

Water diversion can pose a risk to resident or migratory fishes, but fish-protection devices that reduce the entrainment of native fishes and maintain normal diversion activities exist.
Keywords: Behaviour, conservation, management, sturgeon, swimming physiology, threatened species
Abstract
Diversion (i.e. extraction) of water from rivers and estuaries can potentially affect native wildlife populations if operation is not carefully managed. For example, open, unmodified water diversions can act as a source of injury or mortality to resident or migratory fishes from entrainment and impingement, and can cause habitat degradation and fragmentation. Fish-protection devices, such as exclusion screens, louvres or sensory deterrents, can physically or behaviourally deter fish from approaching or being entrained into water diversions. However, empirical assessment of their efficacy is often lacking or is investigated only for particular economically or culturally important fishes, such as salmonids. The Southern population of anadromous green sturgeon (Acipenser medirostris) is listed as threatened in California, and there is a high density of water diversions located within their native range (the Sacramento–San Joaquin watershed). Coupled with their unique physiology and behaviour compared with many other fishes native to California, the green sturgeon is susceptible to entrainment into diversions and is an ideal species with which to study the efficacy of mitigation techniques. Therefore, we investigated juvenile green sturgeon (188–202 days post-hatch) in the presence of several fish-protection devices to assess behaviour and entrainment risk. Using a large experimental flume (∼500 kl), we found that compared with an open diversion pipe (control), the addition of a trash-rack box, louvre box, or perforated cylinder on the pipe inlet all significantly reduced the proportion of fish that were entrained through the pipe (P = 0.03, P = 0.028, and P = 0.028, respectively). Likewise, these devices decreased entrainment risk during a single movement past the pipe by between 60 and 96%. These fish-protection devices should decrease the risk of fish entrainment during water-diversion activities.
Introduction
Worldwide, water diversions have been identified as a potential source of injury and mortality (Kimmerer, 2008; Baumgartner et al., 2009; Grimaldo et al., 2009), reduced fitness (Bennett, 2005; Kimmerer, 2008), or habitat degradation (Drinkwater and Frank, 1994; Kingsford, 2000) for many fish species. In particular, mortality can be caused indirectly as a result of habitat fragmentation, degradation or alteration (Liermann et al., 2012; Sheer and Steel, 2006), or directly from interactions with water diversions and associated structures (Kimmerer, 2008). Owing to the high density of water diversions in some locations (Herren and Kawasaki, 2001) and the large amount of water diverted from some river systems, entrainment into water diversions can pose a risk to several fishes, including those listed under the Endangered Species Act (ESA), such as delta smelt (Hypomesus transpacificus; Bennett, 2005) or green sturgeon (Acipenser medirostris; Mussen et al., 2014a). As a result, fish passage in proximity to water diversions has been a concern for scientists and managers, and guidance systems that confer fish protection at operational water diversions have been developed. Examples of fish-protection devices include positive barriers, such as exclusion screens, and behavioural barriers, such as louvres or sensory deterrents. Positive barriers are designed to physically prevent a fish from being entrained into a water diversion (Taft, 2000; USBR, 2006), whereas sensory deterrents and louvres modify the behaviour of fish to deter interactions between the fish and the diversion (USBR, 2006; Noatch and Suski, 2012).
Unfortunately, in some systems heavily altered by water diversions, the vast majority of diversion pipes remain unmodified by fish-protection devices (i.e. Herren and Kawasaki, 2001). For example, of the more than 3300 water diversions that extract water from the Sacramento–San Joaquin watershed, ∼98% of them are open, unmodified pipes that offer little to no protection for passing fishes (Herren and Kawasaki, 2001).
Despite the relatively widespread acceptance of the effectiveness of some fish-protection systems by managers, empirical investigations of the efficacy of many devices are lacking. While fish-exclusion screens can reduce the entrainment of fishes (Simpson and Ostrand, 2012; Boys et al., 2013), screens can be costly (McMichael et al., 2004; Moyle and Israel, 2005), difficult to maintain and install (USBR, 2006), and studies investigating screen-diversion impacts on fish populations are relatively scarce (Moyle and Israel, 2005). Additionally, some evidence suggests that traditional fish-protection systems, such as trash (or bar) racks or louvres, can be inefficient for certain species, particularly young-of-the-year juveniles or species without robust biomechanical and physiological capabilities (such as swimming performance; Kynard and Horgan, 2001; Amaral et al., 2002). For example, laboratory investigations of delta smelt and Chinook salmon (Oncorhynchus tshawytscha) behaviour near fish screens revealed that delta smelt were more likely to be injured from physical contact with screen faces and showed high rates of post-experimental mortality following repeated screen contacts or impingement (Swanson et al., 2004, 2005). Additionally, Chinook salmon were capable of increasing their swimming velocities with greater flow velocities and thus avoided screen contacts at high water velocities, while delta smelt were unable to do so. The performance differences observed between these two species underscore the importance of species-specific biomechanical and physiological functions in designing fish-protection systems that are effective for targeted fish species. Previous work has shown that closely related species (e.g. green vs. white sturgeon, Acipenser transmontanus) can have differential performance in the presence of fish-protection devices, with green sturgeon showing increased susceptibility to contact with and impingement upon fish-exclusion screens (Poletto et al., 2014a). Additionally, studies have investigated the importance of ontogeny in fish responses to water diversions (e.g. Nobriga et al., 2004) and have highlighted the importance of considering life stage and size-dependent physiology or behaviour when designing fish-protection devices.
Given that fishes have unique behavioural and physiological characteristics that make certain species more susceptible to entrainment, species-specific information is often required when implementing conservation and management practices. Detailed investigations into the behavioural, biomechanical, or physiological responses of fishes to such devices are needed, particularly for understudied fish species that often lack commercial or recreational value. For example, the green sturgeon is a species native to California with unique biomechanical, behavioural, and physiological characteristics when compared with salmonid species. Green sturgeon have two distinct population segments (DPS; the Northern and Southern DPS; Israel et al., 2004), the more southern of which is listed as ‘threatened’ under the ESA and is the focus of ongoing conservation and management efforts. Southern DPS green sturgeon spawn in the Sacramento–San Joaquin watershed in the Central Valley of California (Israel et al., 2004; Seesholtz et al., 2015), which is heavily modified by water projects (Herren and Kawasaki, 2001; Cloern and Jassby, 2012). Furthermore, their anadromous life-history strategy (Doroshov, 1985; Allen and Cech, 2006) may expose juveniles to thousands of large-scale water exports as they outmigrate from adult spawning grounds to juvenile rearing habitats. Previous investigations have shown that green sturgeon are susceptible to entrainment into unmodified water diversions and are limited in their ability to escape entrainment velocities (Mussen et al., 2014a). This may in part be due to their reduced swimming capabilities compared with salmonids (Peake et al., 1997) and other sturgeon species, such as shortnose (Acipenser brevirostrum), Adriatic (Acipenser naccarii) and lake sturgeon (Acipenser fulvescens), among others (Deslauriers and Kieffer, 2011). However, our previous work investigating the efficacy of a sensory deterrent and two structural pipe modifications (an upturned pipe and a terminal pipe plate) in reducing entrainment through a simulated water-diversion pipe showed that reduction of juvenile green sturgeon entrainment risk was possible (Poletto et al., 2014b). Given their propensity for becoming entrained into open water diversions (Mussen et al., 2014a, b) and impinged on traditional fish-protection devices (Poletto et al., 2014a), green sturgeon are an important candidate species for investigating entrainment and fish-protection devices in the laboratory.
