

Abstract
PGC-1α4, a novel isoform of the transcriptional coactivator PGC-1α, was recently postulated to modulate the expression of anabolic and catabolic genes and therefore regulate skeletal muscle hypertrophy. Resting levels of PGC-1α4 mRNA expression were found to increase in healthy adults after resistance training. However, the acute effect of resistance exercise (RE) on PGC-1α4 expression in populations prone to progressive muscle loss, such as postmenopausal women, has not been evaluated. Here we investigated alterations in mRNA expression of PGC-1α4 and PGC-1α1, a regulator of muscle oxidative changes, in postmenopausal women following high-intensity eccentric RE, and analyzed these findings with respect to changes in IGF-1 and catabolic gene expression. Nine postmenopausal women (57.9 ± 3.2 yr) performed 10 sets of 10 maximal eccentric repetitions of single-leg extension with 20 second rest periods between sets. Muscle biopsies were obtained from the vastus lateralis of the exercised leg before and 4 hours after the RE bout with mRNA expression determined by qRT-PCR. No significant changes in the mRNA expression of either PGC-1α isoform were observed following acute eccentric RE (P > 0.05). IGF-1Ea mRNA expression significantly increased (P < 0.05) while IGF-1Eb and MGF did not significantly change (P > 0.05). PGC-1α4 mRNA expression was associated with reduced mRNA expression of the catabolic gene myostatin (R = −.88, P < 0.01), while MGF mRNA expression was associated with reduced mRNA expression of the catabolic gene FOXO3A (R = −.81, P < 0.05). These data demonstrate an attenuated response of PGC-1α isoforms to an acute bout of maximal eccentric exercise with short rest periods in postmenopausal women.
Keywords: PGC-1α4, MGF, MYOSTATIN, RESISTANCE EXERCISE, SARCOPENIA
INTRODUCTION
Peroxisome proliferator-activated receptor gamma (PPARG) coactivator 1 alpha (PGC-1α) is a transcriptional coactivator that has been identified in oxidative tissues such as brown adipose (33), heart, and skeletal muscle (43). Due to its ability to bind to and coactivate numerous transcription factors involved with oxidative metabolism, PGC-1α was ascribed the role of a master regulator of mitochondrial biogenesis (23). Endurance exercise elicits the activation of PGC-1α in skeletal muscle through the upstream signaling cascades of AMP-activated protein kinase (AMPK), calcium/calmodulin-dependent protein kinase II (CAMKII), and p38 mitogen-activated protein kinase (MAPK) (3). Once activated, PGC-1α induces expression of mitochondrial and metabolic genes that result in the phenotypic changes in muscle associated with endurance training, including mitochondrial proliferation, fast-to-slow fiber type characteristic transitions, and angiogenesis (3).
Increases in the mRNA expression of PGC-1α have been observed in human skeletal muscle 2-3 hours after an acute bout of aerobic exercise (AE) in both trained and untrained individuals (30). Furthermore, PGC-1α mRNA expression is elevated after acute and chronic bouts of high-intensity interval training (HIIT) (32), which, over the long term, promote the oxidative remodeling of skeletal muscle similar to endurance training (13). However, the response of PGC-1α to resistance exercise (RE) is less defined. In one investigation, no change in PGC-1α mRNA expression was found in strength- and endurance-trained athletes post-acute RE (6), while in another, significant increases were observed after RE in the untrained state (10). Collectively, these findings preclude the role of PGC-1α in hypertrophy-stimulating exercise, such as long term RE.
Recently, Ruas et al. (38) demonstrated that resistance training does indeed influence PGC-1α expression in the form of a truncated version of the PGC-1α transcript, which they named PGC-1α4. PGC-1α4 originates from an alternative promoter ~13kb upstream of the canonical proximal promoter, and is functionally distinct from a previously identified truncated transcript, NT-PGC-α, that originates from the proximal promoter. In particular, PGC-1α4 has been shown to regulate anabolic and catabolic skeletal muscle signaling pathways without the activation of classical PGC-1α oxidative target genes (38), while NT-PGC-α was shown to be induced by hypoxia in muscle cells, and preferentially mediate angiogenesis via vascular endothelial growth factor (VEGF) expression with little effect on mitochondrial biogenesis (42). Ruas et al. (38) observed elevations in resting human skeletal muscle PGC-1α4 expression in association with increases in the anabolic hormone insulin-like growth factor-1 (IGF-1) and decreases in the muscle-secreted catabolic factor myostatin following an 8-week RE protocol. Furthermore, tumor-bearing mice with transgenic expression of PGC-1α4 exhibited resistance to the muscle-wasting of cancer. Thus, these authors concluded that PGC-1α4 may regulate a genetic program of muscle hypertrophy, likely through the activation of IGF-1 and suppression of myostatin signaling pathways.
IGF-1 is expressed in the muscle as several isoforms, IGF-1Ea, IGF-1Eb and IGF-1Ec, with each isoform described to contribute in some way to muscle hypertrophy or muscle regeneration (15, 44). IGF-1Ea is similar to IGF-1 synthesized by the liver, and when injected into mouse muscle, prevents the age-related loss of muscle, or sarcopenia, in old mice (4). IGF-1Ec stimulates satellite cell proliferation and muscle hypertrophy following stretch or damage to muscle, and hence is termed mechano-growth factor (MGF) (44). Increases in IGF-1Ea and MGF fold change were observed following a RE bout in young men (27), but mRNA expression of both isoforms were unchanged in older men (15). On the other hand, less is known about the exact physiological role of IGF-1Eb, although fold change increases in IGF-1Eb were found to coincide with peak myogenin fold change following exercise in young males (27).
The significance of the association of PGC-1α4 to the IGF-1 and myostatin pathways lies in its application to conditions of progressive muscle loss, such as sarcopenia. The age-related loss of muscle mass and function is associated with impairments in the performance of activities of daily living, particularly in older, postmenopausal women (19). In addition, menopause has been associated with a decline in lean mass related to a reduction in ovarian hormones independent of aging (8). The loss of muscle mass in older adults is attributed to the impaired ability of skeletal muscle to respond to anabolic stimuli and the increased activation of proteolytic signaling pathways (16). Specifically, anabolic stimuli, such as an acute bout of RE, stimulates robust protein synthesis in young individuals; however, the same bout of RE produces a blunted response in older adults (11). In addition, age-related muscle atrophy has been associated with increased signaling in the myostatin/activin pathway, which mediates muscle protein breakdown via the ubiquitin-proteasome system (16). The binding of myostatin to its muscle surface receptor dephosphorylates the fork-head box O (FOXO) family of transcription factors, which translocate to the nucleus to activate transcription of atrophy-specific genes (16). In particular, FOXO3A activates key ligases in the rate-limiting step of ubiquination, including atrogin-1/muscle atrophy F-box (MAFbx) and muscle RING finger 1 (MuRF1) (39). Therefore, the suppression of myostatin levels observed in young adults in association with increased PGC-1α4 expression following a RE program (38), in principle, makes PGC-1α4 an attractive target for conditions of progressive muscle loss. Indeed, older women express higher levels of myostatin at rest compared to young women (34) and myostatin-deficient mice do not experience the same age-related muscle atrophy as wild-type controls (40). Hence, an examination of the acute effects of RE on PGC-1α4, and its relationship to IGF-1 isoform and proteolytic mRNA expression, is warranted in an older population of women who may benefit from therapeutic interventions for muscle loss.
