

Abstract
Background
Age-related changes in nociception have been extensively studied in the past decades. However, it remains unclear whether in addition to the increased incidence of chronic illness, age-related changes in nociception contribute to increased prevalence of pain in the elderly. Although a great deal of evidence suggests that nociception thresholds increase with aging, other studies yield disparate results. The aim of this investigation was to longitudinally determine the effect of aging on nociception.
Methods
The authors developed a nociception assay for mice using electrical stimuli at 2,000, 250, and 5 Hz that reportedly stimulate Aβ, Aδ, and C sensory nerve fibers, respectively. A system was designed to automate a method that elicits and detects pain-avoiding behavior in mice. Using a Latin square design, the authors measured current vocalization thresholds serially over the course of mice’s life span.
Results
For 2,000-Hz (Aβ), 250-Hz (Aδ), and 5-Hz (C) electrical stimuli, current vocalization thresholds first decreases and then increases with aging following a U-shaped pattern (P < 0.001). In addition, average current vocalization thresholds at youth and senescence are significantly higher than those at middle age for the 250-Hz (Aδ) and 5-Hz (C fiber) electrical stimulus (P < 0.05).
Conclusions
Using a novel and noninjurious nociception assay, the authors showed that over the life span of mice, current vocalization threshold to electrical stimuli changes in a U-shaped pattern. The findings support the notion that age-related changes in nociception are curvilinear, and to properly study and treat pain, the age of subjects should be considered.
AGE-RELATED changes in nociception has been an area of significant clinical and scientific interest over the past decades.1–6 One reason for such interest is that in the growing elderly population, pain is often associated with chronic illnesses and is one of the most common reasons prompting the need for medical care. However, it is unclear whether age-related physiologic changes in nociception contribute to increased prevalence of pain in the elderly.1,6 Another reason to study age-related changes in nociception is that when pain occurs at younger ages, it can lead to long-term changes in pain perception, processing, and development.7–11 Therefore, a better understanding of age-related changes in nociception would enable the development and delivery of age-appropriate therapy.
Although the effects of aging are incompletely understood, there is evidence from human and animal studies suggesting that age-related changes in nociception have a curvilinear pattern. For example, population-based studies indicate that prevalence rates of chronic pain peak in midlife12,13 and that middle-aged individuals report larger number of pain locations, higher prevalence of fibromyalgia, and higher rates of undiagnosed causes of pain.14 In animals, age-related changes in sensitivity to longer-lasting and tonic pain peak during midlife.15 In a study evaluating pain behavior after formalin injection (a model of tissue injury and inflammation), rats displayed an age-related curvilinear pattern of sensitivity to tonic pain. Middle-aged rats showed the greatest sensitivity, whereas young and older animals displayed similar and lower sensitivity to the formalin test.15 Therefore, although the pattern of age-related changes in nociception is incompletely understood, there is growing evidence showing that it may be curvilinear.
Similarly, the effects of aging in each of the individual sensory nerve fibers—Aβ (pressure sensation), Aδ (well-localized sharp pain), and C (diffuse burning pain)—are also incompletely understood. Some investigations in humans suggest that aging might differentially affect sensory nerves. Researchers have suggested that in the elderly, there is impairment of Aδ16 –18 and Aβ fibers2 manifested by abnormalities in thermal and pressure nociception. Therefore, the development of a method that allows for longitudinal studies of the effects of aging on each sensory nerve fiber is needed and could contribute to the understanding of age-related changes in nociception and to the development of age-specific therapy.
In this study, we developed an assay to evaluate pain thresholds using electrical stimuli at 2,000, 250, and 5 Hz to preferentially stimulate Aβ, Aδ, and C sensory nerve fibers, respectively. In addition, contrary to most studies of nociception using methods that measure simple reflexive behavior in response to phasic nociceptive stimulation (tail-flick test, hot plate), we developed a technique that allows for the longitudinal study of complex pain-avoiding behavior in response to noxious stimulation. We defined vocalization as the pain-avoiding behavior to electrical stimuli and measured current vocalization threshold in 36 mice starting at the age of 10 weeks through their life span or 104 weeks.
Materials and Methods
We studied vocalization threshold to electrical stimuli in 36 female B6129SF2/J mice (Jackson Laboratory, Bar Harbor, ME) and longitudinally measured pain thresholds over the life span of each mouse or until 104 weeks of life. To study animals throughout the aging process, as recommended by gerontology researchers, we selected ages that represent youth, middle age, and senescence for this mouse strain.19,20 To eliminate operator variation, we developed this assay such that delivery, intensity, duration, and type of electrical stimulus as well as detection of pain-avoiding behavior were automated. Daily and before each experiment, animals were weighed and visually examined for signs of poor health. Mice were housed in ventilated cages in a temperature-controlled environment (21°C) and kept in a standard 12-h light– dark cycle, with food pellets (National Institutes of Health diet) and water available ad libitum. Mice were housed on Tek-Fresh bedding (Harlan Teklad, Madison, WI), with paper tubes, nestlets, and nutritional supplements (fresh fruit or bacon softies; BioServ, Frenchtown, NJ) for enrichment. All experiments were performed between 7:30 and 10:30 AM in a quiet room with only one animal present during measurements. Animals were acclimatized to the holder during three 10-min sessions 1–2 weeks before the initial study. The investigational protocol was approved by the Clinical Center Animal Care and Use Committee, National Institutes of Health, Bethesda, Maryland.
Nociception Assay
Figure 1 illustrates the components of the nociception assay. Custom hardware and software were designed to control and automate electrical stimulation frequency, intensity, duration, and duty cycle. The system automates a large portion of the protocol to facilitate the conduct of experiments, reduces user fatigue by sequencing the protocol, providing automated and manual detection of vocalizations, providing a handheld remote control and display (enables user to focus on mouse instead of laptop screen), and saving experiment data to a file for subsequent analysis. The application-specific software program controls all components of the system. The electrical stimulus is generated by a neurostimulator (Neurometer; Neurotron, Inc., Baltimore, MD), and is controlled through a standard RS-232 serial port. A custom-built handheld remote control and display device is connected through a custom cable to a digital I/O PCMCIA card (6533; National Instruments, Austin, TX). A microphone (AT943-SP; Sound Professionals, Mt. Laurel, NJ) is placed on a rubber mount in front of the mouse and is connected to a custom-built preamplifier circuit. The amplified audio signal is digitized with a data acquisition PCMCIA card (National Instruments). Animals were gently restrained in a mouse holder (Kent Scientific Corporation, Torrington, CT) such that the tail was accessible to the investigator. The mouse holder was slightly modified to minimize mouse chewing and scratching on hard surfaces, which can be a source of problematic audio noise. A grounding (SDE44; Neurotron Inc.) and a stimulating electrode (ATE1925; Neurotron Inc.) were attached to the tail approximately 1 cm apart.