Investigations of fish behaviour near fish-protection devices and quantification of how effective specific designs may be at preventing entrainment or injury to passing fish can help to improve mitigation efforts at water diversions and inform more effective conservation and management strategies. Therefore, we examined the efficacy of three fish-protection devices in reducing the entrainment of juvenile green sturgeon using a large river-simulation flume and an ‘over-the-levee’-style water-diversion pipe. The behaviour of juvenile green sturgeon in the presence of a trash-rack box, a louvre box, and a perforated cylinder was quantified and compared with that in the presence of a control (i.e. an open unmodified pipe). We hypothesized that the addition of each of the three protection devices would change fish passage past the pipe by altering entrainment and entrainment risk per pipe passage, as well as the number of successful escape attempts. Our results are informative to scientists and managers looking to mitigate the impacts of water-diversion activities on juvenile green sturgeon through the use of fish-protection devices.
Materials and methods
Fish tested
Northern DPS green sturgeon (F2) were spawned from green sturgeon broodstock at the University of California Davis (UC Davis) in November 2011 using established tank spawning methodologies (Van Eenennaam et al., 2001, 2012). Southern DPS fish are listed under the ESA as threatened, and no broodstock fish exist for this population. Fish were reared at the UC Davis Center for Aquatic Biology and Aquaculture in 815 l round fibreglass flow-through tanks at ambient well-water temperatures (18.0 ± 1.0°C) with continuous flows of aerated [dissolved oxygen (DO) 7.5 ± 1.0 mg O2/l], non-chlorinated fresh water. Water temperatures and DO levels were monitored daily, and fish were fed daily with semi-moist pellets (Rangen, Inc., Buhl, ID, USA), before transitioning to a dry pelleted diet (SilverCup™) at ∼60 days post-hatch. Daily food quantities were calculated using an optimal growth feeding model developed for the closely related white sturgeon (A. transmontanus; Deng et al., 2003; Lee et al., 2014) that describes the daily food ratios (as a percentage of body weight per day) necessary for optimal growth. All handling, rearing and experimental procedures used were approved by the UC Davis Institutional Animal Care and Use Committee (IACUC #15836).
Experimental flume
A large (∼500 kl), outdoor, recirculating flume was used to investigate the efficacy of structural fish-protection devices at reducing the entrainment of juvenile green sturgeon. The flume was constructed of a reinforced concrete floor with painted steel walls. The testing area within the flume was 18.29 m long, 3.05 m wide and 3.20 m deep (for detailed flume specifications see Mussen et al., 2013). To simulate an ‘over-the-levee’-style water diversion that is commonly found in the Sacramento–San Joaquin watershed (Dan Meier, United States Fish and Wildlife Service, personal communication), a painted steel diversion pipe 0.46 m in diameter was placed along one wall of the flume at approximately one-half the length of the flume, and projected into the flume at an angle of 26.6° over a simulated riverbank with a 26.6° angled bank (ramp; see Fig. 1). During experiments, the sweeping velocity of water moving through the flume was 0.15 m/s, the depth was 2.2 m, and the water-diversion rate through the diversion was maintained at 0.57 m3/s for all treatment conditions. These hydraulic conditions were found to entrain the highest number of juvenile green sturgeon in previous experiments (Mussen et al., 2014a) and allowed for comparisons with other fish species native to California (i.e. Chinook salmon; Mussen et al., 2014b, 2015), as well as with previously tested structural modifications and sensory deterrents (Poletto et al., 2014b).
Figure 1:

Images of modifications tested in each treatment condition. All images were taken downstream of the diversion pipe from a stationary underwater camera. (a) Control: no modifications were made to the diversion pipe. (b) Trash-rack box: a widened rectangular inlet was added to the end of the diversion pipe, and a trash rack consisting of 21 vertical steel bars was affixed to the inlet. (c) Louvre box: the same rectangular inlet as in the trash-rack box condition was added to the end of the pipe, and an angled louvre array consisting of 67 vertical steel louvres was affixed to the inlet. (d) Perforated cylinder: a cylindrical pipe extension was added to the end of the diversion pipe, and contained 25 rows of 12 circular holes covering two-thirds of the surface area of the cylinder.
Experiments
Experimental methodologies and behavioural quantifications followed those detailed by Poletto et al. (2014b) and Mussen et al. (2014a). Briefly, fish were tested during the day from late May until June 2012. For each 1 h experiment, 60 (±3) naïve juvenile green sturgeon aged 188–202 days post-hatch were tested within the flume at the same time. Six replicates were conducted for each experimental condition. Fish were either diverted through the pipe or remained within the flume testing area. At the conclusion of each experimental trial, fish were collected and individually weighed and measured [wet mass and fork length (FL)] separately. Underwater cameras (Speco CVC 320) were positioned at four locations within the flume to record fish behaviour near the diversion pipe, and an additional fifth camera (Sony CCD-TRV 108) was positioned directly above the centre of the pipe intake and used in combination with a clear Plexiglass acrylic 1.2 m2 viewing plate to provide an overhead view. Videos taken during each experiment were later analysed using a video-editing program (Sony Movie Studio 10), following procedures described by Mussen et al. (2013).
Fish were tested within the flume with the open, unmodified pipe (control) and three different fish-protection devices attached to the diversion pipe inlet: a trash-rack box, a louvre box, and a perforated cylinder. In control conditions, no modifications were made to the pipe inlet, and the diversion pipe remained open (Fig. 1a). The trash-rack box condition included the addition of a pipe extension that terminated in a widened rectangular opening with a vertical trash rack (Fig. 1b). The widened rectangular opening was 1.7 m wide, 0.76 m tall and 0.76 m deep. The trash rack was attached to the rectangular inlet and consisted of 21 vertical steel bars (1.3 cm wide × 0.32 cm thick) evenly spaced along the inlet opening every 7.6 cm. To compensate for the addition of the trash-rack box, the diversion pipe was shorted by 0.76 m relative to the control pipe length, which allowed the inlet opening for the trash-rack box to be in the identical location within the flume as was the control pipe. The louvre box condition used the same widened, rectangular extension as the trash-rack box, but included the addition of a vertical louvre array in place of the trash rack at the pipe inlet (Fig. 1c). Sixty-seven individual vertical steel louvres (6.35 cm wide × 0.32 cm thick) were evenly spaced 2.5 cm apart and positioned with the widest area of the louvre roughly perpendicular to that of the sweeping river velocity. The entire louvre array was positioned at a 15° angle to the sweeping river velocity, such that the downstream portion of the array extended an additional 47.5 cm into the main channel of the flume relative to the upstream portion. The perforated cylinder condition included a cylindrical pipe extension that was attached to the shortened diversion pipe in the same orientation (Fig. 1d). The cylindrical pipe extension was 0.91 m in diameter and 0.94 m long, with 25 rows of 12 circular holes, 5 cm in diameter and spaced 2.5 cm apart. The perforated circles covered roughly two-thirds of the surface of the cylinder, with the bottom third of the cylinder (closest to floor of the flume) remaining fully intact (i.e. unperforated).
Multiple distinct metrics related to entrainment were quantified, including fish passage and impingement. The number of fish that were entrained through the diversion pipe and the timing of each entrainment event was recorded and quantified. Additionally, the total number of times that fish swam past the pipe, regardless of direction (i.e. upstream to downstream of the pipe or downstream to upstream of the pipe) or orientation (i.e. positive or negative rheotaxis) was recorded, as well as the timing of each passage event. Therefore, entrainment was expressed in two ways, i.e. the proportion of fish entrained and the entrainment risk per pipe passage (EPP). The proportion of fish entrained through the diversion pipe for each trial was calculated as the total number of fish that were entrained relative to the total number of fish in the flume at the beginning of the experiment. The EPP for each trial was calculated as the total number of entrainment events relative to the total number of times that fish passed the diversion pipe. Escape attempts were also noted, and successful escape attempts that resulted in the fish avoiding entrainment once an entrainment event began (as determined by changes in body position or velocity) were also quantified. Owing to the physical nature of the structural fish-protection modifications used, it was possible for fish to become impinged on the louvre box, trash-rack box and perforated cylinder, but not the open-pipe control. An impingement event was considered to occur when more than two-thirds of the body of the fish remained in contact with the structure for >1 s. The number of impingement events was quantified in addition to the impingement escape rate (IER) and impingement duration. The IER was calculated as the number of times that fish were able to escape impingement successfully without becoming entrained through the diversion pipe, and the impingement duration was quantified for each impingement event as the length of time the fish remained impinged on the structure.
