

Abstract
Yeasts are unicellular fungi that do not form fruiting bodies. Although the yeast lifestyle has evolved multiple times, most known species belong to the subphylum Saccharomycotina (syn. Hemiascomycota, hereafter yeasts). This diverse group includes the premier eukaryotic model system, Saccharomyces cerevisiae; the common human commensal and opportunistic pathogen, Candida albicans; and over 1,000 other known species (with more continuing to be discovered). Yeasts are found in every biome and continent and are more genetically diverse than angiosperms or chordates. Ease of culture, simple life cycles, and small genomes (~10–20 Mbp) have made yeasts exceptional models for molecular genetics, biotechnology, and evolutionary genomics. Here we discuss recent developments in understanding the genomic underpinnings of the making of yeast biodiversity, comparing and contrasting natural and human-associated evolutionary processes. Only a tiny fraction of yeast biodiversity and metabolic capabilities has been tapped by industry and science. Expanding the taxonomic breadth of deep genomic investigations will further illuminate how genome function evolves to encode their diverse metabolisms and ecologies.
Graphical abstract
A brief history of yeast evolutionary genomics
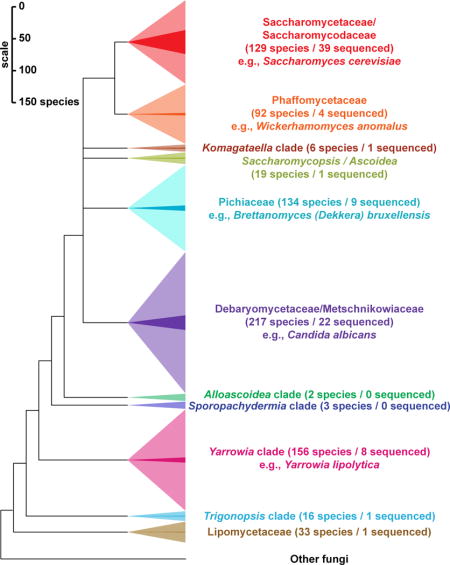
The Saccharomyces cerevisiae genome was the first eukaryotic genome sequenced [1], a collaborative feat that enabled two decades of innovation and discovery. C. albicans soon followed [2], as well as a handful of relatives in both clades and a smattering of taxonomically diverse species [3–8]. S. cerevisiae became a proving ground for new genomic technologies, such as deleting and barcoding genes for functional profiling [9]. Early comparative studies pioneered the now commonplace use of genome sequencing to address specific functional and evolutionary hypotheses, such as using conserved genomic regions to identify functional DNA sequence elements [3,4]. These genomes began to catalyze research in many other fields, resulting in important advances to our understanding of the evolution of genome content and organismal traits [10,11], phylogenetics [12], and cis-regulatory element prediction [13]. By the last comprehensive review of yeast evolutionary genomics in 2010 [14], the genomes of 26 yeast species had been sequenced, still mostly in the two clades containing S. cerevisiae and C. albicans. With the popularization of next-generation sequencing, the genomes of 40 additional species have been published in the last five years, while 20 more are publicly available but await formal publications (Figure 1). Nonetheless, most yeast biodiversity remains unexplored at the genomic level (Figure 2).
Figure 1. Key traits and phylogenetic relationships of the 86 yeasts of the subphylum Saccharomycotina whose genomes have been sequenced.

The topology of the cladogram has been estimated conservatively from previous analyses using genome [12,23] or multi-locus sequence data [27,36]. Major clades [36] are color-coded (clade names are shown in Figure 2). Only one reference genome per species is included with preference given to the highest quality and/or most widely used reference genome. Only publicly available genome assemblies are included. Interspecies hybrids are discussed in the text but are not shown here. WGD, whole genome duplication; recent work has shown that the WGD was caused by an allopolyploidization event that occurred between an early member of the Zygosaccharomyces/Torulaspora clade and an early member of the Kluyveromyces/Lachancea/Eremothecium clade, thus making this part of the phylogeny a network, rather than a tree [73]. CTG clade, yeasts using an alternate codon table where CTG encodes serine, instead of leucine. JGI, genomes publicly available on MycoCosm at http://genome.jgi-psf.org/programs/fungi/index.jsf, which are subject to the usage terms of the DOE Joint Genome Institute until formal publication. GB1, GenBank Accession CCBQ000000000; GB2, Genbank Accession JPPO00000000; GB3, GenBank Accession AEOI00000000; GB4, GenBank Accession LCTY00000000.
Figure 2. Most yeast clades remain underrepresented by genome sequence data.
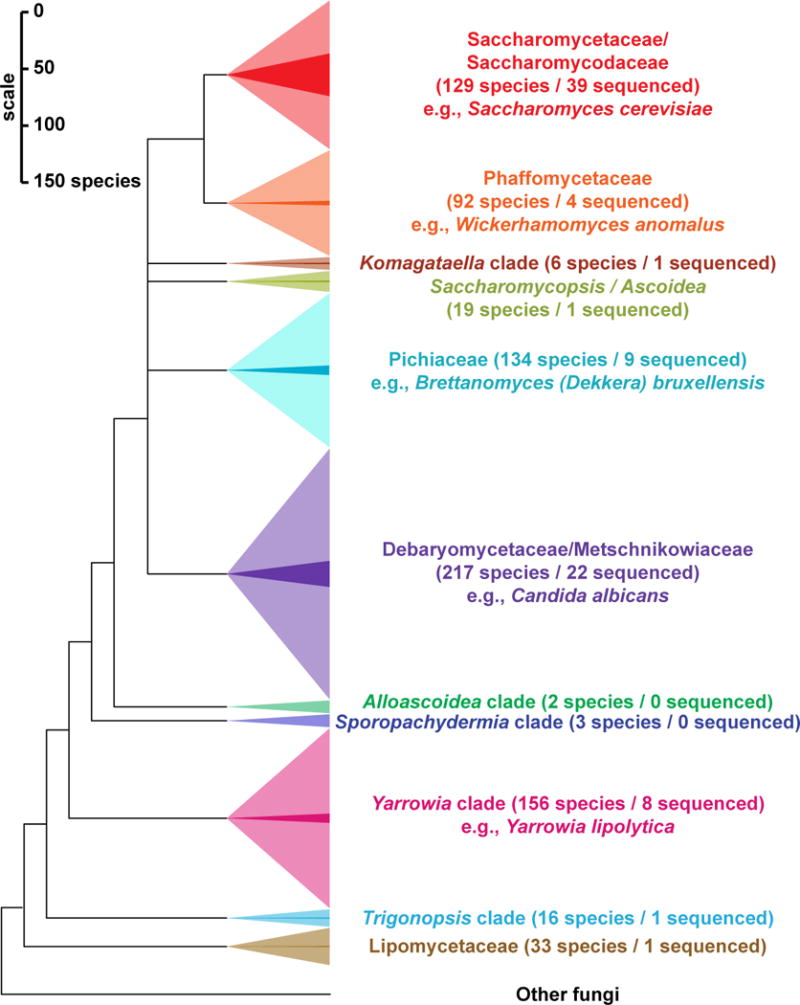
Estimated species counts for each major yeast clade [36] (light colors) are compared with the number of publicly available genomes from each clade (dark sliver within larger triangles). In addition to the 807 species shown, approximately 400 anamorphic species (e.g. Candida spp.) currently lack a clear phylogenetic placement. As the phylogeny is resolved, these species will be reassigned to genera and higher taxonomic ranks consistent with their phylogeny (Box 1). Color codes are the same as in Figure 1.