Fig. 1.
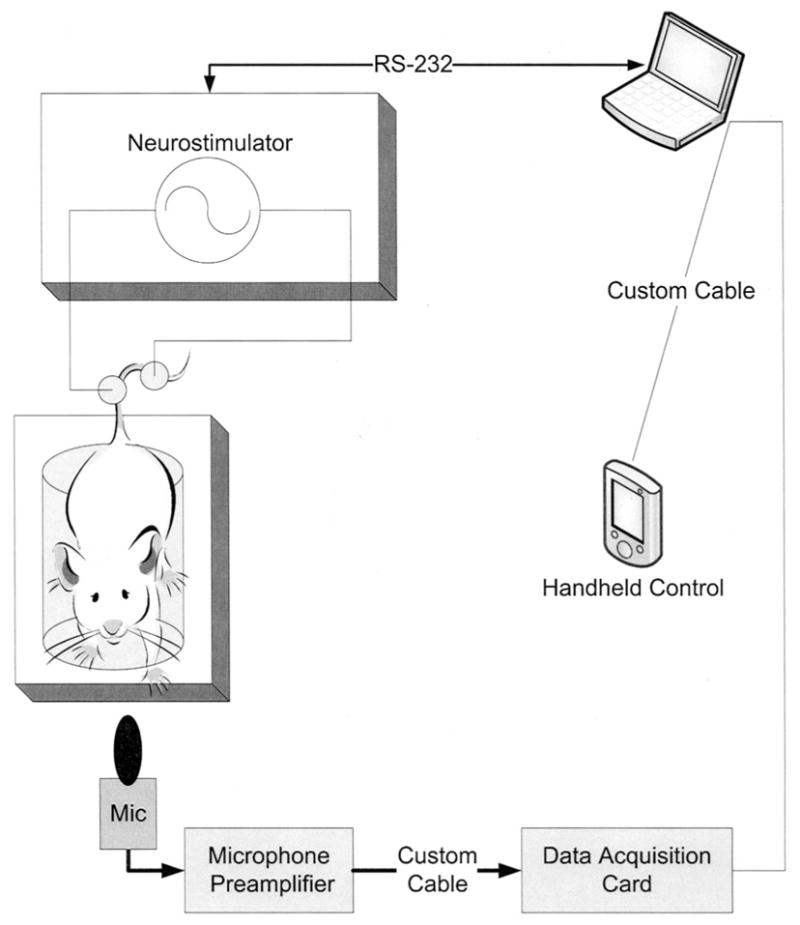
Components of our noninjurious nociception assay in mice used to study age-related changes in current vocalization threshold to electrical stimuli. The investigator can initiate and conduct the entire experiment through computer control. The neurostimulator used was the Neurometer (Neurotron, Inc., Baltimore, MD). Mic = microphone; RS-232 = standard serial communication port.
Current Vocalization Threshold
In this nociception assay, vocalization was the pain-avoiding behavior used to terminate delivery of electrical stimulus. The amperage of the electrical stimulus that elicits vocalization is defined as current vocalization threshold. For ease of presentation, we chose to define the unit of measurement of current vocalization threshold as the unit that corresponds to 100 times the intensity (amperage) that elicited vocalization (table 1). To develop a protocol (i.e., hardware and software) to detect mouse vocalizations, we conducted pilot studies to determine the characteristics of vocalization in response to the electrical stimulus. Because movements by the mouse during the experiment are picked up by the microphone, they must be distinguished from vocalizations. Typically, these signals are much lower in frequency and do not share the same characteristics as a vocalization. The software program accepts audio data from the microphone and performs filtering operations to remove background noise and spikes caused by scratching or chewing. After filtering, calculations are performed to find any audio segment that contains significant energy to be considered a possible vocalization. Energy segments of short duration (< 5 ms) are rejected as noise. Further processing is performed on the remaining segments to determine any periodicity present in the signal. If a segment is determined to be reasonably periodic and within a predefined repetition rate, that segment is classified as a vocalization, and the electrical stimulus is immediately terminated.
Table 1.
Intensities of Electrical Stimuli at Respective Frequencies Delivered to Determine Current Vocalization Threshold*
| Stimulus | Frequency and Intensities of Electrical Stimuli
|
||
|---|---|---|---|
| 2,000 Hz | 250 Hz | 5 Hz | |
| Current vocalization threshold units/corresponding mA | Current vocalization threshold units/corresponding mA | Current vocalization threshold units/corresponding mA | |
| Minimum | 40/0.4 | 14/0.14 | 5/0.05 |
| Maximum | 130/1.3 | 50/0.5 | 50/0.5 |
| Increment | 10/0.1 | 4/0.04 | 5/0.05 |
One current vocalization threshold unit equates to 100 mA.
Delivery of Electrical Stimulus
Electrical stimuli are delivered at three different frequencies, 2,000, 250, and 5 Hz, which are believed to preferentially stimulate Aβ, Aδ, and C fibers, respectively.21 For a given frequency, stimuli are delivered at predetermined intervals and in incremental intensities within ranges shown in table 1. In a given day, vocalization threshold for each frequency was measured five times, and for analyses of all factors except repetition, we used the average of these five measurements. For a given frequency, each stepwise intensity increase of electrical stimulus lasted 1 s, and there was a 1-s interval between each stimulus delivered (50% duty cycle). The electrical stimulus pulse that elicits vocalization could last less than 1 s if detection of vocalization occurs before completion of the normal pulse. There is a 1-min interval between deliveries of electrical stimuli of different frequencies.
The computer program reads in the desired experimental protocol, which is stored in a spreadsheet format (see experimental design). This file contains the permutation of electrical stimulus frequencies for each mouse for each day of the protocol. It also contains minimum and maximum amperages for each frequency, as well as the duration and pause timings for each stimulus. The program provides visual feedback to the user as to the electrical stimulus frequency, intensity, and repetition. The investigator can manually stop a stimulus (due to missed automatic vocalization detection), inform the program of false automatic vocalization detection, or abort the entire procedure if necessary. The handheld remote control and display device allows a single person to operate the system while still paying close attention to the mouse.
