

Abstract
AtGRP3 is a glycine-rich protein (GRP) from Arabidopsis thaliana shown to interact with the receptor-like kinase AtWAK1 in yeast, in vitro and in planta. In this work, phenotypic analyses using transgenic plants were performed in order to better characterize this GRP. Plants of two independent knockout alleles of AtGRP3 develop longer roots suggesting its involvement in root size determination. Confocal microscopy analysis showed an abnormal cell division and elongation in grp3-1 knockout mutants. Moreover, we also show that grp3-1 exhibits an enhanced Aluminum (Al) tolerance, a feature also described in AtWAK1 overexpressing plants. Together, these results implicate AtGRP3 function root size determination during development and in Al stress.
Introduction
The plant glycine-rich proteins (GRPs) superfamily is characterized by the presence of variable semi-repetitive glycine-rich motifs. Based on these variations, this superfamily has been further divided into five distinct classes. This classification does not consider these protein functions due to the fact that only recently the first functional characterization studies were performed, elucidating some plant GRP activities [1].
Plant GRPs have been of scientific interest due to their tissue-specific, developmentally and/or stress modulated expression patterns (reviewed in [2]). GRPs have been identified in various plant species, and over 150 GRP genes were found in the transcriptome or whole-genome analysis of sugarcane, Eucalyptus, Arabidopsis and rice [3, 4]; V. Galvão, V. Cardeal and G. Sachetto-Martins, personal communication).
Functional characterization approaches have been conducted in order to study plant GRP function (reviewed in [1]). Most of these studies focused on Arabidopsis GRPs and have implicated plant GRPs in pollen hydration and competition [5], flowering [6]; [7], plant defense [8], RNA splicing [9], cell elongation [10], pri-miRNA processing [11] and various responses including cold and osmotic stress [12–20].
The AtGRP3 gene (At2g05520) was first isolated as a cDNA clone from Arabidopsis and Northern blot analysis indicated strong expression of this gene in leaves and inflorescence axis. The protein sequence contains a putative signal peptide, followed by a glycine-rich region with GGXXXGG motif and a cysteine-rich C-terminus [21]. This structure classifies AtGRP3 as a Class II GRP [1]. The cysteine-rich domain is necessary for the interaction of AtGRP3 with the extracellular domain of the wall associated kinase AtWAK1 [22]. AtWAK1 (At1g21250) is a receptor-like kinase (RLK) containing an extracellular, a transmembrane and a cytoplasmic kinase domain [23]. This gene is expressed throughout plant development and is induced by an analog of salicylic acid [24]. Sub-cellular localization experiments using GFP fusion indicated that AtWAK1 is initially localized to endomembrane system and then transported to the cell surface where it is co-localizes with pectin as shown by protoplast experiments [25, 26]. Domain swap studies showed that binding of oligogalacturonides to the extracellular domain of AtWAK1 triggers activation of the kinase domain eliciting defense responses against fungi and bacteria. Accordingly, plants overexpressing AtWAK1 are more resistant to the fungus Botrytis cinerea [27]. These plants also display an enhanced Al tolerance suggesting a role for AtWAK1 in Al signaling pathway [28].
AtGRP3/AtWAK1 binding has been shown not only through yeast two-hybrid experiments, but has also been confirmed in vitro and in planta. In addition, a protein complex involving AtGRP3/AtWAK1 and the kinase-associated protein phosphatase (KAPP) is formed in planta. AtGRP3 expression is induced by salicylic acid resulting in a positive feedback that stimulates further its expression as well as the expression of AtWAK1 and PR-1 in protoplasts, suggesting a role for AtGRP3 in plant defense and signaling [22].
Here, in order to elucidate the functional role of AtGRP3 throughout plant development and its possible involvement in AtWAK1-mediated Al signaling, knockout plants were characterized. Our results propose the participation of AtGRP3 in determining root size. These results are confirmed by confocal microscopy analysis, which indicates an abnormal cell division and cell elongation in grp3-1 knockout mutants. Finally, grp3-1 knockout plants presented enhanced Al tolerance, suggesting that AtGRP3 and AtWAK1 function in the same signaling pathway.
Material and Methods
Plant material
Growth conditions, root growth analysis were performed according to Mangeon and collaborators [10]. Root growth experiments in Al were performed according to Sivaguru and collaborators [28]. The growth measurements were performed 10 days after seedling transfer to plates containing Aluminum chloride hexahydrate, 99% (hereafter, Al).