Yeast mating systems
Most yeasts have true sexual cycles, but some manifest complex parasexual cycles (non-meiotic processes involving mitotic recombination and chromosome loss), whereas others are thought to be asexual. This multiplicity of forms and sexual cycles led to a byzantine nomenclature system that is undergoing a radical simplification, which will ultimately result in a taxonomy based on phylogeny (Box 1). Although asexual strains achieve higher laboratory fitness [15], they seem not to persist or undergo frequent cladogenesis. Sex may increase the rate of adaptation to novel environments and help purge deleterious alleles [16,17]. Thus, most yeast clades harbor at least some sexual taxa, which likely reflect their ancestral states. Sexual cycles exist in both obligately outcrossing (heterothallic) species, where mating types reside in different individuals, as well as in species that can self-fertilize (homothallic).
Box 1. One Fungus, One Name (1F1N) and the Phylogenomic Future of Yeast Taxonomy.
Despite the considerable progress made classifying yeasts using multi-locus DNA sequence data, critical gaps remain. Many genera are paraphyletic or polyphyletic, while many taxonomic circumscriptions at or above the family level are poorly supported or completely lacking. Further complicating matters, many species are assigned to large, polyphyletic, anamorphic (no known sexual state) genera (e.g. Candida), rather than to allied teleomorphic (sexual) genera. Due to a recent revision in the International Code of Nomenclature for algae, fungi, and plants (formerly the International Code of Botanical Nomenclature), anamorphs and teleomorphs can now, for the first time, be reassigned to monophyletic genera that contain species both with and without known sexual cycles. This long-overdue change presents a timely opportunity to formulate a comprehensive and stable yeast taxonomy based on complete genetic data. Well-circumscribed monophyletic genera (and higher order taxa) will finally provide evolutionary geneticists, mycologists, biotechnologists, and clinicians with phylogenetically informative names to aid in the design of experiments and the interpretation of data. In this review, the current, formally recognized genus names are shown first with alternative names in parentheses. In some cases, such as the affinity of C. glabrata with Nakaseomyces, there is strong phylogenomic support and a virtual certainty that the species will be ultimately reassigned to a particular genus; in other cases, phylogenomic analyses and further taxonomic consensus will be required.
Self-fertilization in S. cerevisiae and relatives is achieved through the tight regulation of the HO gene, which encodes a homing endonuclease co-opted from an intein selfish element [18]. The Ho protein cuts the active mating cassette, allowing for homology repair by one of two silent mating cassettes, thereby switching the mating type and enabling clonal mother-daughter mating. Interestingly, Kluyveromyces lactis HO has been replaced by MATα3, another selfish element co-opted from a DNA transposon [19,20]. A simpler two-locus system that switches mating types by inversion has evolved in the clade containing Ogataea (Hansenula) polymorpha and Komagataella (Pichia) pastoris [21]. Here inversion simply switches which of the two mating loci is present in active chromatin, and thus determines which is expressed. These genetic systems have profound effects on maintaining local genetic variation [22] and generating genome instability [23] at the mating-type locus.
The genetic regulatory networks that control the batteries of genes necessary for each mating type have been investigated extensively in several species, establishing a paradigm for both the logic and molecular mechanisms by which such networks evolve. Several genetic changes can facilitate the evolutionary rewiring of regulatory networks, including the duplication of regulators and partitioning of target genes [24], intercalation of new regulators [25], and alteration of binding site specificity [26]. Most such changes are thought to occur through selectively neutral mechanisms, often resulting in the same network output. These studies have established rewiring as a common feature in regulatory network evolution and have shown how neutral changes can profoundly shape evolutionary outcomes.
Metabolic diversity, ecology, and biotechnology
Thanks to their diverse metabolisms, yeasts exhibit a remarkable range of ecologies, including methanol consumption in O. polymorpha and K. pastoris; xylose fermentation in Scheffersomyces (P.) stipitis and Spathaspora passalidarum; lipid production in Yarrowia lipolytica and Lipomyces starkeyi; human pathogenesis in C. albicans and C. (Nakaseomyces) glabrata; and cotton pathogenesis in Eremothecium (Ashbya) gossypii [27] (Figure 1). Many of these metabolic capabilities are exploited in various biotechnological, food, and beverage industries. For example, S. cerevisiae is the workhorse of the multibillion-dollar brewing, wine-making, baking, and biofuel industries, but other commonly used species include Kazachstania exigua (sourdough), Cyberlindnera jadinii (syn. C. utilis, food additives), K. pastoris (heterologous protein production), and E. gossypii (riboflavin). Nonetheless, these well-known yeasts represent a scant slice of yeast metabolisms and ecologies, and most species remain unharnessed by industry and only modestly explored by science.
Yeast ecological niches require more study, especially in natural settings, but what limited information exists suggests that yeast niches are partitioned by many parameters, including temperature, pH, radiation, insect and plant hosts, and metabolism [27–29]. For example, Saccharomyces and relatives have evolved an extreme preference for fermenting glucose into ethanol, even in the presence of oxygen, a process known as Crabtree-Warburg Effect or aerobic fermentation. This “make-accumulate-consume” strategy provides a powerful ecological advantage by exploiting the rich reserves of simple sugars in sap, fruit, and other sources [30–33]. Other genera, such as Scheffersomyces and Spathaspora, have adapted to living in the guts of wood-consuming beetles and are capable of fermenting xylose, the second most abundant monosaccharide in woody plant material, which is of critical importance to the lignocellulosic biofuel industry [11,34,35]. Some traits are unique, such as a requirement for carbon dioxide in Cyniclomyces [36], while others, such as temperature preferences, evolve quickly [27–29]. A handful of lineages have evolved into commensals and/or pathogens of mammals and birds, exhibiting increased thermophily, the loss of key metabolic functions, increased biofilm-forming ability, and the acquisition of new traits to evade the host immune system [2,7,8,37].
The genetic outlines underlying some of these remarkable innovations are beginning to be understood. Some differences evolve rapidly and can be explained by one or two genes. For example, there is widespread variation in the catabolism of disaccharides, perhaps because extracellular cleavage often sets up a “prisoner’s dilemma” where competitors can consume the monosaccharide intermediates [29]. Other traits, such as galactose catabolism, seem to be lost frequently [10,38] (and potentially regained [39]), correlating with the loss of several genes. Aerobic fermentation is thought to involve many genetic changes in glycolytic and mitochondrial pathways and has evolved only a handful of times [30–33]. Strikingly, many of these pathways underwent parallel evolutionary changes during the acquisition of aerobic fermentation by the lineage leading to S. cerevisiae and the highly divergent lineage leading to Brettanomyces (Dekkera) bruxellensis [30,32].
Much remains to be understood about the forces driving most yeast radiations (Figure 2). For example, dozens of species evolved from clades associated with fruit-rot and tree-flux habitats to exploit the diverse chemistries associated with different species of cacti, which, in turn, have been exploited by several different cactophilic Drosophila and sap beetle species that feed on these yeasts [27,28]. Most species of the Starmerella clade are associated with Hymenoptera species, while yeasts of the genus Ogataea frequently inhabit leaf litter where methanol consumption may be beneficial [21,27,40]. Little is known about the genetic underpinnings of these ecological innovations, except that they must rely, at least partly, on metabolism. Metabolic traits, which can be easily studied in the lab and whose genetic basis is often well understood from model organisms, are therefore uniquely suited to provide the thin end of a wedge to open an understanding of yeast diversification.
