

Intercellular communication between neurons and glial cells via extracellular vesicles (EVs) as a novel mechanism of information transfer has been shown to be involved in regeneration processes within the central nervous system (CNS) (Rajendran et al., 2014). Hence, to take advantage of EV signaling for therapeutic applications appears to be a completely new approach to promote regeneration. One fundamental reason why influencing neuronal recovery after damage remains problematic is due to limited access to molecular processes, which are essential for the transcription of growth-promoting cues. Interestingly, EVs are currently under investigation for their use as hybrid vectors containing both therapeutic agents and superparamagnetic iron oxide nanoparticles (SPIOs) that can be remote guided using an external magnetic field (Silva et al., 2015).
In general, EVs are defined by their size and type of release mechanism from the cell of origin. EVs are secreted by neurons, astrocytes, microglia and oligodendrocytes in the CNS, as well as by neurons and glia in the peripheral nervous system (PNS). They are referred to as microvesicles with sizes ranging from 100 to 1,000 nm when derived from direct budding of the plasma membrane, and as exosomes with sizes ranging from 40 to 100 nm when arising from ectocytosis of multivesicular bodies (Turturici et al., 2014). The latter have been described more intensively because they were shown to be implicated in various cellular functions and disease states and, therefore, could constitute valuable biomarkers (Simons and Raposo, 2009). EVs carry distinct, functionally active cargos such as genetic material, proteins or lipids, which depend on the cell type they were secreted from (Bellingham et al., 2012; Kalani et al., 2014). The transfer of these types of information is thought to influence cellular phenotypes through reprogramming local or distal recipient cells. This certainly applies to exosomes transporting mRNA or miRNA, which can regulate gene expression or silencing in the target cell (Camussi et al., 2010; Kawikova and Askenase, 2015). Furthermore, it has been shown that exosome membranes contain different lipids such as phosphatidylcholine, phosphatidylserine or sphingomyelin that are recognized by recipient cells, providing potential markers for targeting (Kalani et al., 2014).
The intercellular transfer of disease particles within the CNS via exosomes has been shown to substantially contribute to the progression of neurodegeneration (Kalani et al., 2014). In neurodegenerative disorders, including Alzheimer's, Parkinson's, and Huntington's diseases, as well as amyotrophic lateral sclerosis EVs have been implicated in the delivery of mutated or misfolded proteins. For instance, in Alzheimer's disease the spread of neurotoxic oligomeric fibrils of the amyloid-beta (Aβ) peptide involves EVs signaling. This is due to the fact that the amyloid precursor protein is proteolytically cleaved at the plasma membrane with subsequent uptake into endosomes that can enter multivesicular bodies. It has been proposed that the secretion of exosomes containing Aβ peptides could therefore be involved in the extracellular accumulation of neuropathologic plaques that are a typical feature in Alzheimer's disease. It has also been discussed that EVs might be involved in the spatiotemporal seeding of the pathology within the CNS. Studies using mouse models for Parkinson's disease showed that EVs are possibly implicated in the transfer of aggregated alpha-synuclein between brain cells (Lai et al., 2012). However, the actual impact of EV signaling in disease progression is not yet understood and still is under intensive investigation. In contrast, when shed from endothelial cells and astrocytes as a consequence of blood-brain barrier breakdown in ischemia, nucleoside triphosphate diphosphohydrolase containing EVs are capable of providing neuroprotection by degrading toxic ATP (Lai et al., 2012). Therefore, it seems plausible to interfere with detrimental EV signaling for treatment of CNS diseases by manipulating their cargo.