Thus, we investigated the mRNA expression of two transcriptional coactivators – PGC-1α4, a regulator of hypertrophy, and PGC-1α1, a regulator of oxidative changes – in skeletal muscle of postmenopausal women following an acute bout of maximal eccentric exercise. Changes in the mRNA expression of the IGF-1 isoforms IGF-1Ea, IGF-1Eb and MGF, and proteolytic genes including myostatin, MAFbx, FOXO3A and MuRF1 were also examined to further distinguish the regulatory roles of each PGC-1α isoform in skeletal muscle. We hypothesized that a single bout of maximal eccentric resistance exercise would elicit increases in the mRNA expression of PGC-1α4 and IGF-1 isoforms, while PGC-1α1 would remain unchanged in healthy, sedentary postmenopausal women.
METHODS
Experimental Approach to the Problem
Muscle samples collected in a previous investigation were used to generate data for the analyses in this study. The present study furthers the approach utilized in the prior investigation in postmenopausal women through the examination of additional gene targets and the retrospective comparison to proteolytic gene expression. Participants completed 10 sets of 10 maximal eccentric knee extensions, which have been shown to stimulate changes in muscle gene expression (20). Muscle samples were collected before and 4 hours following the acute exercise bout and were analyzed for PGC-1α4, PGC-1α1, IGF-1Ea, IGF-1Eb, and MGF mRNA expression using reverse transcriptase polymerase chain reaction (RT-PCR).
Subjects
A priori analysis determined that a sample size of 4 participants would have 80% power to detect a change from pre- to post-exercise for PGC-1α mRNA expression fold change of 3.6 with standard deviation 1.7 using a paired t-test with a 0.05 two-sided significance level. This was based on prior literature investigating PGC-1α via muscle biopsy (12).
Eligibility inclusion criteria included the following: a) subjects were postmenopausal women, 55-65 years of age, b) subjects were apparently healthy but could not have participated in more than 60 minutes of total exercise per week, and c) subjects were free of any musculoskeletal conditions that would prevent successful completion of the exercise protocol. This study was submitted to the University of Southern California Institutional Review Board for approval and all subjects signed a written informed consent form.
Nine healthy, postmenopausal women, 55 to 65 years of age (57.9 ± 3.2 yr) were enrolled in the study. All participants were sedentary at the time of study enrollment, which was determined by phone interview conducted by a study investigator (CDC), prior to study enrollment. A complete physical activity history was not obtained. The participants were classified as postmenopausal if they had not experienced spontaneous menstrual bleeding for at least 6 months. Use of hormone therapy (HT) was permitted as long as the participant remained on the drug for the duration of the study. Data was collected on the type, history, and duration of HT use. Two of the nine participants in the study were on HT, which has been shown to influence proteolytic gene expression following RE (7). However, when data from the HT participants were excluded, no changes in statistical significance were observed for any of our results. Therefore, the HT data was included in the final analyses.
Participants underwent an initial screening for eligibility with a history and physical examination performed by the study physician at the general clinical research center at the USC. Following the physician's examination, a nurse performed a single blood draw of the antecubital vein under fasting conditions from each subject which was analyzed by the Endocrinology Laboratory in the general clinical research center for the following measures: estrogen, glucose, insulin, thyroid hormone, and blood cell counts. Exclusion criteria included cancer within the last 5 years, diabetes or fasting blood glucose ≥126 mg/dL, cirrhosis or active hepatitis, uncontrolled thyroid function, clearance < 30 cc/min, ALT (alanine aminotransferase) > 1.5× upper limit of normal, or chronic lung problems or cardiovascular disease (myocardial infarction, heart failure, or active angina within the prior six months before enrollment), musculoskeletal disease or injury (rheumatoid arthritis, neuropathy, back injury, etc.) that would prevent resistance training, use of testosterone or other anabolic therapies, heparin, or coumadin within the last 6 months.
Procedures
Body composition
At baseline, total body mass, fat mass, lean mass, and percent fat were measured using a total body DEXA (dual-energy x-ray absorptiometer; model DPX-IQ 2288 with Smart Scan version 4.7e; Lunar Corporation, Madison, WI, USA). The same experienced investigator (CDC) was responsible for performing and analyzing all scans.
Physical activity assessment
All potential participants were pre-screened over the phone by the principal investigator (CDC) to ensure that they were sedentary and not participating in regular strenuous physical activity. Baseline, including current and past year, physical activity levels were assessed for each participant using the Entry Questionnaire and Physical Activity Scale (1). Lifetime physical activity patterns were not obtained. Participants were instructed to refrain from exercise during the duration of the study.
Dietary analysis
All participants completed a three-day dietary food record and were instructed to record their dietary intake over the course of 2 typical weekdays and 1 weekend day. Dietary entries were reviewed with a registered dietician to determine accuracy in portion size estimates and detail of food intake. Nutritionist ProTM (Nutrition Analysis Software Version 1.3, Jones and Bartlett Publishers, Inc., Sudbury, MA) was used for dietary analysis of intake data.
Strength testing
Strength testing was performed at baseline using the Cybex NormTM dynamometer (Cybex International Inc. Ronkonkoma, New York). A five minute warm-up session on a cycle ergometer preceded the strength testing. Participants received detailed instructions on the eccentric and concentric leg extension exercises, and performed 10 repetitions of each exercise at maximal effort. The maximum torque (effort measured) of the knee extensors for eccentric and concentric leg extension was performed with the dominant leg at 60°/s, a common setting utilized in clinical exercise physiology research studies (20). Participants were positioned on the Cybex by visually aligning the placement of the lateral femoral condyle with the axis of rotation of the lever arm. A strap was placed distally on the dominant leg at the level of the load cell. The load cell was positioned 3 cm proximal to the talocural joint. During maximal loading, a shoulder harness, hip restraint, and thigh strap (exercised leg) were used to limit excessive movement and secure the participant to the device. The settings were recorded to ensure exact placement for each acute eccentric exercise bout.
The tester determined the participants’ eccentric and concentric maximum torque as the highest produced torque. The maximum torque was determined to assess whether participants were generating maximal effort during each set of the acute training bout. All participants performed one session of strength testing prior to the acute training bout. Participants filled out and orally confirmed a modified Borg soreness scale 2-3 days following the strength testing.