Hybridization, introgression, and horizontal gene transfer
One of the most important paradigm shifts catalyzed by broader yeast genome sequencing is a realization of the prevalence of gene sharing through hybridization, introgression, and horizontal gene transfer (HGT), especially among domesticated strains. Perhaps the most iconic examples are the lager-brewing yeasts, which were recently shown to be allopolyploids of S. cerevisiae and the recently discovered species S. eubayanus [41,42]. Alloploids of S. cerevisiae x S. kudriavzevii are also used in brewing, wine fermentation, and cider fermentation [43]. Conversely, strains of S. uvarum used in cider and champagne fermentation have acquired genes from several Saccharomyces species, but most wild strains lack these introgressions [44].
Fewer alloploids are known outside of the genus Saccharomyces, and all are from industrial or clinical settings. In the genus Zygosaccharomyces, a miso production strain [45] and a sparkling wine contaminant [46] are allopolyploids. Echoing the recent status of S. eubayanus, genome sequencing showed Millerozyma (P.) sorbitophila, a sorbitol contaminant, to be an interspecies hybrid of Millerozyma (P.) farinosa and a yet-to-be-identified congeneric species [47]. Similarly, genome sequencing of a wine spoilage strain purported to be B. bruxellensis revealed it to be allotriploid [48]. Several clinical isolates from the C. parapsilosis species complex are also interspecies hybrids [49].
In addition to hybridization, many strains of S. cerevisiae used in industry have picked up genes through introgression from various Saccharomyces species [44,50,51], as well as from more distantly related yeasts through HGT [52–54]. HGT is not restricted to strains grown in artificial environments but also occurs at a low, but significant, rate across the yeast phylogeny, influencing the content of both the nuclear [55] and mitochondrial genomes [56]. For example, a recent examination of fungal metabolic pathways inferred that 1.8% of yeast metabolic genes have undergone HGT at some point in their history [57]. HGT has influenced individual genes, sometimes repeatedly [33,53,58]; entire metabolic pathways [59]; and larger genomic fragments [52], although the latter has only been observed in industrial strains [52,55]. Although it is thought that conjugation, transformation, and viral transduction are involved in filamentous fungi [60], less is known about the mechanisms that facilitate yeast HGT. Interestingly, a recent study showed that experimental rates of bacteria-to-yeast HGT can be strongly influenced by gene presence/absence polymorphisms in non-essential yeast genes [61].
Resequencing dynamic genomes
Yeast genomics has greatly enhanced our understanding of the evolutionary dynamics of natural populations, among domesticated strains, during infections, and during laboratory experiments. Aside from S. cerevisiae [51], population genomic studies have characterized the metabolic, genetic, and biogeographic diversity of S. paradoxus [62], S. kudriavzevii [38], and S. uvarum [44], making this genus one of the few where population genomic datasets exist for the majority of known species [63]. Large-scale population genomic studies in other yeasts remain rare, but a recent investigation of the stability of the unusual GC content of a chromosome-arm in Lachancea kluyveri is an undeniable harbinger of the future [64]. Remarkably, estimates of the frequency of outcrossing (~10−5) in L. kluyveri are similar to Saccharomyces [22], suggesting that key lifestyle parameters may be conserved across vast timespans, different ecologies, and different mating-type control systems.
Genome resequencing has allowed experimental evolution studies in S. cerevisiae to rapidly move from identifying specific adaptive mutations [15,65,66] to examining pools of variants undergoing clonal interference [67,68] to massively parallelized tracking of individually tagged lineages [69]. Although the most advanced genetic tricks are not available for other yeasts, some of these approaches have even been extended to study the evolution of clinical isolates of C. albicans during the course of infection [70,71]. Collectively, these studies paint the portrait of a dynamic genome that can be pushed easily toward evolving a variety of traits. Indeed, a provocative corollary is that the trillions of scattered cells belonging to each yeast species are being exposed to potentially quite different selective pressures, ultimately engendering different genomic outcomes that must be reconciled.
Which microevolutionary processes explain macroevolutionary patterns?
Juxtaposing data from laboratory evolution and population genomic studies to the data from comparisons of distantly related genomes raises an interesting paradox: rapid, radical, and sometimes irreversible changes regularly occur across microevolutionary timescales, and yet, relative stability persists over macroevolutionary ones. Some common types of mutations, such as aneuploidy [66], are unlikely to be fixed in a species because these low frequency polymorphisms are easily reversed. Others, such as interspecies hybridization and the loss of mating pathways, can expose lineages to Muller’s Ratchet and eventual extinction. Many of the most readily acquired mutations are further predicted to reduce or eliminate gene activity, especially in regulatory genes [15,65,66,71].
Certainly, many lineages have lost specific genes, networks, and traits, but even when they arise as conditionally advantageous mutations, several forces likely act to limit the impact of the most radical changes over geological timescales. First, species are seldom exposed to static environments that allow conditionally useful genes to be tossed aside. Rather, they must be able to thrive and complete their life cycles in fluctuating environments. Second, individuals exchange genes with other members of their species, which may exist across a range of environments, allowing genes to be reintroduced if a loss was only transiently beneficial. Third, although the mutations may be rarer than nonsense or frameshift mutations, more specific regulatory mutations are likely to arise eventually [72]. Fourth, horizontal gene transfer (HGT) may allow genes to be reacquired from other organisms [55,59]. Finally, lineages that have lost key functions may be less likely to undergo cladogenesis or be more susceptible to extinction.
Given the currently limited sampling of yeast genomes, one might justly ask whether we are missing a trend of widespread, but undetected, gene loss and gene sharing that will be revealed as all the branches of the phylogeny are filled in. Many additional congeneric hybrids at the tips of the phylogeny will undoubtedly be discovered. Nonetheless, as with most gene loss events, hybridization events are generally neutral or short-lived adaptations to extreme conditions. Allopolyploidization results in whole genome duplications (WGDs) [73], which are relatively easy to detect, thanks to pioneering work that inferred the WGD in the lineage leading to S. cerevisiae by comparing this genome to itself [74]. Although that solitary WGD event had a tremendous and lasting impact on duplicate gene function and genome structure in over 50 species of Saccharomycetaceae, including S. cerevisiae [75], the frequency with which yeast WGD descendants persist is clearly much lower than in angiosperms, where most lineages have undergone multiple rounds of WGD [76]. Finally, HGT is responsible for a non-trivial number of yeast adaptations, but the rate remains much lower than in bacteria [57].
Prospects for the future
As with all organisms, yeast genome sequences comprehensively describe their genetic makeups, but historical and genetic processes that have sculpted their evolution are best understood through a comparative lens. Remarkably, several ambitious projects promise to enhance dramatically the genome sampling of fungal biodiversity at both the population and species levels, including the 1KFG Project surveying the entire spectrum of fungal diversity (http://1000.fungalgenomes.org), the 1002 Yeast Genomes Project focusing on S. cerevisiae (http://1002genomes.u-strasbg.fr), the Dikaryome Consortium (http://dikaryome.org), the iGénolevures Consortium (http://gryc.inra.fr), and the Y1000+ Project focusing on the subphylum Saccharomycotina (http://y1000plus.org).