Advantages EVs exhibit for therapeutic applications are their low immunogenicity, their unique delivery capability including their long circulation half-life as well as their blood-brain barrier passage. One approach already successfully applied for targeting brain cells of mice was to use self-derived dendritic cells to produce exosomes (Kalani et al., 2014; Kawikova and Askenase, 2015). These cells were genetically engineered to express Lamp2b, an exosomal membrane protein, fused with rabies glycoprotein for targeting brain cells. Following isolation, exosomes were loaded with siRNA by electroporation and intravenously injected into mouse to specifically deliver siRNA (for GAPDH) to the neurons, microglia, and oligodendrocytes in the brain, resulting in specific gene knockdown. Another therapeutic approach used exosomes as carriers for targeted drug delivery in a mouse model for Multiple Sclerosis, the myelin oligodendrocyte glycoprotein-induced experimental autoimmune encephalomyelitis (Kalani et al., 2014). Here, exosomal curcumin and JS1124 (a signal transducer and activator of stat3 inhibitor) was delivered to the brain via the intranasal route, which significantly decreased inflammation.
The encapsulation of biomolecules using exosomes or EVs in general constitutes a novel concept for influencing neuronal regeneration because, as mentioned before, their membrane composition identifies their origin and determines their fate (Kalani et al., 2014). One promising strategy for using exosomes or EVs in therapeutic applications is to isolate and then loading them with substances that can be easily tracked and guided for targeted delivery. In this context, the approach using superparamagnetic iron oxide nanoparticles as cargo together with therapeutic agents for effective targeting on the molecular level via an external magnetic field shows great potential in terms of neuroregeneration and neuroprotection (Silva et al., 2015).
SPIOs are ideal cargo candidates due to their small size, biocompatibility and unique magnetic properties. They have been specifically applied as contrast agents in magnetic resonance imaging (MRI) (Weinstein et al., 2010). SPIOs are defined as particles with at least one dimension between 1 to 100 nm in size that exhibit characteristic features depending on their surface coating, electrical charge, shape and hydrodynamic diameter. These properties furthermore determine SPIO behavior in biological fluids, their interaction with cells and the extracellular matrix, and their uptake and degradation (Lunov et al., 2010; Ludwig et al., 2013; Neubert et al., 2015). Furthermore, SPIOs can be surface functionalized to enhance their cellular uptake and transport drugs that can be subsequently released in high concentrations at the site of interest (Silva et al., 2015).
A recent study by Silva et al. (2015) demonstrated that macrophage-derived EVs can be loaded with both SPIOs and chemotherapeutic agents, and remote guided by an external magnetic force to be released at cancer cells. This approach took advantage of microvesicles that are, like exosomes, predominantly secreted as a consequence of physical or chemical cell stress (Turturici et al., 2014). In the study, an immortalized cell line of human monocytes, namely THP1 macrophages, was co-incubated with nanoparticles and chemotherapeutic agents. Subsequently, macrophages were stressed by serum depletion, leading to shedding of microvesicles that were also loaded with nanoparticles as verified by magnetic sorting. These experiments showed that SPIO-loaded microvesicles can be magnetically targeted to cancer cells as a naturally derived drug delivery system that is spatially controllable. In addition, detrimental off-target effects can be avoided and drug-related toxicity reduced (Silva et al., 2015).
SPIOs have also been shown to be taken up by different brain cells in vitro and in vivo. In our recent study, we systematically analyzed the effects of four different types of clinically relevant SPIOs on murine primary brain cells (Neubert et al., 2015). We applied the European Medicines Agency-approved ferucarbotran and the US Food and Drug Administration (FDA)-approved ferumoxytol. Carboxydextran-coated ferucarbotran is a contrast agent for liver imaging in humans that exhibits a hydrodynamic diameter of 60 nm. Carboxymethyldextran-coated ferumoxytol, with a hydrodynamic diameter of 30 nm, was originally developed for treating iron-deficiency anemia in patients with chronic kidney disease, and is currently used as a blood pool contrast agent for visualizing brain vascular malformations and creating cerebral blood volume maps with MRI. Furthermore, in our study we included two different types of novel, citrate-coated very small iron oxide particles (VSOP-R1, VSOP-R2) that have been tested in human Phase II clinical trials (Taupitz et al., 2004). These nanoparticles are of special interest because their small size of around 7 nm prolongs their blood half-life and facilitates their cellular incorporation, which could be beneficial for therapeutic interventions.
