

Abstract
Varicocele, the most important treatable cause of male infertility, is present in 15% of adult males, 35% of men with primary infertility, and 80% of men with secondary infertility. On the other hand, 80% of these men will not present infertility. Therefore, there is a need to differentiate a varicocele that is exerting a deleterious effect that is treatable from a “silent” varicocele. Despite the growing evidence of the cellular effects of varicocele, its underlying molecular mechanisms are still eluding. Proteomics has become a promising area to determine the reproductive biology of semen as well as to improve diagnosis of male infertility. This review aims to discuss the state-of-art in seminal plasma proteomics in patients with varicocele to discuss the challenges in undertaking these studies, as well as the future outlook derived from the growing body of evidence on the seminal proteome.
Keywords: male infertility, proteomics, seminal plasma, varicocele, varicocelectomy
INTRODUCTION
Varicocele is the most important treatable cause of male infertility. Defined as the presence of dilated veins within the pampiniform plexus with blood reflux,1 its reported prevalence is 15% of the adult male population.2 The fact that it is detected in 35% of men with primary infertility, and in up to 80% of men with secondary infertility,2,3 has led to the proposal that varicocele progressively causes a decline in male fertility potential.2 Moreover, because adolescents with varicocele may present with decreased ipsilateral testicular volume, it has also been proposed that varicocele initiates its effect on puberty onset.4
Treatment of varicocele, usually through surgical ligation of the dilated veins,5 is associated to a catch-up growth of the affected testis in adolescents,6 and to improve semen quality and fertility in adult men.7,8,9 However, some conflicting results have been reported which, associated to the fact that 80% of adult men with varicocele will not present with infertility,10 has led to the need to differentiate a varicocele that is exerting a deleterious effect (and that is treatable) from a “silent” varicocele.
While diagnosis of varicocele is performed by a clinical work-up that includes a physical exam and may include scrotal ultrasonography,11 diagnosing a varicocele that is indeed determining infertility or, in adolescents, which will lead to infertility, is not so easily achieved. Therefore, a number of reports have attempted to increase sensitivity in diagnosing an infertility-defining varicocele, mainly through the use of semen oxidative stress12,13,14,15 and sperm functional integrity testing.6,15,16,17,18 While these studies have increased sensitivity in detecting testicular dysfunction in varicocele, there is still a need to determine possible effective markers of a deleterious varicocele, in order to improve patient screening for treatment.
Despite the growing evidence of the cellular effects of varicocele, its underlying molecular mechanisms still elude clinicians and scientists.19 Many studies have thus focused on determining the molecular pathways associated to varicocele and to male infertility. Studying the secreted proteins in the male reproductive tract has become a promising area to determine the reproductive biology of semen as well as to improve the diagnosis of male infertility.20 Specifically, proteins present in the seminal plasma may contain important information regarding testicular function, especially because around 10% of the proteins found in seminal plasma arise from the testes.21
Therefore, we have reviewed the importance of studying the seminal plasma protein profile, as well as thoroughly reviewed the current literature on seminal plasma proteomics in male infertility, in general, with a specific focus on varicocele and its treatment. Moreover, we have discussed current challenges in undertaking these studies, as well as the future outlook derived from the growing body of evidence on the seminal proteome.
REVIEW CRITERIA
We have performed a literature search using PubMed and Google Scholar electronic databases, with the following keywords: “seminal plasma AND proteomics;” “Seminal plasma AND proteomic;” “Seminal plasma AND proteome;” “Seminal plasma proteomics;” “Seminal plasma proteomic;” “Seminal plasma proteome;” “Varicocele AND proteomics;” “Varicocele AND proteomic;” “Varicocele AND proteome;” “Varicocele proteomics;” “Varicocele proteomic;” and “Varicocele proteome.” Only articles written in English published in peer reviewed journals were selected. The search was performed in June and July, 2015, and covered all articles published until this date.
PROTEOMICS AS A TOOL FOR THE STUDY OF MALE INFERTILITY
Proteomics is a term used to define the comprehensive study of all the proteins of any given fluid or a cell under a specific biological condition.22 Proteins are effector molecules within a general cascade that initiates with Genomics studies, through Transcriptomics down to Proteomics and Metabolomics.23 Because proteins integrate an individual response to environmental conditions, as well as effect a given biological response,23 recent studies which focused on understanding the mechanisms of diseases have deployed great efforts toward understanding not only the proteins present in a given situation, but also their quantities, interactions, and post-translational modifications, as well as the pathways they enrich.24,25,26,27
Current developments in separation techniques as well as in mass spectrometry equipment have greatly increased our ability to delve deep into the proteome of cells and fluids.28 This is of particular interest to biological conditions in which phenotypic differences are determined by proteins in lower quantities. In seminal plasma, 15 proteins may account to up to 90% of the amount of protein (data not published), which means that more often the proteins that determine male infertility will be present in the remaining 10%. Moreover, seminal plasma is composed of fluids secreted from the seminal vesicles, prostate, bulbourethral glands and the vas deferens, epididymides, and seminiferous tubule lumen.29 Therefore, proteins secreted from the testes, which more likely represent testicular function, will be present at very low quantities, and increased sensitivity and dynamic ranges are essential in order to understand the molecular mechanisms of male infertility.
Shotgun proteomics studies, which are untargeted studies based on extensive mapping of generated protein digests, allow identification of a large number of expressed proteins in the seminal plasma, focusing on understanding their functions, interactions, and contribution to biological processes.30 These studies are based on mass spectrometry (MS) identification and quantification of proteins.
Briefly, mass spectrometers are subdivided into (i) an ionization source, such as matrix-assisted laser desorption ionization (MALDI) and electrospray ionization (ESI), (ii) mass analyzer(s), such as quadrupole (Q) and time-of-flight (TOF), and (iii) a detector. For peptide identification, ionized peptides can be fragmented in a collision chamber coupled to a high resolution mass analyzer, in an approach termed tandem MS (MS/MS).31 In addition, MS can be coupled to liquid chromatography (LC-MS/MS), enabling separation (fractionation) of peptides to decrease sample complexity, as well as generating quantitative information.32 Prior to MS, samples may be separated by one-dimensional (1DE) or two-dimensional electrophoresis (2DE), in order to reduce sample complexity in the protein level, leading to the possibility of observing proteins present at smaller amounts. In 1DE, proteins are separated by their molecular masses, whereas in 2DE proteins are also separated by their isoelectric point (pH in which the protein has no electrical charge).33,34 In both cases, the bands or spots of interest are excised from the gel and analyzed by MS. For proteomic data validation, selected reaction monitoring (SRM) allows for absolute protein quantification, in a targeted MS approach.35 The different techniques used for proteomics studies were recently reviewed by Scherl.31 Shotgun proteomics can be applied both to describe the whole seminal plasma proteome and to compare the proteomes between different infertility conditions, which will be further described below.