Eccentric resistance exercise
All participants completed one acute bout of maximal single-leg eccentric knee extension exercise on the dominant leg using the Cybex NormTM dynamometer at 60°/s one week following strength testing. Participants fasted for 8 hours prior to the bout and testing was initiated at approximately 0800 AM. Information on sleep patterns, arousal states and hydration levels were not assessed prior to the exercise bout. Participants were positioned on the Cybex using the settings recorded from the strength testing visit. The exercise protocol consisted of 10 sets of 10 maximal eccentric repetitions with the participants performing the eccentric component while the investigator assisted the participant during the concentric component by moving the limb back to the starting position (15 degrees before full extension). The exercise protocol was designed to initiate skeletal muscle damage necessary to induce myogenic gene expression changes (20). Each repetition was separated by the time it took the testing investigator to manually return the lever arm back to 75 degrees of knee flexion (i.e. starting position). A 20 second rest period was given between each of the sets. Biofeedback was provided on the computer screen and verbal encouragement was provided by the tester during each repetition. The acute eccentric exercise bout occurred approximately 1 week after strength testing and the baseline muscle biopsy to allow recovery from soreness. Participants filled out and orally confirmed a modified Borg soreness scale 2-3 days following the acute eccentric exercise bout.
Muscle biopsies
Percutaneous muscle biopsies (150-200 mg) were obtained from the vastus lateralis of the exercised leg, one week prior to the acute eccentric exercise bout and 4 hours post-exercise. Four hours following exercise is an optimal time point to assess changes in myogenic gene expression stimulated by exercise (25). Biopsy specimens were collected using sterile conditions and local anesthesia (1% lidocaine) with a 5-mm Bergstrom muscle biopsy needle (Micrins Surgical, Lake Forest, Illinois) from the mid-portion of the vastus lateralis muscle. The post-exercise biopsy was performed at a distance of 2-4 cm proximal to the first site. Muscle tissue samples were immediately flash-frozen in liquid nitrogen and stored at −80°C until processed for analysis.
RNA extraction and cDNA synthesis
Total RNA was isolated following homogenization (Kinematica Polytron PT1200C, Switzerland) of 30-40 mg of muscle tissue with a monophasic solution containing guanidine-isothiocyanate-containing lysis buffer and β-Mercaptoethanol (Qiagen RNeasy Tissue Kit, Valencia, CA). After thorough homogenization, samples were centrifuged at 12,000 rpm at 4°C for 10 minutes and the resulting supernatant (free of insoluble protein and high molecular weight DNA) was transferred to a new microcentrifuge tube. Approximately 600 μL of Buffer RLT was added to these samples, samples were homogenized by passing the lysate 5 times though a 20-gauge needle fitted to an RNase-free syringe. Approximately 600 μL 70% ethanol was added to these samples. Samples were centrigued for 15 s at 12,000 rpm. The supernatant was transferred to a new microcentrifuge tube. The RNA pellet was then exposed to subsequent washes including steps one wash of 700 μL Buffer RW1 and two washes of Buffer RPE. Finally, the resultant air-dried RNA pellet was dissolved in 40 μL RNase-free water. The diluted RNA samples were then stored at −80°C until later analyses. The concentration and purity of the RNA was determined using a UV spectrophotometer (NanoDrop ND-1000, Thermo Scientific, Waltham, MA) by measuring absorbance at 260 nm and 280 nm.
After total RNA concentration determination, 500 nanograms of total skeletal muscle RNA was reverse transcribed to synthesize cDNA using Taqman reverse transcription reagents, according to the manufacturer's instructions (Applied Biosystems, Branchburg, NJ). The following reverse transcription reaction mixture was prepared (50 μL total): a) 10× RT buffer, b) MgCl2, c) dNTPs, d) random hexamers, e) RNase inhibitor, f) reverse transcriptase, g) RNase free water, and h) 500 ng total cellular RNA. RNA was reverse transcribed into cDNA with the following temperature/time protocol: 25°C for 10min, 48°C for 30min, 95°C for 5min, and 4°C infinite (Mycycler, BioRad, Hercules, CA).
Oligonucleotide primers for PCR
Forward and reverse oligonucleotide primers were used to amplify the gene expression of PGC-1α1, PGC-1α4, IGF-1Ea, IGF-1Eb, MGF, myostatin, FOXO3A, MAFbx, and MuRF1. The primer sequences for the specific target genes and melt curve temperatures are shown in Table 1. Primer sequences were designed using Primer3 program for myostatin, FOXO3A, MAFbx and MuRF1 (37), and as previously described for IGF-1Ea, IGF-1Eb, and MGF (15). Primer sequences for PGC-1α1 and PGC-1α4 were designed as previously described (38) (Figure 1). In particular, forward primers for detecting PGC-1α1 were located at exon 1a and reverse primers were located at exon 2; for PGC-1α4, forward primers were located at exon 5 and reverse primers were located at exon 7a. Melt curves were determined for each primer set to ensure amplification of pure PCR products. GAPDH was used as an internal control for detecting changes in gene expression using quantitative real-time PCR (qRT-PCR), which has previously been used as an internal control to examine myogenic gene expression following resistance exercise (25, 26, 35). In addition, we repeated all PCR analysis using cyclophilin as an alternative internal control gene. Results using cyclophilin (data not shown) and GAPDH did not differ, thus data pertaining to GAPDH only is presented.
Table 1.
Primer sequences used for qRT-PCR
| PCR Primer Sequence 5’ → 3” | Melt Curve Temperature | |
|---|---|---|
| PGC-1α1 | F: ATGGAGTGACATCGAGTGTGCT R: GAGTCCACCCAGAAAGCTGT |
58.7°C |
| PGC-1α4 | F: TCACACCAAACCCACAGAGA R: CTGGAAGATATGGCACAT |
60°C |
| IGF-1Ea | F: GCCTGCTCACCTTCACCAGC R: TCAAATGTACTTCCTTCTGGGTCTTG |
83°C |
| IGF-1Eb | F: ATCTACCAACAAGAACACG R: TACTTCCAATCTCCCTCC |
83°C |
| MGF | F: CGAAGTCTCAGAGAAGGAAAGG R: ACAGGTAACTCGTGCAGAGC |
84°C |
| Myostatin | F: CTGTAACCTTCCCAGGACCA R: CCCATCCAAAAGCTCAAAA |
82°C |
| FOXO3A | F: GAACGTGGGAACTTCACTGGTGCTA R: GGTCTGCTTTGCCCACTTCCCCTT |
84°C |
| MuRF1 | F: CTCAGTGTCCATGTCTGGAGGCCGTT R: GGCCGACTGGAGCACTCCTGTTTGTA |
79°C |
| MAFbx | F: CATCCTTATGTACACTGGTCCAAAGA R: TCCGATGATACACCCACATGTTAATG |
82°C |
| GAPDH | F: AGCCACATCGCTCAGACA R: GCCCAATACGACCAAATCC |
60°C |
F: Forward; R: Reverse;
Figure 1.