Study Design and Experimental Protocol
Table 2 shows the protocol and order of electrical stimulus delivery. Measurements were obtained during each age study period at intervals described in table 3 in awake and nonsedated animals. To evaluate whether mice develop habituation to electrical stimulus, during each age period indicated in table 3, we obtained repeated measurements of current vocalization thresholds for each frequency in six different sessions (every other day for 2 consecutive weeks). Further, to evaluate the impact of exposure to each electrical frequency on the vocalization threshold of the other two frequencies, we used a Latin square design where each animal was exposed to all possible permutations of electrical frequencies. Animals were studied in groups of six, and the assigned permutations were maintained throughout the study.
Table 2.
Latin Square Experimental Design with the Randomization of Stimuli to Be Delivered to Individual Mice during Each and All Age Study Periods*
| Mouse | Week 1 Sessions
|
Week 2 Sessions
|
||||
|---|---|---|---|---|---|---|
| Monday | Wednesday | Friday | Monday | Wednesday | Friday | |
| 1 | 2,000, 250, 5 Hz | 250, 5, 2,000 Hz | 5, 2,000, 250 Hz | 250, 2,000, 5 Hz | 5, 250, 2,000 Hz | 2,000, 5, 250 Hz |
| 2 | 250, 5, 2,000 Hz | 5, 2,000, 250 Hz | 5, 250, 2,000 Hz | 2,000, 5, 250 Hz | 250, 2,000, 5 Hz | 2,000, 250, 5 Hz |
| 3 | 5, 2,000, 250 Hz | 2,000, 250, 5 Hz | 2,000, 5, 250 Hz | 5, 250, 2,000 Hz | 250, 5, 2,000 Hz | 250, 2,000, 5 Hz |
| 4 | 250, 2,000, 5 Hz | 5, 250, 2,000 Hz | 2,000, 250, 5 Hz | 250, 5, 2,000 Hz | 2,000, 5, 250 Hz | 5, 2,000, 250 Hz |
| 5 | 5, 250, 2,000 Hz | 2,000, 5, 250 Hz | 250, 2,000, 5 Hz | 5, 2,000, 250 Hz | 2,000, 250, 5 Hz | 250, 5, 2,000 Hz |
| 6 | 2,000, 5, 250 Hz | 250, 2,000, 5 Hz | 250, 5, 2,000 Hz | 2,000, 250, 5 Hz | 5, 2,000, 250 Hz | 5, 250, 2,000 Hz |
Thirty-six animals were enrolled in this study and followed for the duration of their lives or until 104 weeks. Animals were studied in groups of six (a total of six groups of six animals). The assignment of their random order of electrical stimulus was maintained for each and all age study periods. During each age study period (table 3), measurements of current vocalization threshold were obtained over 2 weeks.
Table 3.
Number of Animals, Age, and Weight for Each Age Study Period*
| Age Study Period | Number of Alive Animals | Age, Median (Range), weeks | Weight, Mean ± SEM, g |
|---|---|---|---|
| 1 | 36 | 14 (10–16) | 23 ± 0.1 |
| 2 | 34 | 44 (40–46) | 35 ± 0.2 |
| 3 | 32 | 64 (64–68) | 39 ± 0.2 |
| 4 | 30 | 84 (84–86) | 40 ± 0.2 |
| 5 | 21 | 103 (101–103) | 40 ± 0.4 |
Overall, during this investigation 15 animals died of natural death as indicated by the decrease in the number of animals in respective age study periods.
Statistical Methods
Data were analyzed using SAS Version 8.2 (SAS Institute Inc., Cary, NC). Each frequency vocalization threshold (2,000, 250, and 5 Hz) was analyzed separately using repeated-measures analysis of variance (ANOVA) with the procedure for generalized linear models; the Wilks lambda multivariate test for within-subjects effects was used. The experimental design (Latin square) allowed us to control for sequence of electrical stimuli (six permutations of the order of delivery of the three frequencies) and session (six sessions, Monday, Wednesday, and Friday for 2 weeks) for each of the five age periods studied. Separately, at each of the five age periods studied, we also performed a two-way analysis using the means of five repetitions, to examine the effect of sequence and session. To determine the effect of sequence and session at each period, the Tukey honestly significant difference was used for post hoc testing. Using repeated-measures ANOVA, we also evaluated the effect of repetition, because each frequency at each session was repeated five times. We used contrasts for post hoc testing to compare the five repetitions among each other (10 pairwise comparisons); P values were adjusted using the Holm adjustment for multiple comparisons. We used repeated-measures ANOVA to determine whether there was a difference among the five age periods studied. Contrasts among all five periods (10 pairwise comparisons) were used for post hoc testing, and these P values were adjusted using the Holm method. P values less than 0.05 were considered significant.
Results
Throughout the study, animals showed no signs of distress nor incurred any injury from the electrical stimuli. Twenty-one of 36 mice survived for the entirety of the study, and 15 died of natural deaths. Previous pilot studies showed that a mouse’s vocalization in response to electrical stimulus can present with a range of frequencies (2–15 kHz) and amplitudes. Some vocalizations are sine waves of a single frequency, others resemble a chirp (the frequency slowly increases/decreases during the duration of the vocalization), and still others are more complex and do not fit simple descriptions, although almost all contain a strong sinusoidal component. Over time, there were no noticeable differences in animals’ audible vocalizations in response to electrical stimulus.
Throughout the duration of the study, no audible vocalizations were heard or recorded while animals were in the restraint device or their cages, except when electrical stimuli were delivered. In addition, the false trigger rate (inaccurate detection of vocalization by the software when it did not occur) was less than 2% of measurements and was usually elicited by environmental noise (refrigerators, knocks on doors, centrifuges), and that of missed vocalizations (when the software did not detect a vocalization that did occur) was less than 1%. During experiments, we observed that in response to electrical stimuli of intensities lower than that which elicited vocalization (current vocalization threshold), animals displayed variable random tail movements suggesting that perception of stimulus preceded vocalization.
