗∗∗∗ | 55.76 ± 5.03∗∗∗∗ |
| 0.64 | 92.83 ± 1.04 | 86.25 ± 5.78 | 60.73 ± 6.51∗∗∗ | 76.30 ± 0.54∗∗∗∗ |
| 0.32 | 123.60 ± 15.28 | 110.60 ± 5.72 | 99.40 ± 5.49 | 85.21 ± 2.35∗∗ |
| 0.16 | 141.50 ± 14.56∗ | 130.80 ± 9.96∗ | 108.80 ± 4.81 | 94.44 ± 2.94 |
| 0.08 | 154.60 ± 15.55∗∗ | 201.90 ± 19.43∗∗∗∗ | 122.60 ± 10.43∗ | 104.23 ± 2.10 |
Results expressed as percentage of resazurin reduction compared to control cells maintained in culture medium. Each value represents mean ± SEM of at least three independent experiments done in duplicate. Statistical differences compared to control cells (∗ p < 0.05, ∗∗ p < 0.01, ∗∗∗ p < 0.001, and ∗∗∗∗ p < 0.0001 using one-way ANOVA followed by Dunnett's multiple comparison test).
4. Conclusions
This study allowed a better understanding of the bioactivities of D. carota subsp. carota essential oil. The results showed that this oil had a significant activity towards the inhibition of Gram-positive bacteria, Cryptococcus neoformans, and dermatophytes. Importantly, the oil was also efficient in inhibiting the germ tube formation and the preformed biofilms of Candida albicans. Despite the oil exhibiting no considerable antiradical activity, it reduced about 20% NO release in LPS-stimulated macrophages, at concentrations devoid of toxicity to these cells. It is reasonable to conclude that concentrations lower than 0.64 μL/mL present a safety profile for different human cell types unveiling the potential application of the essential oil for therapeutical purposes, with a special focus on fungal infections associated with a proinflammatory status. Further experiments disclosing the mechanism of action and in vivo tests are of utmost importance to further support the benefit and safety of D. carota subsp. carota essential oil.
Acknowledgments
The authors thank Otília Vieira (Center for Neuroscience and Cell Biology, University of Coimbra, Portugal) for providing the Raw 264.7 cell line, Eugénia Carvalho (Centre for Neuroscience and Cell Biology, University of Coimbra, Portugal) for the kind gift of the HaCat cell line, and Conceição Pedroso Lima (Centre for Neuroscience and Cell Biology, University of Coimbra, Portugal) for the HepG2 cell line.
Conflict of Interests
The authors declare that there is no conflict of interests regarding the publication of this paper.
References
- 1.Bakkali F., Averbeck S., Averbeck D., Idaomar M. Biological effects of essential oils—a review. Food and Chemical Toxicology. 2008;46(2):446–475. doi: 10.1016/j.fct.2007.09.106. [DOI] [PubMed] [Google Scholar]
- 2.Gonçalves M. J., Cruz M. T., Tavares A. C., et al. Composition and biological activity of the essential oil from Thapsia minor, a new source of geranyl acetate. Industrial Crops and Products. 2012;35(1):166–171. doi: 10.1016/j.indcrop.2011.06.030. [DOI] [Google Scholar]
- 3.Chalchat J. C., Chiron F., Garry R. P., Lacoste J., Sautou Y. Photochemical hydroperoxidation of terpenes. Antimicrobial activity of α-pinene, β-pinene and limonene hydroperoxides. Journal of Essential Oil Research. 2000;12(1):125–134. doi: 10.1080/10412905.2000.9712059. [DOI] [Google Scholar]
- 4.Soković M., Stojković D., Glamočlija J., Ćirić A., Ristić M., Grubišić D. Susceptibility of pathogenic bacteria and fungi to essential oils of wild Daucus carota . Pharmaceutical Biology. 2009;47(1):38–43. doi: 10.1080/13880200802400535. [DOI] [Google Scholar]
- 5.Guinoiseau E., Luciani A., Casanova J., Tomi F., Bolla J. M., Berti L. Daucus carota L.: a common plant with a potentially large medicinal application-field. In: Govil J. N., Singh V. K., editors. Drug Plants III. Vol. 29. Houston, Tex, USA: Studium Press LLC; 2010. pp. 385–411. (Recent Progress in Medicinal Plants). [Google Scholar]