Data analyses
Data were analysed using R Studio version 2.15.2 software package (R Core Team, 2012). Statistical analyses and graphical depiction of data in R were performed using the R core package (R Core Team, 2012), ‘car’ (Fox and Weisberg, 2011), ‘plyr’ (Wickham, 2011), ‘lme4’ (Bates et al., 2014) and ‘ggplot2’ packages (Wickham, 2009). Fish mass was compared among treatments and entrainment status (i.e. diverted through the pipe or remaining in the flume) using a two-way ANOVA of ‘treatment’ and ‘entrainment status’. Simple effects of treatment and entrainment status on mass were analysed using one-way ANOVAs with subsequent Tukey's post hoc tests. The FL of fish was analysed using a generalized linear model, because of residual heteroscedasticity, using the same predictor variables (‘treatment’ and ‘entrainment status’) and a γ distribution. Model fit was evaluated graphically. Simple effects of treatment and entrainment status on FL were analysed using pairwise comparisons of means. The number of times that fish passed the pipe and the number of successful escape attempts were analysed using one-way ANOVAs with subsequent Tukey's post hoc tests for multiple comparisons of means. The proportion of fish entrained, EPP, number and duration of impingement events and IER were all analysed using individual Kruskal–Wallis rank sum tests and subsequent pairwise comparisons of means with Bonferroni-adjusted P-values to correct for multiple comparisons. The number and duration of impingement events as well as the IER were only compared among the three structural modifications, because the open pipe control was unable to cause impingement events. Passage over time was analysed using two-way ANOVAs, with ‘time’ and ‘treatment’ as factors, and subsequent Tukey's post hoc tests. Time was a categorical variable with three levels: 0–20 min into the trial, 20–40, and 40–60 min. Entrainment over time was analysed using a generalized linear mixed model with a Poisson distribution and an offset term equal to the logarithm of the total number of fish entrained. ‘Treatment’ and ‘time’ were fixed effects, while time within treatment within each experiment, experiment number within each treatment, and experiment number were all considered random effects. Significance was considered at α ≤ 0.05 for all comparisons.
Results
Fish size
A summary of fish sizes according to treatment and entrainment status is shown in Table 1. Fish mass was significantly different among treatments (F3,654 = 10.1; P = 1.7 × 10−6) and entrainment status (i.e. whether or not fish were entrained; F1,654 = 29.4; P = 8.5 × 10−8), and there was a significant interaction between treatment and entrainment status (F3,654 = 4.4; P = 0.004). Likewise, fish length (FL) was also significantly different among treatments (d.f. = 3; P = 0.01) and entrainment status (d.f. = 1; P = 1.1 × 10−9), and there was a significant interaction between the two (d.f. = 3; P = 1.9 × 10−7; Table 1). Overall, fish that were entrained through the diversion pipe were smaller in length and mass than those that were not.
Table 1:
The mass and fork length of fish that were or were not entrained through the diversion pipe for each treatment
| Entrainment status |
||||
|---|---|---|---|---|
| Mass (g) |
Fork length (cm) |
|||
| Treatment | Entrained | Not entrained | Entrained | Not entrained |
| Control | 115.4 ± 3.5a* | 126.9 ± 3.6a | 28.4 ± 0.3a | 29.2 ± 0.3 |
| Louvre box | 56.1 ± 2.5b* | 110.6 ± 4.2b | 22.7 ± 0.4b* | 28.9 ± 0.4 |
| Perforated cylinder | 112.0 ± 14.3ac | 119.7 ± 4.0ab | 28.3 ± 1.2ac | 29.1 ± 0.3 |
| Trash-rack box | 85.9 ± 4.6c* | 110.0 ± 4.1b | 26.1 ± 0.5c* | 28.7 ± 0.4 |
Lowercase letters indicate significant differences among treatments for each individual entrainment status (i.e. entrained or not entrained), whereas asterisks indicate significant differences (P < 0.05) between entrainment statuses for each treatment. Details of statistical analyses are given in the main text.
Total passages
The number of times that fish were observed to move past the pipe was not significantly different among treatments (F3,20 = 1.6, P = 0.2). During control trials, fish moved past the pipe 102.5 ± 14.6 times (mean ± SEM) during the hour-long experimental period. Likewise, fish moved past the pipe 91.3 ± 6.5 times during trash-rack box experiment, 121.7 ± 17.2 times during louvre box experiments, and 125.7 ± 10.2 times during perforated cylinder experiments.
Proportion of fish entrained
The effect of treatment on the proportion of fish that were entrained through the diversion pipe was significant (χ2 = 20.8, d.f. = 3, P = 0.0001; Fig. 2). The trash-rack box, louvre box and perforated cylinder all significantly reduced the proportion of fish that were entrained relative to the control (P = 0.028, P = 0.03 and P = 0.028, respectively). More than 40% of fish tested in control conditions were entrained through the diversion pipe (0.41 ± 0.06, mean proportion ± SEM). Of the three modifications, the trash-rack box entrained the highest proportion of fish compared with the louvre box and perforated cylinder (P = 0.029 and P = 0.027, respectively). The trash-rack box entrained 15% of the fish tested (0.15 ± 0.01, mean proportion ± SEM), while the louvre box and perforated cylinder reduced entrainment to only 5 and 2%, respectively (0.05 ± 0.01 and 0.02 ± 0.01, mean proportion ± SEM, respectively).
Figure 2:
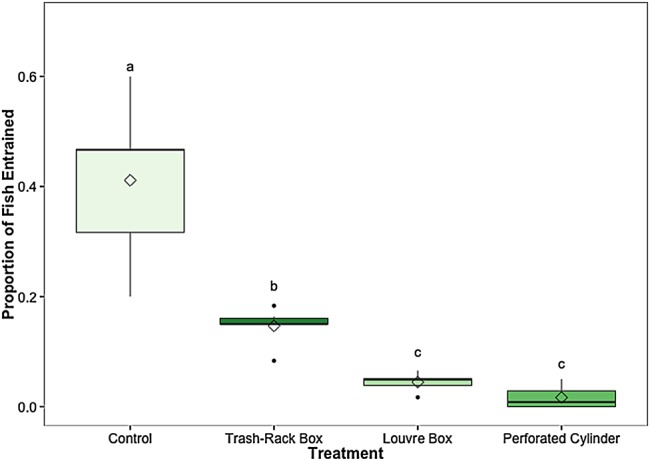
The proportion of fish that were entrained through the diversion pipe out of the total number of fish tested for each treatment. Different lowercase letters indicate statistically significant differences among treatments. Black line represents the median, box the interquartile range (IQR), whiskers = 1.5 IQR, filled circles represent outliers, and diamond indicates the mean. Mean proportions of fish diverted for each treatment (±SEM) are reported in the text. n = 6 trials for each treatment, 60 (±3) fish per trial.