T-DNA lines
The grp3-1 T-DNA mutant, SALK_084685, was isolated from the Salk Institute Genomic Analysis Laboratory collection [29]. Homozygous mutants were isolated by PCR-based genotyping using gene specific PCR primers G3 LP (5’CCAACGCTTTGAAAAAGTTAAA3´) and G3 RP (5´tgaattcactgtggctgtccaaa3´) together with LBa1 (5’TGGTTCACGTAGTGGGCCATCG3’). A second T-DNA insertion line, grp3-2 T-DNA mutant, SALK_012941c, was isolated from the Salk Institute Genomic Analysis Laboratory collection as an homozygous line.
Real-time quantitative PCR (RT-qPCR)
The RT-qPCR experiments were carried out on cDNAs synthesized from total RNA extracted from 5 days-old seedlings using Trizol (Thermo-Fischer) according to the manufacturer’s instructions. One μg of total RNA was pre-digested with RQ1 RNase-free DNase (Promega) following manufacturer’s protocol and was used to synthesize cDNA using Superscript III (Thermo-Fischer) according the manufacturer’s instructions. Real-time quantitative PCR reactions were performed using SYBR Select Master Mix (Thermo-Fischer) in standard conditions. TIP41 (At4g34270) and FDH (At5g43940) were used as reference genes. A list of primers and concentrations used is presented in S1 Table. Reactions were performed in an Applied Biosystems 7500 Fast real-time PCR system and results were analyzed according to LinReg PCR (HFRC) and qBase (Biogazelle).
For the expression analysis, five pools containing 10 plants each were used in the experiments. The plotted data is an arithmetic mean of the three pools presenting the observed trend, excluding the outliers. For each sample, three technical replicates were performed.
Confocal microscopy analyses
For confocal visualization of root cells, plants were stained with propidium iodide according to Truernit and collaborators [30]. Analyses were performed in a Leica TCS SPE instrument using settings for propidium iodide according to the manufacturer (Leica Microsystems). Measurements were performed using ImageJ software (NIH).
For root diameter and number of cell rows analysis, eight plants of each background were used. For root length analysis, one hundred cells for each background were measured at the root hair zone.
Statistical analysis
The phenotypic parameters were analyzed according to the Student test (t test) for comparison between arithmetic means of samples in which the variances are different. The probability of random events is 95% and only values of P<0.05 were considered.
Results
Phenotypic analyses indicate that AtGRP3 is involved in determining root size
In order to characterize the functional role of AtGRP3, a loss-of-function line was obtained. This T-DNA line from the Salk Collection presenting insertion in the 5’UTR was genotyped and homozygous lines were selected. Quantitative real-time PCR analysis demonstrated that this line, named grp3-1, corresponds to an effective knockout without detectable levels of transcripts (Fig 1A).
Fig 1. grp3-1 loss-of-function mutant analysis.
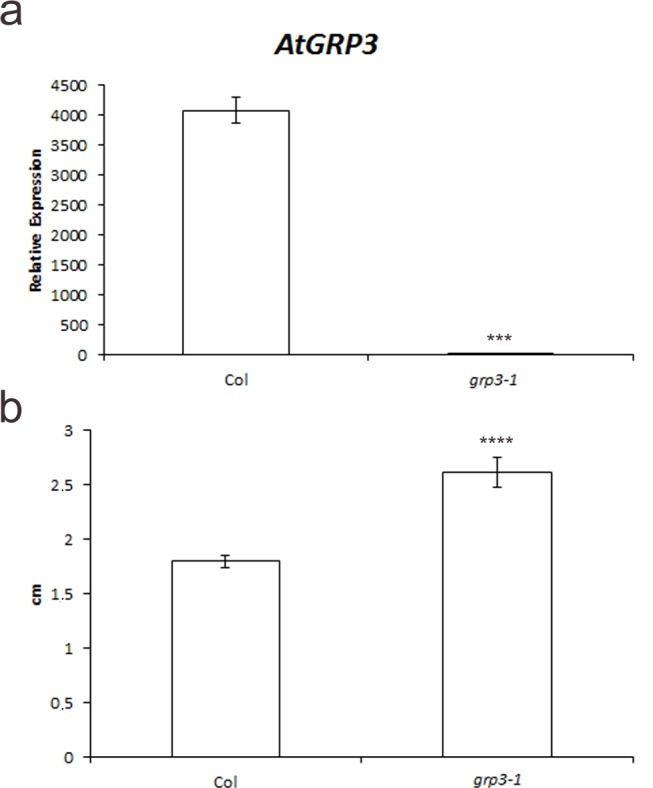
a Relative expression of AtGRP3 transcripts analyzed through real-time quantitative PCR of Col and grp3-1 mutant. b Summarized data for root length measurements of 2-week-old plants. Error bars indicate standard error. *** indicates p≤ 0.005 and **** indicates p≤ 0.001.