ORIGIN AND CHARACTERIZATION OF THE SEMINAL PLASMA PROTEOME
The seminal plasma components, secreted from the testes, the cauda epididymis and the accessory sexual glands, protect the sperm during their transit through the female reproductive tract, playing major roles for sperm survival, function, motility and viability, as well as for successful fertilization.29,36,37,38,39 In particular, seminal plasma proteins are capable of binding and fusing to the sperm membrane to regulate its survival after ejaculation,40 motility,41,42,43 capacitation40,44 and interaction and fusion with the oocyte.45 Moreover, seminal plasma proteins are capable of interacting with the female genital tract epithelium, to regulate sperm transport and female immune response.46 The main functions of seminal plasma proteins were reviewed in depth by Rodríguez-Martínez et al.47
Seminal plasma is rich in extra and intracellular proteins, with a mean total protein concentration of 35–55 g l−1.39 The majority of seminal plasma proteins are spermadhesins, cysteine-rich secretory proteins (CRISPs), proteins carrying fibronectin type II domains and enzymes.48 Approximately, 70% of the protein content of seminal plasma is secreted from the seminal vesicles, such as semenogelin-1 (SEMG1) and semenogelin-2 (SEMG2), important proteins for semen coagulation after ejaculation, lactotransferrin (LTF), and fibronectin (FN1).49 Another 20% of seminal plasma proteins are secreted from the prostate (three main proteins regulated by hormones: prostate-specific antigen – KLK3, important for semen liquefaction, prostatic acid phosphatase – ACPP, and beta-microseminoprotein - MSMB) and the bulbourethral glands (mainly mucins).48,50,51 Finally, approximately 10% of seminal plasma proteins are of testicular or epididymal origin.21 It has already been shown that proteins associated with testicular function, as well as proteins from altered sperm present throughout the male reproductive tract, are observed in the seminal plasma.39,52,53 Therefore, the seminal plasma protein profile reflects not only the activity of the male sexual accessory glands, but also spermatogenesis, epididymal maturation and sperm integrity.
SEMINAL PLASMA PROTEOMICS AND MALE INFERTILITY
A summary of studies focusing on the seminal plasma proteome in fertile men and men with varicocele is presented in Tables 1 and 2, respectively. Characterization of the seminal plasma proteome is important in order to determine, as best as possible, a normal state (Table 1). In men with normal semen analysis, Fung et al.54 identified, utilizing different approaches, over 100 proteins, such as SEMG1, SEMG2, KLK3, ACPP, albumin (ALB), and prolactin inducible protein (PIP). Furthermore, Pilch and Mann39 later reported, by the use of 1DE followed by LC-MS/MS, 923 proteins in seminal plasma of one healthy donor. From the 52% of proteins with an assigned subcellular location, 78% were cellular proteins and 25% were extracellular or secreted proteins (including proteins from prostasomes), demonstrating that the seminal plasma is composed of many more secreted proteins than other fluids. The seminal plasma proteome described in that study presented with 66% homology with the prostate fluid and 18% with the epididymal fluid, while 38% were common to all fluids.55
Table 1.
Descriptive seminal plasma proteomics studies in men with normal semen analysis or of proven fertility

Table 2.
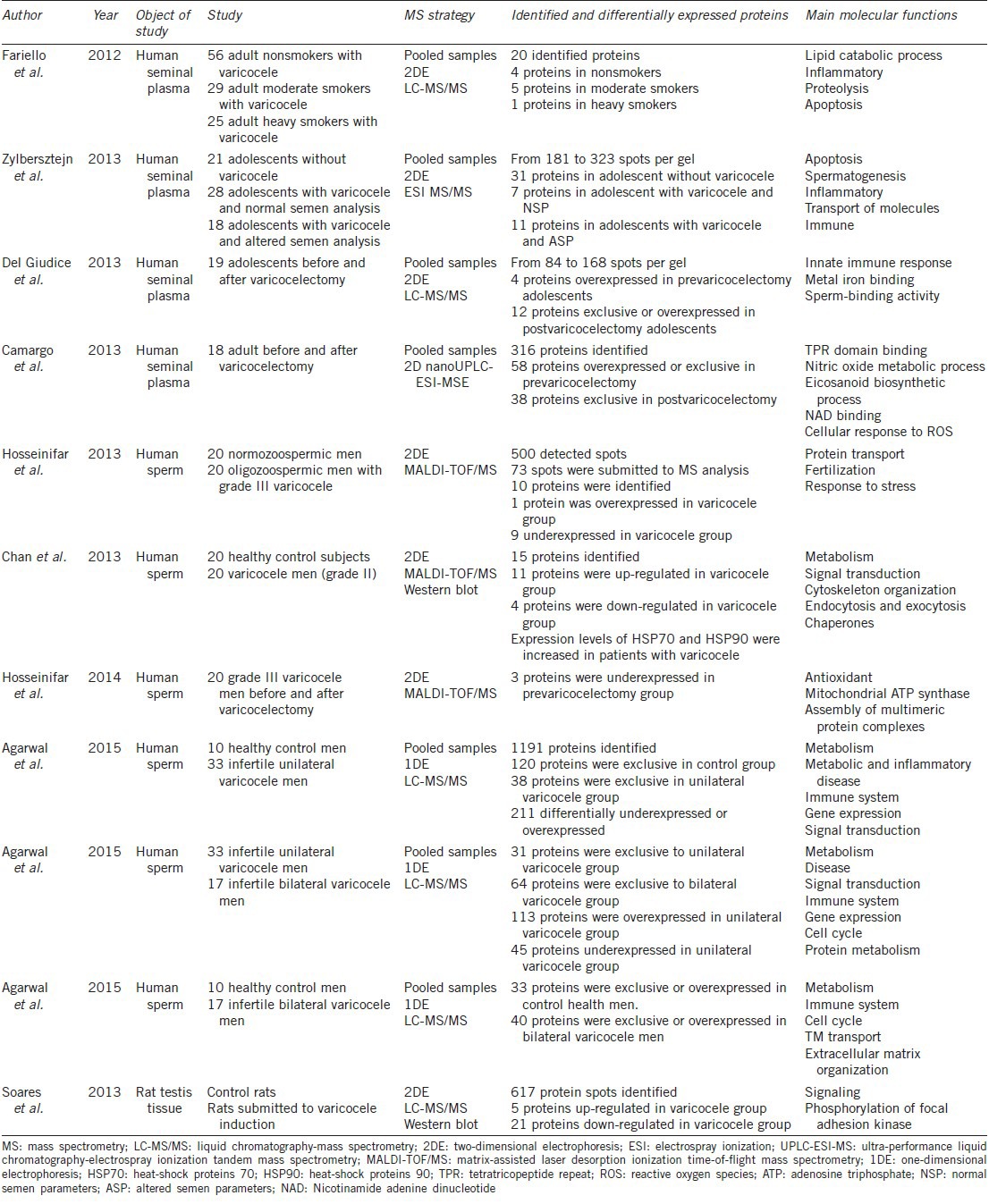
Descriptive proteomics studies in adolescents and adult men with varicocele
In 2012, Milardi et al.52 evaluated the seminal plasma of 5 men with proven fertility, and identified between 919 and 1487 proteins per sample, of which 83 were observed in all five samples, such as SEMG1, SEMG2, LTF, olfactory receptor 5R1 (OR5R1), clusterin (CLU), and E3 ubiquitin-protein ligase UBR5 (UBR5). Rolland et al.56 then demonstrated, in a healthy donor, a total of 669 proteins which integrated to data from previous studies, accounted for 2545 seminal plasma proteins. These proteins were mostly involved in protein binding (~50%) and catalytic activity (~39%), and were mainly cytoplasmic (58.7%, such as proteins found in vesicles and the endomembrane system or in the cytoskeleton) or extracellular (21.2%) proteins. The authors suggested that seminal plasma lacks nucleic acid-binding proteins, transcription regulators, and membrane receptors and channels. These proteomic results were also compared to gene expression data from testis, epididymis and prostate, available in public databases, to identify the contribution of these organs to the seminal plasma proteome. Using this approach, the authors identified 83 testicular proteins, 42 epididymal proteins, 7 seminal vesicles proteins, and 17 prostatic proteins.56
These studies have established the seminal plasma proteome associated with men presenting with normal semen analysis or of proven fertility. To compare the seminal plasma proteome between fertile men and men with different infertility conditions, several studies were performed, as described in Supplementary Table 1 (44.1KB, pdf) . These studies have been performed mainly in the search for biomarkers of infertility, and, in this context, seminal plasma is a promising source, because it presents high concentrations of proteins, many of which are tissue-specific (testicular proteins, for instance), as well as non secreted proteins (arriving, for example, from altered sperm).57
Comparative seminal plasma proteomics studies in male infertility
One of the most studied causes of infertility using a proteomics approach thus far is azoospermia. In men after vasectomy (mimicking obstructive azoospermia) and with nonobstructive azoospermia (Sertoli cell-only syndrome – SCOS), several proteins, such as CLU, were underexpressed compared to controls, and are potential biomarkers of impaired spermatogenesis.58,59 To identify the contribution of the testis and epididymis to the human seminal plasma proteome, fertile and vasectomized (mimicking obstructive azoospermia) men were compared.21 Thirty-two proteins were uniquely presented in the fertile group (testicular and epididymal proteins). In the vasectomy group, 49 proteins were underexpressed (proteins expressed in the testis and epididymis, but also in other regions of the male reproductive tract). The identified testicular and epididymal proteins, such as testis-expressed sequence 101 protein (TEX101), have important reproductive functions and may be potential biomarkers of obstructive azoospermia.