Entrainment risk per pipe passage
The EPP was significantly different among treatments (χ2 = 20.6, d.f. = 3, P = 0.0001), with all three modifications significantly lowering the EPP compared with the control treatment (P = 0.03 for all comparisons to control; Fig. 3). The EPP was 0.25 ± 0.06 (mean ± SEM) for fish tested in control conditions, 0.10 ± 0.01 for the trash-rack box, 0.03 ± 0.01 for the louvre box and 0.01 ± 0.01 for the perforated cylinder. The EPP for the trash-rack box was significantly higher than that of the louvre box or perforated cylinder (P = 0.03 and P = 0.029, respectively).
Figure 3:
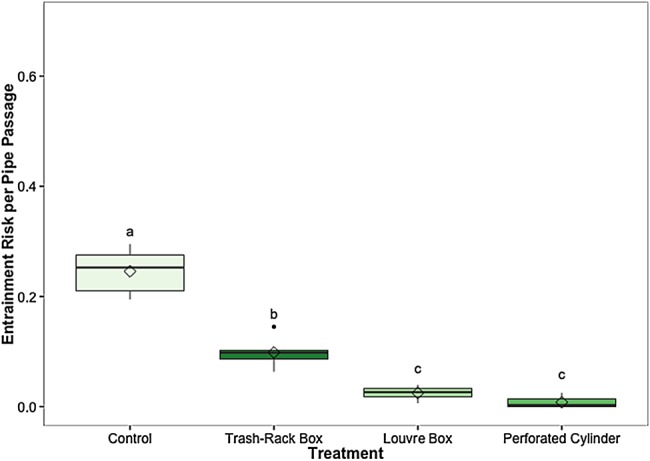
The entrainment risk per pipe passage for each treatment condition. This represents the chance of a fish becoming entrained into the diversion pipe after swimming past the pipe a single time. Different lowercase letters indicate statistically significant differences among treatments. Black line represents the median, box the interquartile range (IQR), whiskers = 1.5 IQR, filled circles represent outliers, and diamond indicates the mean. Mean proportions of fish diverted for each treatment (±SEM) are reported in the text. n = 6 trials for each treatment, 60 (±3) fish per trial.
Successful entrainment escapes
The number of times that fish were able to successfully avoid entrainment once an entrainment event began (escapes) was significantly different among treatments (F3,20 = 4.6, P = 0.01). Fish were able to escape entrainment from the trash-rack box and the louvre box a significantly greater number of times than they did from the control (6.3 ± 1.1 vs. 1.2 ± 0.3, mean ± SEM, P = 0.04; and 7.2 ± 1.6 vs. 1.2 ± 0.3, P = 0.01, respectively). There were no significant differences in successful escape attempts among the three modifications, and the number of escapes ranged from 5.2 to 7.2 for the modifications.
Impingement: number of events, escape rates and duration
Number of events
While there was no significant difference among treatments in the number of times that fish became impinged on the structural modifications, the effect approached significance (χ2 = 5.8, d.f. = 2, P = 0.054). Fish became impinged on the trash-rack box an intermediate number of times (4.7 ± 0.7 times, mean ± SEM), the louvre box most frequently (6.0 ± 1.0) and the perforated cylinder the lowest number of times (3.2 ± 0.4).
Impingement escape rate
The number of times that fish were able to escape impingement without becoming entrained (IER) was significantly different among treatments (χ2 = 8.2, d.f. = 2, P = 0.02). However, post hoc pairwise comparisons revealed no significant differences in the IERs of the trash-rack box (0.77 ± 0.01), louvre box (0.91 ± 0.06, mean ± SEM) or perforated cylinder (0.57 ± 0.12) treatments (all comparisons P > 0.05).
Impingement duration
The amount of time that fish spent impinged upon a structural modification differed significantly among treatments (χ2 = 9.1, d.f. = 2, P = 0.01; Fig. 4). The duration of impingement was significantly less for the trash-rack box compared with the louvre box (1.8 ± 0.2 vs. 13.4 ± 9.5 s; P = 0.025), but there were no differences in impingement time between the louvre box and perforated cylinder (13.4 ± 9.5 vs. 3.8 ± 1.7 s; P = 0.14) or between the trash-rack box and the perforated cylinder (1.7 ± 0.2 vs. 3.8 ± 1.7 s; P = 0.69).
Figure 4:
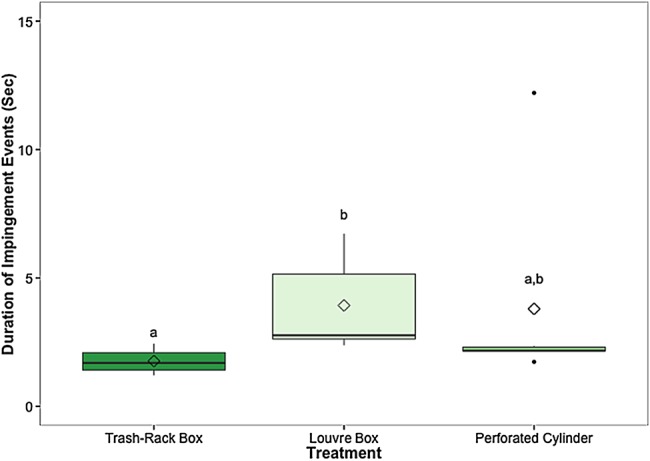
The mean duration of time each fish spent impinged upon the structure for each modification. The control treatment was not included in this comparison, because fish were unable to become impinged upon the pipe itself (with no modification). Different lowercase letters indicate statistically significant differences among treatments. Black line represents the median, box the interquartile range (IQR), whiskers = 1.5 IQR, filled circles represent outliers, and diamond indicates the mean. Mean proportions of fish diverted for each treatment (±SEM) are reported in the text. n = 6 trials for each treatment, 60 (±3) fish per trial.
Passage and entrainment over time
There was a significant effect of time on the number of times that fish moved past the diversion pipe (F3,60 = 13.8, P = 1.2 × 10−5). As experimental time increased, the number of passage events also increased (Fig. 5), with the greatest number of fish moving past the pipe from 40 to 60 min into the experiment. The number of pipe passages during the 0–20 min period of the experiment was significantly lower than those observed during the 20–40 and 40–60 min periods (P = 6.2 × 10−6 and P = 0.02, respectively). While there was no significant difference in the number of passages during the 20–40 and 40–60 min periods, the difference approached significance (P = 0.053). There was also no significant interaction between time and treatment on the number of pipe passages (F6,60 = 1.2, P = 0.3).
Figure 5:
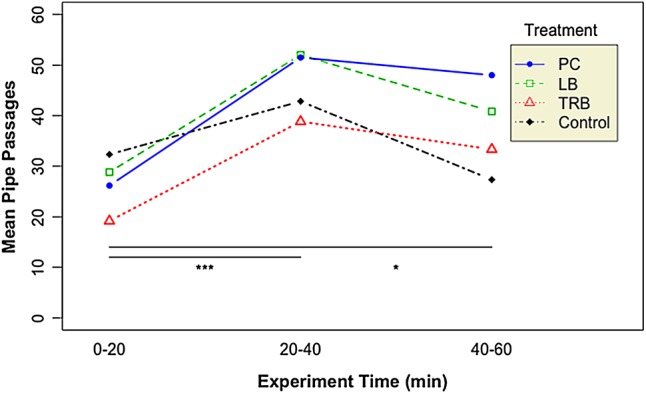
The number of times that green sturgeon swam past the pipe over the duration of the experimental trial. Asterisks indicate statistically significant differences between time periods during the experimental trial: *P < 0.05 and ***P < 0.001. Abbreviations: LB, louvre box; PC, perforated cylinder; and TRB, trash-rack box.