Phenotypical analyses of grp3-1 knockout plants were carried out throughout plant development and we observed that grp3-1 knockout plants presented a 45% increase in root length compared to Col, used as controls (Fig 1B).
In order to confirm if the observed phenotype was due to loss of AtGRP3 function, a second T-DNA line (named grp3-2) was also analyzed. Quantitative real-time PCR analysis indicates that this line is also a knockout allele (S1A Fig). Phenotypical analyses of grp3-2 were carried out in order to confirm the increase in root length observed in the other AtGPR3 mutant allele. Indeed, grp3-2 also present longer roots compared to Col (S1B Fig) corroborating the hypothesis that AtGRP3 is involved in determining root size.
Cell elongation and division markers are induced in grp3-1 knockout plants
The size of plant organs are controlled by two main processes: cell elongation and cell division [31–34]. In order to verify the cause for the enhanced root size observed in grp3-1 plants, the expression of genes known to be involved in these two processes was assessed.
First, genes involved in cell wall biosynthesis [35–38] and modification [39, 40] were tested. In grp3-1 plants, a 2-fold induction in the expression of both the cellulose biosynthesis regulator gene COBRA (COB, At5g60920) and the endo-1,4-β-glucanase gene KORRIGAN1 (KOR1, At5g49720) was observed (Fig 2A and 2B). Furthermore, a 220% increase in the cellulose synthase gene (CESA6, At5g64740) expression was also seen in grp3-1 compared to wild-type (Fig 2C). For the chitinase-like gene POM1 (At1g05850), an increase of 35% was detected in grp3-1 (Fig 2D).
Fig 2. Relative expression of cell elongation and/or division molecular markers in Col and grp3-1.
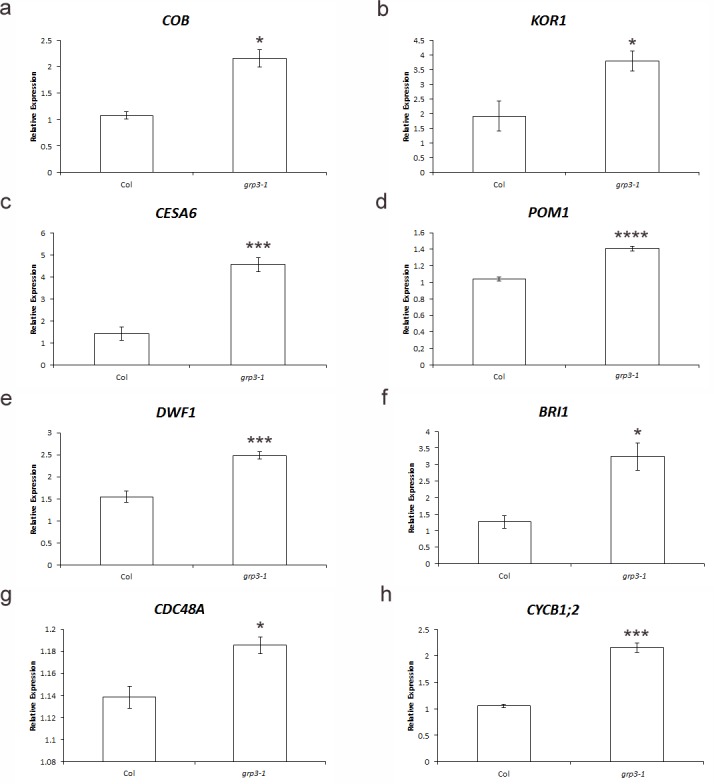
Quantitative real time PCR for a COB (At5g60920). b KOR1 (At5g49720). c CESA6 (At5g64740). d POM1 (At1g05850). e DWF1 (At3g19820). f BRI1 (At4g39400). g CDC48 (At3g09840). h CYCB1;2 (At5g06150). Error bars indicate standard error. * indicates p≤ 0.05, *** indicates p≤ 0.005 and **** indicates p≤ 0.001.
Since the phytohormone brassinosteroid is involved in cell elongation processes among other functions [41], genes involved in brassinosteroid biosynthesis [42] and signaling [43, 44] such as the brassinosteroid receptor gene BRI1 (At4g39400) and the brassinosteroid biosynthesis gene DWF1 (At3g19820) were also analyzed. A 60% and 157% increase over Col expression were detected for DWF1 and BRI1 in grp3-1, respectively (Fig 2E and 2F).
Genes involved in cell division [45] and cell cycle [46] were also tested. The cell division cycle gene CDC48A (At3g09840) presented a modest, but significant induction (3%) (Fig 2G) while the mitotic cyclin CYCB1;2 (At5g06150) had a 2-fold induction (Fig 2H).