In a follow-up study,20 5 men with nonobstructive azoospermia were evaluated and the obtained data were compared to their previous study.21 Sorbitol dehydrogenase (SORD) was able to differentiate between obstructive and nonobstructive azoospermia and was thus suggested as a potential biomarker, in addition to several proteins suggested in another study as biomarkers of nonobstructive azoospermia and obstructive azoospermia, such as NPC2 and PIP, respectively.60 Furthermore, a study was performed to narrow down the potential biomarker candidates for differential diagnosis of azoospermia,61 and 18 proteins were selected to be further analyzed by multiplex SRM.62 Extracellular matrix protein 1 (ECM1) levels were higher in fertile men and in men with nonobstructive azoospermia, but highly decreased in vasectomized men, differentiating these conditions with high specificity and sensitivity.62 On the other hand, TEX101 levels were higher in fertile men and undetectable in SCOS and postvasectomy samples.62 These data might increase the confidence in nonobstructive azoospermia and obstructive azoospermia diagnosis using these two biomarkers, and facilitate the prediction of testicular sperm extraction (TESE) outcome.
To study only nonobstructive azoospermia, Freour et al.63 classified men as Sperm Positive (presenting with sperm in TESE) or Sperm Negative (absence of sperm in TESE). Sixty-eight proteins were differentially expressed, of which they selected CLU, PIP and galectin-3-binding protein (LGALS3BP) as potential spermatogenesis biomarkers in the seminal plasma. LGALS3BP was further confirmed by ELISA and presented in higher levels in Sperm Positive samples, with a concentration lower than 153 ng ml−1 associated with a negative TESE outcome. Finally, Rolland et al.56 demonstrated that L-lactate dehydrogenase C chain (LDHC), phosphoglycerate kinase 2 (PGK2), and transketolase-like protein 1 (TKTL1) proteins are not (or less) observed in the seminal plasma from nonobstructive azoospermia, obstructive azoospermia and vasectomized men, suggesting that these proteins may be biomarkers of male fertility.
In addition to studies focusing on azoospermia, other infertility conditions were also evaluated. In smokers with varicocele, several proteins were related to smoking, such as annexin A3 (ANXA3) and extracellular superoxide dismutase [Cu-Zn] (SOD3) in moderate smokers (<20 cigarettes a day), and zinc-alpha-2-glycoprotein (AZGP1) in heavy smokers.64 In men with impaired spermatogenesis (oligozoospermia, asthenozoospermia, and/or teratozoospermia), several proteins were altered, and, interestingly, oxidative stress was the major mechanism suggested to play a role in these alterations.41,65,66,67 In men with spinal cord injury, overexpressed proteins were mostly involved with smooth muscle function, cytoskeletal binding, homeostasis, oxidation of iron and response to calcium. This study demonstrated that the method of semen retrieval in these patients may also alter the obtained proteome.68 Finally, in androgen deficiency, it was demonstrated that the semen of these patients lacks proteins mostly involved with catalytic and binding activities, and hydrolase is the most impaired enzyme by testosterone deficiency.69
Several studies were also carried out in order to verify if the seminal plasma proteome reflects oxidative status and sperm function. Seminal oxidative stress was associated to specific enriched functions such as transition metal ion response/homeostasis, reactive oxygen species (ROS) detoxification, aging, and cell morphogenesis, differentiation, motility and cycle.70,71 Utilizing a similar approach in order to verify mechanisms underlying male infertility, another study analyzed the seminal plasma proteome in relation to sperm DNA fragmentation in normozoospermic men.53 Thirty proteins were altered in men presenting high sperm DNA fragmentation, which evidenced acute-phase response, fatty acid-binding, vesicle lumen and endoribonuclease activity. Finally, fertile and infertile men, irrespective of the cause for the infertility, were compared, and 10 proteins were overexpressed in the infertile group, although these proteins were not identified by MS/MS.72
More recently, a review was performed to search for all seminal plasma proteomics studies in order to obtain the whole list of seminal plasma proteins identified so far using different MS approaches. A total of 4,188 proteins were identified, of which 2,168 were non-redundant proteins (identified using UniProtKB/Swiss-Prot reviewed database). These proteins were mostly involved with proteolysis and carbohydrate catabolism. The authors also demonstrated that different MS techniques might complement each other. Moreover, by analyzing several publications on post-translation modifications, it was possible to highlight that phosphorylation, acetylation, glycosylation and disulfide bonds are the majority of post-translation modifications present in seminal plasma proteins.73
SEMINAL PLASMA PROTEOMICS AND VARICOCELE
Because varicocele is a complex disease with a number of different mechanisms that may determine infertility, the paradigm-shifting approach of studying its postgenomic pathways brings the promise of understanding, in a more precise manner, the effector pathways of varicocele-derived male infertility, as well as predict treatment outcomes.74 A summary of studies focusing on the seminal plasma proteome in varicocele men and the analysis of its intervention is presented in Table 2. Because there are only a few studies regarding seminal plasma proteomics in men with varicocele, proteomic analysis of spermatozoa is also discussed in this topic.
Proteomic studies regarding the effect of varicocele and its treatment
In adolescents
Zylbersztejn et al.75 compared the proteomic profile of seminal plasma of adolescents without varicocele, with varicocele and normal semen analysis, and with varicocele and altered semen analysis, using a 2DE approach. Forty-seven spots were extracted and submitted to mass spectrometry identification. Proteins such as SEMG1 and Insulin-like growth factor-binding protein 3 (IGFBP3) were observed in seminal plasma of adolescents with varicocele and altered semen, and are linked to regulation of apoptosis. Conversely, E3 ubiquitin-protein ligase BRE1B (RNF40), Proactivator polypeptide-like 1 (PSAPL1) and Epididymal secretory protein E3-beta (EDDM3B) were observed in seminal plasma of adolescents with varicocele and normal seminal analysis. The authors concluded that proteins exclusively expressed in seminal plasma of adolescents with varicocele could be potential biomarkers for early varicocele diagnosis regarding the negative impact of varicocele in the testes, even those found in patients with normal seminal parameters.75
To study the effect of varicocelectomy in seminal plasma proteome of adolescents, Del Giudice et al.76 used 2DE analysis followed by LC-MS/MS for spot identification. The authors observed that 19 spots were differentially or exclusively expressed, and determined that two proteins were overexpressed before varicocelectomy: DNA-directed RNA polymerase III subunit RPC2 (POLR3B) and Negative elongation factor E (NELFE). At 3 months postvaricocelectomy, 6 proteins were exclusively expressed or overexpressed, including SEMG1, SEMG2, ACPP, KLK3, TGM4, and alpha-1-antitrypsin (SERPINA1).76 All proteins found differentially expressed in postvaricocelectomy group play a crucial role in the physiological process of ejaculation, thus a shift-back to homeostasis can be observed after varicocelectomy.77
In adults
In order to verify the effect of varicocelectomy in men, Camargo et al.74 studied seminal plasma of men who underwent varicocelectomy using a label-free shotgun proteomics approach. In total, 316 proteins were identified and quantified, of which 58 proteins were overexpressed or exclusively expressed in the prevaricocelectomy men, and 38 were exclusively expressed in the postvaricocelectomy men. In prevaricocelectomy group, the proteins CLU, annexin A5 (ANXA5), L-lactate dehydrogenase A chain (LDHA), 14-3-3 protein epsilon (YWHAE), 14-3-3 protein theta (YWHAQ), Heat shock protein HSP 90-beta (HSP90AB1), and Heat shock protein HSP 90-alpha (HSP90AA1) presented enriched functions such as metabolism and regulation of nitric oxide (NO), and of TPR domain-binding. After varicocelectomy, Protein deglycase DJ-1 (PARK7), Superoxide dismutase [Cu-Zn] (SOD1), Protein S100-A9 (S100A9), Annexin A1 (ANXA1), Glyceraldehyde 3-phosphate dehydrogenase (GAPDH), and cytoplasmic malate dehydrogenase (MDH1) enriched functions such as cellular response to ROS, gluconeogenesis, NAD-binding and protein stabilization.