Our results showed that these four different types of SPIOs have different effects on the morphology of primary hippocampal neurons depending on their surface coating and respective charge, size, and concentration, as well as on the cell culture and incubation condition. Interestingly, the applied SPIOs induced degeneration of neurons in monoculture, whereas they promoted neurite outgrowth in neurons from neuron-glia co-cultures in a concentration and exposure time-dependent manner. For instance, lower concentrations of ferucarbotran and high concentrations of VSOP-R2 stimulated neurite outgrowth (Neubert et al., 2015). Based on our findings and those of other investigators, this effect might be due to the characteristic physicochemical properties of the SPIOs themselves, but could otherwise also be due to EV signaling within the neuron-glia co-culture. There is a considerable possibility that SPIOs were incorporated into EVs, constituting a positive stimulus for promoting neurite outgrowth. In this regard, it has been shown that one SPIO type we applied, namely VSOP, was internalized after being attached to the extracellular membrane (Lunov et al., 2010; Ludwig et al., 2013). In our study, we observed that VSOPs are attached to or internalized by primary neurons in monocultures, showing SPIO-neuron interaction (Figure 1A–C). This increases the probability that SPIOs are encapsulated in vesicles due to EV production through membrane invaginations, forming multivesicular bodies and subsequently exosomes, or by direct budding of the cell membrane, generating microvesicles. It would therefore be possible to interfere with EV signaling for therapeutic applications, for instance, by using cargos that contain specifically functionalized SPIOs that promote neuronal regeneration. Furthermore, EVs could be manipulated to carry both substances stimulating neuronal regeneration such as growth factors and magnetic SPIOs that can be easily tracked and remote guided. For example EVs containing SPIOs and substances to promote neurite outgrowth can be remote guided by an external magnetic stimulus to be released at recipient neurons. Following EV uptake by the recipient functionally restricted neurons and SPIO unpacking, therapeutic substances are systemically released and SPIOs systemically degraded. Free iron is thought to enter cellular iron metabolism. Within the intracellular space, therapeutic substances could subsequently activate intracellular signaling cascades or the transcription or silencing of corresponding genes, leading to increased neurite outgrowth. The elevated number of neurites could induce increased synaptic connectivity between neurons and result in functional recovery.
Figure 1.
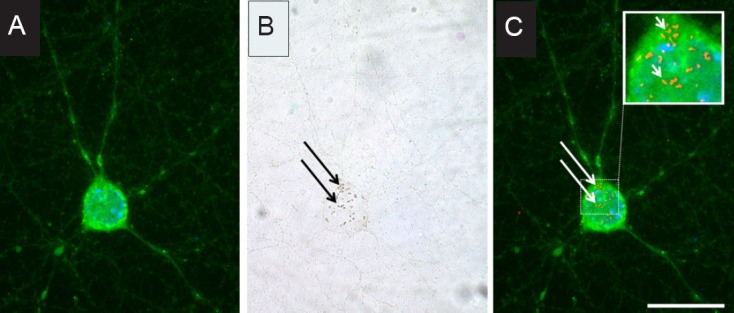
Representative images of murine primary hippocampal neurons (10 DIV) incubated with 0.5 mM of VSOP-R1 for 24 hours.
(A) Following VSOP exposure, neurons were stained with neuron-specific class III β-tubulin antibody Tuj1 (green fluorescence) and the nucleic acid stain Hoechst 33258 (blue fluorescence). (B) The neuron from (A) was captured as bright field image to show the localization of VSOPs that are visible as brown spots and are pointed out by arrows. The image shows VSOPs being attached to or internalized by neurites and the soma. (C) The overlay of pseudo-colored VSOPs in red (arrows) with green fluorescent neurons demonstrates particle localization and suggests possible ways of interaction. Scale bar: 40 μm. VSOP: Very small superparamagnetic iron oxide particles.