Microscopy analysis reveals enhanced cell elongation and abnormal cell division in grp3-1 roots
In order to verify if cell division and elongation could be accounted for the enlarged root size phenotype seen in grp3-1 mutants, confocal microscopy analysis was conducted. For that matter, root cells were measured and counted. The first noticeable difference was in the division pattern of stele cells in grp3-1 compared to Col plants. While a small proportion of grp3-1 individuals presented a pattern of division similar to Col (Fig 3A and 3B), over 70% of the individuals presented disorganized stele cell rows (Fig 3C). It is important to note that, even when the pattern of division was normal, all grp3-1 plants presented extra rows of stele cells. On average, grp3-1 plants presents two extra rows of stele cells compared to Col (Fig 3D). This increase is reflected in a 20% increase of the root diameter of grp3-1 plants (Fig 3E).
Fig 3. Confocal analysis of root cells.

a-c Division pattern of stele cells. * labels stele rows. a Col wild-type. b grp3-1 individual presenting normal division pattern. c grp3-1 individual presenting abnormal division pattern. d Counting of number of stele cell rows. e Root diameter measurements. f Cell length measurements. Error bars indicate standard error. * indicates p≤ 0.05, ** indicates p≤ 0.01, *** indicates p≤ 0.005 and **** indicates p≤ 0.001.
In order to verify if cell elongation is also disturbed in grp3-1 mutants, root cell length in the maturation zone was measured. Root cells in grp3-1 plants were 35% longer than Col root cells (Fig 3F), indicating that cell elongation is indeed contributing for the increase in root size. These observations corroborate the data shown above of higher expression levels of several cell elongation molecular markers in the grp3-1 mutant.
grp3-1 presents an increased tolerance to Al
In a previous work, Sivaguru and collaborators [28] have reported that plants overexpressing AtWAK1 presented an increased tolerance to Al. Since AtGRP3 is capable of binding to AtWAK1 extracellular domain [22], we investigated if AtGRP3 was also involved in Al signaling by testing grp3-1 plants for Al tolerance.
Col plants submitted to Al presented an inhibition in root growth of 54%, while in grp3-1 plants this inhibition was reduced to 27% (Fig 4A). This data suggests, therefore, that as observed for plants overexpressing AtWAK1, grp3-1 knockout plants also present an increased tolerance to Al.
Fig 4. Analyses regarding implication of AtGRP3 in Al signaling in Col wild-type and grp3-1 knockout plants.

a Reduction in root growth resulting from Al exposure. Error bars indicate standard error. b Relative expression of AtWAK1 transcripts analyzed through real-time quantitative PCR of Col and grp3-1 mutant. Error bars indicate standard error. *** indicates p≤ 0.005 and **** indicates p≤ 0.001. c Quantitative real time PCR for AtGRP3 and AtWAK1 in Col plants submitted to Al for 0h and 8h. Error bars indicate standard error. * indicates p≤ 0.05 and **** indicates p≤ 0.001.
Park and collaborators [22] have shown that addition of AtGRP3 to protoplasts led to AtWAK1 expression induction. In order to check if AtGRP3 is involved in AtWAK1 endogenous expression, the levels of AtWAK1 were analyzed in the grp3-1 mutant. In fact, a 43% reduction of AtWAK1 expression levels was observed in the grp3-1 mutant compared to Col wild-type (Fig 4B).
Sivaguru and collaborators [28] have shown that AtWAK1 expression levels were induced in the presence of Al. In order to verify if AtGRP3 is also modulated by Al, Col plants were submitted to 8h of 100μM Al and the levels of AtGRP3 were checked. Differently from AtWAK1, AtGRP3 was not significantly modulated by Al (Fig 4C).
Discussion
The analysis of two mutant alleles hints for a possible role of AtGRP3 in determining root size. In plants, this control is regulated by two major events–cell division [33, 34] and cell elongation [31, 32]. A GRP gene from a different class–AtGRP5 –presented organ size phenotypes that were caused mainly by altering cell elongation [10]. We found a similar phenotype of altered cell length in grp3-1 knockout roots (Fig 3F) indicating that AtGRP3 is another GRP gene involved in regulating cell elongation processes. Functional analyses though, suggest that they have opposing roles in cell elongation, since AtGRP5 is a promoter [10] while AtGRP3 works as a repressor of cell elongation.
Corroborating AtGRP3/AtWAK1 interaction previously reported [22] and the possible role of AtGRP3 in root size determination, plants overexpressing AtWAK1 also present shorter roots compared to wild-type [28]. Interestingly, the levels of AtWAK1 are reduced in grp3-1 mutant (Fig 4B) which displays longer roots. This suggests that both AtGRP3 and AtWAK1 work as repressors of root growth.