- 6.da Silva Dias J. C. Nutritional and health benefits of carrots and their seed extracts. Food and Nutrition Sciences. 2014;5(22):2147–2156. doi: 10.4236/fns.2014.522227. [DOI] [Google Scholar]
- 7.Tavares A. C., Gonçalves M. J., Cavaleiro C., et al. Essential oil of Daucus carota subsp. halophilus: composition, antifungal activity and cytotoxicity. Journal of Ethnopharmacology. 2008;119(1):129–134. doi: 10.1016/j.jep.2008.06.012. [DOI] [PubMed] [Google Scholar]
- 8.Chizzola R. Composition of the essential oil from Daucus carota ssp. carota growing wild in Vienna. Journal of Essential Oil-Bearing Plants. 2010;13(1):12–19. doi: 10.1080/0972060x.2010.10643785. [DOI] [Google Scholar]
- 9.Staniszewska M., Kula J., Wieczorkiewicz M., Kusewicz D. Essential oils of wild and cultivated carrots—the chemical composition and antimicrobial activity. Journal of Essential Oil Research. 2005;17(5):579–583. doi: 10.1080/10412905.2005.9699002. [DOI] [Google Scholar]
- 10.Valente J., Zuzarte M., Resende R., et al. Daucus carota subsp. gummifer essential oil as a natural source of antifungal and anti-inflammatory drugs. Industrial Crops and Products. 2015;65:361–366. doi: 10.1016/j.indcrop.2014.11.014. [DOI] [Google Scholar]
- 11.Jabrane A., Jannet H. B., Harzallah-Skhiri F., Mastouri M., Casanova J., Mighri Z. Flower and root oils of the Tunisian Daucus carota L. ssp. maritimus (Apiaceae): integrated analyses by GC, GC/MS, and 13C-NMR spectroscopy, and in vitro antibacterial activity. Chemistry and Biodiversity. 2009;6(6):881–889. doi: 10.1002/cbdv.200800144. [DOI] [PubMed] [Google Scholar]
- 12.Maxia A., Marongiu B., Piras A., et al. Chemical characterization and biological activity of essential oils from Daucus carota L. subsp. carota growing wild on the Mediterranean coast and on the Atlantic coast. Fitoterapia. 2009;80(1):57–61. doi: 10.1016/j.fitote.2008.09.008. [DOI] [PubMed] [Google Scholar]
- 13.Council of Europe. European Pharmacopoeia. 7th. Strasbourg, France: Directorate for the Quality of Medicines & HealthCare of the Council of Europe; 2010. [Google Scholar]
- 14.Cavaleiro C., Salgueiro L. R., Miguel M. G., Proença da Cunha A. Analysis by gas chromatography-mass spectrometry of the volatile components of Teucrium lusitanicum and Teucrium algarbiensis . Journal of Chromatography A. 2004;1033(1):187–190. doi: 10.1016/j.chroma.2004.01.005. [DOI] [PubMed] [Google Scholar]
- 15.Adams R. P. Identification of Essential Oil Components by Gas Chromatography/Quadrupole Mass Spectroscopy. 4th. Carol Stream, Ill, USA: Allured; 2007. [Google Scholar]
- 16.Joulain D., König W. A. The Atlas of Spectral Data of Sesquiterpene Hydrocarbons. Hamburg, Germany: E.B.-Verlag; 1998. https://books.google.pt/books?id=j3RgQgAACAAJ. [Google Scholar]
- 17.CLSI. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically. 9th. Wayne, Pa, USA: Clinical & Laboratory Standards Institute; 2012. (Standard M07-A9). [Google Scholar]
- 18.CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts; Approved Standard M27-A3. 3rd. Wayne, Pa, USA: Clinical and Laboratory Standards Institute (CLSI); 2008. [Google Scholar]
- 19.CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi; Approved Standard M38-A2. 2nd. Wayne, Pa, USA: Clinical and Laboratory Standards Institute (CLSI); 2008. [Google Scholar]
- 20.Zuzarte M., Gonçalves M. J., Cruz M. T., et al. Lavandula luisieri essential oil as a source of antifungal drugs. Food Chemistry. 2012;135(3):1505–1510. doi: 10.1016/j.foodchem.2012.05.090. [DOI] [PubMed] [Google Scholar]