Effect of Aging on Current Vocalization Thresholds
Table 3 outlines the number of animals and ages at each age study period. Repeated-measures ANOVA indicated that mean current vocalization thresholds in response to 2,000-, 250-, and 5-Hz electrical stimulus changes over time in a U-shaped pattern (all P ≤ 0.001; fig. 2). Post hoc analysis with the Holm correction for multiple comparisons indicated that for 2,000 Hz, the mean current vocalization threshold was similar at younger and older ages (14 and 103 weeks, respectively; P = not significant), and that at younger age (14 weeks) was higher than that at middle ages (P < 0.05). For 250 Hz, mean current vocalization thresholds at younger and older ages were higher than those of middle ages (44, 64, and 84 weeks; P < 0.04). Finally, for 5 Hz, the mean current vocalization threshold was similar at younger and older ages (14 and 103 weeks respectively; P = not significant), and both were higher than those at middle ages (44, 64, and 84 weeks; all P < 0.04).
Fig. 2.
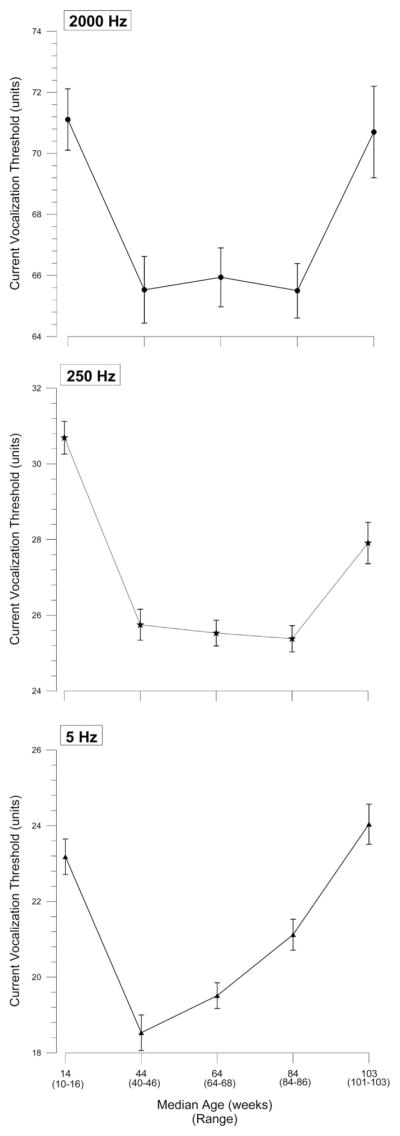
Mean ± SEM current vocalization thresholds for 2,000-, 250-, and 5-Hz electrical stimuli for 21 mice that lived through the entire duration of the experiment. Over time, for all three electrical frequencies, 2,000, 250, and 5 Hz, and in turn, preferentially for Aβ, Aδ, and C sensory nerve fibers, average current vocalization threshold to electrical stimulus changes in an U-shaped pattern, i.e., it initially decreases and then increases (all P < 0.001). Average current vocalization threshold at young and old ages (14 and 103 weeks) are higher than those at middle ages for 250- and 5-Hz frequencies (all P < 0.05).
Effect of Order of Delivery of Electrical Stimulus Frequencies on Current Vocalization Thresholds
To evaluate the impact of electrical stimulation with a given frequency on current vocalization threshold for other frequencies, we used a Latin square design where animals were exposed to all possible permutations of electrical stimulus. Repeated-measures ANOVA indicated that in the first study period (14 weeks), vocalization thresholds varied with the sequence in which electrical stimulus (2,000, 250, and 5 Hz) was delivered (P < 0.008). Post hoc analysis with the Tukey honestly significant difference test showed that mean vocalization thresholds for 2,000- and 250-Hz frequencies were lower when they were obtained after compared with when they were obtained before the 5-Hz electrical stimulus (P < 0.05). There was no impact of the sequence of delivery of any electrical stimuli on the vocalization threshold for the 5-Hz frequency (table 4; P = not significant).
Table 4.
Mean ± SEM Vocalization Thresholds for Each Frequency of Electrical Stimulus According to the Sequence of Stimulus Delivered during the First Age Study Period*
| Sequence of Frequency Delivered | Frequency of Electrical Stimulus
|
||
|---|---|---|---|
| 2,000 Hz | 250 Hz | 5 Hz | |
| 2,000, 250, 5 Hz | 74 ± 0.9 | 32 ± 0.3 | 22 ± 0.4 |
| 2,000, 5, 250 Hz | 72 ± 0.9 | 30 ± 0.4 | 23 ± 0.5 |
| 250, 2,000, 5 Hz | 70 ± 0.9 | 29 ± 0.4 | 21 ± 0.4 |
| 250, 5, 2,000 Hz | 69 ± 1.0 | 31 ± 0.4 | 23 ± 0.4 |
| 5, 2,000, 250 Hz | 62 ± 1.0 | 28 ± 0.4 | 23 ± 0.7 |
| 5, 250, 2,000 Hz | 65 ± 1.0 | 28 ± 0.4 | 24 ± 0.5 |
During the first age study period (mean age, 14 weeks; range, 10–16 weeks), vocalization threshold varied according to the sequence in which each stimulus (2,000, 250, and 5 Hz) was delivered (P < 0.008). For 2,000 and 250 Hz, vocalization thresholds were lower when they were obtained after compared with when they were obtained before the 5-Hz electrical stimulus (P < 0.05). There was no impact of the sequence of delivery of electrical stimuli for the 5-Hz frequency (P = not significant).
Effect of Repetitions of Current Vocalization Threshold to Electrical Stimulus Obtained within 1 Day
Repeated-measures ANOVA indicated that there were differences among the five measurements that yielded the mean current vocalization threshold for 2,000 and 250 Hz (P < 0.0001 for both) and 5 Hz (P < 0.01) over all age periods. We were primarily interested in comparing the first repetition to subsequent measurements, although we adjusted P values for all 10 pairwise comparisons. Post hoc analysis using the Holm adjusted P values for multiple comparisons indicated that for 2,000-and 250-Hz frequencies, the first vocalization threshold was higher than each of the four subsequent measurements (all P = 0.001; fig. 3). For the 5-Hz frequency, the first repetition was significantly different from the third, fourth, and fifth repetitions (all P < 0.05) but not the second (P = 0.17) repetition.
Fig. 3.