Hosseinifar et al.78 characterized the sperm proteome in patients with varicocele. The authors studied sperm from normozoospermic men (control) and compared these with sperm from oligozoospermic men with varicocele using a 2DE approach. From 500 spots initially observed, 73 were selected for MALDI-TOF/MS identification. The results showed 15 differentially expressed proteins, which include SEMG2 and its precursor PARK7, CLU, and 78 kDa glucose-regulated protein (HSPA5), underexpressed in patients with varicocele when compared with control men. The authors concluded that heat-shock proteins, mitochondrial proteins, and cytoskeleton proteins in sperm are affected by varicocele.78 Using the same approach, Chan et al.79 also determined 15 proteins differentially expressed between men with and without varicocele. After confirmation by Western blotting, heat-shock proteins 70 and 90 (HSP70 and HSP90) were found overexpressed in sperm of men with varicocele.79
Furthermore, Hosseinifar et al.80 identified, by using 2DE, differences in protein expression in sperm of men submitted to varicocelectomy. Three proteins were underexpressed in the prevaricocelectomy samples: SOD1, HSPA5, and ATP synthase subunit delta, mitochondrial (ATP5D). HSPA5 is a member of HSP70 family,12 which agrees with their previous results.79 Moreover, Chan et al.79 and Camargo et al.74 also observed these proteins in patients with varicocele and in prevaricocelectomy patients, respectively, as discussed above. Therefore, heat shock proteins seem to be key elements in the adult varicocele.
Agarwal et al. recently performed three different studies with 1DE and LC-MS/MS to better understand unilateral and bilateral varicocele. When sperm proteomics from men with unilateral varicocele and men with proven fertility (controls) were compared, 1191 proteins were observed, of which 369 proteins were exclusive or differentially expressed between the groups.81 From these, 15 proteins were involved in reproductive functions such as: (i) motility, overrepresented by calcium-binding tyrosine phosphorylation-regulated protein (CABIR), prostate and testis expressed protein 4 (PATE4), radial spoke head protein 9 homolog isoform 1 (RSPH 9), dynein heavy chain 17, axonemal (DNAH17), apolipoprotein A-I (APOA1), sodium/potassium transporting ATPase subunit alpha-4 isoform 1 (ATP1A4) and A-kinase anchor protein 3 (AKAP3), which were all underexpressed in men with varicocele; (ii) capacitation, overrepresented by SEMG2 and ATP1A4, underexpressed in varicocele men; (iii) hyperactivation and acrosome reaction, overrepresented by NPC2, dihydrolipoyl dehydrogenase, mitochondrial (DLD), acrosin precursor (ACR), and cysteine-rich secretory protein 2 (CRISP2), which were overexpressed in patients with varicocele, and (iv) zona-pellucida binding, overrepresented by sperm surface protein Sp17 (SPA17), which was underexpressed in varicocele group and acrosome formation associated factor isoform 1 (EQTN), exclusive in controls.81
In another study, the sperm proteomic profile of men with bilateral varicocele was compared to men with proven fertility (controls).83 In that study, 73 differentially expressed proteins were observed between the groups: 8 exclusive and 25 overexpressed in fertile controls and 7 exclusive and 33 overexpressed in bilateral varicocele. Two main proteins were found and confirmed by Western blotting: (i) T-complex protein 11 homolog (TCP11), previously located to germ cells of fertile men, and (ii) Tektin-3 (TEKT3), which is believed to play a role in low sperm motility in men with bilateral varicocele.82 In the third study, when sperm from unilateral and bilateral varicocele patients were compared, 31 proteins were exclusively expressed in unilateral varicocele group and 64 were exclusively expressed in bilateral varicocele.83 The authors demonstrated that specific proteins in seminal plasma could perhaps explain why bilateral varicocele is more aggressive, such as SEMG1, SEMG2, fibrous sheath CABYR-binding protein (FSCB), enkurin (ENKUR), and hyaluronidase PH-20 (SPAM1).83 However, Hosseinifar et al.78 found SEMG2 underexpressed in patients with varicocele when compared with healthy control men, although the authors utilized a different technique which may be sensitive to protease activity and thus lead to altered results.
In addition, Soares et al.84 studied the effect of varicocele in a rat animal model. After varicocele induction, the testes were removed and the homogenates were submitted to 2DE analysis followed by LC-MS/MS. Six proteins were up-regulated and 23 were down-regulated in varicocele-induced rats when compared to sham-operated rats. The authors observed guanine nucleotide binding protein subunit beta-2-like 1 (Gnb2l1 – homologous to GNB2L1 in human) and G protein coupled estrogen receptor 1 (Gper1 – homologous to GPER1 in human) up-regulated in testes of rats with induced varicocele, which was confirmed by Western blotting analysis. These proteins are linked to reduced expression of androgen receptor (AR), suggesting that varicocele affects mechanisms that control AR expression and function.86
FINAL REMARKS/EXPERT COMMENTARY
Seminal plasma is a complex mixture with different origins86 and differences in composition that may arise from various reasons. Isolating factors that arise from fertility-related causes, such as molecular pathways of sperm function, is not easily achieved, because many diverse conditions potentially alter simply the contribution of individual components to the seminal plasma, such as alteration in seminal vesicle or prostate function.68 This is an important challenge faced by studies in seminal plasma proteomics – understanding the origin of the studied proteome. Furthermore, semen is immediately coagulated upon ejaculation, followed by proteolytic activity leading to liquefaction.87,88 Thus, when the sample is made available for studies, important protease and phosphatase activity has usually already taken place, which in turn affects the ability of current studies to observe peptides and their posttranslational modifications.89
Varicocele remains an elusive disease, in that quite often it will be detected in fertile men.10 Differentiating a fertility-compromising varicocele, on the other hand, is imperative for the general medical practice, if intervention in patients who indeed require an intervention is to be achieved. Patient (and disease) individualization is one of the important promises proteomics studies bring,24,90 and in the case of varicocele it may assist in differentiating deleterious from “silent” varicoceles.
In general, it seems that varicocele leads to a state of homeorhesis, or a deviation from homeostasis toward a dynamic equilibrium in an altered state, characterized by enrichment of functions such as immune response and apoptosis, somewhat competing with important functions (such as fertilization, motility, and zona binding).76 In adolescents, for example, correcting varicocele shifts the seminal plasma proteome back to its regular state, enriching expected functions for semen associated to preserved reproductive function. In adults, a general dysfunction is observed in the seminal plasma proteome, and functions important for fertilization are underrepresented in these patients.