Kohorn [47] has proposed a model in which WAKs, GRPs and pectin together regulate cell expansion. Years later, corroborating Kohorn’s model, Decreux and Messiaen [48] have demonstrated that AtWAK1 binds pectin in vitro. Our results are in agreement with this model since grp3-1 knockout plants present increased root cell length.
The most prominent phenotype of grp3-1 mutant is its root length (Fig 1B). The analysis of expression levels of genes known to be involved in cell wall deposition (COB, KOR1, CESA6), cell wall modification (POM1) and brassinosteroid signaling (BRI1) has shown to be upregulated in grp3-1 knockouts (Fig 2) which presents longer roots (Fig 1B). Agreeing with these data, null mutants for all those genes present shorter roots [35, 36, 39, 49].
Besides cell elongation, division also can be accounted for organ size [31–34]. In order to analyze if cell division markers are deregulated in the grp3-1 knockout mutant, the expression of several cell cycle-related genes was assigned (Fig 2, S1 Table). CYCB1;2 and CDC48A expression were in fact up-regulated in the grp3-1 mutant (Fig 2G and 2H). Corroborating these data, confocal microscopy analysis has shown that grp3-1 mutant present more stele cell rows (Fig 3A–3D). Interestingly, the analysis of a CDC48A mutant—a gene upregulated in the grp3-1 background—revealed a root tip free of stele cells [45].
Plants overexpressing the RLK AtWAK1 presented increased Al tolerance [28]. Since AtGRP3 binds to the extracellular domain of this protein [22], the idea that this signaling was also dependent of AtGRP3 is very tempting. It is expected that AtWAK1 overexpression plants contain an excess of AtWAK1 free of AtGRP3. With that idea in mind, grp3-1 plants, in which AtWAK1 free of AtGRP3 is also present, were tested for Al tolerance. In fact, grp3-1 knockout plants also displayed increased Al tolerance (Fig 4A). One hypothesis is that, in the presence of Al, AtGRP3 binding to AtWAK1 leads to physiological and morphological responses that result in root growth inhibition. Therefore, in the event of accumulation of AtWAK1 free of AtGRP3 (AtWAK1 overexpression or grp3-1 plants), this signaling is impaired resulting in Al tolerance.
Interestingly, the levels of AtWAK1 are induced by Al, while AtGRP3 levels are not significantly induced (Fig 4C). This could be a strategy to accumulate AtWAK1 free of AtGRP3 that, according to Kohorn [47], would lead to more cell expansion. The first symptom of Al toxicity is the inhibition of root elongation, which occurs around 1–2 h after exposition to Al [50]. This fast response indicates that Al primarily inhibits cell elongation and expansion, although, in the long term, cell division is also affected [50, 51]. By increasing AtWAK1 levels, the plant would enhance root elongation at least to a minimum, trying to overcome Al toxicity to some extent.
Our data indicates AtGRP3 as a repressor of root growth during plant development and upon Al stress. Collectively, these results points for functional orthologues of AtGRP3 as good targets for biotechnological approaches for Al tolerance, since knocking down these genes would not only lead to higher tolerance but also longer roots which could increases productivity.
Supporting Information
a Relative expression of AtGRP3 transcripts analyzed through real-time quantitative PCR of Col and grp3-2 mutant. b Summarized data for root length measurements of 1-week-old plants. Error bars indicate standard error. * indicates p≤ 0.05 and *** indicates p≤ 0.005.
(TIF)
(DOCX)
Acknowledgments
The authors would like to thank Ana Lucia Giannini and Régis Lopes Corrêa for critical reading of the manuscript and Luiza da Silva (FAPERJ technical assistant fellowship) for general technical support.