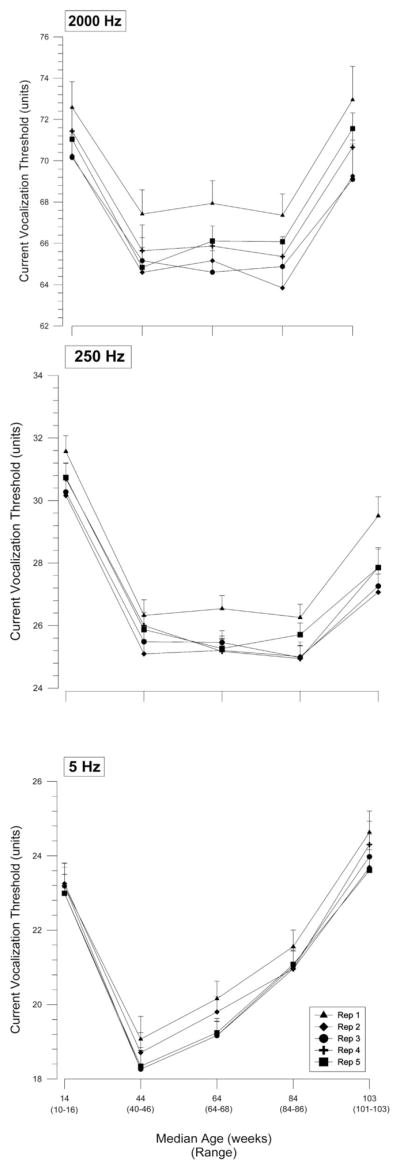
Mean ± SEM of each of five repeated measurements of current vocalization threshold to electrical stimulus for 2,000, 250, and 5 Hz (preferentially Aβ, Aδ, and C fibers) in animals that survived the entire experiment. Over the duration of the experiment, for all three frequencies, the mean first vocalization threshold was higher than the mean of some or all four subsequent measurements obtained in a given day (all P < 0.05). Rep = repetition.
Effect of Repeated Current Vocalization Threshold Measurements Obtained in Different Days over 2 Weeks at Each Age Study Period
To investigate whether animals develop habituation to electrical stimuli, current vocalization threshold for all frequencies was determined every other day (session) for 6 days, in random order, over a 2-week interval for each age study period as described in table 2. We evaluated whether there was a difference among mean current vocalization threshold for each frequency obtained on 6 different days (sessions) during a given age study period. At any and all study periods, for 2,000-, 250-, and 5-Hz frequencies, there were no differences among mean vocalization threshold obtained on different days (all P = not significant; data not shown).
Discussion
In this study, we found that in female mice, current vocalization threshold to electrical stimulation changes with aging in a U-shaped pattern. This U-shaped pattern of response was observed with electrical stimulation at 2,000, 250, and 5 Hz, which are believed to preferentially stimulate Aβ, Aδ, and C sensory nerve fibers, respectively. Specifically, our findings indicate that in midlife compared with youth and senescence, pain-avoiding behavior is elicited with lower intensity electrical stimuli (fig. 2), suggesting that middle-aged mice have lower tolerance to noxious stimuli. The fact that random variable tail movements occur with electrical stimuli of intensities lower than that which elicits vocalization might suggest that perception of stimulus precedes vocalization and that vocalization reflects pain tolerance and not perception threshold to electrical stimulation. We conclude that over the life span of mice, there are age-related changes in nociception that impact on the animals’ display of pain-avoiding behavior in a U-shaped pattern.
The impact of aging on nociception has been investigated in human and animal studies with various noxious stimuli.5,22 Although the majority of nociception studies using various methods suggest that sensitivity to noxious stimulation might decrease with aging in both humans and animals,4,22,23 other investigations show disparate results.24,25 For example, in humans, somatosensory thresholds for warmth, cold, and vibration were shown to increase, pressure pain threshold to decrease, and heat pain threshold to remain unchanged with aging.4 Studies in rats have shown that aging is associated with unchanged thermal reaction latency (tail immersion in 48°C water) and decreased mechanical pressure thresholds (paw pressure/struggle threshold, von Frey test).24 –26 In rats, adults (125 days old) had longer withdrawal latencies to conductive thermal stimuli at 40°–45°C, but not 35°C or 50°C, compared with very young pups (5 and 10 days old).27 In other studies, older rats (> 22 months) had decreased nociceptive response to high-intensity heat26 and increased pressure pain threshold compared with younger (3 months) animals.28 In rats, vocalization or jump threshold to nonneurospecific electrical stimulation decreased with aging.29,30 Researchers have suggested that these inconsistent results could be explained by variability of experimental designs, endpoints, and methodologies used (intensity and duration of noxious stimulus) and differences in strains and age groups studied.4,5,22,31 Nevertheless, these conflicting results suggest that age-related changes in nociception are incompletely understood.
Although several studies suggest that aging impacts nociception in a linear pattern, there is growing evidence indicating that age-related changes in pain behavior are nonlinear. For example, data derived from population-based studies suggest that prevalence rates for chronic pain peaks in midlife.12–14 In animals, age-related changes in sensitivity to longer-lasting and tonic pain were shown to have a curvilinear pattern and to peak in midlife in rodent models of tissue injury and inflammation.15 Along with studies of behavior response to pain, investigations of age-related anatomical and physiologic changes of the peripheral and central nervous systems of humans and animals suggest that age-related changes in nociceptive fibers are curvilinear throughout life.5,32,33 In murine peripheral nervous system, myelin thickness increases until 12 months and decreases between 12 and 22 months, nerve degeneration appears between 12 and 20 months, and a general disorganization and marked nerve fiber loss become evident beyond 20 months.33 Another study shows that in mice, nerve conduction velocity increases from 2 to 6 months, remains unchanged during adulthood, and subsequently decreases with aging, following an inverted U–shaped pattern.32,34 The current investigation shows that in response to electrical stimulation, of possibly both myelinated (Aβ, Aδ) and unmyelinated (C) fibers, mice display an age-related U-shaped pattern of response that closely mirrors age-related anatomical and neurophysiological changes in their peripheral nervous system. Although it is unclear whether a mechanistic relation between changes we observed in current vocalization threshold and physiologic age-related changes in myelinization and nerve conduction velocity in mice exists, our study adds to the body of literature suggesting that age-related changes in nociception are curvilinear.
Most studies of age-related changes in nociception and pain threshold use phasic stimulus in the form of thermal and mechanical stimulation in rats.5,22,35 Although traditional methods have proven valuable for the study of pain, some have limitations for longitudinal studies with repeated measurements. One such limitation is development of habituation to the stimulus. In a longitudinal study of the effect of caloric restriction in nociception in mice (42–100 weeks), latency response times in the 50°C hot plate changed with age. In that study, latency times decreased sharply over a few weeks and remained lower for several weeks, a phenomenon thought by the authors to represent habituation learning.36 Another limitation of currently used methods is the nonspecific nature of the stimulus. For example, with thermal stimuli (hot plate test or tail flick), temperatures greater than 45°C stimulate both Aδ and C fibers.37 Cold thermal testing is not standardized and is nonspecific at temperatures lower than 10°C.38 Further, the response to thermal and mechanical pressure stimuli can be altered by the animals’ skin color, thickness, and body temperature.27,39,40 Therefore, a method without these limitations would greatly add to available tools for the study of nociception.