Representation of exons in the human PGC-1α gene as previously described (38).
Exon 1a represents the canonical proximal promoter, exon 1b represents the upstream alternative promoter, exon 7a results in truncated PGC-1α transcripts, and exon 7b results in nontruncated PGC-1α transcripts. Depicted above the exons is a schematic of splice variants with corresponding nomenclature. Listed below the exons are arrows indicating the primer pairs, along with the splice variants measured.
Quantitative Real-Time PCR
A qRT-PCR method was applied to determine relative gene expression levels for PGC-1α1, PGC-1α 4, IGF-1Ea, IGF-1Eb, MGF, myostatin, FOXO3A, MAFbx, and MuRF1. A total of 10ng of cDNA was added to each of the 20μl PCR reaction for each gene, including GAPDH. Each PCR reaction contained the following mixture: 10μl 2.5× iQSYBRgreen supermix (BioRad, Hercules, CA); 7μl RNase free water, 10ng cDNA and 10 ρmol of each primer (Forward and Reverse) of interest. All samples were run in quadruplicate. Each PCR reaction was amplified using BioRad iCycler iQ thermal cycler (BioRad, Hercules, CA). Thermal cycling conditions were as specified by the manufacturer: The amplification profile involved the initial denaturation step at 95°C for 3 min, followed by 60 cycles consisting of denaturation at 95°C for 15 s, primer annealing (60°C for PGC-1α4, 83°C for IGF-1Ea and IGF-1Eb; 58.7°C for PGC-1α1; 82°C for myostatin and MAFbx; 84°C for FOXO3A and MGF; and 79°C for MuRF1), and extension at 56°C for 1 min.
mRNA Quantification
All data were determined by normalizing the cDNA measured in 8 replicates (4 replicates repeated) for each participant sample to GAPDH (internal control) and then averaging the data to account for the change in mRNA expression as a result of the exercise stimulus. mRNA expression levels were determined using the 2−ΔCT method (24), where the GOI (gene of interest) expression was normalized to ICG (internal control gene) expression. For correlations between mRNA expressions, fold change was calculated using the 2−ΔΔCT method (24), where the GOI expression was normalized to ICG expression and normalized to preexercise value [ΔΔCT = (CT GOI POSTEXERCISE – CT ICG POSTEXERCISE) – (CT GOI PREEXERCISE – CT ICG PREEXERCISE)].
Statistical Analyses
Statistical analyses were performed using SPSS, version 16.0 (SPSS Inc., Chicago, IL). Data were normally distributed, and parametric analyses were used to compare gene expression. Paired t-tests were used to compare mRNA expression before and after exercise for each gene of interest. Pearson's product moment correlations were used to analyze the relationship between fold changes of PGC-1α transcripts, IGF-1 isoforms, and proteolytic markers including myostatin, FOXO3A, MAFbx, and MuRF1. A probability level of p < 0.05 was used to determine statistical significance for all analyses. Intraclass correlation coefficients (ICC) were calculated to assess the test-retest reliability of gene expression variables.
RESULTS
Baseline characteristics of the 9 participants are shown in Table 2. All participants had low activity levels with an average of 2.5 ± 0.5 hours of low-intensity exercise per week, consisting of mainly household chores and leisure walking. Average ratings of perceived exertion were 5.0 ± 0.5 (strong perceived soreness) out of 10 for the 1-RM testing and 8.0 ± 0.5 (very strong perceived exertion) out of 10 for the acute exercise bout.
Table 2.
Participant Characteristics (n=9)‡
| Age (yrs) | 57.9±3.2 |
| Height (cm) | 157.6±3.7 |
| Weight (kg) | 87.4±23.1 |
| BMI (kg/cm2) | 35.9±1.4 |
| Percent Fat (%) | 39.1±11.6 |
| LBM (kg) | 46.9±7.0 |
| Caloric intake (kcal)† | 1541±498 |
| Fat intake (g)† | 66.1±32.3 |
| Protein intake (g)† | 78.2±30.9 |
| Carbohydrate intake (g)† | 210.7±31.5 |
| 1-RM Eccentric (Nm) | 126.7±49.4 |
| 1-RM Concentric (Nm) | 88.0±25.1 |
Data presented as Mean ± SD.
Expressed as daily averages.
BMI, Body Mass Index; LBM, Lean Body Mass; kcal, kilocalories; g, grams; NM, Newton meters.
Relative change in mRNA expression
Compared to pre-exercise levels, PGC-1α1 and PGC-1α4 mRNA expressions were not significantly different after the acute exercise bout (P > 0.05; Figure 2).
Figure 2.
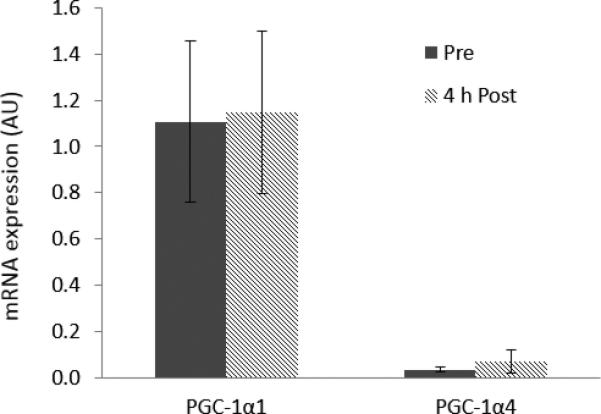
mRNA expression of PGC-1α1 and PGC-1α4 from human skeletal muscle before (pre) and 4 hours after (4 hr post) a single bout of maximal eccentric resistance exercise. Data represent arbitrary units normalized to GAPDH. Values are means ± SE. PGC-1α, peroxisome proliferator-activated receptor-γ coactivator-1α.
IGF-1 isoform mRNA expression was measured in a subset of participants (n = 7; data was not available in 2 participants due to lack of sampling). IGF-1Ea mRNA expression significantly increased following acute resistance exercise (P < 0.05; Figure 3). IGF-1Eb and MGF mRNA expression were not significantly different after the acute exercise bout (P > 0.05; Figure 3).
Figure 3.