Abbreviations
- GRP
glycine-rich protein
- Al
Aluminum
- RLK
receptor-like kinase
- GFP
green fluorescence protein
- KAPP
kinase-associated protein phosphatase
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was supported by C/5249-1, http://www.ifs.se/; E26/110.117/2010, http://www.faperj.br/; 474392/2008-2, http://www.cnpq.br; PNPD2691094, http://www.capes.gov.br. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Mangeon A, Junqueira RM, Sachetto-Martins G. Functional diversity of the plant glycine-rich proteins superfamily. Plant Signal Behav. 2010; 5: 99–104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Sachetto-Martins G, Franco L, de Oliveira D. Plant glycine-rich proteins: a family or just proteins with a common motif? Bioch Biophys Acta. 2000; 1492: 1–14. [DOI] [PubMed] [Google Scholar]
- 3.Fusaro A, Mangeon A, Rocha C, Junqueira R, Coutinho T, Margis R, et al. Classification, expression pattern and comparative analysis of sugarcane expressed sequences tags (ESTs) encoding glycine-rich proteins (GRPs). Gen Mol Biol. 2001; 24: 263–273. [Google Scholar]
- 4.Bocca S.N, Magioli C, Mangeon A, Junqueira RM, Cardeal V, Margis R, et al. Survey of glycine-rich proteins (GRPs) in the Eucalyptus expressed sequence tag database (ForEST). Gen Mol Biol. 2005; 228: 608–624. [Google Scholar]
- 5.Mayfield JA, Preuss D. Rapid initiation of Arabidopsis pollination requires the oleosin-domain protein GRP17. Nat Cell Biol. 2000; 2: 128–130. [DOI] [PubMed] [Google Scholar]
- 6.Fusaro AF, Bocca SN, Ramos RL, Barrôco RM, Magioli C, Jorge VC, et al. AtGRP2, a cold-induced nucleo-cytoplasmic RNA-binding protein, has a role in flower and seed development. Planta. 2007; 225: 1339–1351. [DOI] [PubMed] [Google Scholar]
- 7.Streitner C, Danisman S, Wehrle F, Schoning JC, Alfano JR, Staiger D. The small glycine-rich RNA binding protein AtGRP7 promotes floral transition in Arabidopsis thaliana. Plant J. 2008; 56: 239–250. 10.1111/j.1365-313X.2008.03591.x [DOI] [PubMed] [Google Scholar]
- 8.Fu ZQ, Guo M, Jeong BR, Tian F, Elthon TE, Cerny RL, et al. A type III effector ADP-ribosylates RNA-binding proteins and quells plant immunity. Nature. 2007; 447: 284–288. [DOI] [PubMed] [Google Scholar]
- 9.Schoning JC, Streitner C, Meyer IM, Gao Y, Staiger D. Reciprocal regulation of glycine-rich RNA-binding proteins via an interlocked feedback loop coupling alternative splicing to nonsense-mediated decay in Arabidopsis. Nucleic Acids Res. 2008; 36: 6977–6987. 10.1093/nar/gkn847 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Mangeon A, Magioli C, Menezes-Salgueiro AD, Cardeal V, de Oliveira C, Galvao VC, et al. AtGRP5, a vacuole-located glycine-rich protein involved in cell elongation. Planta. 2009; 230: 253–265. 10.1007/s00425-009-0940-4 [DOI] [PubMed] [Google Scholar]
- 11.Köster T, Meyer K, Weinholdt C, Smith LM, Lummer M, Speth C, et al. Regulation of pri-miRNA processing by the hnRNP-like protein AtGRP7 in Arabidopsis. Nucl Acids Res. 2014; 42: 9925–9936. 10.1093/nar/gku716 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kwak KJ, Kim YO, Kang H. Characterization of transgenic Arabidopsis plants overexpressing GR-RBP4 under high salinity, dehydration, or cold stress. J Exp Bot. 2005; 56: 3007–3016. [DOI] [PubMed] [Google Scholar]
- 13.Kim YO, Kim JS, Kang H. Cold-inducible zinc finger-containing glycine-rich RNA-binding protein contributes to the enhancement of freezing tolerance in Arabidopsis thaliana. Plant J. 2005; 42: 890–900. [DOI] [PubMed] [Google Scholar]
- 14.Kim YO, Kang H. The role of a zinc finger-containing glycine-rich RNA-binding protein during the cold adaptation process in Arabidopsis thaliana. Plant Cell Physiol. 2006; 47: 793–798. [DOI] [PubMed] [Google Scholar]
- 15.Kim JS, Park SJ, Kwak KJ, Kim YO, Kim JY, Song J, et al. Cold shock domain proteins and glycine-rich RNA-binding proteins from Arabidopsis thaliana can promote the cold adaptation process in Escherichia coli. Nucleic Acids Res. 2007; 35: 506–516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kim JY, Park SJ, Jang B, Jung CH, Ahn SJ, Goh CH, et al. Functional characterization of a glycine-rich RNA-binding protein 2 in Arabidopsis thaliana under abiotic stress conditions. Plant J. 2007; 50: 439–451. [DOI] [PubMed] [Google Scholar]