At least for the yeasts, these projects stand a chance of saturating two key dimensions of known biodiversity: genetic depth and taxonomic breadth. Just as a genome sequence definitively describes what genes are present in an organism, only a complete clade of genomes can fully chronicle their evolution and enable the study of genetic and functional diversification across taxa, niches, and time. With these projects underway and with perhaps the most advanced armamentarium for functional dissection at hand, the genetic features that cause some species to make beer, others to inhabit cacti, and still others to cause lethal blood infections are finally coming into focus. The yeasts are rising to the challenge to create an unparalleled model of eukaryotic genome evolution.
Acknowledgments
We thank the many colleagues who alerted us to relevant literature and apologize to those whose work was excluded due to space limitations or oversight. Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture. USDA is an equal opportunity provider and employer. Research in the Hittinger Lab is supported in part by the National Science Foundation (Grant Nos. DEB-1442148 to CTH and CPK, DEB-1253634 to CTH); the DOE Great Lakes Bioenergy Research Center (DOE Office of Science BER DE-FC02-07ER64494); the USDA National Institute of Food and Agriculture (Hatch project 1003258); the Alexander von Humboldt Foundation (CTH is an Alfred Toepfer Faculty Fellow); and the Pew Charitable Trusts (CTH is a Pew Scholar in the Biomedical Sciences). Research in the Rokas Lab is supported by the National Science Foundation (DEB-1442113); the National Institutes of Health (NIAID, AI105619); and the March of Dimes. Research in the Lachance Lab is supported by the Natural Science and Engineering Research Council of Canada. Research in the Libkind Lab is supported by CONICET, Universidad Nacional del Comahue (B171), and FONCyT (PICT 2014-2542). Research in the Rosa Lab is supported by the Conselho Nacional de Desenvolvimento Cientifico e Tecnológico (CNPq)-Brazil.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Literature Cited
- 1.Goffeau A, Barrell BG, Bussey H, Davis RW, Dujon B, Feldmann H, Galibert F, Hoheisel JD, Jacq C, Johnston M, et al. Life with 6000 genes. Science. 1996;274:546,563–567. doi: 10.1126/science.274.5287.546. [DOI] [PubMed] [Google Scholar]
- 2.Jones T, Federspiel NA, Chibana H, Dungan J, Kalman S, Magee BB, Newport G, Thorstenson YR, Agabian N, Magee PT, et al. The diploid genome sequence of Candida albicans. Proc Natl Acad Sci U S A. 2004;101:7329–7334. doi: 10.1073/pnas.0401648101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Cliften P, Sudarsanam P, Desikan A, Fulton L, Fulton B, Majors J, Waterston R, Cohen BA, Johnston M. Finding functional features in Saccharomyces genomes by phylogenetic footprinting. Science. 2003;301:71–76. doi: 10.1126/science.1084337. [DOI] [PubMed] [Google Scholar]
- 4.Kellis M, Patterson N, Endrizzi M, Birren B, Lander ES. Sequencing and comparison of yeast species to identify genes and regulatory elements. Nature. 2003;423:241–254. doi: 10.1038/nature01644. [DOI] [PubMed] [Google Scholar]
- 5.Kellis M, Birren BW, Lander ES. Proof and evolutionary analysis of ancient genome duplication in the yeast Saccharomyces cerevisiae. Nature. 2004;428:617–624. doi: 10.1038/nature02424. [DOI] [PubMed] [Google Scholar]
- 6.Dietrich FS, Voegeli S, Brachat S, Lerch A, Gates K, Steiner S, Mohr C, Pohlmann R, Luedi P, Choi S, et al. The Ashbya gossypii genome as a tool for mapping the ancient Saccharomyces cerevisiae genome. Science. 2004;304:304–307. doi: 10.1126/science.1095781. [DOI] [PubMed] [Google Scholar]
- 7.Butler G, Rasmussen MD, Lin MF, Santos MA, Sakthikumar S, Munro CA, Rheinbay E, Grabherr M, Forche A, Reedy JL, et al. Evolution of pathogenicity and sexual reproduction in eight Candida genomes. Nature. 2009;459:657–662. doi: 10.1038/nature08064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Dujon B, Sherman D, Fischer G, Durrens P, Casaregola S, Lafontaine I, De Montigny J, Marck C, Neuveglise C, Talla E, et al. Genome evolution in yeasts. Nature. 2004;430:35–44. doi: 10.1038/nature02579. [DOI] [PubMed] [Google Scholar]
- 9.Giaever G, Chu AM, Ni L, Connelly C, Riles L, Veronneau S, Dow S, Lucau-Danila A, Anderson K, Andre B, et al. Functional profiling of the Saccharomyces cerevisiae genome. Nature. 2002;418:387–391. doi: 10.1038/nature00935. [DOI] [PubMed] [Google Scholar]
- 10.Hittinger CT, Rokas A, Carroll SB. Parallel inactivation of multiple GAL pathway genes and ecological diversification in yeasts. Proc Natl Acad Sci U S A. 2004;101:14144–14149. doi: 10.1073/pnas.0404319101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11**.Wohlbach DJ, Kuo A, Sato TK, Potts KM, Salamov AA, Labutti KM, Sun H, Clum A, Pangilinan JL, Lindquist EA, et al. Comparative genomics of xylose-fermenting fungi for enhanced biofuel production. Proc Natl Acad Sci U S A. 2011;108:13212–13217. doi: 10.1073/pnas.1103039108. The first comparative genomics study of xylose-fermenting yeasts revealed several common genetic pathways and physiological strategies that have evolved to enable xylose fermentation. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12*.Salichos L, Rokas A. Inferring ancient divergences requires genes with strong phylogenetic signal. Nature. 2013;497:327–31. doi: 10.1038/nature12130. The latest in a series of phylogenomic methodological advances catalyzed by yeast genome sequences. This paper introduced the internode certainty and tree certainty indices, which formally measure phylogenetic conflict. [DOI] [PubMed] [Google Scholar]
- 13.MacIsaac KD, Wang T, Gordon DB, Gifford DK, Stormo GD, Fraenkel E. An improved map of conserved regulatory sites for Saccharomyces cerevisiae. BMC Bioinformatics. 2006;7:113. doi: 10.1186/1471-2105-7-113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Dujon B. Yeast evolutionary genomics. Nat Rev Genet. 2010;11:512–524. doi: 10.1038/nrg2811. [DOI] [PubMed] [Google Scholar]
- 15.Lang GI, Murray AW, Botstein D. The cost of gene expression underlies a fitness trade-off in yeast. Proc Natl Acad Sci U S A. 2009;106:5755–5760. doi: 10.1073/pnas.0901620106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Goddard MR, Godfray HCJ, Burt A. Sex increases the efficacy of natural selection in experimental yeast populations. Nature. 2005;434:636–40. doi: 10.1038/nature03405. [DOI] [PubMed] [Google Scholar]
- 17.Gray JC, Goddard MR. Sex enhances adaptation by unlinking beneficial from detrimental mutations in experimental yeast populations. BMC Evol Biol. 2012;12:43. doi: 10.1186/1471-2148-12-43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Butler G, Kenny C, Fagan A, Kurischko C, Gaillardin C, Wolfe KH. Evolution of the MAT locus and its Ho endonuclease in yeast species. Proc Natl Acad Sci U S A. 2004;101:1632–1637. doi: 10.1073/pnas.0304170101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Fabre E, Muller H, Therizols P, Lafontaine I, Dujon B, Fairhead C. Comparative genomics in hemiascomycete yeasts: evolution of sex, silencing, and subtelomeres. Mol Biol Evol. 2005;22:856–73. doi: 10.1093/molbev/msi070. [DOI] [PubMed] [Google Scholar]