Previous studies already demonstrated that of EVs are found in synapses, which could constitute another, so far unanticipated mechanisms involved in neurite outgrowth (Smalheiser, 2007; Lachenal et al., 2011). It has been shown that neuronal exosome secretion is regulated by calcium influx and by glutamatergic synaptic activity affecting both, presynaptic and postsynaptic events of cortical and hippocampal neurons. At the presynaptic side as a consequence of depolarization, calcium is thought to be responsible for the fusion of multivesicular bodies, subsequently leading to exosome secretion. Neuronal exosomes have been shown to contain the neuronal cell adhesion protein L1 and GluR2/3 subunits of glutamate AMPA receptors, which affect the excitability, receptor availability and plasticity (Lai et al., 2012). Furthermore, it has been proposed that exosomes are involved in retrograde signaling across synapses, where they are released from the lipid raft region of the postsynaptic membrane following stimuli that elicit long-term potentiation (Smalheiser, 2007). Here, the transfer of synaptic proteins (such as CAM kinase II alpha) and synaptic RNAs to the presynaptic terminal enables synaptic plasticity. These findings support the fact that exosomes are influencing intercellular connections, thereby promoting the recovery of neuronal signaling that is one among other fundamental requirements for neurite outgrowth.
However, the potential application of EVs or exosomes that carry therapeutic biomolecules and are labeled with SPIOs for visual tracking should be thoroughly investigated to verify target specific substance release as well as coordinated SPIO uptake and secretion. On the one hand, it is necessary to check for intrinsic mechanisms preventing the degradation of encapsulated SPIOs in multivesicular bodies that might not undergo ectocytosis but could also be prone to uptake by acidophilic lysosomes. The degradation or uncontrolled accumulation of SPIOs within cells of the CNS can potentially cause adverse effects (Neubert et al., 2015). Cellular reactions critically depend on the respective nanoparticle properties, including composition, size, and surface coating. For instance, high surface-to-volume ratios cause increased reactivity of SPIOs with surrounding tissue which can influence cell physiology and morphology. Following SPIO degradation, release of free iron ions affects distinct subcellular processes and can enforce mitochondrial dysfunction through the production of reactive oxygen species (Neubert et al., 2015). Therefore, the physicochemical properties of SPIOs and their interaction with biological tissue have to be thoroughly investigated in advance.
On the other hand, SPIO binding and accumulation at the extracellular membrane and the subsequent internalization in EVs have to be ensured. It is also certainly necessary to characterize SPIO behavior under standardized conditions in cell cultures of individual cells as well as mixed cell cultures, for instance, containing neurons, astrocytes and microglial cells. Under these conditions, the possibility of influencing EV cargos and their delivery could indeed open up new strategies for affecting information transfer within the brain to promote neuroregeneration.
This study was supported by DFG Grant KFO 213 to JG.
References
- Bellingham SA, Guo BB, Coleman BM, Hill AF. Exosomes: vehicles for the transfer of toxic proteins associated with neurodegenerative diseases? Front Physiol. 2012;3:124. doi: 10.3389/fphys.2012.00124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Camussi G, Deregibus MC, Bruno S, Cantaluppi V, Biancone L. Exosomes/microvesicles as a mechanism of cell-to-cell communication. Kidney Int. 2010;78:838–848. doi: 10.1038/ki.2010.278. [DOI] [PubMed] [Google Scholar]