- 17.Kim YO, Pan S, Jung CH, Kang H. A zinc finger-containing glycine-rich RNA-binding protein, atRZ-1a, has a negative impact on seed germination and seedling growth of Arabidopsis thaliana under salt or drought stress conditions. Plant Cell Physiol. 2007; 48: 1170–1181. [DOI] [PubMed] [Google Scholar]
- 18.Sasaki K, Kim MH, Imai R. Arabidopsis COLD SHOCK DOMAIN PROTEIN2 is a RNA chaperone that is regulated by cold and developmental signals. Biochem Biophys Res Commun. 2007; 364: 633–638. [DOI] [PubMed] [Google Scholar]
- 19.Kim JS, Jung HJ, Lee HJ, Kim KA, Goh CH, Woo Y, et al. Glycine-rich RNA-binding protein 7 affects abiotic stress responses by regulating stomata opening and closing in Arabidopsis thaliana. Plant J. 2008; 55: 455–466. 10.1111/j.1365-313X.2008.03518.x [DOI] [PubMed] [Google Scholar]
- 20.Park SJ, Kwak KJ, Oh TR, Kim YO, Kang H. old shock domain proteins affect seed germination and growth of Arabidopsis thaliana under abiotic stress conditions. Plant Cell Physiol. 2009; 50: 869–878. 10.1093/pcp/pcp037 [DOI] [PubMed] [Google Scholar]
- 21.de Oliveira DE, Seurinck J, Inzé D, Van Montagu M, Botterman J. Differential expression of five Arabidopsis genes encoding glycine-rich proteins. Plant Cell. 1990; 2: 427–436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Park AR, Cho SK, Yun UJ, Jin MY, Lee SH, Sachetto-Martins G, et al. Interaction of the Arabidopsis receptor kinase Wak1 with a glycine-rich protein, AtGRP-3. J Biol Chem. 2001; 276: 26688–26693. [DOI] [PubMed] [Google Scholar]
- 23.He Z-H, Cheeseman I, He D, Kohorn BD. A cluster of five cell wall-associated receptor kinase genes, Wak1–5, are expressed in specific organs of Arabidopsis. Plant Mol Biol. 1999; 39: 1189–1196. [DOI] [PubMed] [Google Scholar]
- 24.Wagner TA, Kohorn BD. Wall-associated kinases are expressed throughout plant development and are required for cell expansion. Plant Cell. 2001; 13: 303–318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kohorn B, Kobayashi M, Johansen S, Riese J, Huang LF, Koch K, et al. An Arabidopsis cell wall-associated kinase required for invertase activity and cell growth. Plant J. 2006; 46: 307–316. [DOI] [PubMed] [Google Scholar]
- 26.Kohorn B, Kobayashi M, Johansen S, Friedman HP, Fischer A, Byers N. Wall-associated kinase 1 (WAK1) is crosslinked in endomembranes, and transport to the cell surface requires correct cell-wall synthesis. J Cell Sci. 2006; 119: 2282–2290. [DOI] [PubMed] [Google Scholar]
- 27.Brutus A, Sicilia F, Macone A, Cervone F, de Lorenzo G. A domain swap approach reveals a role of the plant wall-associated kinase 1 (WAK1) as a receptor of oligogalacturonides. Proc Natl Acad Sci USA. 2010; 107: 9452–9457. 10.1073/pnas.1000675107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Sivaguru M, Ezaki B, He Z-H, Tong H, Osawa H, Baluška F, et al. Aluminum-induced gene expression and protein localization of a cell wall-associated receptor kinase in Arabidopsis. Plant Physiol. 2003; 132: 2256–2266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen H, Shinn P, et al. Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science. 2003; 301: 653–657. [DOI] [PubMed] [Google Scholar]
- 30.Truernit E, Bauby H, Dubreucq B, Grandjean O, Runions J, Barthélémy J, et al. High-resolution whole-mount imaging of three-dimensional tissue organization and gene expression enables the study of phloem development and structure in Arabidopsis. Plant Cell. 2008; 20: 1494–1503. 10.1105/tpc.107.056069 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Dolan L, Davies J. Cell expansion in roots. Curr Opin Plant Biol. 2004; 7: 33–39. [DOI] [PubMed] [Google Scholar]
- 32.Becker B. Function and evolution of the vacuolar compartment in green algae and land plants (Viridiplantae). Int Rev Cytol. 2007; 264: 1–24. [DOI] [PubMed] [Google Scholar]
- 33.Masuda HP, Cabral LM, de Veylder L, Tanurdzic M, Engler JA, Geelen D, et al. ABAP1 is a novel plant Armadillo BTB protein involved in DNA replication and transcription. EMBO J. 2008; 27: 2746–2756. 10.1038/emboj.2008.191 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Rojas CA, Eloy NB, Lima MF, Rodrigues RL, Franco LO, Himanen K, et al. Overexpression of the Arabidopsis anaphase promoting complex subunit CDC27a increases growth rate and organ size. Plant Mol Biol. 2009; 71: 307–318. 10.1007/s11103-009-9525-7 [DOI] [PubMed] [Google Scholar]