- 20.Barsoum E, Martinez P, Aström SU. Alpha3, a transposable element that promotes host sexual reproduction. Genes Dev. 2010;24:33–44. doi: 10.1101/gad.557310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21**.Hanson SJ, Byrne KP, Wolfe KH. Mating-type switching by chromosomal inversion in methylotrophic yeasts suggests an origin for the three-locus Saccharomyces cerevisiae system. Proc Natl Acad Sci U S A. 2014;111:E4851–8. doi: 10.1073/pnas.1416014111. Discovered and investigated an inversion-based mechanism for mating-type switching that may represent the ancestral state for many yeasts, likely indicating that the use of homing endonucleases co-opted from selfish elements, as in S. cerevisiae, is evolutionarily derived. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Tsai IJ, Bensasson D, Burt A, Koufopanou V. Population genomics of the wild yeast Saccharomyces paradoxus: Quantifying the life cycle. Proc Natl Acad Sci U S A. 2008;105:4957–4962. doi: 10.1073/pnas.0707314105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23*.Gordon JL, Armisen D, Proux-Wera E, OhEigeartaigh SS, Byrne KP, Wolfe KH. Evolutionary erosion of yeast sex chromosomes by mating-type switching accidents. Proc Natl Acad Sci U S A. 2011;108:20024–20029. doi: 10.1073/pnas.1112808108. Sequenced several phylogenetically well-distributed Saccharomycetaceae to show that genes adjacent to the mating-type locus are prone to loss or translocation in species with homing endonuclease-based switching mechanisms. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Pérez JC, Fordyce PM, Lohse MB, Hanson-Smith V, DeRisi JL, Johnson AD. How duplicated transcription regulators can diversify to govern the expression of nonoverlapping sets of genes. Genes Dev. 2014;28:1272–7. doi: 10.1101/gad.242271.114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Booth LN, Tuch BB, Johnson AD. Intercalation of a new tier of transcription regulation into an ancient circuit. Nature. 2010;468:959–963. doi: 10.1038/nature09560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26*.Baker CR, Tuch BB, Johnson AD. Extensive DNA-binding specificity divergence of a conserved transcription regulator. Proc Natl Acad Sci U S A. 2011;108:7493–8. doi: 10.1073/pnas.1019177108. An excellent example of hypothesis-driven genome sequencing where two species were sequenced to study the evolution of the binding site motifs of a mating-type transcription factor. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kurtzman CP, Fell JW, Boekhout T. The Yeasts: A Taxonomic Study. Elsevier Science; 2011. [Google Scholar]
- 28.Lachance M. The Biodiversity, Ecology, and Biogeography of Ascomycetous Yeasts. In: Wiley-Blackwell Martin F., editor. The Ecological Genomics of Fungi. 2014. pp. 355–370. [Google Scholar]
- 29.Greig D, Travisano M. The Prisoner’s Dilemma and polymorphism in yeast SUC genes. Proc Biol Sci. 2004;271(Suppl):S25–6. doi: 10.1098/rsbl.2003.0083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30**.Thompson DA, Roy S, Chan M, Styczynsky MP, Pfiffner J, French C, Socha A, Thielke A, Napolitano S, Muller P, et al. Evolutionary principles of modular gene regulation in yeasts. Elife. 2013;2:e00603. doi: 10.7554/eLife.00603. Landmark study that explored the evolution of aerobic fermentation across several species of yeasts by examining the genetic regulatory networks governing diauxic shift, the transition from the consumption of fermentable to respiratory carbon sources. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31*.Hagman A, Säll T, Compagno C, Piskur J. Yeast “make-accumulate-consume” life strategy evolved as a multi-step process that predates the whole genome duplication. PLoS One. 2013;8:e68734. doi: 10.1371/journal.pone.0068734. This physiological study of several yeasts suggested that the timing and trajectory of the evolution of aerobic fermentation by the lineage leading to S. cerevisiae was much more complex than has been assumed. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32**.Rozpedowska E, Hellborg L, Ishchuk OP, Orhan F, Galafassi S, Merico A, Woolfit M, Compagno C, Piskur J. Parallel evolution of the make-accumulate-consume strategy in Saccharomyces and Dekkera yeasts. Nat Commun. 2011;2:302. doi: 10.1038/ncomms1305. Demonstrated that B. bruxellensis independently evolved aerobic fermentation in part through the same molecular mechanisms as in S. cerevisiae, including the massive loss of a cis-acting element regulating genes involved in respiration. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Gojkovic Z, Knecht W, Zameitat E, Warneboldt J, Coutelis JB, Pynyaha Y, Neuveglise C, Moller K, Loffler M, Piskur J. Horizontal gene transfer promoted evolution of the ability to propagate under anaerobic conditions in yeasts. Mol Genet Genomics. 2004;271:387–393. doi: 10.1007/s00438-004-0995-7. [DOI] [PubMed] [Google Scholar]
- 34.Lobo FP, Gonçalves DL, Alves SL, Gerber AL, de Vasconcelos ATR, Basso LC, Franco GR, Soares MA, Cadete RM, Rosa CA, et al. Draft Genome Sequence of the D-Xylose-Fermenting Yeast Spathaspora arborariae UFMG-HM19.1AT. Genome Announc. 2014;2:e01163–13. doi: 10.1128/genomeA.01163-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Jeffries TW, Grigoriev IV, Grimwood J, Laplaza JM, Aerts A, Salamov A, Schmutz J, Lindquist E, Dehal P, Shapiro H, et al. Genome sequence of the lignocellulose-bioconverting and xylose-fermenting yeast Pichia stipitis. Nat Biotechnol. 2007;25:319–326. doi: 10.1038/nbt1290. [DOI] [PubMed] [Google Scholar]
- 36.Kurtzman CP, Robnett CJ. Relationships among genera of the Saccharomycotina (Ascomycota) from multigene phylogenetic analysis of type species. FEMS Yeast Res. 2013;13:23–33. doi: 10.1111/1567-1364.12006. [DOI] [PubMed] [Google Scholar]
- 37.Gabaldón T, Martin T, Marcet-Houben M, Durrens P, Bolotin-Fukuhara M, Lespinet O, Arnaise S, Boisnard S, Aguileta G, Atanasova R, et al. Comparative genomics of emerging pathogens in the Candida glabrata clade. BMC Genomics. 2013;14:623. doi: 10.1186/1471-2164-14-623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Hittinger CT, Gonçalves P, Sampaio JP, Dover J, Johnston M, Rokas A. Remarkably ancient balanced polymorphisms in a multi-locus gene network. Nature. 2010;464:54–8. doi: 10.1038/nature08791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Slot JC, Rokas A. Multiple GAL pathway gene clusters evolved independently and by different mechanisms in fungi. Proc Natl Acad Sci U S A. 2010;107:10136–10141. doi: 10.1073/pnas.0914418107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Ramezani-Rad M, Hollenberg CP, Lauber J, Wedler H, Griess E, Wagner C, Albermann K, Hani J, Piontek M, Dahlems U, et al. The Hansenula polymorpha (strain CBS4732) genome sequencing and analysis. FEMS Yeast Res. 2003;4:207–15. doi: 10.1016/S1567-1356(03)00125-9. [DOI] [PubMed] [Google Scholar]