- 21.Pinto E., Gonçalves M. J., Hrimpeng K., et al. Antifungal activity of the essential oil of Thymus villosus subsp. lusitanicus against Candida, Cryptococcus, Aspergillus and dermatophyte species. Industrial Crops and Products. 2013;51:93–99. doi: 10.1016/j.indcrop.2013.08.033. [DOI] [Google Scholar]
- 22.Taweechaisupapong S., Ngaonee P., Patsuk P., Pitiphat W., Khunkitti W. Antibiofilm activity and post antifungal effect of lemongrass oil on clinical Candida dubliniensis isolate. South African Journal of Botany. 2012;78:37–43. doi: 10.1016/j.sajb.2011.04.003. [DOI] [Google Scholar]
- 23.Raut J. S., Shinde R. B., Chauhan N. M., Karuppayil S. M. Terpenoids of plant origin inhibit morphogenesis, adhesion, and biofilm formation by Candida albicans . Biofouling. 2013;29(1):87–96. doi: 10.1080/08927014.2012.749398. [DOI] [PubMed] [Google Scholar]
- 24.Saharkhiz M. J., Motamedi M., Zomorodian K., Pakshir K., Miri R., Hemyari K. Chemical composition, antifungal and antibiofilm activities of the essential oil of Mentha piperita L. ISRN Pharmaceutics. 2012;2012:6. doi: 10.5402/2012/718645.718645 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Re R., Pellegrini N., Proteggente A., Pannala A., Yang M., Rice-Evans C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radical Biology and Medicine. 1999;26(9-10):1231–1237. doi: 10.1016/S0891-5849(98)00315-3. [DOI] [PubMed] [Google Scholar]
- 26.Garrett A. R., Murray B. K., Robison R. A., O'Neill K. L. Measuring antioxidant capacity using the ORAC and TOSC assays. Methods in Molecular Biology. 2010;594(12):251–262. doi: 10.1007/978-1-60761-411-1-17. [DOI] [PubMed] [Google Scholar]
- 27.Cao G., Prior R. L. Measurement of oxygen radical absorbance capacity in biological samples. Methods in Enzymology. 1998;299:50–62. doi: 10.1016/S0076-6879(99)99008-0. [DOI] [PubMed] [Google Scholar]
- 28.Green L. C., Wagner D. A., Glogowski J., Skipper P. L., Wishnok J. S., Tannenbaum S. R. Analysis of nitrate, nitrite, and [15N]nitrate in biological fluids. Analytical Biochemistry. 1982;126(1):131–138. doi: 10.1016/0003-2697(82)90118-X. [DOI] [PubMed] [Google Scholar]
- 29.Riss T. L., Moravec R. A., Niles A. L., et al. Assay Guidance Manual. National Center for Advancing Translational Sciences; 2004. Cell viability assays; p. p. 21. [Google Scholar]
- 30.Inouye S., Takizawa T., Yamaguchi H. Antibacterial activity of essential oils and their major constituents against respiratory tract pathogens by gaseous contact. Journal of Antimicrobial Chemotherapy. 2001;47(5):565–573. doi: 10.1093/jac/47.5.565. [DOI] [PubMed] [Google Scholar]
- 31.Filipowicz N., Kamiński M., Kurlenda J., Asztemborska M., Ochocka J. R. Antibacterial and antifungal activity of juniper berry oil and its selected components. Phytotherapy Research. 2003;17(3):227–231. doi: 10.1002/ptr.1110. [DOI] [PubMed] [Google Scholar]
- 32.Dai J., Zhu L., Yang L., Qiu J. Chemical composition, antioxidant and antimicrobial activities of essential oil from Wedelia prostrata . EXCLI Journal. 2013;12:479–490. [PMC free article] [PubMed] [Google Scholar]
- 33.Wang W., Li N., Luo M., Zu Y., Efferth T. Antibacterial activity and anticancer activity of Rosmarinus officinalis L. essential oil compared to that of its main components. Molecules. 2012;17(3):2704–2713. doi: 10.3390/molecules17032704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Matsuzaki Y., Tsujisawa T., Nishihara T., Nakamura M., Kakinoki Y. Antifungal activity of chemotype essential oils from rosemary against Candida albicans . Open Journal of Stomatology. 2013;3(2):176–182. doi: 10.4236/ojst.2013.32031. [DOI] [Google Scholar]