To study age-related changes in nociception, we developed an assay that enables longitudinal investigations of complex pain-avoiding behavior. Unlike methods that measure simple reflexive responses to noxious stimuli to phasic nociceptive stimulation, we developed a behavior test where vocalization is the endpoint to terminate the noxious stimulus. As shown by others, audible vocalization is a behavior known to occur in response to electrical stimulus in rodents and to be a reliable supraspinal complex response to noxious stimulation.39,41 In addition, we were able to eliminate variability of operator interpretations of animal response to stimulus, standardize measurements, and automate the entire method. Another potential advantage of our method for longitudinal animal studies is that the electrical sine-wave stimuli delivered to the skin has constant current output and therefore directly stimulates nerve fibers independent of physical factors at the site of stimulation.39,42 That way, changes in skin thickness, temperature, and water content, which might occur with the aging process and might affect transduction of noxious stimuli, are unlikely to impact on the animal’s response to electrical stimuli.39,42 Therefore, this assay may be a useful tool for studies of the physiology and pharmacology of nociception because it does not produce injury or habituation, controls for changes that might impact on function of peripheral nociceptors, and yet elicits complex response to noxious stimuli.
Neurospecificity of the electrical stimulus at the frequencies used in this investigation is a matter of debate. To deliver electrical stimuli at 5, 250, and 2,000 Hz, we used a device that has been used for in vivo and in vitro studies, and in clinical settings for diagnosis of neuropathies43– 45 and investigations of pharmacodynamics of topical analgesics46 and of mechanisms of nociception.47,48 Although a matter of controversy, there is in vivo and in vitro evidence supporting the notion that electrical stimuli delivered at 2,000, 250, and 5 Hz preferentially activate Aβ, Aδ, and C fibers, respectively.21 Using action potential intracellular recordings from isolated dorsal root ganglia neurons with attached dorsal roots, researchers showed that C fibers were activated by 5-Hz and not by 250- or 2,000-Hz sine-wave stimuli.21 In addition, 2,000-Hz stimulation at low intensity selectively activated Aβ, and at significantly higher intensity, it stimulated Aδ neurons.21 Lastly, 250-Hz stimulation was shown to stimulate Aδ and Aβ but not C fibers.21 In that same study, in vivo patch clamp recordings showed that in response to cutaneous stimulation at 250 and 5 Hz, large-amplitude excitatory postsynaptic currents in substantia gelatinosa neurons (known to receive synaptic input from Aδ and C fibers) were observed. This finding suggests that 250-Hz and high-intensity 2,000-Hz electrical stimuli activate Aδ-fiber neurons, and 5-Hz stimulus specifically activates C-fiber neurons.21 Our finding that vocalization with 2,000-Hz stimulation (not necessarily a noxious stimulus) occurred only at much higher amperage than with 250 and 5 Hz might suggests that pain-avoiding behavior resulted from stimulation of Aδ fibers, which is in agreement with those in vitro findings. Other studies in rats showing that after intrathecal morphine animals had increased threshold only to the 5-Hz electrical stimulus applied to the hind paw also support the concept that 5-Hz stimulus activates C fibers.49 Another study showed that in rats receiving 2% intrathecal lidocaine, the time for return to baseline values of cutaneous sensory threshold was longer for 5-Hz (C fiber) and 250-Hz (Aδ fiber) than for 2,000-Hz (Aβ) stimulation.50 Such pattern of recovery from a nerve block with local anesthetics is similar to that observed in humans undergoing spinal anesthesia—small-diameter fibers (C and Aδ fibers) recover later than large-diameter fibers (Aβ). Therefore, although neurospecificity of the electrical stimuli used in this investigation is incompletely established and still a matter of controversy, there is evidence to suggest that 2,000, 250, and 5 Hz preferentially activate Aβ, Aδ, and C sensory nerve fibers, respectively. With this investigation we showed that our assay, with its noninjurious and automated properties, adds to the armamentarium of tools to study nociception, possibly allowing for serial measurements of specific sensory fiber current vocalization thresholds.
We evaluated whether animals develop habituation to electrical stimuli and whether stimulation at a given frequency impacts on current vocalization threshold of the other frequencies used. With regard to habituation to the electrical stimulus, we found that the first vocalization threshold was higher than all or some of the subsequent four measurements for 2,000-, 250-, and 5-Hz stimulations. However, during each age study period, we measured vocalization threshold every other day within a period of 2 weeks and observed no significant changes on current vocalization threshold from one day to the next. The fact that variations within daily measurements were not observed from one day to the next and that the changes over all age study periods were nonlinear suggest that even if there is some habituation learning, it is a very short-lasting phenomenon. Unlike what is reported with thermal and mechanical stimulus,25,36 mice are unlikely to develop habituation to electrical stimulation, and this supports the suitability of this assay for serial measurements of complex pain-avoiding behavior elicited by noxious stimuli.
With regard to the question of whether stimulation with a given frequency impacts on vocalization threshold to other frequencies, we found that current vocalization threshold to 5-Hz stimulation (C fiber) was not affected by sequence of electrical stimuli. Conversely, at the youngest age, current vocalization thresholds for 2,000 Hz (preferentially Aβ) or 250 (Aδ) were lower when 5 Hz (preferentially C fiber) preceded compared with when it followed 2,000- or 250-Hz stimulations. One possible explanation for this finding is that repeated stimulation with 5-Hz frequency leads to windup phenomenon, consequent heightened sensitivity, and in turn lower current vocalization threshold to 2,000-Hz (Aβ) and 250-Hz (Aδ fibers) stimuli.51–55 That this phenomenon was observed only in young mice could possibly be due to morphologic changes that occur with aging, such as a relative reduction in C and Aδ fibers16,32,56 or reduction of the N-methyl-D-aspartate receptor, which is known to be involved in windup and central sensitization and to be reduced in senescence.53,54,57,58 However, our findings are in conflict with those in rats indicating that windup phenomenon seen with C-fiber activation on the spinal nociceptive pathways on the flexor reflexes is more slowly attenuated in aged than in adult rats.59 Although the effect of aging in windup phenomenon was not tested in our study, these discrepant results may be explained by the differences in species and methodologies used. However, it is conceivable that our nociception assay could be used for research studies of events leading to windup and central sensitization.