Defining end-points in studies regarding the seminal plasma proteome is also important, as these will, in essence, impact the importance of the findings. Simply utilizing semen analysis or testicular volume to separate groups may lead findings to fall short in sensitivity. On the other hand, defining fertility as an end-point will insert variability from the female partner into studies, which may limit their applicability. Under the same rationale, defining what is to be studied is just as important. If, say, one is interested in studying testicular function, normalizing the quantitative proteomics studies to constitutive testicular proteins may be fundamental to focus in on proteins that reflect the testicular and epididymal environments. Studying epididymal microvesicles and exosomes (epididymosomes) is also a novel approach that has been producing interesting results, because epididymal fluid is an important environment to transfer proteins participating in key sperm events (including capacitation), which has been shown in animals and in humans.91,92,93 ESPB1 and BSP1, for example, bind to specific phospholipids in the sperm membrane (presenting a phosphorylcholine moiety) in order to localize to specific sites, where it will mark dead or altered sperm.94 In humans, epididymosomes have been shown to present proteins with a diverse array of functions, namely as signal transducers, adhesion, transporter and protein trafficking, defense, chaperones, enzymes and structural proteins. RAB proteins, for example, seem to participate in determining membrane location targeting for individual epididymosomes proteins. Because these are docked onto membranes and form an integral part of the mature sperm membrane, epididymosomes transfer of proteins characterizes an important step in sperm maturation.92
As the seminal plasma proteomics data are amounting to the point that tens of thousands of peptides and thousands of proteins are observed in small portions of the seminal plasma, it will be essential to bear in mind that (i) these studies originate from a clinical setting and produce results with a clinical impact, based on medical information and a true Cartesian hypothesis, and (ii) data filtering and analysis will be of essence if these data are to be transformed into actionable information, such as protein diagnostic and therapeutic targets. Functional studies that characterize the events downstream of protein pathways will further help in determining biomarkers of male fertility.95 This translates to the fact that, as is the case with medicine, in general, the study of varicocele and male infertility will require a multidisciplinary effort and an integrative approach toward data analysis. It may be that the understanding of varicocele and its underlying molecular mechanisms is more at hand now than it ever was, as long as all the generated data don’t get in the way!
AUTHOR CONTRIBUTIONS
All the authors contributed equally in the planning, literature review, drafting, writing, revision, and approval of this manuscript.
COMPETING INTERESTS
None of the authors declare competing financial interests.
ACKNOWLEDGMENTS
The authors wish to acknowledge support from the Sao Paulo Research Foundation - FAPESP (process number 2012/15039-7, 2014/11493-0 and 2014/17185-6) and from the National Council for Scientific and Technological Development (CNPq, process 306616/2013-0).
Supplementary information is linked to the online version of the paper on the Asian Journal of Andrology website.
REFERENCES
- 1.Brugh VM, Matschke HM, Lipshultz LI. Male factor infertility. Endocrinol Metab Clin North Am. 2003;32:689–707. doi: 10.1016/s0889-8529(03)00047-1. [DOI] [PubMed] [Google Scholar]
- 2.Gorelick JI, Goldstein M. Loss of fertility in men with varicocele. Fertil Steril. 1993;59:613–6. [PubMed] [Google Scholar]
- 3.Witt MA, Lipshultz LI. Varicocele: a progressive or static lesion? Urology. 1993;42:541–3. doi: 10.1016/0090-4295(93)90268-f. [DOI] [PubMed] [Google Scholar]
- 4.Steeno O, Knops J, Declerck L, Adimoelja A, van de Voorde H. Prevention of fertility disorders by detection and treatment of varicocele at school and college age. Andrologia. 1976;8:47–53. doi: 10.1111/j.1439-0272.1976.tb01645.x. [DOI] [PubMed] [Google Scholar]
- 5.Marmar JL, DeBenedictis TJ, Praiss D. The management of varicoceles by microdissection of the spermatic cord at the external inguinal ring. Fertil Steril. 1985;43:583–8. doi: 10.1016/s0015-0282(16)48501-8. [DOI] [PubMed] [Google Scholar]
- 6.Lacerda JI, Del Giudice PT, da Silva BF, Nichi M, Fariello RM, et al. Adolescent varicocele: improved sperm function after varicocelectomy. Fertil Steril. 2011;95:994–9. doi: 10.1016/j.fertnstert.2010.10.031. [DOI] [PubMed] [Google Scholar]
- 7.Schlegel PN. Is assisted reproduction the optimal treatment for varicocele-associated male infertility?. A cost-effectiveness analysis. Urology. 1997;49:83–90. doi: 10.1016/S0090-4295(96)00379-2. [DOI] [PubMed] [Google Scholar]
- 8.Nieschlag E, Hertle L, Fischedick A, Abshagen K, Behre HM. Update on treatment of varicocele: counselling as effective as occlusion of the vena spermatica. Hum Reprod Oxf Engl. 1998;13:2147–50. doi: 10.1093/humrep/13.8.2147. [DOI] [PubMed] [Google Scholar]
- 9.Madgar I, Weissenberg R, Lunenfeld B, Karasik A, Goldwasser B. Controlled trial of high spermatic vein ligation for varicocele in infertile men. Fertil Steril. 1995;63:120–4. doi: 10.1016/s0015-0282(16)57306-3. [DOI] [PubMed] [Google Scholar]
- 10.Pinto KJ, Kroovand RL, Jarow JP. Varicocele related testicular atrophy and its predictive effect upon fertility. J Urol. 1994;152:788–90. doi: 10.1016/s0022-5347(17)32710-6. [DOI] [PubMed] [Google Scholar]
- 11.Masson P, Brannigan RE. The varicocele. Urol Clin North Am. 2014;41:129–44. doi: 10.1016/j.ucl.2013.08.001. [DOI] [PubMed] [Google Scholar]
- 12.Lima SB, Cenedeze MA, Bertolla RP, Filho PA, Oehninger S, et al. Expression of the HSPA2 gene in ejaculated spermatozoa from adolescents with and without varicocele. Fertil Steril. 2006;86:1659–63. doi: 10.1016/j.fertnstert.2006.05.030. [DOI] [PubMed] [Google Scholar]
- 13.Del Giudice PT, Lima SB, Cenedeze MA, Pacheco-Silva A, Bertolla RP, et al. Expression of the Fas-ligand gene in ejaculated sperm from adolescents with and without varicocele. J Assist Reprod Genet. 2010;27:103–9. doi: 10.1007/s10815-010-9384-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Kiziler AR, Aydemir B, Guzel S, Yazici CM, Gulyasar T, et al. Comparison of before and after varicocelectomy levels of trace elements, nitric oxide, asymmetric dimethylarginine and malondialdehyde in the seminal plasma and peripheral and spermatic veins. Biol Trace Elem Res. 2015;167:172–8. doi: 10.1007/s12011-015-0303-x. [DOI] [PubMed] [Google Scholar]