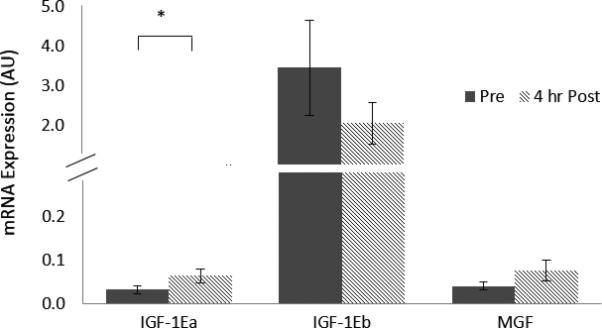
mRNA expression of IGF-1Ea, IGF-1Eb and MGF from human skeletal muscle before (pre) and 4 hours after (4 h post) a single bout of maximal eccentric resistance exercise. Data represent arbitrary units normalized to GAPDH. Values are means ± SE. IGF-1, insulin-like growth factor 1; MGF, mechano-growth factor. * P < 0.05
Fold change correlations with proteolytic genes
Fold changes of proteolytic mRNA expression of myostatin, FOXO3A, MAFbx, and MuRF1 were significantly decreased following an acute bout of RE as previously reported (7), These values were used to assess correlations between proteolytic and anabolic genes. A significant negative correlation was observed between exercise-induced fold change in PGC-1α4 and decrease in myostatin fold change (R = −0.85, P <0.01). In addition, the fold change in MGF was significantly correlated with decreases in the fold change in FOXO3A (R = −0.81, P < 0.05). No other significant correlations were observed between gene fold changes.
DISCUSSION
We investigated PGC-1α isoforms, and IGF-1 isoforms mRNA expression in skeletal muscle of postmenopausal women in response to a single bout of maximal eccentric RE. Additionally we examined the correlations between fold changes in mRNA expression of PGC-1α isoforms and IGF-1 isoforms, with fold changes in proteolytic genes. We report that mRNA expression of both PGC-1α4 and PGC-1α1 were not significantly different following RE. Furthermore, we observed a significant increase in the mRNA expression of IGF-1Ea after RE, but no significant changes were found in IGF-1Eb and MGF mRNA expression after RE. Finally, fold changes in myostatin negatively correlated with fold changes in PGC-1α4, and fold changes in FOXO3A negatively correlated with fold changes in MGF.
While numerous studies have examined the acute effects of RE on PGC-1α mRNA expression in relation to other molecular markers (10, 28, 32), this study is the first to relate PGC-1α isoforms to IGF-1 isoforms and proteolytic genes after a single bout of RE in postmenopausal women. The work of Ruas et al. (38) similarly investigated the mRNA expression of PGC-1α isoforms, IGF-1 and myostatin, but rather focused on the chronic effect of an 8 wk training program on resting levels of mRNA expression in a cohort of men and women of varying age. These authors noted the greatest increases in PGC-1α4 expression and decreases in myostatin expression in groups that performed resistance training or combined resistance + aerobic training. In contrast to these results, we report no change in PGC-1α4 expression after acute RE. A seemingly obvious explanation for the discrepant PGC-1 α4 expression between the present study and the work of Ruas et al. (38) are differences in the acute response to exercise versus chronic adaptations to training. However, several investigations measuring the acute response of PGC-1 α isoforms to RE have reported transient elevations at 2 h (32, 45), 3 h (10, 28), and 6 h (32) post-RE. Therefore, the lack of change in PGC-1α1 and PGC-1 α4 in the present study compared to previous exercise investigations on PGC-1α isoforms may be attributed to factors other than training status, such as participant characteristics or nature of the exercise.
Participant age may be expected to influence the response of PGC-1α isoforms following an acute bout of exercise, as aging has been shown to alter signaling events in skeletal muscle that lead to the suppression of PGC-1α, such as increases in reactive oxygen species that activate NF-ΚB, a negative regulator of PGC-1α (2). Evidence from rat models supports this age-associated alteration in PGC-1α mRNA expression, as old rats exhibited significantly lower resting PGC-1α expression compared to young rats (21). However, in humans, older age does not appear to influence the PGC-1α response to an acute bout of either aerobic or resistance exercise. Elderly men (>70 years of age) who performed aerobic exercise expressed significantly higher levels of PGC-1α after exercise compared to rest (18), while older men (>60 years of age) responded to aerobic exercise with PGC-1α mRNA expression comparable to that of younger men regardless of training status (5). Only one study has examined the response of both PGC-1α1 and PGC-1α4 after acute RE in older adults, and found that levels of PGC-1α4, but not PGC-1α1, increased above rest 3 h after exercise in a cohort of older men (70±4 yr) (28). These authors utilized a protocol of four sets of 10 repetitions at 75%-1RM with a 2 min rest period between sets for the leg press and leg extension exercises. As we employed 10 sets of 10 repetitions of maximal eccentric leg extension with 20 sec rest periods between sets, compared to the exercise stimulus utilized by Ogborn et al. (28), our protocol reflects shorter rest periods, greater intensity, and higher volume. Such a protocol would be expected to elicit greater increases in PGC-1α isoform expression as evidence from both mice (41) and human (32) studies indicate an intensity-dependent response in PGC-1α isoform to exercise. In particular, Pugh et al (32) observed greater increases in PGC-1α-ex1b, a splice variant originating from the alternative promoter that is synonymous with PGC-1α4, following a combined protocol of RE + HIIT compared to RE alone, while PGC-1α-ex1b expression was increased in a dose-dependent manner in mice that ran at faster speeds for 30 min (41). Thus, the lack of change in PGC-1α isoforms in the present study despite adequate RE stimulus may be attributed to gender and postmenopausal status of our participants rather than age.
As this study is the first to assess PGC-1α isoforms following acute RE in postmenopausal women, no direct comparisons can be made with studies of the same exercise stimulus in the same subject population. However, one investigation on the influence of estrogen on PGC-1α1 and other metabolic genes following acute aerobic exercise may lend insight to our findings on PGC-1α isoforms in postmenopausal women. Ninety minutes of moderate intensity cycling exercise significantly increased PGC-1α mRNA expression in the skeletal muscle of young women who had higher levels of circulating estrogen (luteal phase of menstrual cycle), but not in young women with lower levels of circulating estrogen (follicular phase of menstrual cycle) (12). This finding suggests that lower estrogen levels or lower ratios of progesterone to estrogen may be responsible for the lack of increase in PGC-1α mRNA expression. Thus, it is plausible that estrogen levels in postmenopausal women may blunt the exercise-induced increase in both PGC-1α1 and PGC-1α4 through mechanisms that have yet to be identified.
We also correlated previously reported myostatin expression (7) with PGC-1α4 expression and observed a significant negative relationship, implying that downregulation of myostatin may be related to increases in PGC-1α4 expression. Studies that have examined the response of myostatin to a single bout of RE have reported a diminished ability of postmenopausal women to downregulate myostatin expression following a RE stimulus compared to young women or men (22). Furthermore, postmenopausal women not using HT exhibited smaller suppression of myostatin following acute RE compared to postmenopausal women on HT (7). As the majority of participants in the present study were postmenopausal women not using HT (n=7 of 9), the lack of change in PGC-1α4 we observed post-RE may also be associated with the blunted downregulation of myostatin in this population. Support for this hypothesis is tenuously provided by a study examining the acute response of PGC-1α to RE versus aerobic +RE in young men, where significantly greater PGC-1α expression was observed in the RE group, which also displayed a trend of greater myostatin suppression (10). Future exploration into the relationship between PGC-1α isoforms and myostatin expression in response to acute RE is warranted as limited data exists.