- 35.Rivas da Silva A. C., Lopes P. M., Barros de Azevedo M. M., Costa D. C. M., Alviano C. S., Alviano D. S. Biological activities of α-pinene and β-pinene enantiomers. Molecules. 2012;17(6):6305–6316. doi: 10.3390/molecules17066305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Cavaleiro C., Pinto E., Gonçalves M. J., Salgueiro L. Antifungal activity of Juniperus essential oils against dermatophyte, Aspergillus and Candida strains . Journal of Applied Microbiology. 2006;100(6):1333–1338. doi: 10.1111/j.1365-2672.2006.02862.x. [DOI] [PubMed] [Google Scholar]
- 37.Singh P., Shukla R., Prakash B., et al. Chemical profile, antifungal, antiaflatoxigenic and antioxidant activity of Citrus maxima Burm. and Citrus sinensis (L.) Osbeck essential oils and their cyclic monoterpene, DL-limonene. Food and Chemical Toxicology. 2010;48(6):1734–1740. doi: 10.1016/j.fct.2010.04.001. [DOI] [PubMed] [Google Scholar]
- 38.Ünal M. Ü., Uçan F., Şener A., Dinçer S. Research on antifungal and inhibitory effects of DL-limonene on some yeasts. Turkish Journal of Agriculture and Forestry. 2012;36(5):576–582. doi: 10.3906/tar-1104-41. [DOI] [Google Scholar]
- 39.Marei G. I. K., Abdel Rasoul M. A., Abdelgaleil S. A. M. Comparative antifungal activities and biochemical effects of monoterpenes on plant pathogenic fungi. Pesticide Biochemistry and Physiology. 2012;103(1):56–61. doi: 10.1016/j.pestbp.2012.03.004. [DOI] [Google Scholar]
- 40.Mitchell A. P. Dimorphism and virulence in Candida albicans . Current Opinion in Microbiology. 1998;1(6):687–692. doi: 10.1016/s1369-5274(98)80116-1. [DOI] [PubMed] [Google Scholar]
- 41.Saville S. P., Lazzell A. L., Bryant A. P., et al. Inhibition of filamentation can be used to treat disseminated candidiasis. Antimicrobial Agents and Chemotherapy. 2006;50(10):3312–3316. doi: 10.1128/AAC.00628-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Zore G. B., Thakre A. D., Rathod V., Karuppayil S. M. Evaluation of anti-Candida potential of geranium oil constituents against clinical isolates of Candida albicans differentially sensitive to fluconazole: inhibition of growth, dimorphism and sensitization. Mycoses. 2011;54(4):e99–e109. doi: 10.1111/j.1439-0507.2009.01852.x. [DOI] [PubMed] [Google Scholar]
- 43.Vuong C., Kocianova S., Voyich J. M., et al. A crucial role for exopolysaccharide modification in bacterial biofilm formation, immune evasion, and virulence. The Journal of Biological Chemistry. 2004;279(52):54881–54886. doi: 10.1074/jbc.m411374200. [DOI] [PubMed] [Google Scholar]
- 44.Soto S. M., Smithson A., Horcajada J. P., Martinez J. A., Mensa J. P., Vila J. Implication of biofilm formation in the persistence of urinary tract infection caused by uropathogenic Escherichia coli . Clinical Microbiology and Infection. 2006;12(10):1034–1036. doi: 10.1111/j.1469-0691.2006.01543.x. [DOI] [PubMed] [Google Scholar]
- 45.Neves Â., Rosa S., Gonçalves J., et al. Screening of five essential oils for identification of potential inhibitors of IL-1-induced Nf-κB activation and NO production in human chondrocytes: characterization of the inhibitory activity of α-pinene. Planta Medica. 2010;76(3):303–308. doi: 10.1055/s-0029-1186085. [DOI] [PubMed] [Google Scholar]
- 46.Quintans-Júnior L., Moreira J. C. F., Pasquali M. A. B., et al. Antinociceptive activity and redox profile of the monoterpenes (+)-camphene, p-cymene, and geranyl acetate in experimental models. ISRN Toxicology. 2013;2013:11. doi: 10.1155/2013/459530.459530 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Rufino A. T., Ribeiro M., Judas F., et al. Anti-inflammatory and chondroprotective activity of (+)-α-pinene: structural and enantiomeric selectivity. Journal of Natural Products. 2014;77(2):264–269. doi: 10.1021/np400828x. [DOI] [PubMed] [Google Scholar]