- 35.Roudier F, Fernandez AG, Fujita M, Himmelspach R, Borner GHH, Schindelman G, et al. COBRA, an Arabidopsis extracellular glycosyl-phosphatidyl inositol-anchored protein, specifically controls highly anisotropic expansion through its involvement in cellulose microfibril orientation. Plant Cell. 2005; 17: 1749–1763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Paredez AR, Persson S, Ehrhardt DW, Somerville CR. Genetic evidence that cellulose synthase activity influences microtubule cortical array organization. Plant Physiol. 2008; 147: 1723–1734. 10.1104/pp.108.120196 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Gutierrez R, Lindeboom JJ, Paredez AR, Emons AMC, Ehrhardt DW. Arabidopsis cortical microtubules position cellulose synthase delivery to the plasma membrane and interact with cellulose synthase trafficking compartments. Nat Cell Biol. 2009; 11: 797–806. 10.1038/ncb1886 [DOI] [PubMed] [Google Scholar]
- 38.Chebli Y, Kaneda M, Zerzour R, Geitmann A. The cell wall of the Arabidopsis pollen tube—spatial distribution, recycling, and network formation of polysaccharides. Plant Physiol. 2012; 160: 1940–1955. 10.1104/pp.112.199729 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Hauser M-T, Morikami A, Benfey PN. Conditional root expansion mutants of Arabidopsis. Development. 1995; 121: 1237–1252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Zhong R, Kays SJ, Schroeder BP, Ye Z-H. Mutation of a chitinase-like gene causes ectopic deposition of lignin, aberrant cell shapes, and overproduction of ethylene. Plant Cell. 2002; 14: 165–179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Belkhadir Y, Wang X, Chory J. Arabidopsis brassinosteroid signaling pathway. Sci STKE. 2006; 364: cm5. [DOI] [PubMed] [Google Scholar]
- 42.Choe S, Dilkes BP, Gregory BD, Ross AS, Yuan H, Noguchi T, et al. The Arabidopsis dwarf1 mutant is defective in the conversion of 24-methylenecholesterol to campesterol in brassinosteroid biosynthesis. Plant Physiol. 1999; 119: 897–907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.He Z, Wang ZY, Li J, Zhu Q, Lamb C, Ronald P, et al. Perception of brassinosteroids by the extracellular domain of the receptor kinase BRI1. Science. 2000; 288: 2360–2363. [DOI] [PubMed] [Google Scholar]
- 44.Kinoshita T, Caño-Delgado A, Seto H, Hiranuma S, Fujioka S, Yoshida S, et al. Binding of brassinosteroids to the extracellular domain of plant receptor kinase BRI1. Nature. 2005; 433: 167–171. [DOI] [PubMed] [Google Scholar]
- 45.Park S, Rancour DM, Bednarek SY. In planta analysis of the cell cycle-dependent localization of AtCDC48A and its critical roles in cell division, expansion, and differentiation. Plant Physiol. 2008; 148: 246–258. 10.1104/pp.108.121897 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Vandepoele K, Raes J, De Veylder L, Rouzé P, Rombauts S, Inzé D. Genome-wide analysis of core cell cycle genes in Arabidopsis. Plant Cell. 2002; 14: 903–916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Kohorn B. WAKs; cell wall associated kinases. Curr Opin Cell Biol. 2001; 13: 529–533. [DOI] [PubMed] [Google Scholar]
- 48.Decreux A, Messiaen J. Wall-associated kinase WAK1 interacts with cell wall pectins in a calcium-induced conformation. Plant Cell Physiol. 2005; 46: 268–278. [DOI] [PubMed] [Google Scholar]
- 49.Clouse SD, Langford M, McMorris TC. A brassinosteroid-insensitive mutant in Arabidopsis thaliana exhibits multiple defects in growth and development. Plant Physiol. 1996; 111: 671–678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Kochian LV. Cellular mechanisms of aluminium toxicity and resistance in plants. Ann Rev Plant Phys Plant Mol Biol. 1995; 46: 237–260. [Google Scholar]
- 51.Matsumoto H. Cell biology of aluminum toxicity and tolerance in higher plants. Int Rev Cytol. 2000; 200: 1–46. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
a Relative expression of AtGRP3 transcripts analyzed through real-time quantitative PCR of Col and grp3-2 mutant. b Summarized data for root length measurements of 1-week-old plants. Error bars indicate standard error. * indicates p≤ 0.05 and *** indicates p≤ 0.005.
(TIF)
(DOCX)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.