We noted an increase in IGF-1Ea mRNA expression following RE with no changes in IGF-1Eb and MGF. This is in contrast to the work of Ruas et al. (38), who observed no changes in the chronic response of IGF-1 following 8 wk of resistance training or combined resistance+aerobic training. The divergent IGF-1 results may again be due to differences in the chronic versus acute response to exercise, or it may be due to differences in the primer pair used to detect IGF-1. We measured mRNA expression of 3 isoforms of IGF-1 – IGF-1Ea, IGF-1Eb, and MGF -- using previously described primer sets (15), while it is unknown if Ruas et al. (38) measured total IGF-1 or splice variant expression. Our findings are supported when compared to studies investigating the acute response of IGF-1 isoforms to eccentric RE (27, 29). The increase we observed in IGF-1Ea after RE is similar to results in young men, where IGF-1Ea fold change was found to increase following heavy eccentric RE at 6 h (29), 48 h (29), and 72 h after exercise (27). However, our results on IGF-1Ea differ when compared to conventional RE utilizing both concentric and eccentric movements, as IGF-IEa expression has been observed to decrease in young men < 6 h after exercise (31) or remain the same in both young and older males 48 h after exercise (36). Thus, despite differences in biopsy time points, our findings on IGF-1Ea are in line previous work utilizing a protocol of eccentric RE.
On the other hand, age appears to influence expression of IGF-1Ec (MGF) as young subjects exhibit increases in MGF expression as early as 2.5 h (15) and up to 24 h after heavy RE (22, 27), while older subjects show no change at either time point (15, 22). MGF expression has been shown to increase in older adults following a program of resistance training at 5 and 12 weeks (14), implying that habitual mechanical loading may be necessary to stimulate a response in older skeletal muscle. As MGF is implicated in the early stages of satellite cell activation (17), lack of change in MGF expression following an acute bout of RE, such as that observed in the present study, may reflect a reduced ability of older skeletal muscle to activate and proliferate satellite cells. Indeed, diminished numbers of satellite cells have been observed in older men compared to young men following a protocol of maximal eccentric exercise similar to the protocol used in the present study (9).
In addition to a small sample size, this study had several potential limitations. First, the untrained, sedentary cohort used in this study may limit generalizability to a larger population. Second, a third PGC-1α isoform, NT-PGC-1α, was inherently included in the mRNA expression of PGC-1α4 and PGC-1α1. This occurred as a result of the primer pairs selected, which were chosen to match those from a previous investigation (38) for the purpose of comparison. The inclusion of NT-PGC-1 α in both primer sets likely acted to obscure the effect of the acute RE bout on each PGC-1α isoform. As NT-PGC-1α has been shown to be activated by hypoxia (42), and our exercise protocol utilized minimal rest periods (20 sec), expression of NT- PGC-1α was plausibly increased as a result of RE and contributed to the responses of both PGC-1α4 and PGC-1α1. If a distinction between the responses of the two isoforms is desired, then primer pairs directed at the proximal versus alternative promoter on the PGC-1α gene as described by Ydfors et al. (45) might be considered instead. Third, our single biopsy at 4 h after exercise limits the observation of acute molecular markers that may peak before or after this time point.
In conclusion, this study demonstrates that in postmenopausal women, acute eccentric exercise elicits no change in a transcriptional coactivator responsible for oxidative alterations in muscle (PGC-1α1) or in a splice variant involved in the regulation of muscle hypertrophy (PGC-1α4). Nevertheless, the changes in PGC-1α4 following exercise were found to correlate to decreases in the atrophy-related factor myostatin. Furthermore, muscle growth factors associated with myogenic activity were found to increase (IGF-1Ea) or remain the same (IGF-1Eb and MGF) in response to RE, with changes in MGF correlating to decreases in FOXO3A. Future research is needed to elucidate the mechanisms underlying exercise-induced elevations in PGC-1α isoforms and signaling attenuation in the ubiquitin-proteasome pathway, particularly in populations who may benefit from age-related muscle loss prevention strategies.
PRACTICAL APPLICATIONS
Our findings highlight the growing importance of elucidating the role of emerging molecular regulators of skeletal muscle hypertrophy such as PGC-1α4 in response to various types of exercise. Resistance exercise has been shown to alter signaling pathways in skeletal muscle in favor of protein synthesis. Yet, in populations vulnerable to progressive skeletal muscle loss, signaling can become impaired, shifting the balance toward protein degradation. Identifying exercise that optimizes the transcription of myogenic targets may provide a strategy for increasing muscle anabolism, which has particular significance for attenuating skeletal muscle loss in clinical populations such as the sarcopenic elderly, hypogonadal men, and postmenopausal women. This research sheds light on the importance of the manipulation of exercise prescription as a means of increasing the acute expression of molecular markers associated with hypertrophy, in the context that maximal eccentric exercise provided a stimulus for the downregulation of proteolytic genes, but not anabolic genes. Fitness professionals may consider these findings when constructing exercise prescriptions that aim to counter muscle loss for postmenopausal women.
ACKNOWLEDGEMENTS
We would like to thank the subjects who participated in this study. We wish to thank the nurses and personnel of the General Clinical Research Center of the University of Southern California for their help with the clinical portion of this study.
The Clinical Exercise Research Center at the University of Southern California and NIH NCRR GCRC M0I RR000043 provided funds for this project.
Footnotes
The authors report no conflicts of interest. The results of the present study do not constitute endorsement by the NSCA.