Despite several potential advantages of this novel method, our findings should be interpreted with caution because our study has limitations. One such limitation is the lack of cross-sectional controls for each age group. Given our longitudinal study design, animals were studied repeatedly throughout the aging process. One could postulate that the changes observed are confounded by repeated experience of pain throughout life and not entirely reflective of effects of aging in nociception. Although this possibility cannot be ruled out, one would expect that linear, instead of curvilinear, changes in current vocalization threshold would result from repeated exposure to electrical stimuli. The fact that we observed a U-shaped instead of linear pattern of change in vocalization thresholds possibly suggests that repeated exposure to electrical stimulation is an unlikely reason for the changes observed. Further, the fact that there were no differences in current vocalization threshold from one day to the next within each of the 2 weeks of every-other-day testing conducted during each age period evaluation might suggest that repeated exposure to stimuli insignificantly contributes to our long-term findings. In addition, our finding of a U-shaped pattern for vocalization threshold in response to electrical stimulation over the life span of mice is in agreement with that of others measuring complex behavior responses to tonic pain.15 This congruence in results suggests that our findings are likely explained by age-related changes in nociception. Another limitation of our study is that we did not include male mice. One might argue that variability resulting from uncontrolled estrous phase and status among female mice could have contributed to the results. Although this possibility cannot be eliminated, the fact that there were no significant differences in daily vocalization thresholds within each of the 2-week evaluations at each age study period (when female mice could have gone through multiple estrous cycles) suggests that the impact of estrous cyclicity was likely insignificant. In a recent meta-analysis of studies of nociception, researchers showed that estrous cyclicity in female rodent adds no significant variability to data obtained from male counterparts, a result that we believe supports the validity of our findings and the use of female mice.60
In summary, we developed a novel nociception assay and showed age-related changes in current vocalization threshold to electrical stimuli believed to preferentially stimulate Aβ, Aδ, and C sensory nerve fibers. We showed that over the life span of mice, current vocalization threshold to electrical stimulus has a U-shaped pattern that is possibly age related, and middle-aged mice have higher sensitivity to noxious stimulus than young and senescent mice. Therefore, our findings add to the body of literature indicating that there are age-related changes in nociception, that these changes are curvilinear, and that in the study and treatment of pain, age of subjects should be considered.
Acknowledgments
Supported by the Intramural Research Program of the National Institutes of Health, National Institutes of Health Clinical Center and Center for Information Technology, National Institutes of Health, Bethesda, Maryland. The National Institutes of Health holds a biological materials license with Neurotron, Baltimore, Maryland, for use of the assay described in the article. The following authors will receive royalty income from this licensing agreement held by the Federal Government: Quezado, Finkel, Besch, Pohida, and Kakareka. None of the authors have personal financial interest in the subject matter or equipment discussed in the article. In addition, the entire work was funded solely by the National Institutes of Health intramural program.
The authors thank Robert W. Finkel, Ph.D. (Professor of Physics, St. John’s University, Queens, New York), and Olavo Vasconcelos, M.D. (Assistant Professor of Medicine, Uniformed Services University of the Health Sciences), for their helpful comments and review of the manuscript.
Footnotes
Presented in part at the Annual Meeting of the American Society of Anesthesiologists, Atlanta, Georgia, October 15–19, 2005.
References
- 1.Helme RD, Gibson SJ. The epidemiology of pain in elderly people. Clin Geriatr Med. 2001;17:417–31. v. doi: 10.1016/s0749-0690(05)70078-1. [DOI] [PubMed] [Google Scholar]
- 2.Pickering G, Jourdan D, Eschalier A, Dubray C. Impact of age, gender and cognitive functioning on pain perception. Gerontology. 2002;48:112–8. doi: 10.1159/000048937. [DOI] [PubMed] [Google Scholar]
- 3.Edwards RR, Fillingim RB, Ness TJ. Age-related differences in endogenous pain modulation: A comparison of diffuse noxious inhibitory controls in healthy older and younger adults. Pain. 2003;101:155–65. doi: 10.1016/s0304-3959(02)00324-x. [DOI] [PubMed] [Google Scholar]
- 4.Lautenbacher S, Kunz M, Strate P, Nielsen J, Arendt-Nielsen L. Age effects on pain thresholds, temporal summation and spatial summation of heat and pressure pain. Pain. 2005;115:410–8. doi: 10.1016/j.pain.2005.03.025. [DOI] [PubMed] [Google Scholar]
- 5.Gibson SJ, Farrell M. A review of age differences in the neurophysiology of nociception and the perceptual experience of pain. Clin J Pain. 2004;20:227–39. doi: 10.1097/00002508-200407000-00004. [DOI] [PubMed] [Google Scholar]
- 6.Gibson SJ, Helme RD. Age-related differences in pain perception and report. Clin Geriatr Med. 2001;17:433–56. v–vi. doi: 10.1016/s0749-0690(05)70079-3. [DOI] [PubMed] [Google Scholar]
- 7.Porter FL, Grunau RE, Anand KJ. Long-term effects of pain in infants. J Dev Behav Pediatr. 1999;20:253–61. doi: 10.1097/00004703-199908000-00008. [DOI] [PubMed] [Google Scholar]