- 41.Libkind D, Hittinger CT, Valerio E, Goncalves C, Dover J, Johnston M, Goncalves P, Sampaio JP. Microbe domestication and the identification of the wild genetic stock of lager-brewing yeast. Proc Natl Acad Sci U S A. 2011;108:14539–14544. doi: 10.1073/pnas.1105430108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Nakao Y, Kanamori T, Itoh T, Kodama Y, Rainieri S, Nakamura N, Shimonaga T, Hattori M, Ashikari T. Genome sequence of the lager brewing yeast, an interspecies hybrid. DNA Res. 2009;16:115–129. doi: 10.1093/dnares/dsp003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Peris D, Lopes CA, Belloch C, Querol A, Barrio E. Comparative genomics among Saccharomyces cerevisiae x Saccharomyces kudriavzevii natural hybrid strains isolated from wine and beer reveals different origins. BMC Genomics. 2012;13:407. doi: 10.1186/1471-2164-13-407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44*.Almeida P, Goncalves C, Teixeira S, Libkind D, Bontrager M, Albertin W, Durrens P, Sherman D, Marullo P, Masneuf-pomare I, et al. A Gondwanan imprint on global diversity and domestication of wine and cider yeast Saccharomyces uvarum. Nat Commun. 2014;5:4044. doi: 10.1038/ncomms5044. Genome sequence data from over 50 strains showed that many cider and champagne yeasts are domesticated strains of S. uvarum, which have often acquired genes from other species of Saccharomyces. Wild strains of S. uvarum generally lacked introgressions. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Gordon JL, Wolfe KH. Recent allopolyploid origin of Zygosaccharomyces rouxii strain ATCC 42981. Yeast. 2008;25:449–56. doi: 10.1002/yea.1598. [DOI] [PubMed] [Google Scholar]
- 46.Mira NP, Münsterkötter M, Dias-Valada F, Santos J, Palma M, Roque FC, Guerreiro JF, Rodrigues F, Sousa MJ, Leão C, et al. The genome sequence of the highly acetic acid-tolerant Zygosaccharomyces bailii-derived interspecies hybrid strain ISA1307, isolated from a sparkling wine plant. DNA Res. 2014;21:299–313. doi: 10.1093/dnares/dst058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Louis VL, Despons L, Friedrich A, Martin T, Durrens P, Casarégola S, Neuvéglise C, Fairhead C, Marck C, Cruz JA, et al. Pichia sorbitophila, an Interspecies Yeast Hybrid, Reveals Early Steps of Genome Resolution After Polyploidization. G3 (Bethesda) 2012;2:299–311. doi: 10.1534/g3.111.000745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Curtin CD, Borneman AR, Chambers PJ, Pretorius IS. De-Novo Assembly and Analysis of the Heterozygous Triploid Genome of the Wine Spoilage Yeast Dekkera bruxellensis AWRI1499. PLoS One. 2012;7:e33840. doi: 10.1371/journal.pone.0033840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49*.Pryszcz LP, Németh T, Gácser A, Gabaldón T. Genome comparison of Candida orthopsilosis clinical strains reveals the existence of hybrids between two distinct subspecies. Genome Biol Evol. 2014;6:1069–78. doi: 10.1093/gbe/evu082. Genome sequence data showed that several clinical isolates from the C. parapsilosis species complex are interspecies hybrids, extending the possible link between humans and interspecies hybridization to a clinical setting. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Dunn B, Richter C, Kvitek DJ, Pugh T, Sherlock G. Analysis of the Saccharomyces cerevisiae pan-genome reveals a pool of copy number variants distributed in diverse yeast strains from differing industrial environments. Genome Res. 2012;22:908–924. doi: 10.1101/gr.130310.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51*.Strope PK, Skelly DA, Kozmin SG, Mahadevan G, Stone EA, Magwene PM, Dietrich FS, McCusker JH. The 100-genomes strains, an S. cerevisiae resource that illuminates its natural phenotypic and genotypic variation and emergence as an opportunistic pathogen. Genome Res. 2015;25:762–74. doi: 10.1101/gr.185538.114. Landmark study of 100 S. cerevisiae genome sequences, which includes considerable evidence of introgression and horizontal gene transfer. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Novo M, Bigey F, Beyne E, Galeote V, Gavory F, Mallet S, Cambon B, Legras JL, Wincker P, Casaregola S, et al. Eukaryote-to-eukaryote gene transfer events revealed by the genome sequence of the wine yeast Saccharomyces cerevisiae EC1118. Proc Natl Acad Sci U S A. 2009;106:16333–16338. doi: 10.1073/pnas.0904673106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53*.Coelho MA, Gonçalves C, Sampaio JP, Gonçalves P. Extensive intra-kingdom horizontal gene transfer converging on a fungal fructose transporter gene. PLoS Genet. 2013;9:e1003587. doi: 10.1371/journal.pgen.1003587. Demonstrated the repeated HGT of the FSY1 fructose symporter into multiple divergent clades of yeasts, including wine-associated lineages of S. cerevisiae. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54*.Marsit S, Mena A, Bigey F, Sauvage F-X, Couloux A, Guy J, Legras J-L, Barrio E, Dequin S, Galeote V. Evolutionary advantage conferred by an eukaryote-to-eukaryote gene transfer event in wine yeasts. Mol Biol Evol. 2015;32:1695–707. doi: 10.1093/molbev/msv057. Determined that the FOT oligopeptide transporters conferred a fitness advantage in grape musts. Along with FSY1 [53], two of three major HGT events from other yeasts into wine-associated lineages of S. cerevisiae have now been shown to confer conditional fitness advantages. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Fitzpatrick DA. Horizontal gene transfer in fungi. FEMS Microbiol Lett. 2012;329:1–8. doi: 10.1111/j.1574-6968.2011.02465.x. [DOI] [PubMed] [Google Scholar]
- 56.Wu B, Buljic A, Hao W. Extensive Horizontal Transfer and Homologous Recombination Generate Highly Chimeric Mitochondrial Genomes in Yeast. Mol Biol Evol. 2015 doi: 10.1093/molbev/msv127. [DOI] [PubMed] [Google Scholar]
- 57**.Wisecaver JH, Slot JC, Rokas A. The evolution of fungal metabolic pathways. PLoS Genet. 2014;10:e1004816. doi: 10.1371/journal.pgen.1004816. Quantified the rate of horizontal gene transfer across various fungal lineages and showed it to be lower in the Saccharomycotina than in several clades of Pezizomycotina, the subphylum to which most filamentous ascomycetes belong. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Bruto M, Prigent-Combaret C, Luis P, Moënne-Loccoz Y, Muller D. Frequent, independent transfers of a catabolic gene from bacteria to contrasted filamentous eukaryotes. Proc Biol Sci. 2014;281:20140848. doi: 10.1098/rspb.2014.0848. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Wisecaver JH, Rokas A. Fungal metabolic gene clusters-caravans traveling across genomes and environments. Front Microbiol. 2015;6:161. doi: 10.3389/fmicb.2015.00161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Soanes D, Richards TA. Horizontal gene transfer in eukaryotic plant pathogens. Annu Rev Phytopathol. 2014;52:583–614. doi: 10.1146/annurev-phyto-102313-050127. [DOI] [PubMed] [Google Scholar]