REFERENCES
- 1.Aadahl M, Jorgensen T. Validation of a new self-report instrument for measuring physical activity. Med Sci Sports Exerc. 2003;35:1196–1202. doi: 10.1249/01.MSS.0000074446.02192.14. [DOI] [PubMed] [Google Scholar]
- 2.Alvarez-Guardia D, Palomer X, Coll T, Davidson MM, Chan TO, Feldman AM, Laguna JC, Vazquez-Carrera M. The p65 subunit of NF-kappaB binds to PGC-1alpha, linking inflammation and metabolic disturbances in cardiac cells. Cardiovasc Res. 2010;87:449–458. doi: 10.1093/cvr/cvq080. [DOI] [PubMed] [Google Scholar]
- 3.Arany Z. PGC-1 coactivators and skeletal muscle adaptations in health and disease. Curr Opin Genet Dev. 2008;18:426–434. doi: 10.1016/j.gde.2008.07.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Barton-Davis ER, Shoturma DI, Musaro A, Rosenthal N, Sweeney HL. Viral mediated expression of insulin-like growth factor I blocks the aging-related loss of skeletal muscle function. Proceedings of the National Academy of Sciences of the United States of America. 1998;95:15603–15607. doi: 10.1073/pnas.95.26.15603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Cobley JN, Bartlett JD, Kayani A, Murray SW, Louhelainen J, Donovan T, Waldron S, Gregson W, Burniston JG, Morton JP, Close GL. PGC-1alpha transcriptional response and mitochondrial adaptation to acute exercise is maintained in skeletal muscle of sedentary elderly males. Biogerontology. 2012;13:621–631. doi: 10.1007/s10522-012-9408-1. [DOI] [PubMed] [Google Scholar]
- 6.Coffey VG, Zhong Z, Shield A, Canny BJ, Chibalin AV, Zierath JR, Hawley JA. Early signaling responses to divergent exercise stimuli in skeletal muscle from well-trained humans. FASEB J. 2005;20:190–192. doi: 10.1096/fj.05-4809fje. [DOI] [PubMed] [Google Scholar]
- 7.Dieli-Conwright CM, Spektor TM, Rice JC, Sattler FR, Schroeder ET. Influence of hormone replacement therapy on eccentric exercise induced myogenic gene expression in postmenopausal women. J Appl Physiol. 2009;107:1381–1388. doi: 10.1152/japplphysiol.00590.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Douchi T, Iemura A, Matsuo T, Kuwahata T, Oki T, Yoshimitsu N, Nagata Y. Relationship of head lean mass to regional bone mineral density in elderly postmenopausal women. Maturitas. 2003;46:225–230. doi: 10.1016/s0378-5122(03)00195-6. [DOI] [PubMed] [Google Scholar]
- 9.Dreyer HC, Blanco CE, Sattler FR, Schroeder ET, Wiswell RA. Satellite cell numbers in young and older men 24 hours after eccentric exercise. Muscle & nerve. 2006;33:242–253. doi: 10.1002/mus.20461. [DOI] [PubMed] [Google Scholar]
- 10.Fernandez-Gonzalo R, Lundberg TR, Alvarez-Alvarez L, de Paz JA. Muscle damage responses and adaptations to eccentric-overload resistance exercise in men and women. Eur J Appl Physiol. 2014;114:1075–1084. doi: 10.1007/s00421-014-2836-7. [DOI] [PubMed] [Google Scholar]
- 11.Fry CS, Drummond MJ, Glynn EL, Dickinson JM, Gundermann DM, Timmerman KL, Walker DK, Dhanani S, Volpi E, Rasmussen BB. Aging impairs contraction-induced human skeletal muscle mTORC1 signaling and protein synthesis. Skelet Muscle. 2011;1:11. doi: 10.1186/2044-5040-1-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Fu MH, Maher AC, Hamadeh MJ, Ye C, Tarnopolsky MA. Exercise, sex, menstrual cycle phase, and 17beta-estradiol influence metabolism-related genes in human skeletal muscle. Physiol Genomics. 2009;40:34–47. doi: 10.1152/physiolgenomics.00115.2009. [DOI] [PubMed] [Google Scholar]
- 13.Gibala MJ, McGee SL. Metabolic adaptations to short-term high-intensity interval training: a little pain for a lot of gain? Exerc Sport Sci Rev. 2008;36:58–63. doi: 10.1097/JES.0b013e318168ec1f. [DOI] [PubMed] [Google Scholar]
- 14.Hameed M, Lange KH, Andersen JL, Schjerling P, Kjaer M, Harridge SD, Goldspink G. The effect of recombinant human growth hormone and resistance training on IGF-I mRNA expression in the muscles of elderly men. J Physiol. 2004;555:231–240. doi: 10.1113/jphysiol.2003.051722. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hameed M, Orrell RW, Cobbold M, Goldspink G, Harridge SD. Expression of IGF-I splice variants in young and old human skeletal muscle after high resistance exercise. J Physiol. 2003;547:247–254. doi: 10.1113/jphysiol.2002.032136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Han HQ, Zhou X, Mitch WE, Goldberg AL. Myostatin/activin pathway antagonism: Molecular basis and therapeutic potential. Int J Biochem Cell B. 2013;45:2333–2347. doi: 10.1016/j.biocel.2013.05.019. [DOI] [PubMed] [Google Scholar]
- 17.Hill M, Wernig A, Goldspink G. Muscle satellite (stem) cell activation during local tissue injury and repair. J Anat. 2003;203:89–99. doi: 10.1046/j.1469-7580.2003.00195.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Iversen N, Krustrup P, Rasmussen HN, Rasmussen UF, Saltin B, Pilegaard H. Mitochondrial biogenesis and angiogenesis in skeletal muscle of the elderly. Exp Gerontol. 2011;46:670–678. doi: 10.1016/j.exger.2011.03.004. [DOI] [PubMed] [Google Scholar]
- 19.Janssen I, Heymsfield SB, Ross R. Low relative skeletal muscle mass (sarcopenia) in older persons is associated with functional impairment and physical disability. J Am Geriatr Soc. 2002;50:889–896. doi: 10.1046/j.1532-5415.2002.50216.x. [DOI] [PubMed] [Google Scholar]
- 20.Jensky NE, Sims JK, Dieli-Conwright CM, Sattler FR, Rice JC, Schroeder ET. Exercise does not influence myostatin and follistatin messenger RNA expression in young women. J Strength Cond Res. 2010;24:522–530. doi: 10.1519/JSC.0b013e3181c8664f. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kang C, Chung E, Diffee G, Ji LL. Exercise training attenuates aging-associated mitochondrial dysfunction in rat skeletal muscle: role of PGC-1alpha. Exp Gerontol. 2013;48:1343–1350. doi: 10.1016/j.exger.2013.08.004. [DOI] [PubMed] [Google Scholar]
- 22.Kim JS, Cross JM, Bamman MM. Impact of resistance loading on myostatin expression and cell cycle regulation in young and older men and women. Am J Physiol Endocrinol Metab. 2005;288:E1110–1119. doi: 10.1152/ajpendo.00464.2004. [DOI] [PubMed] [Google Scholar]
- 23.Lin J, Handschin C, Spiegelman BM. Metabolic control through the PGC-1 family of transcription coactivators. Cell Metab. 2005;1:361–370. doi: 10.1016/j.cmet.2005.05.004. [DOI] [PubMed] [Google Scholar]