There was no significant effect of time on the number of entrainment events (χ2 = 4.9, P = 0.2), and the inclusion of ‘time’ as a variable did not significantly improve model fit. There was also no significant interaction between time and treatment on the number of fish that were entrained through the diversion (χ2 = 1.3, P = 0.3).
Discussion
Our results indicate that the tested fish-protection modifications of a water-diversion pipe can successfully reduce the entrainment risk of a threatened juvenile fish species while still maintaining water-diversion activities. Some of these devices (i.e. louvres and trash racks) have been used to reduce fish entrainment on large hydropower projects and pumping facilities (USBR, 2006; Kimmerer, 2008), but may also represent alternatives to fish screens on smaller water-diversion pipes, such as exclusion screens and sensory deterrents. Reconciliation between the needs of water diverters and native fish species is crucial for effective fish conservation and management, and our results suggest that such solutions are possible.
We have previously assessed the entrainment risk of slightly larger juvenile green sturgeon (mean FL = 39.4 vs. 28.5 cm) using a different set of structural modifications or sensory deterrents (Poletto et al., 2014b). In control conditions matching those tested in the present study, we showed that an average of 44% (±4.0% SEM) of fish tested within the flume were diverted through the pipe (Poletto et al., 2014b), which was very similar to the mean of 41% (±6.0% SEM) obtained in the present study. Likewise, the previously obtained EPP was 0.25 ± 0.03 (mean ± SEM) for the unmodified control pipe (Poletto et al., 2014b), which was the same as that obtained for the control conditions in the present study (0.25 ± 0.06, mean ± SEM). This entrainment risk is relatively high; an individual fish moving past the unmodified diversion pipe had a one in four chance of becoming entrained after swimming past the pipe a single time. In both investigations, the number of fish diverted through the unmodified diversion pipe, as well as the EPP, were consistent over two year classes, tested at relatively similar size classes, and this underscores the susceptibility of this species to entrainment in the wild. We do caution, however, that the entrainment rates observed within the experimental flume were gathered during the daytime in one set of hydraulic conditions (i.e. sweeping flow and diversion rate), and any extrapolation into field entrainment rates should be made qualitatively and not quantitatively. However, our data highlight the presumed entrainment risk that this species faces when encountering unmodified water diversions, especially as the sturgeon outmigrate through the watershed as juveniles. Further studies are needed to determine how frequently these fish may encounter active water diversions during their outmigration period in the environment.
In the hydraulic conditions tested in our experimental flume (15 cm/s river, or sweeping velocity; 0.53 m3/s diversion volume), all three modifications in the present study significantly reduced the proportion of fish that were entrained into the diversion pipe (Fig. 2) and significantly reduced the EPP (Fig. 3) relative to the unmodified control conditions. The proportion of fish that were entrained through the diversion pipe was reduced by as much as 95% with the addition of the perforated cylinder, and EPP was reduced as much as 96% with the same modification. Previous work also revealed a significant reduction (between 70 and 93% reduction) in the number of fish that were entrained though the pipe, and in the EPP (between 76 and 92% reduction) when structural modifications (a change in the orientation of the pipe intake or the addition of a pipe cap) were tested. In both investigations, the total number of times that fish swam past the pipe was consistent among experiments, and the opportunity for fish to be diverted through the pipe was statistically similar for each treatment. Therefore, the overall reduction in entrainment for the structural modifications was not a function of a reduced number of fish interacting with the devices relative to control. The consistency of our results between investigations and over several years suggests that quantifying juvenile green sturgeon behaviour in the presence of a water diversion is a tractable model for investigating the efficacy of several different fish-protection devices that may assist with future management actions. Furthermore, given the high entrainment rates observed in the laboratory in comparison with another fish species native to California (i.e. Chinook salmon; Mussen et al., 2013), conferring protection to green sturgeon may also provide protection to additional fish species with greater physiological resistance to entrainment and higher swimming capabilities.
Each fish-protection device tested in the present study is presumed to have reduced entrainment as a result of a combination of physical and hydraulic features, although it is plausible that the changes in the water velocity surrounding the protection devices predominately mediated the reduced observed entrainment rates. The diversion modifications introduced additional physical structure to the area surrounding the pipe and widened the area over which water was drawn into the pipe, altering flow velocities at the upstream end of the modification and changing water acceleration over the area of the device. For the trash-rack box, the widened inlet reduced flow velocities near the pipe by introducing additional structure parallel or nearly parallel to the sweeping flow within the flume (the widened inlet; Fig. 1b). The average intake velocities at the face of the trash rack approached 0.56 m/s, but rapidly increased within the trash-rack box as fish moved closer to the centre of the pipe inlet. At the centre of the pipe inlet within the trash-rack box, average water velocities approached those of the unmodified control pipe, 3.4 m/s. This rapid acceleration in water velocity over a short distance is likely to have overwhelmed the swimming abilities of juvenile green sturgeon (see review by Verhille et al., 2014), such that any fish entering the device between the vertical trash-rack bars was likely to be entrained. However, the relatively low water velocities near the face of the trash rack reduced the likelihood of sturgeon experiencing intake velocities as high as those of the unmodified control, and resulted in fewer entrainments.
Likewise, the louvre box reduced velocities around the pipe, particularly at the upstream portion of the louvre array and at the face of louvres. The position of the individual louvres and the angle of orientation relative to the flow used here have been recommended for optimal efficiency (Taft, 2000) and have been previously shown to guide fish suitably across a louvre array and downstream of a diversion (Bates and Vinsonhaler, 1957). Louvres modify the hydraulics surrounding a water diversion, and thus the behavioural response of passing fishes (Bates and Vinsonhaler, 1957; USBR, 2006). It is likely that this additional modification of water flow surrounding the diversion contributed to the reduction in entrainment seen with the louvre box. In previous experiments, a comparison of a vertical bar rack (similar to the trash rack used here) and a louvre array at guiding shortnose (A. brevirostrum) and pallid sturgeon (Scaphirhynchus albus) revealed a much lower guidance efficiency for the bar rack, which the authors attributed at least partly to the lack of velocity stimuli associated with the bar rack (Kynard and Horgan, 2001). Indeed, our louvre box was significantly more effective at reducing entrainment compared with the trash-rack box, though both were successful compared with the control. As with the trash-rack box, the intake velocity at the centre of the pipe inlet within the louvre box was similar to that of the unmodified control pipe, although the average intake velocity over the entire louvre array approached ∼0.64 m/s. The intake velocity changed across the louvre array such that the downstream portion of the array exhibited higher intake velocities, owing to the angle of projection into the main channel of the sweeping flow and, while more fish were observed to contact the louvre array at the downstream portion, the device was still successful at reducing entrainment.
As with the two other two pipe-modification treatments, the perforated cylinder modified water velocities around the water-diversion pipe, and resulted in the lowest proportion of entrainment and EPP of any of the devices tested. While the extended structure, widened inlet and intake surface area from the perforations is likely to have contributed to the modified water velocities around the device, it is likely that the spatial orientation of the perforated circles in the cylindrical pipe had the largest impact on reducing green sturgeon entrainment. Green sturgeon are largely benthic organisms (Moyle, 2002), and while some individuals used the entire water column within the flume, the majority of fish swam at or near the bottom of the flume. The intact portion of the perforated cylinder prevented the entrainment of individuals swimming directly underneath the diversion pipe, which was commonly observed in the control treatment. A similar phenomenon was observed in an earlier study of juvenile green sturgeon, in which a partial pipe plate that covered the bottom portion of the pipe intake significantly reduced entrainment (Poletto et al., 2014b). These results suggest that the intake orientation or placement within the water column is an important feature to consider when designing fish-protection devices for single species or types of species.