- 61.Moriguchi K, Yamamoto S, Tanaka K, Kurata N, Suzuki K. Trans-kingdom horizontal DNA transfer from bacteria to yeast is highly plastic due to natural polymorphisms in auxiliary nonessential recipient genes. PLoS One. 2013;8:e74590. doi: 10.1371/journal.pone.0074590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62*.Liti G, Carter DM, Moses AM, Warringer J, Parts L, James SA, Davey RP, Roberts IN, Burt A, Koufopanou V, et al. Population genomics of domestic and wild yeasts. Nature. 2009;458:337–341. doi: 10.1038/nature07743. The first population genomics study in yeasts, which included a comparison of S. cerevisiae with its many domesticated lineages to its sister species, S. paradoxus, whose natural history and genetic makeup have been much less impacted by humans. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Lawrie DS, Petrov DA. Comparative population genomics: power and principles for the inference of functionality. Trends Genet. 2014;30:133–9. doi: 10.1016/j.tig.2014.02.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64**.Friedrich A, Jung P, Reisser C, Fischer G, Schacherer J. Population genomics reveals chromosome-scale heterogeneous evolution in a protoploid yeast. Mol Biol Evol. 2015;32:184–92. doi: 10.1093/molbev/msu295. The first yeast population genomics study outside of the Saccharomyces suggested that the frequency of outcrossing may be conserved across vast timescales. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.New AM, Cerulus B, Govers SK, Perez-Samper G, Zhu B, Boogmans S, Xavier JB, Verstrepen KJ. Different Levels of Catabolite Repression Optimize Growth in Stable and Variable Environments. PLoS Biol. 2014;12:e1001764. doi: 10.1371/journal.pbio.1001764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Selmecki AM, Maruvka YE, Richmond PA, Guillet M, Shoresh N, Sorenson AL, De S, Kishony R, Michor F, Dowell R, et al. Polyploidy can drive rapid adaptation in yeast. Nature. 2015;519:349–52. doi: 10.1038/nature14187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67**.Kvitek DJ, Sherlock G. Whole genome, whole population sequencing reveals that loss of signaling networks is the major adaptive strategy in a constant environment. PLoS Genet. 2013;9:e1003972. doi: 10.1371/journal.pgen.1003972. This experimental evolution study showed that loss-of-function alleles, especially in regulatory pathways, frequently evolve in constant environmental conditions. Conditional fitness experiments verfied the tradeoffs under selection. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Lang GI, Rice DP, Hickman MJ, Sodergren E, Weinstock GM, Botstein D, Desai MM. Pervasive genetic hitchhiking and clonal interference in forty evolving yeast populations. Nature. 2013;500:571–4. doi: 10.1038/nature12344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69*.Levy SF, Blundell JR, Venkataram S, Petrov DA, Fisher DS, Sherlock G. Quantitative evolutionary dynamics using high-resolution lineage tracking. Nature. 2015;519:181–6. doi: 10.1038/nature14279. Developed a highly parallelized system that tracks the evolution of cellular lineages through next-generation sequencing of DNA barcodes. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Ford CB, Funt JM, Abbey D, Issi L, Guiducci C, Martinez DA, Delorey T, Li BY, White TC, Cuomo C, et al. The evolution of drug resistance in clinical isolates of Candida albicans. Elife. 2015;4:e00662. doi: 10.7554/eLife.00662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Hirakawa MP, Martinez DA, Sakthikumar S, Anderson MZ, Berlin A, Gujja S, Zeng Q, Zisson E, Wang JM, Greenberg JM, et al. Genetic and phenotypic intra-species variation in Candida albicans. Genome Res. 2015;25:413–25. doi: 10.1101/gr.174623.114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Stern DL, Orgogozo V. The loci of evolution: how predictable is genetic evolution? Evolution (N Y) 2008;62:2155–2177. doi: 10.1111/j.1558-5646.2008.00450.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73**.Marcet-Houben M, Gabaldón T. Beyond the whole genome duplication: phylogenetic evidence for an ancient inter-species hybridization in the baker’s yeast lineage. PLoS Biol. 2015;13:e1002220. doi: 10.1371/journal.pbio.1002220. Provided the first concrete phylogenomic evidence that the WGD in the lineage leading to S. cerevisiae was due to an allopolyploidization event, rather than an autopolyploidization event. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Wolfe KH, Shields DC. Molecular evidence for an ancient duplication of the entire yeast genome. Nature. 1997;387:708–713. doi: 10.1038/42711. [DOI] [PubMed] [Google Scholar]
- 75.Conant GC, Wolfe KH. Turning a hobby into a job: how duplicated genes find new functions. Nat Rev Genet. 2008;9:938–50. doi: 10.1038/nrg2482. [DOI] [PubMed] [Google Scholar]
- 76.Soltis DE, Bell CD, Kim S, Soltis PS. Origin and early evolution of angiosperms. Ann N Y Acad Sci. 2008;1133:3–25. doi: 10.1196/annals.1438.005. [DOI] [PubMed] [Google Scholar]
- 77.Scannell DR, Zill OA, Rokas A, Payen C, Dunham MJ, Eisen MB, Rine J, Johnston M, Hittinger CT. The awesome power of yeast evolutionary genetics: new genome sequences and strain resources for the Saccharomyces sensu stricto genus. G3 Genes Genomes Genet. 2011;1:11–25. doi: 10.1534/g3.111.000273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Liti G, Nguyen Ba AN, Blythe M, Müller CA, Bergström A, Cubillos FA, Dafhnis-Calas F, Khoshraftar S, Malla S, Mehta N, et al. High quality de novo sequencing and assembly of the Saccharomyces arboricolus genome. BMC Genomics. 2013;14:69. doi: 10.1186/1471-2164-14-69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Baker E, Wang B, Bellora N, Peris D, Hulfachor AB, Koshalek JA, Adams M, Libkind D, Hittinger CT. The genome sequence of Saccharomyces eubayanus and the domestication of lager-brewing yeasts. Mol Biol Evol. 2015;32:2818–31. doi: 10.1093/molbev/msv168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Scannell DR, Frank AC, Conant GC, Byrne KP, Woolfit M, Wolfe KH. Independent sorting-out of thousands of duplicated gene pairs in two yeast species descended from a whole-genome duplication. Proc Natl Acad Sci U S A. 2007;104:8397–8402. doi: 10.1073/pnas.0608218104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Souciet JL, Dujon B, Gaillardin C, Johnston M, Baret PV, Cliften P, Sherman DJ, Weissenbach J, Westhof E, Wincker P, et al. Comparative genomics of protoploid Saccharomycetaceae. Genome Res. 2009;19:1696–1709. doi: 10.1101/gr.091546.109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Galeote V, Bigey F, Devillers H, Neuvéglise C, Dequin S. Genome Sequence of the Food Spoilage Yeast Zygosaccharomyces bailii CLIB 213T. Genome Announc. 