- 24.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 25.Louis E, Raue U, Yang Y, Jemiolo B, Trappe S. Time course of proteolytic, cytokine, and myostatin gene expression after acute exercise in human skeletal muscle. J Appl Physiol. 2007;103:1744–1751. doi: 10.1152/japplphysiol.00679.2007. [DOI] [PubMed] [Google Scholar]
- 26.Mahoney DJ, Carey K, Fu MH, Snow R, Cameron-Smith D, Parise G, Tarnopolsky MA. Real-time RT-PCR analysis of housekeeping genes in human skeletal muscle following acute exercise. Physiol Genomics. 2004;18:226–231. doi: 10.1152/physiolgenomics.00067.2004. [DOI] [PubMed] [Google Scholar]
- 27.McKay BR, O'Reilly CE, Phillips SM, Tarnopolsky MA, Parise G. Co-expression of IGF-1 family members with myogenic regulatory factors following acute damaging muscle-lengthening contractions in humans. J Physiol. 2008;586:5549–5560. doi: 10.1113/jphysiol.2008.160176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ogborn DI, McKay BR, Crane JD, Safdar A, Akhtar M, Parise G, Tarnopolsky MA. Effects of age and unaccustomed resistance exercise on mitochondrial transcript and protein abundance in skeletal muscle of men. Am J Physiol Regul Integr Comp Physiol. 2015;308:R734–741. doi: 10.1152/ajpregu.00005.2014. [DOI] [PubMed] [Google Scholar]
- 29.Philippou A, Papageorgiou E, Bogdanis G, Halapas A, Sourla A, Maridaki M, Pissimissis N, Koutsilieris M. Expression of IGF-1 isoforms after exercise-induced muscle damage in humans: characterization of the MGF E peptide actions in vitro. In vivo. 2009;23:567–575. [PubMed] [Google Scholar]
- 30.Pilegaard H, Saltin B, Neufer PD. Exercise induces transient transcriptional activation of the PGC-1α gene in human skeletal muscle. J Physiol. 2003;546:851–858. doi: 10.1113/jphysiol.2002.034850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Psilander N, Damsgaard R, Pilegaard H. Resistance exercise alters MRF and IGF-I mRNA content in human skeletal muscle. J Appl Physiol. 2003;95:1038–1044. doi: 10.1152/japplphysiol.00903.2002. [DOI] [PubMed] [Google Scholar]
- 32.Pugh JK, Faulkner SH, Jackson AP, King JA, Nimmo MA. Acute molecular responses to concurrent resistance and high-intensity interval exercise in untrained skeletal muscle. Physiol Rep. 2015;3 doi: 10.14814/phy2.12364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Puigserver P, Wu Z, Park CW, Graves R, Wright M, Spiegelman BM. A Cold-Inducible Coactivator of Nuclear Receptors Linked to Adaptive Thermogenesis. Cell. 1998;92:829–839. doi: 10.1016/s0092-8674(00)81410-5. [DOI] [PubMed] [Google Scholar]
- 34.Raue U, Slivka D, Jemiolo B, Hollon C, Trappe S. Myogenic gene expression at rest and after a bout of resistance exercise in young (18–30 yr) and old (80–89 yr) women. J Appl Physiol. 2006;101:53–59. doi: 10.1152/japplphysiol.01616.2005. [DOI] [PubMed] [Google Scholar]
- 35.Raue U, Slivka D, Jemiolo B, Hollon C, Trappe S. Proteolytic gene expression differs at rest and after resistance exercise between young and old women. J Gerontol A Biol Sci Med Sci. 2007;62:1407–1412. doi: 10.1093/gerona/62.12.1407. [DOI] [PubMed] [Google Scholar]
- 36.Roberts MD, Dalbo VJ, Sunderland KL, Poole CN, Hassell SE, Bemben D, Cramer J, Stout J, Kerksick CM. IGF-1 splice variant and IGF-1 peptide expression patterns in young and old human skeletal muscle prior to and following sequential exercise bouts. Eur J Appl Physiol. 2010;110:961–969. doi: 10.1007/s00421-010-1588-2. [DOI] [PubMed] [Google Scholar]
- 37.Rozen S, Skaletsky H. Primer3 on the WWW for general users and for biologist programmers. Methods in molecular biology. 2000;132:365–386. doi: 10.1385/1-59259-192-2:365. [DOI] [PubMed] [Google Scholar]
- 38.Ruas JL, White JP, Rao RR, Kleiner S, Brannan KT, Harrison BC, Greene NP, Wu J, Estall JL, Irving BA, Lanza IR, Rasbach KA, Okutsu M, Nair KS, Yan Z, Leinwand LA, Spiegelman BM. A PGC-1alpha isoform induced by resistance training regulates skeletal muscle hypertrophy. Cell. 2012;151:1319–1331. doi: 10.1016/j.cell.2012.10.050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Sandri M, Sandri C, Gilbert A, Skurk C, Calabria E, Picard A, Walsh K, Schiaffino S, Lecker SH, Goldberg AL. Foxo transcription factors induce the atrophy-related ubiquitin ligase atrogin-1 and cause skeletal muscle atrophy. Cell. 2004;117:399–412. doi: 10.1016/s0092-8674(04)00400-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Siriett V, Platt L, Salerno MS, Ling N, Kambadur R, Sharma M. Prolonged absence of myostatin reduces sarcopenia. J Cell Physiol. 2006;209:866–873. doi: 10.1002/jcp.20778. [DOI] [PubMed] [Google Scholar]
- 41.Tadaishi M, Miura S, Kai Y, Kawasaki E, Koshinaka K, Kawanaka K, Nagata J, Oishi Y, Ezaki O. Effect of exercise intensity and AICAR on isoform-specific expressions of murine skeletal muscle PGC-1alpha mRNA: a role of beta(2)-adrenergic receptor activation. Am J Physiol Endocrinol Metab. 2011;300:E341–349. doi: 10.1152/ajpendo.00400.2010. [DOI] [PubMed] [Google Scholar]
- 42.Thom R, Rowe GC, Jang C, Safdar A, Arany Z. Hypoxic induction of vascular endothelial growth factor (VEGF) and angiogenesis in muscle by truncated peroxisome proliferator-activated receptor gamma coactivator (PGC)-1alpha. J Biol Chem. 2014;289:8810–8817. doi: 10.1074/jbc.M114.554394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Wu Z, Puigserver P, Andersson U, Zhang C, Adelmant G, Mootha V, Troy A, Cinti S, Lowell B, Scarpulla RC, Spiegelman BM. Mechanisms Controlling Mitochondrial Biogenesis and Respiration through the Thermogenic Coactivator PGC-1. Cell. 1999;98:115–124. doi: 10.1016/S0092-8674(00)80611-X. [DOI] [PubMed] [Google Scholar]
- 44.Yang S, Alnaqeeb M, Simpson H, Goldspink G. Cloning and characterization of an IGF-1 isoform expressed in skeletal muscle subjected to stretch. J Muscle Res Cell Motil. 1996;17:487–495. doi: 10.1007/BF00123364. [DOI] [PubMed] [Google Scholar]
- 45.Ydfors M, Fischer H, Mascher H, Blomstrand E, Norrbom J, Gustafsson T. The truncated splice variants, NT-PGC-1alpha and PGC-1alpha4, increase with both endurance and resistance exercise in human skeletal muscle. Physiol Rep. 2013;1:e00140. doi: 10.1002/phy2.140. [DOI] [PMC free article] [PubMed] [Google Scholar]