Changes in water velocity have been shown to be an important factor in mediating the behaviour and physiology of fishes, particularly during downstream migrations (Arnold, 1974; Heggenes and Traaen, 1987; Tiffan et al., 2009). Anadromous fishes, in particular, are responsive to changes in water velocity (i.e. acceleration or deceleration; Haro et al., 1998; Enders et al., 2012) and turbulence (Odeh et al., 2002), probably owing to their highly migratory behaviour and the importance of flow cues in selecting successful passage routes (Enders et al., 2009). Juvenile Pacific salmon smolts have been shown to avoid areas with rapid changes in water velocities (Kemp et al., 2005; Enders et al., 2009), and changes in water velocity have resulted in behavioural changes of juvenile green sturgeon in previous laboratory experiments (Poletto et al., 2014b), suggesting the importance of this cue in mediating swimming movements. Hydraulic conditions have also been shown to affect entrainment of fish species native to California into large water-pumping facilities (Kimmerer, 2008; Grimaldo et al., 2009), and modification of water flows has been suggested as a way in which to attenuate losses at these facilities (Grimaldo et al., 2009; Verhille et al., 2014). In the present study, the changes in water velocities surrounding the water diversion caused by the addition of the fish-protection devices is likely to have induced behavioural avoidance in green sturgeon approaching the pipe, which contributed to the reduced numbers of entrainment events.
In addition to modifying the behaviour of fishes, physiological responses to water velocity are important to consider as well, especially in the context of life-history strategies of anadromous fishes, such as green sturgeon. As evidenced for some anadromous salmonids (Flagg and Smith, 1982; Katzman and Cech, 2001), changes in swimming physiology and performance accompany the physiological transition from freshwater to saltwater tolerance for green sturgeon (Allen et al., 2006), that mediate successful outmigration. These changes include muscle fibre activity (Katzman and Cech, 2001) and changes in critical swimming velocities (Ucrit values), such that absolute Ucrit (i.e. swimming capability) decreases as size increases in green sturgeon preparing to migrate (Katzman and Cech, 2001; Allen et al., 2006). The green sturgeon tested in the present study (and the previous study) were within the size range of fish capable of migrating and entering saltwater, and thus they might have an even further reduced swimming capability compared with smaller juveniles of the same species. Indeed, in a laboratory study investigating the susceptibility of juvenile green sturgeon to contacting or becoming impinged on fish screens, larger sturgeon were more likely to contact screens or become impinged on them (J. Poletto, D. Cocherell, and N. Fangue, unpublished data).
The changes in water velocities surrounding the fish-protection devices may have reduced velocities such that green sturgeon were able to overcome them. For control fish swimming near the pipe, 3.4 m/s is nearly seven times faster than the Ucrit of fish of the size that were tested (∼0.50 m/s; Allen et al., 2006). At their faces, the trash-rack box and the louvre box reduced intake velocities (0.56 and 0.64 m/s, respectively) to values closer to the mean Ucrit values for fish of the size tested (∼0.4–0.6 m/s; Allen et al., 2006), which allowed fish to avoid entrainment more easily.
In addition to the changes in water velocities created by the protection devices, it is possible that these devices served as partial physical barriers for fish entrainment. For example, the trash-rack box introduced a vertical barrier to the pipe inlet through the use of vertical steel bars evenly spaced every 7.6 cm, and the louvre box consisted of vertical louvres every 2.5 cm. The perforated cylinder, while enlarging the opening of the pipe inlet, provided a barrier to fish entrainment via the steel space in between the evenly spaced circular holes. Given the average mass and length of the fish tested (mean FL 28.5 cm and mean mass 112.6 g) these openings provided an opportunity for fish to become impinged on the structures without being entrained. Given that no physical barrier existed around the pipe inlet for the unmodified control conditions, no impingement events were observed. Impingement events occurred, however, for each of the fish-protection devices, although in low numbers. While there was no difference among the devices for the number of impingements, there was a difference in the length of time that fish spent impinged upon the devices (Fig. 4), with the louvre box causing the longest impingements. Repeated contact or impingements on fish-protection devices may result in increased predation owing to heightened susceptibility (Office of Technology Assessment, 1995), elevated cortisol and stress hormones that may increase predation risk (Olla et al., 1992), or exhaustion following sustained attempts to escape impingement. Indeed, Kieffer et al. (2001) demonstrated that juvenile Atlantic sturgeon (Acipenser oxyrinchus) exhibited prolonged physiological stress following exhaustive exercise, which may potentially affect subsequent migratory movements or predatory escape attempts.
Additionally, the size of the fish differed both for the fish that were entrained among treatment types and for entrainment status (i.e. entrained or not) within treatments (Table 1). Generally, fish that were entrained were both shorter and lower in mass than those that were not, although mass was much more variable than length. Among treatments, the smallest fish were entrained through the trash-rack box and louvre box, which could indicate that vertical bars and vertical louvres, respectively, restricted the larger fish physically from becoming entrained. Another possibility is that these devices reduced the intake velocities to such an extent that only the smallest fish were unable to avoid entrainment. The dramatic differences in mass observed between those entrained or not through the trash-rack and louvre boxes might indicate differential health or condition factors that may have contributed to the observed differences. Alternatively, these larger sturgeon may possess increased burst-swimming capabilities over short distances, associated with increased white (anaerobic) muscle fibres (e.g. as in smolting coho salmon, Oncorhynchus kisutch; Katzman and Cech, 2001). This ‘remodelling’ of locomotory muscle has not been investigated in anadromous sturgeons.
The devices tested herein have the potential to reduce entrainment of passing fishes successfully, particularly for smaller, privately owned water-diversion pipes used mainly for irrigation. Several large-scale water-export projects operated by federal and state governments currently use louvre and trash-rack systems, notably diversions to the Delta-Mendota Canal and California Aqueduct in the Central Valley of California (USBR, 2006). These diversions export water from the Sacramento–San Joaquin River Delta, which is part of the native range of the threatened Southern DPS green sturgeon. Louvres have proved successful in reducing the entrainment of several fish species (see Table 6 in USBR, 2006), although their efficacy is dependent upon the hydraulic conditions preceding the louvre array and the species and life stage of the fish in question. In contrast, smaller diversions generally rely on screening or other methods to reduce entrainment, or largely remain unmodified (Herren and Kawasaki, 2001). Fish exclusion screens can often be cost prohibitive (McMichael et al., 2004; Moyle and Israel, 2005), require regular maintenance (USBR, 2006), and can be potentially injurious to certain species of fishes (Swanson et al., 2004, 2005; Young et al., 2010). Here, we suggest that behavioural fish-guidance devices, which reduce inflow velocities and distribute them across a greater area (e.g. the perforated cylinder), and that alternatives to commonly used methods (e.g. the perforated cylinder), can be modified for use on water-diversion pipes and should be investigated further and considered in future water-management discussions.
Overall, our results indicate that the entrainment risk of juvenile green sturgeon at active water-diversion pipes can be reduced through the use of fish-protection devices, and offer an empirical investigation of their efficacy. Conferring protection to green sturgeon may also provide protection to additional fish species affected by entrainment. We suggest that development of fish entrainment-reduction devices for small-scale water diversions should continue, including further development of variations on current fish-protection technologies. Furthermore, we urge that rigorous testing of such technologies should be conducted prior to implementation and that consideration of the behaviour and physiology of target species should be included to create the most effective designs.
Funding
This work was supported by the California Department of Fish and Wildlife Ecosystem Restoration Program (agreement # E0783004 to N.A.F. and J.J.C.) and the University of California Agricultural Experiment Station (grant no. 2098-H to N.A.F.). This research indirectly benefited from funding from the US Department of Interior's Anadromous Fish Screen Program and the National Science Foundation Graduate Fellowship Program (grant to J.B.P.).