2013;1:e00606–13. doi: 10.1128/genomeA.00606-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Sarilar V, Devillers H, Freel KC, Schacherer J, Neuvéglise C. Draft Genome Sequence of Lachancea lanzarotensis CBS 12615T, an Ascomycetous Yeast Isolated from Grapes. Genome Announc. 2015;3:e00292–15. doi: 10.1128/genomeA.00292-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Dietrich FS, Voegeli S, Kuo S, Philippsen P. Genomes of Ashbya fungi isolated from insects reveal four mating-type loci, numerous translocations, lack of transposons, and distinct gene duplications. G3 (Bethesda) 2013;3:1225–39. doi: 10.1534/g3.112.002881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Wendland J, Walther A. Genome evolution in the Eremothecium clade of the Saccharomyces complex revealed by comparative genomics. G3 (Bethesda) 2011;1:539–48. doi: 10.1534/g3.111.001032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Wendland J, Walther A. Chromosome number reduction in Eremothecium coryli by two telomere-to-telomere fusions. Genome Biol Evol. 2014;6:1186–98. doi: 10.1093/gbe/evu089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Lertwattanasakul N, Kosaka T, Hosoyama A, Suzuki Y, Rodrussamee N, Matsutani M, Murata M, Fujimoto N, Suprayogi, Tsuchikane K, et al. Genetic basis of the highly efficient yeast Kluyveromyces marxianus: complete genome sequence and transcriptome analyses. Biotechnol Biofuels. 2015;8:47. doi: 10.1186/s13068-015-0227-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Giorello FM, Berná L, Greif G, Camesasca L, Salzman V, Medina K, Robello C, Gaggero C, Aguilar PS, Carrau F. Genome Sequence of the Native Apiculate Wine Yeast Hanseniaspora vineae T02/19AF. Genome Announc. 2014;2:e00530–14. doi: 10.1128/genomeA.00530-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Freel KC, Sarilar V, Neuvéglise C, Devillers H, Friedrich A, Schacherer J. Genome Sequence of the Yeast Cyberlindnera fabianii (Hansenula fabianii) Genome Announc. 2014;2:e00638–14. doi: 10.1128/genomeA.00638-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Tomita Y, Ikeo K, Tamakawa H, Gojobori T, Ikushima S. Genome and transcriptome analysis of the food-yeast Candida utilis. PLoS One. 2012;7:e37226. doi: 10.1371/journal.pone.0037226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Schneider J, Rupp O, Trost E, Jaenicke S, Passoth V, Goesmann A, Tauch A, Brinkrolf K. Genome sequence of Wickerhamomyces anomalus DSM 6766 reveals genetic basis of biotechnologically important antimicrobial activities. FEMS Yeast Res. 2012;12:382–6. doi: 10.1111/j.1567-1364.2012.00791.x. [DOI] [PubMed] [Google Scholar]
- 92.Schneider J, Andrea H, Blom J, Jaenicke S, Rückert C, Schorsch C, Szczepanowski R, Farwick M, Goesmann A, Pühler A, et al. Draft genome sequence of Wickerhamomyces ciferrii NRRL Y-1031 F-60-10. Eukaryot Cell. 2012;11:1582–3. doi: 10.1128/EC.00258-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.De Schutter K, Lin Y-C, Tiels P, Van Hecke A, Glinka S, Weber-Lehmann J, Rouzé P, Van de Peer Y, Callewaert N. Genome sequence of the recombinant protein production host Pichia pastoris. Nat Biotechnol. 2009;27:561–6. doi: 10.1038/nbt.1544. [DOI] [PubMed] [Google Scholar]
- 94.Liu X, Kaas RS, Jensen PR, Workman M. Draft genome sequence of the yeast Pachysolen tannophilus CBS 4044/NRRL Y-2460. Eukaryot Cell. 2012;11:827. doi: 10.1128/EC.00114-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Morales L, Noel B, Porcel B, Marcet-Houben M, Hullo M-F, Sacerdot C, Tekaia F, Leh-Louis V, Despons L, Khanna V, et al. Complete DNA sequence of Kuraishia capsulata illustrates novel genomic features among budding yeasts (Saccharomycotina) Genome Biol Evol. 2013;5:2524–39. doi: 10.1093/gbe/evt201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Piškur J, Ling Z, Marcet-Houben M, Ishchuk OP, Aerts A, LaButti K, Copeland A, Lindquist E, Barry K, Compagno C, et al. The genome of wine yeast Dekkera bruxellensis provides a tool to explore its food-related properties. Int J Food Microbiol. 2012;157:202–9. doi: 10.1016/j.ijfoodmicro.2012.05.008. [DOI] [PubMed] [Google Scholar]
- 97.Chan GF, Gan HM, Ling HL, Rashid NAA. Genome sequence of Pichia kudriavzevii M12, a potential producer of bioethanol and phytase. Eukaryot Cell. 2012;11:1300–1. doi: 10.1128/EC.00229-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Riccombeni A, Vidanes G, Proux-Wéra E, Wolfe KH, Butler G. Sequence and Analysis of the Genome of the Pathogenic Yeast Candida orthopsilosis. PLoS One. 2012;7:e35750. doi: 10.1371/journal.pone.0035750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Kim J-S, Baek J-H, Park N-H, Kim C. Complete Genome Sequence of Halophilic Yeast Meyerozyma caribbica MG20W Isolated from Rhizosphere Soil. Genome Announc. 2015;3:e00127–15. doi: 10.1128/genomeA.00127-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Chatterjee S, Alampalli SV, Nageshan RK, Chettiar ST, Joshi S, Tatu US. Draft genome of a commonly misdiagnosed multidrug resistant pathogen Candida auris. BMC Genomics. 2015;16:686. doi: 10.1186/s12864-015-1863-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Hershkovitz V, Sela N, Taha-Salaime L, Liu J, Rafael G, Kessler C, Aly R, Levy M, Wisniewski M, Droby S. De-novo assembly and characterization of the transcriptome of Metschnikowia fructicola reveals differences in gene expression following interaction with Penicillium digitatum and grapefruit peel. BMC Genomics. 2013;14:168. doi: 10.1186/1471-2164-14-168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Kunze G, Gaillardin C, Czernicka M, Durrens P, Martin T, Böer E, Gabaldón T, Cruz JA, Talla E, Marck C, et al. The complete genome of Blastobotrys (Arxula) adeninivorans LS3 – a yeast of biotechnological interest. Biotechnol Biofuels. 2014;7:66. doi: 10.1186/1754-6834-7-66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Polev DE, Bobrov KS, Eneyskaya EV, Kulminskaya AA. Draft Genome Sequence of Geotrichum candidum Strain 3C. Genome Announc. 2014;2:e00956–14. doi: 10.1128/genomeA.00956-14. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 104.Vega-Alvarado L, Gómez-Angulo J, Escalante-García Z, Grande R, Gschaedler-Mathis A, Amaya-Delgado L, Sanchez-Flores A, Arrizon J. High-Quality Draft Genome Sequence of Candida apicola NRRL Y-50540. Genome Announc. 2015;3:e00437–15. doi: 10.1128/genomeA.00437-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Matsuzawa T, Koike H, Saika A, Fukuoka T, Sato S, Habe H, Kitamoto D, Morita T. Draft Genome Sequence of the Yeast Starmerella bombicola NBRC10243, a Producer of Sophorolipids, Glycolipid Biosurfactants. Genome Announc. 2015;3:e00176–15. doi: 10.1128/genomeA.00176-15. [DOI] [PMC free article] [PubMed] [Google Scholar]