- Kalani A, Tyagi A, Tyagi N. Exosomes: mediators of neurodegeneration, neuroprotection and, therapeutics. Mol Neurobiol. 2014;49:590–600. doi: 10.1007/s12035-013-8544-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kawikova I, Askenase PW. Diagnostic and therapeutic potentials of exosomes in CNS diseases. Brain Res. 2015;1617:63–71. doi: 10.1016/j.brainres.2014.09.070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lachenal G, Pernet-Gallay K, Chivet M, Hemming FJ, Belly A, Bodon G, Blot B, Haase G, Goldberg Y, Sadoul R. Release of exosomes from differentiated neurons and its regulation by synaptic glutamatergic activity. Mol Cell Neurosci. 2011;46:409–418. doi: 10.1016/j.mcn.2010.11.004. [DOI] [PubMed] [Google Scholar]
- Lai CP, Breakefield XO. Role of exosomes/microvesicles in the nervous system and use in emerging therapies. Front Physiol. 2012;3:228. doi: 10.3389/fphys.2012.00228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ludwig A, Poller WC, Westphal K, Minkwitz S, Lattig-Tunnemann G, Metzkow S, Stangl K, Baumann G, Taupitz M, Wagner S, Schnorr J, Stangl V. Rapid binding of electrostatically stabilized iron oxide nanoparticles to THP-1 monocytic cells via interaction with glycosaminoglycans. Basic Res Cardiol. 2013;108:328. doi: 10.1007/s00395-013-0328-2. [DOI] [PubMed] [Google Scholar]
- Lunov O, Syrovets T, Rocker C, Tron K, Nienhaus GU, Rasche V, Mailander V, Landfester K, Simmet T. Lysosomal degradation of the carboxydextran shell of coated superparamagnetic iron oxide nanoparticles and the fate of professional phagocytes. Biomaterials. 2010;31:9015–9022. doi: 10.1016/j.biomaterials.2010.08.003. [DOI] [PubMed] [Google Scholar]
- Neubert J, Wagner S, Kiwit J, Brauer AU, Glumm J. New findings about iron oxide nanoparticles and their different effects on murine primary brain cells. Int J Nanomedicine. 2015;10:2033–2049. doi: 10.2147/IJN.S74404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rajendran L, Bali J, Barr MM, Court FA, Kramer-Albers EM, Picou F, Raposo G, van der Vos KE, van Niel G, Wang J, Breakefield XO. Emerging roles of extracellular vesicles in the nervous system. J Neurosci. 2014;34:15482–15489. doi: 10.1523/JNEUROSCI.3258-14.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silva AK, Luciani N, Gazeau F, Aubertin K, Bonneau S, Chauvierre C, Letourneur D, Wilhelm C. Combining magnetic nanoparticles with cell derived microvesicles for drug loading and targeting. Nanomedicine. 2015;11:645–655. doi: 10.1016/j.nano.2014.11.009. [DOI] [PubMed] [Google Scholar]
- Simons M, Raposo G. Exosomes--vesicular carriers for intercellular communication. Curr Opin Cell Biol. 2009;21:575–581. doi: 10.1016/j.ceb.2009.03.007. [DOI] [PubMed] [Google Scholar]
- Smalheiser NR. Exosomal transfer of proteins and RNAs at synapses in the nervous system. Biol Direct. 2007;2:35. doi: 10.1186/1745-6150-2-35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taupitz M, Wagner S, Schnorr J, Kravec I, Pilgrimm H, Bergmann-Fritsch H, Hamm B. Phase I clinical evaluation of citrate-coated monocrystalline very small superparamagnetic iron oxide particles as a new contrast medium for magnetic resonance imaging. Invest Radiol. 2004;39:394–405. doi: 10.1097/01.rli.0000129472.45832.b0. [DOI] [PubMed] [Google Scholar]
- Turturici G, Tinnirello R, Sconzo G, Geraci F. Extracellular membrane vesicles as a mechanism of cell-to-cell communication: advantages and disadvantages. Am J Physiol Cell Physiol. 2014;306:C621–633. doi: 10.1152/ajpcell.00228.2013. [DOI] [PubMed] [Google Scholar]
- Weinstein JS, Varallyay CG, Dosa E, Gahramanov S, Hamilton B, Rooney WD, Muldoon LL, Neuwelt EA. Superparamagnetic iron oxide nanoparticles: diagnostic magnetic resonance imaging and potential therapeutic applications in neurooncology and central nervous system inflammatory pathologies, a, review. J Cereb Blood Flow Metab. 2010;30:15–35. doi: 10.1038/jcbfm.2009.192. [DOI] [PMC free article] [PubMed] [Google Scholar]