- 15.Blumer CG, Restelli AE, Giudice PT, Soler TB, Fraietta R, et al. Effect of varicocele on sperm function and semen oxidative stress. BJU Int. 2012;109:259–65. doi: 10.1111/j.1464-410X.2011.10240.x. [DOI] [PubMed] [Google Scholar]
- 16.Bertolla RP, Cedenho AP, Hassun Filho PA, Lima SB, Ortiz V, et al. Sperm nuclear DNA fragmentation in adolescents with varicocele. Fertil Steril. 2006;85:625–8. doi: 10.1016/j.fertnstert.2005.08.032. [DOI] [PubMed] [Google Scholar]
- 17.Pajovic B, Radojevic N, Terzic N, Dimitrovski A. Correlation between nitric oxide levels, the hypo-osmotic swelling test for sperm membranes and semen analysis in patients with varicocele. Scand J Urol. 2013;47:404–10. doi: 10.3109/21681805.2013.769459. [DOI] [PubMed] [Google Scholar]
- 18.Blumer CG, Fariello RM, Restelli AE, Spaine DM, Bertolla RP, et al. Sperm nuclear DNA fragmentation and mitochondrial activity in men with varicocele. Fertil Steril. 2008;90:1716–22. doi: 10.1016/j.fertnstert.2007.09.007. [DOI] [PubMed] [Google Scholar]
- 19.Sheehan MM, Ramasamy R, Lamb DJ. Molecular mechanisms involved in varicocele-associated infertility. J Assist Reprod Genet. 2014;31:521–6. doi: 10.1007/s10815-014-0200-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Batruch I, Smith CR, Mullen BJ, Grober E, Lo KC, et al. Analysis of seminal plasma from patients with non-obstructive azoospermia and identification of candidate biomarkers of male infertility. J Proteome Res. 2012;11:1503–11. doi: 10.1021/pr200812p. [DOI] [PubMed] [Google Scholar]
- 21.Batruch I, Lecker I, Kagedan D, Smith CR, Mullen BJ, et al. Proteomic analysis of seminal plasma from normal volunteers and post-vasectomy patients identifies over 2000 proteins and candidate biomarkers of the urogenital system. J Proteome Res. 2011;10:941–53. doi: 10.1021/pr100745u. [DOI] [PubMed] [Google Scholar]
- 22.Pennington SR, Wilkins MR, Hochstrasser DF, Dunn MJ. Proteome analysis: from protein characterization to biological function. Trends Cell Biol. 1997;7:168–73. doi: 10.1016/S0962-8924(97)01033-7. [DOI] [PubMed] [Google Scholar]
- 23.Zhang Y, Fonslow BR, Shan B, Baek MC, Yates JR. Protein analysis by shotgun/bottom-up proteomics. Chem Rev. 2013;113:2343–94. doi: 10.1021/cr3003533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hood L, Flores M. A personal view on systems medicine and the emergence of proactive P4 medicine: predictive, preventive, personalized and participatory. N Biotechnol. 2012;29:613–24. doi: 10.1016/j.nbt.2012.03.004. [DOI] [PubMed] [Google Scholar]
- 25.Liu Y, Qing H, Deng Y. Biomarkers in Alzheimer's disease analysis by mass spectrometry-based proteomics. Int J Mol Sci. 2014;15:7865–82. doi: 10.3390/ijms15057865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Li JJ, Qi RZ, Ng GK, Xie D. Proteomics in gastric cancer research: benefits and challenges. Proteomics Clin Appl. 2009;3:185–96. doi: 10.1002/prca.200800151. [DOI] [PubMed] [Google Scholar]
- 27.Cutillas PR. Role of phosphoproteomics in the development of personalized cancer therapies. Proteomics Clin Appl. 2015;9:383–95. doi: 10.1002/prca.201400104. [DOI] [PubMed] [Google Scholar]
- 28.Bantscheff M, Lemeer S, Savitski MM, Kuster B. Quantitative mass spectrometry in proteomics: critical review update from 2007 to the present. Anal Bioanal Chem. 2012;404:939–65. doi: 10.1007/s00216-012-6203-4. [DOI] [PubMed] [Google Scholar]
- 29.Mann T. London, New York: John Wiley and Sons, Methuen and Co; 1954. The Biochemistry of Semen; p. 280. [Google Scholar]
- 30.Aitken RJ, Baker MA. The role of proteomics in understanding sperm cell biology. Int J Androl. 2008;31:295–302. doi: 10.1111/j.1365-2605.2007.00851.x. [DOI] [PubMed] [Google Scholar]
- 31.Scherl A. Clinical protein mass spectrometry. Methods San Diego Calif. 2015;81:3–14. doi: 10.1016/j.ymeth.2015.02.015. [DOI] [PubMed] [Google Scholar]
- 32.Whitehouse CM, Dreyer RN, Yamashita M, Fenn JB. Electrospray interface for liquid chromatographs and mass spectrometers. Anal Chem. 1985;57:675–9. doi: 10.1021/ac00280a023. [DOI] [PubMed] [Google Scholar]
- 33.Kim ST, Kim HS, Kim HJ, Kim SG, Kang SY, et al. Prefractionation of protein samples for proteome analysis by sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Mol Cells. 2003;16:316–22. [PubMed] [Google Scholar]
- 34.Pernemalm M, Lehtiö J. A novel prefractionation method combining protein and peptide isoelectric focusing in immobilized pH gradient strips. J Proteome Res. 2013;12:1014–9. doi: 10.1021/pr300817y. [DOI] [PubMed] [Google Scholar]
- 35.Lange V, Picotti P, Domon B, Aebersold R. Selected reaction monitoring for quantitative proteomics: a tutorial. Mol Syst Biol. 2008;4:222. doi: 10.1038/msb.2008.61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Henault MA, Killian GJ. Effect of homologous and heterologous seminal plasma on the fertilizing ability of ejaculated bull spermatozoa assessed by penetration of zona-free bovine oocytes. J Reprod Fertil. 1996;108:199–204. doi: 10.1530/jrf.0.1080199. [DOI] [PubMed] [Google Scholar]
- 37.Brackett NL, Davi RC, Padron OF, Lynne CM. Seminal plasma of spinal cord injured men inhibits sperm motility of normal men. J Urol. 1996;155:1632–5. [PubMed] [Google Scholar]
- 38.Rodriguez-Martinez H, Saravia F, Wallgren M, Martinez EA, Sanz L, et al. Spermadhesin PSP-I/PSP-II heterodimer induces migration of polymorphonuclear neutrophils into the uterine cavity of the sow. J Reprod Immunol. 2010;84:57–65. doi: 10.1016/j.jri.2009.10.007. [DOI] [PubMed] [Google Scholar]
- 39.Pilch B, Mann M. Large-scale and high-confidence proteomic analysis of human seminal plasma. Genome Biol. 2006;7:R40. doi: 10.1186/gb-2006-7-5-r40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Kumar V, Hassan MI, Tomar AK, Kashav T, Nautiyal J, et al. Proteomic analysis of heparin-binding proteins from human seminal plasma: a step towards identification of molecular markers of male fertility. J Biosci. 2009;34:899–908. doi: 10.1007/s12038-009-0104-5. [DOI] [PubMed] [Google Scholar]
- 41.Wang J, Wang J, Zhang HR, Shi HJ, Ma D, et al. Proteomic analysis of seminal plasma from asthenozoospermia patients reveals proteins that affect oxidative stress responses and semen quality. Asian J Androl. 2009;11:484–91. doi: 10.1038/aja.2009.26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Zhao C, Huo R, Wang FQ, Lin M, Zhou ZM, et al. Identification of several proteins involved in regulation of sperm motility by proteomic analysis. Fertil Steril. 2007;87:436–8. doi: 10.1016/j.fertnstert.2006.06.057. [DOI] [PubMed] [Google Scholar]
- 43.Ding Z, Qu F, Guo W, Ying X, Wu M, et al. Identification of sperm forward motility-related proteins in human seminal plasma. Mol Reprod Dev. 2007;74:1124–31. doi: 10.1002/mrd.20624. [DOI] [PubMed] [Google Scholar]