Acknowledgements
We would like to thank the Yurok tribe for their involvement and generous help in acquiring the green sturgeon broodstock, and J. Van Eenennaam, P. Lutes and E. Hallen for spawning and assistance in rearing the fish. We also acknowledge the time and effort that went into completing this project, and thank the student and staff assistants, especially D. Jauregui, O. Patton, B. Williamson, T. Nguyen, F. LaLuz, N. Ho, N. Britton, M. Richmond, T. Agosta, M. Figueroa, D. Cheng, A. Fratzke, I. Chau and M. Zhang. For insightful suggestions and advice in improving the project design, we thank D. Meier, S. Thomas, M. Bowen, T. LaFromboise, S. Hatleberg, D. Lindsay and T. Rust.
References
- Allen PJ, Cech JJ. (2006) Age/size effects on juvenile green sturgeon, Acipenser medirostris, oxygen consumption, growth, and osmoregulation in saline environments . Environ Biol Fish 79: 211–229. [Google Scholar]
- Allen PJ, Hodge B, Werner I, Cech JJ., Jr (2006) Effects of ontogeny, season, and temperature on the swimming performance of juvenile green sturgeon (Acipenser medirostris) . Can J Fish Aquat Sci 63: 1360–1369. [Google Scholar]
- Amaral S, Black J, McMahon B, Dixon D. (2002) Evaluation of angled bar racks and louvers for guidance lake and shortnose sturgeon . Am Fish Soc Symp 28: 197–210. [Google Scholar]
- Arnold GP. (1974) Rheotropism in fishes . Biol Rev 49: 515–576. [DOI] [PubMed] [Google Scholar]
- Bates D, Maechler M, Bolker B, Walker S. (2014) lme4: linear mixed-effects models using Eigen and S4. R package version 1.1–7, http://CRAN.R-project.org/package=lme4. [Google Scholar]
- Bates DW, Vinsonhaler R. (1957) Use of louvers for guiding fish . Trans Am Fish Soc 86: 38–57. [Google Scholar]
- Baumgartner LJ, Reynoldson NK, Cameron L, Stanger JG. (2009) Effects of irrigation pumps on riverine fish . Fish Manag Ecol 16: 429–437. [Google Scholar]
- Bennett WA. (2005) Critical assessment of the delta smelt population in the San Francisco Estuary, California . San Franc Estuary Watershed Sci 3 http://escholarship.org/uc/item/0725n5vk. [Google Scholar]
- Boys CA, Robinson W, Baumgartner LJ, Rampano B, Lowry M. (2013) Influence of approach velocity and mesh size on the entrainment and contact of a lowland river fish assemblage at a screened irrigation pump . PLoS ONE 8: e67026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cloern JE, Jassby AD. (2012) Drivers of change in estuarine-coastal ecosystems: discoveries from four decades of study in San Francisco Bay . Rev Geophys 50: doi:10.1029/2012RG000397. [Google Scholar]
- Deng D-F, Koshio S, Yokoyama S, Bai SC, Shao Q, Cui Y, Hung SS. (2003) Effects of feeding rate on growth performance of white sturgeon (Acipenser transmontanus) larvae . Aquaculture 217: 589–598. [Google Scholar]
- Deslauriers D, Kieffer JD. (2011) The influence of flume length and group size on swimming performance in shortnose sturgeon Acipenser brevirostrum . J Fish Biol 79: 1146–1155. [DOI] [PubMed] [Google Scholar]
- Doroshov SI. (1985) The biology and culture of sturgeon . In Muir J, Roberts R, eds, Recent Advances in Aquaculture, Vol. 2 Croom Helm; , London, UK, pp 251–274. [Google Scholar]
- Drinkwater KF, Frank KT. (1994) Effects of river regulation and diversion on marine fish and invertebrates . Aquat Conserv Mar Freshw Ecosyst 4: 135–151. [Google Scholar]
- Enders EC, Gessel MH, Williams JG. (2009) Development of successful fish passage structures for downstream migrants requires knowledge of their behavioural response to accelerating flow . Can J Fish Aquat Sci 66: 2109–2117. [Google Scholar]
- Enders EC, Gessel MH, Anderson JJ, Williams JG. (2012) Effects of decelerating and accelerating flows on juvenile salmonid behavior . Trans Am Fish Soc 141: 357–364. [Google Scholar]
- Flagg TA, Smith LS. (1982) Changes in swimming behavior and stamina during smolting of coho salmon . In Salmon and Trout Migratory Behavior Symposium Proceedings University of Washington, Seattle, WA, USA, pp. 191–195. [Google Scholar]
- Fox J, Weisberg S. (2011) An {R} Companion to Applied Regression, Ed 2 Sage; , Thousand Oaks, CA, USA: http://socserv.socsci.mcmaster.ca/jfox/Books/Companion. [Google Scholar]
- Grimaldo LF, Sommer T, Van Ark N, Jones G, Holland E, Moyle PB, Herbold B, Smith P. (2009) Factors affecting fish entrainment into massive water diversions in a tidal freshwater estuary: can fish losses be managed? North Am J Fish Manag 29: 1253–1270. [Google Scholar]
- Haro A, Odeh M, Noreika J, Castro-Santos T. (1998) Effect of water acceleration on downstream migratory behavior and passage of Atlantic salmon smolts and juvenile American shad at surface bypasses . Trans Am Fish Soc 127: 118–127. [Google Scholar]
- Heggenes J, Traaen T. (1987) Downstream migration and critical water velocities in stream channels for four salmonid spicies . J Fish Biol 32: 717–727. [Google Scholar]
- Herren, Kawasaki SS. (2001) Inventory of water diversions in four geographic areas in California's Central Valley . Fish Bull 179: 343–355. [Google Scholar]
- Israel JA, Cordes JF, Blumberg MA, May B. (2004) Geographic patterns of genetic differentiation among collections of green sturgeon . North Am J Fish Manag 24: 922–931. [Google Scholar]
- Katzman S, Cech JJ., Jr (2001) Juvenile coho salmon locomotion and mosaic muscle are modified by 3′,3′,5′-tri-iodo-l-thyronine (T3) . J Exp Biol 204: 1711–1717. [DOI] [PubMed] [Google Scholar]
- Kemp PS, Gessel MH, Williams JG. (2005) Fine-scale behavioral responses of Pacific salmonid smolts as they encounter divergence and acceleration of flow . Trans Am Fish Soc 134: 390–398. [Google Scholar]
- Kieffer JD, Wakefield AM, Litvak MK. (2001) Juvenile sturgeon exhibit reduced physiological responses to exercise . J Exp Biol 204: 4281–4289. [DOI] [PubMed] [Google Scholar]
- Kimmerer WJ. (2008) Losses of Sacramento River Chinook salmon and delta smelt to entrainment in water diversions in the Sacramento-San Joaquin Delta . San Franc Estuary Watershed Sci 6 http://escholarship.org/uc/item/7v92h6fs. [Google Scholar]
- Kingsford RT. (2000) Ecological impacts of dams, water diversions and river management on floodplain wetlands in Australia . Austral Ecol 25: 109–127. [Google Scholar]
- Kynard B, Horgan M. (2001) Guidance of yearling shortnose and pallid sturgeon using vertical bar rack and louver arrays . North Am J Fish Manag 21: 561–570. [Google Scholar]
- Lee S, Wang Y, Hung SSO, Strathe AB, Fangue NA, Fadel JG. (2014) Development of optimum feeding rate model for white sturgeon (Acipenser transmontanus) . Aquaculture 433: 411–420. [Google Scholar]
- Liermann CR, Nilsson C, Robertson J, Ng RY. (2012) Implications of dam obstruction for global freshwater fish diversity . Bioscience 62: 539–548. [Google Scholar]