- 44.Miller DJ, Winer MA, Ax RL. Heparin-binding proteins from seminal plasma bind to bovine spermatozoa and modulate capacitation by heparin. Biol Reprod. 1990;42:899–915. doi: 10.1095/biolreprod42.6.899. [DOI] [PubMed] [Google Scholar]
- 45.Primakoff P, Myles DG. Penetration, adhesion, and fusion in mammalian sperm-egg interaction. Science. 2002;296:2183–5. doi: 10.1126/science.1072029. [DOI] [PubMed] [Google Scholar]
- 46.Moldenhauer LM, Diener KR, Thring DM, Brown MP, Hayball JD, et al. Cross-presentation of male seminal fluid antigens elicits T cell activation to initiate the female immune response to pregnancy. J Immunol. 2009;182:8080–93. doi: 10.4049/jimmunol.0804018. [DOI] [PubMed] [Google Scholar]
- 47.Rodríguez-Martínez H, Kvist U, Ernerudh J, Sanz L, Calvete JJ. Seminal plasma proteins: what role do they play? Am J Reprod Immunol. 2011;66(Suppl 1):11–22. doi: 10.1111/j.1600-0897.2011.01033.x. [DOI] [PubMed] [Google Scholar]
- 48.Mann T, Lutwak-Mann C. Male reproductive function and semen. Andrologia. 1982;14:76. [Google Scholar]
- 49.Duncan MW, Thompson HS. Proteomics of semen and its constituents. Proteomics Clin Appl. 2007;1:861–75. doi: 10.1002/prca.200700228. [DOI] [PubMed] [Google Scholar]
- 50.Burden HP, Holmes CH, Persad R, Whittington K. Prostasomes – Their effects on human male reproduction and fertility. Hum Reprod Update. 2006;12:283–92. doi: 10.1093/humupd/dmi052. [DOI] [PubMed] [Google Scholar]
- 51.Utleg AG, Yi EC, Xie T, Shannon P, White JT, et al. Proteomic analysis of human prostasomes. Prostate. 2003;56:150–61. doi: 10.1002/pros.10255. [DOI] [PubMed] [Google Scholar]
- 52.Milardi D, Grande G, Vincenzoni F, Messana I, Pontecorvi A, et al. Proteomic approach in the identification of fertility pattern in seminal plasma of fertile men. Fertil Steril. 2012;97:67–73. doi: 10.1016/j.fertnstert.2011.10.013. [DOI] [PubMed] [Google Scholar]
- 53.Intasqui P, Camargo M, Del Giudice PT, Spaine DM, Carvalho VM, et al. Sperm nuclear DNA fragmentation rate is associated with differential protein expression and enriched functions in human seminal plasma. BJU Int. 2013;112:835–43. doi: 10.1111/bju.12233. [DOI] [PubMed] [Google Scholar]
- 54.Fung KY, Glode LM, Green S, Duncan MW. A comprehensive characterization of the peptide and protein constituents of human seminal fluid. Prostate. 2004;61:171–81. doi: 10.1002/pros.20089. [DOI] [PubMed] [Google Scholar]
- 55.Fu-Jun L, Xiao-Fang S. Comparative analysis of human reproductive proteomes identifies candidate proteins of sperm maturation. Mol Biol Rep. 2012;39:10257–63. doi: 10.1007/s11033-012-1902-7. [DOI] [PubMed] [Google Scholar]
- 56.Rolland AD, Lavigne R, Dauly C, Calvel P, Kervarrec C, et al. Identification of genital tract markers in the human seminal plasma using an integrative genomics approach. Hum Reprod Oxf Engl. 2013;28:199–209. doi: 10.1093/humrep/des360. [DOI] [PubMed] [Google Scholar]
- 57.Drabovich AP, Saraon P, Jarvi K, Diamandis EP. Seminal plasma as a diagnostic fluid for male reproductive system disorders. Nat Rev Urol. 2014;11:278–88. doi: 10.1038/nrurol.2014.74. [DOI] [PubMed] [Google Scholar]
- 58.Starita-Geribaldi M, Poggioli S, Zucchini M, Garin J, Chevallier D, et al. Mapping of seminal plasma proteins by two-dimensional gel electrophoresis in men with normal and impaired spermatogenesis. Mol Hum Reprod. 2001;7:715–22. doi: 10.1093/molehr/7.8.715. [DOI] [PubMed] [Google Scholar]
- 59.Starita-Geribaldi M, Roux F, Garin J, Chevallier D, Fénichel P, et al. Development of narrow immobilized pH gradients covering one pH unit for human seminal plasma proteomic analysis. Proteomics. 2003;3:1611–9. doi: 10.1002/pmic.200300493. [DOI] [PubMed] [Google Scholar]
- 60.Yamakawa K, Yoshida K, Nishikawa H, Kato T, Iwamoto T. Comparative analysis of interindividual variations in the seminal plasma proteome of fertile men with identification of potential markers for azoospermia in infertile patients. J Androl. 2007;28:858–65. doi: 10.2164/jandrol.107.002824. [DOI] [PubMed] [Google Scholar]
- 61.Drabovich AP, Jarvi K, Diamandis EP. Verification of male infertility biomarkers in seminal plasma by multiplex selected reaction monitoring assay. Mol Cell Proteomics. 2011;10:M110004127. doi: 10.1074/mcp.M110.004127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Drabovich AP, Dimitromanolakis A, Saraon P, Soosaipillai A, Batruch I, et al. Differential diagnosis of azoospermia with proteomic biomarkers ECM1 and TEX101 quantified in seminal plasma. Sci Transl Med. 2013;5:212ra160. doi: 10.1126/scitranslmed.3006260. [DOI] [PubMed] [Google Scholar]
- 63.Freour T, Com E, Barriere P, Bouchot O, Jean M, et al. Comparative proteomic analysis coupled with conventional protein assay as a strategy to identify predictors of successful testicular sperm extraction in patients with non-obstructive azoospermia. Andrology. 2013;1:414–20. doi: 10.1111/j.2047-2927.2012.00059.x. [DOI] [PubMed] [Google Scholar]
- 64.Fariello RM, Pariz JR, Spaine DM, Gozzo FC, Pilau EJ, et al. Effect of smoking on the functional aspects of sperm and seminal plasma protein profiles in patients with varicocele. Hum Reprod Oxf Engl. 2012;27:3140–9. doi: 10.1093/humrep/des287. [DOI] [PubMed] [Google Scholar]
- 65.Davalieva K, Kiprijanovska S, Noveski P, Plaseski T, Kocevska B, et al. Proteomic analysis of seminal plasma in men with different spermatogenic impairment. Andrologia. 2012;44:256–64. doi: 10.1111/j.1439-0272.2012.01275.x. [DOI] [PubMed] [Google Scholar]
- 66.Herwig R, Knoll C, Planyavsky M, Pourbiabany A, Greilberger J, et al. Proteomic analysis of seminal plasma from infertile patients with oligoasthenoteratozoospermia due to oxidative stress and comparison with fertile volunteers. Fertil Steril. 2013;100:355–66. doi: 10.1016/j.fertnstert.2013.03.048. [DOI] [PubMed] [Google Scholar]
- 67.Sharma R, Agarwal A, Mohanty G, Jesudasan R, Gopalan B, et al. Functional proteomic analysis of seminal plasma proteins in men with various semen parameters. Reprod Biol Endocrinol. 2013;11:38. doi: 10.1186/1477-7827-11-38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.da Silva BF, Souza GH, Lo Turco EG, Del Giudice PT, Soler TB, et al. Differential seminal plasma proteome according to semen retrieval in men with spinal cord injury. Fertil Steril. 2013;100:959–69. doi: 10.1016/j.fertnstert.2013.06.009. [DOI] [PubMed] [Google Scholar]
- 69.Milardi D, Grande G, Vincenzoni F, Giampietro A, Messana I, et al. Novel biomarkers of androgen deficiency from seminal plasma profiling using high-resolution mass spectrometry. J Clin Endocrinol Metab. 2014;99:2813–20. doi: 10.1210/jc.2013-4148. [DOI] [PubMed] [Google Scholar]
- 70.Sharma R, Agarwal A, Mohanty G, du Plessis SS, Gopalan B, et al. Proteomic analysis of seminal fluid from men exhibiting oxidative stress. Reprod Biol Endocrinol. 2013;11:85. doi: 10.1186/1477-7827-11-85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Intasqui P, Antoniassi MP, Camargo M, Nichi M, Carvalho VM, et al. Differences in the seminal plasma proteome are associated with oxidative stress levels in men with normal semen parameters. Fertil Steril. 2015;104:292–301. doi: 10.1016/j.fertnstert.2015.04.037. [DOI] [PubMed] [Google Scholar]
- 72.Cadavid J AP, Alvarez A, Markert UR, Cardona Maya W. Differential protein expression in seminal plasma from fertile and infertile males. J Hum Reprod Sci. 2014;7:206–11. doi: 10.4103/0974-1208.142485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Gilany K, Minai-Tehrani A, Savadi-Shiraz E, Rezadoost H, Lakpour N. Exploring the human seminal plasma proteome: an unexplored gold mine of biomarker for male infertility and male reproduction disorder. J Reprod Infertil. 2015;16:61–71. [PMC free article] [PubMed] [Google Scholar]
- 74.Camargo M, Intasqui Lopes P, Del Giudice PT, Carvalho VM, Cardozo KH, et al. Unbiased label-free quantitative proteomic profiling and enriched proteomic pathways in seminal plasma of adult men before and after varicocelectomy. Hum Reprod Oxf Engl. 2013;28:33–46. doi: 10.1093/humrep/des357. [DOI] [PubMed] [Google Scholar]
- 75.Zylbersztejn DS, Andreoni C, Del Giudice PT, Spaine DM, Borsari L, et al. Proteomic analysis of seminal plasma in adolescents with and without varicocele. Fertil Steril. 2013;99:92–8. doi: 10.1016/j.fertnstert.2012.08.048. [DOI] [PubMed] [Google Scholar]
- 76.Del Giudice PT, da Silva BF, Lo Turco EG, Fraietta R, Spaine DM, et al. Changes in the seminal plasma proteome of adolescents before and after varicocelectomy. Fertil Steril. 2013;100:667–72. doi: 10.1016/j.fertnstert.2013.04.036. [DOI] [PubMed] [Google Scholar]
- 77.Lilja H, Oldbring J, Rannevik G, Laurell CB. Seminal vesicle-secreted proteins and their reactions during gelation and liquefaction of human semen. J Clin Invest. 1987;80:281–5. doi: 10.1172/JCI113070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Hosseinifar H, Gourabi H, Salekdeh GH, Alikhani M, Mirshahvaladi S, et al. Study of sperm protein profile in men with and without varicocele using two-dimensional gel electrophoresis. Urology. 2013;81:293–300. doi: 10.1016/j.urology.2012.06.027. [DOI] [PubMed] [Google Scholar]
- 79.Chan CC, Sun GH, Shui HA, Wu GJ. Differential spermatozoal protein expression profiles in men with varicocele compared to control subjects: upregulation of heat shock proteins 70 and 90 in varicocele. Urology. 2013;81:1379.e1–8. doi: 10.1016/j.urology.2013.01.031. [DOI] [PubMed] [Google Scholar]
- 80.Hosseinifar H, Sabbaghian M, Nasrabadi D, Modarresi T, Dizaj AV, et al. Study of the effect of varicocelectomy on sperm proteins expression in patients with varicocele and poor sperm quality by using two-dimensional gel electrophoresis. J Assist Reprod Genet. 2014;31:725–9. doi: 10.1007/s10815-014-0209-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Agarwal A, Sharma R, Durairajanayagam D, Ayaz A, Cui Z, et al. Major protein alterations in spermatozoa from infertile men with unilateral varicocele. Reprod Biol Endocrinol. 2015;13:8. doi: 10.1186/s12958-015-0007-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Agarwal A, Sharma R, Durairajanayagam D, Cui Z, Ayaz A, et al. Spermatozoa protein alterations in infertile men with bilateral varicocele. Asian J Androl. 2015 doi: 10.4103/1008-682X.153848. doi: 10.4103/1008-682X.153848. [Epub ahead of print] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Agarwal A, Sharma R, Durairajanayagam D, Cui Z, Ayaz A, et al. Differential proteomic profiling of spermatozoal proteins of infertile men with unilateral or bilateral varicocele. Urology. 2015;85:580–8. doi: 10.1016/j.urology.2014.11.030. [DOI] [PubMed] [Google Scholar]
- 84.Soares TS, Fernandes SA, Lima ML, Stumpp T, Schoorlemmer GH, et al. Experimental varicocoele in rats affects mechanisms that control expression and function of the androgen receptor. Andrology. 2013;1:670–81. doi: 10.1111/j.2047-2927.2013.00103.x. [DOI] [PubMed] [Google Scholar]
- 85.Lucas TF, Royer C, Siu ER, Lazari MF, Porto CS. Expression and signaling of G protein-coupled estrogen receptor 1 (GPER) in rat Sertoli cells. Biol Reprod. 2010;83:307–17. doi: 10.1095/biolreprod.110.084160. [DOI] [PubMed] [Google Scholar]
- 86.Plant TM, Zeleznik AJ. Academic Press; 2014. Knobil and Neill's Physiology of Reproduction: Two-Volume Set; p. 2708. [Google Scholar]
- 87.Robert M, Gagnon C. Semenogelin I: a coagulum forming, multifunctional seminal vesicle protein. Cell Mol Life Sci. 1999;55:944–60. doi: 10.1007/s000180050346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.KLK3 – Prostate-specific antigen precursor – Homo sapiens (Human) [Internet] [Last accessed on 2015 Jul 13]. Available from: http://www.uniprot.org/uniprot/P07288 .
- 89.Mann M, Jensen ON. Proteomic analysis of post-translational modifications. Nat Biotechnol. 2003;21:255–61. doi: 10.1038/nbt0303-255. [DOI] [PubMed] [Google Scholar]
- 90.Merelli I, Pérez-Sánchez H, Gesing S, D’Agostino D. Managing, analysing, and integrating big data in medical bioinformatics: open problems and future perspectives. Biomed Res Int 2014. 2014:134023. doi: 10.1155/2014/134023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.D’Amours O, Frenette G, Bordeleau LJ, Allard N, Leclerc P, et al. Epididymosomes transfer epididymal sperm binding protein 1 (ELSPBP1) to dead spermatozoa during epididymal transit in bovine. Biol Reprod. 2012;87:94. doi: 10.1095/biolreprod.112.100990. [DOI] [PubMed] [Google Scholar]
- 92.Thimon V, Frenette G, Saez F, Thabet M, Sullivan R. Protein composition of human epididymosomes collected during surgical vasectomy reversal: a proteomic and genomic approach. Hum Reprod Oxf Engl. 2008;23:1698–707. doi: 10.1093/humrep/den181. [DOI] [PubMed] [Google Scholar]
- 93.Sullivan R, Saez F. Epididymosomes, prostasomes, and liposomes: their roles in mammalian male reproductive physiology. Reproduction. 2013;146:R21–35. doi: 10.1530/REP-13-0058. [DOI] [PubMed] [Google Scholar]
- 94.D’Amours O, Bordeleau L-J, Frenette G, Blondin P, Leclerc P, et al. Binder of sperm 1 and epididymal sperm binding protein 1 are associated with different bull sperm subpopulations. Reproduction. 2012;143:759–71. doi: 10.1530/REP-11-0392. [DOI] [PubMed] [Google Scholar]
- 95.Milardi D, Grande G, Vincenzoni F, Castagnola M, Marana R. Proteomics of human seminal plasma: identification of biomarker candidates for fertility and infertility and the evolution of technology. Mol Reprod Dev. 2013;80:350–7. doi: 10.1002/mrd.22178. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Comparative seminal plasma proteomics studies in male infertility