- McMichael GA, Vucelick JA, Abernethy CS, Neitzel DA. (2004) Comparing fish screen performance to physical design criteria . Fisheries 29: 10–16. [Google Scholar]
- Moyle PB. (2002) Inland Fishes of California. University of California Press; , Berkeley, CA, USA, 253 pp. [Google Scholar]
- Moyle PB, Israel JA. (2005) Untested assumptions . Fisheries 30: 20–28. [Google Scholar]
- Mussen TD, Cocherell D, Hockett Z, Ercan A, Bandeh H, Kavvas ML, Cech JJ, Fangue NA. (2013) Assessing Juvenile Chinook Salmon Behavior and Entrainment Risk near Unscreened Water Diversions: Large Flume Simulations . Trans Am Fish Soc 142: 130–142. [Google Scholar]
- Mussen TD, Cocherell D, Poletto JB, Reardon JS, Hockett Z, Ercan A, Bandeh H, Kavvas ML, Cech JJ, Fangue NA. (2014a) Unscreened Water-Diversion Pipes Pose an Entrainment Risk to the Threatened Green Sturgeon, Acipenser medirostris . PLoS ONE 9: e86321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mussen TD, Patton O, Cocherell D, Ercan A, Bandeh H, Kavvas ML, Cech JJ, Fangue NA, Post J. (2014b) Can behavioral fish-guidance devices protect juvenile Chinook salmon (Oncorhynchus tshawytscha) from entrainment into unscreened water-diversion pipes? Can J Fish Aquat Sci 71: 1209–1219. [Google Scholar]
- Mussen TD, Cocherell D, Patton O, Jauregui D, Eran A, Bandeh H, Meier D, Thomas S, Kavvas ML, Cech JJ, Jr, et al. (2015) Modified water diversion structures can behaviorally deter Juvenile Chinook Salmon from Entrainment . Trans Am Fish Soc. doi:10.1080/00028487.2015.1064476 [Google Scholar]
- Noatch MR, Suski CD. (2012) Non-physical barriers to deter fish movements . Environ Rev 20: 71–82. [Google Scholar]
- Nobriga ML, Matica Z, Hymanson ZP. (2004) Evaluating entrainment vulnerability to agricultural irrigation diversions: a comparison among open-water fishes . Am Fish Soc Symp 39: 281–295. [Google Scholar]
- Odeh M, Noreika J, Haro A, Maynard A, Castro-Santos T, Cada GF. (2002) Evaluation of the effects of turbulence on the behavior of migratory fish . Final Report 2002, Report to Bonneville Power Administration, http://tethys.pnnl.gov/sites/default/files/publications/Odeh_et_al._2002.pdf.
- Olla BL, Davis MW, Schreck CB. (1992) Notes: Comparison of predator avoidance capabilities with corticosteroid levels induced by stress in juvenile coho salmon . Trans Am Fish Soc 121: 544–547. [Google Scholar]
- Office of Technology Assessment (1995) Fish passage technologies: protection at hydropower facilities. OTAENV-641 . Office of Technology Assessment, US Government Printing Office; , Washington, DC, USA. [Google Scholar]
- Peake S, Beamish FW, McKinley RS, Scruton DA, Katopodis C. (1997) Relating swimming performance of lake sturgeon, Acipenser fulvescens, to fishway design . Can J Fish Aquat Sci 54: 1361–1366. [Google Scholar]
- Poletto JB, Cocherell DE, Ho N, Cech JJ, Klimley AP, Fangue NA, Jonsson B. (2014a) Juvenile green sturgeon (Acipenser medirostris) and white sturgeon (Acipenser transmontanus) behavior near water-diversion fish screens: experiments in a laboratory swimming flume . Can J Fish Aquat Sci 71: 1030–1038. [Google Scholar]
- Poletto JB, Cocherell DE, Mussen TD, Ercan A, Bandeh H, Levent Kavvas M, Cech JJ, Fangue NA. (2014b) Efficacy of a sensory deterrent and pipe modifications in decreasing entrainment of juvenile green sturgeon (Acipenser medirostris) at unscreened water diversions . Conserv Physiol 2: doi:10.1093/conphys/cou056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- R Core Team (2012) R: a Language and Environment for Statistical Computing. R Foundation for Statistical Computing; , Vienna, Austria: ISBN 3-900051-07-0, http://www.R-project.org/. [Google Scholar]
- Seesholtz AM, Manuel MJ, Van Eenennaam JP. (2015) First documented spawning and associated habitat conditions for green sturgeon in the Feather River, California . Environ Biol Fish 98: 905–912. [Google Scholar]
- Sheer MB, Steel EA. (2006) Lost watersheds: barriers, aquatic habitat connectivity, and salmon persistence in the Willamette and Lower Columbia River Basins . Trans Am Fish Soc 135: 1654–1669. [Google Scholar]
- Simpson WG, Ostrand KG. (2012) Effects of entrainment and bypass at screened irrigation canals on juvenile steelhead . Trans Am Fish Soc 141: 599–609. [Google Scholar]
- Swanson C, Young PS, Cech JJ. (2004) Swimming in two-vector flows: performance and behavior of juvenile Chinook salmon near a simulated screened water diversion . Trans Am Fish Soc 133: 265–278. [Google Scholar]
- Swanson C, Young PS, Cech JJ. (2005) Close encounters with a fish screen: integrating physiological and behavioral results to protect endangered species in exploited ecosystems . Trans Am Fish Soc 134: 1111–1123. [Google Scholar]
- Taft EP. (2000) Fish protection technologies: a status report . Environ Sci Policy 3: 349–359. [Google Scholar]
- Tiffan KF, Kock TJ, Haskell CA, Connor WP, Steinhorst RK. (2009) Water velocity, turbulence, and migration rate of subyearling fall chinook salmon in the free-flowing and impounded snake river . Trans Am Fish Soc 138: 373–384. [Google Scholar]
- USBR (2006) Fish protection at water diversions: a guide for planning and designing fish exclusion facilities . US Department of the Interior, Bureau of Reclamation; . http://www.engr.colostate.edu/_pierre/ce_old/classes/ce717/Fish%20Protection%20at%20Water%20Diversions.pdf. [Google Scholar]
- Van Eenennaam JP, Webb MAH, Deng X, Doroshov SI, Mayfield RB, Cech JJ, Jr, Hillemeier DC, Willson TE. (2001) Artificial spawning and larval rearing of Klamath River green sturgeon . Trans Am Fish Soc 130: 159–165. [Google Scholar]
- Van Eenennaam JP, Linares-Casenave J, Doroshov SI. (2012) Tank spawning of first generation domestic green sturgeon: tank spawning of green sturgeon . J Appl Ichthyol 28: 505–511. [Google Scholar]
- Verhille CE, Poletto JB, Cocherell DE, DeCourten B, Baird S, Cech JJ, Jr, Fangue NA. (2014) Larval green and white sturgeon swimming performance in relation to water-diversion flows . Conserv Physiol 2: doi:10.1093/conphys/cou031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wickham H. (2009) ggplot2: Elegant Graphics for Data Analysis. Springer; , New York, NY, USA. [Google Scholar]
- Wickham H. (2011) The split-apply-combine strategy for data analysis . J Stat Softw 40: 1–29. http://www.jstatsoft.org/v40/i01/. [Google Scholar]
- Young PS, Swanson C, Cech JJ. (2010) Close encounters with a fish screen III: behavior, performance, physiological stress responses, and recovery of adult delta smelt exposed to two-vector flows near a fish screen . Trans Am Fish Soc 139: 713–726. [Google Scholar]