- 8.Porter FL, Wolf CM, Miller JP. Procedural pain in newborn infants: The influence of intensity and development. Pediatrics. 1999;104:e13. doi: 10.1542/peds.104.1.e13. [DOI] [PubMed] [Google Scholar]
- 9.Taddio A, Shah V, Gilbert-MacLeod C, Katz J. Conditioning and hyperalgesia in newborns exposed to repeated heel lances. JAMA. 2002;288:857–61. doi: 10.1001/jama.288.7.857. [DOI] [PubMed] [Google Scholar]
- 10.Anand KJ, Scalzo FM. Can adverse neonatal experiences alter brain development and subsequent behavior? Biol Neonate. 2000;77:69–82. doi: 10.1159/000014197. [DOI] [PubMed] [Google Scholar]
- 11.Johnston CC, Stevens BJ. Experience in a neonatal intensive care unit affects pain response. Pediatrics. 1996;98:925–30. [PubMed] [Google Scholar]
- 12.Gagliese L, Melzack R. Chronic pain in elderly people. Pain. 1997;70:3–14. doi: 10.1016/s0304-3959(96)03266-6. [DOI] [PubMed] [Google Scholar]
- 13.Verhaak PF, Kerssens JJ, Dekker J, Sorbi MJ, Bensing JM. Prevalence of chronic benign pain disorder among adults: A review of the literature. Pain. 1998;77:231–9. doi: 10.1016/S0304-3959(98)00117-1. [DOI] [PubMed] [Google Scholar]
- 14.Rustoen T, Wahl AK, Hanestad BR, Lerdal A, Paul S, Miaskowski C. Age and the experience of chronic pain: Differences in health and quality of life among younger, middle-aged, and older adults. Clin J Pain. 2005;21:513–23. doi: 10.1097/01.ajp.0000146217.31780.ef. [DOI] [PubMed] [Google Scholar]
- 15.Gagliese L, Melzack R. Age differences in the response to the formalin test in rats. Neurobiol Aging. 1999;20:699–707. doi: 10.1016/s0197-4580(99)00061-5. [DOI] [PubMed] [Google Scholar]
- 16.Chakour MC, Gibson SJ, Bradbeer M, Helme RD. The effect of age on A delta- and C-fibre thermal pain perception. Pain. 1996;64:143–52. doi: 10.1016/0304-3959(95)00102-6. [DOI] [PubMed] [Google Scholar]
- 17.Harkins SW, Davis MD, Bush FM, Kasberger J. Suppression of first pain and slow temporal summation of second pain in relation to age. J Gerontol A Biol Sci Med Sci. 1996;51:M260–5. doi: 10.1093/gerona/51a.5.m260. [DOI] [PubMed] [Google Scholar]
- 18.Edwards RR, Fillingim RB. Effects of age on temporal summation and habituation of thermal pain: Clinical relevance in healthy older and younger adults. J Pain. 2001;2:307–17. doi: 10.1054/jpai.2001.25525. [DOI] [PubMed] [Google Scholar]
- 19.Nadon NL. Maintaining aged rodents for biogerontology research. Lab Anim (NY) 2004;33:36–41. doi: 10.1038/laban0904-36. [DOI] [PubMed] [Google Scholar]
- 20.Haines DC, Chattopadhyay S, Ward JM. Pathology of aging B6;129 mice. Toxicol Pathol. 2001;29:653–61. doi: 10.1080/019262301753385988. [DOI] [PubMed] [Google Scholar]
- 21.Koga K, Furue H, Rashid MH, Takaki A, Katafuchi T, Yoshimura M. Selective activation of primary afferent fibers evaluated by sine-wave electrical stimulation. Mol Pain. 2005;1:13–23. doi: 10.1186/1744-8069-1-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Gagliese L, Melzack R. Age differences in nociception and pain behaviours in the rat. Neurosci Biobehav Rev. 2000;24:843–54. doi: 10.1016/s0149-7634(00)00041-5. [DOI] [PubMed] [Google Scholar]
- 23.Gagliese L, Farrel MJ. The neurobiology of aging, nociception, and pain: An integration of animal and human experimental evidence. In: Gibson SJ, Weiner DK, editors. Pain in Older Persons. Seattle: IASP Press; 2005. pp. 25–44. [Google Scholar]
- 24.Jourdan D, Boghossian S, Alloui A, Veyrat-Durebex C, Coudore MA, Eschalier A, Alliot J. Age-related changes in nociception and effect of morphine in the Lou rat. Eur J Pain. 2000;4:291–300. doi: 10.1053/eujp.2000.0188. [DOI] [PubMed] [Google Scholar]
- 25.Jourdan D, Pickering G, Marchand F, Gaulier JM, Alliot J, Eschalier A. Impact of ageing on the antinociceptive effect of reference analgesics in the Lou/c rat. Br J Pharmacol. 2002;137:813–20. doi: 10.1038/sj.bjp.0704944. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Goettl VM, Lindsey AE, Neff NH, Hadjiconstantinou M. GM1 ganglioside restores abnormal responses to acute thermal and mechanical stimuli in aged rats. Brain Res. 2000;858:380–5. doi: 10.1016/s0006-8993(00)01960-0. [DOI] [PubMed] [Google Scholar]
- 27.Conway CM, Martinez J, Lytle LD. Maturational changes in the thermal nociceptive responses of developing rats. Dev Psychobiol. 1998;33:47–60. [PubMed] [Google Scholar]
- 28.Akunne HC, Soliman KF. Serotonin modulation of pain responsiveness in the aged rat. Pharmacol Biochem Behav. 1994;48:411–6. doi: 10.1016/0091-3057(94)90545-2. [DOI] [PubMed] [Google Scholar]
- 29.Hess GD, Joseph JA, Roth GS. Effect of age on sensitivity to pain and brain opiate receptors. Neurobiol Aging. 1981;2:49–55. doi: 10.1016/0197-4580(81)90059-2. [DOI] [PubMed] [Google Scholar]
- 30.Gordon WC, Scobie SR, Frankl SE. Age-related differences in electric shock detection and escape thresholds in Sprague-Dawley albino rats. Exp Aging Res. 1978;4:23–35. doi: 10.1080/03610737808257124. [DOI] [PubMed] [Google Scholar]
- 31.Helme RD, Meliala A, Gibson SJ. Methodologic factors which contribute to variations in experimental pain threshold reported for older people. Neurosci Lett. 2004;361:144–6. doi: 10.1016/j.neulet.2003.12.014. [DOI] [PubMed] [Google Scholar]
- 32.Verdu E, Ceballos D, Vilches JJ, Navarro X. Influence of aging on peripheral nerve function and regeneration. J Peripher Nerv Syst. 2000;5:191–208. doi: 10.1046/j.1529-8027.2000.00026.x. [DOI] [PubMed] [Google Scholar]
- 33.Ceballos D, Cuadras J, Verdu E, Navarro X. Morphometric and ultrastructural changes with ageing in mouse peripheral nerve. J Anat. 1999;195(pt 4):563–76. doi: 10.1046/j.1469-7580.1999.19540563.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Verdu E, Buti M, Navarro X. Functional changes of the peripheral nervous system with aging in the mouse. Neurobiol Aging. 1996;17:73–7. doi: 10.1016/0197-4580(95)02010-1. [DOI] [PubMed] [Google Scholar]
- 35.Hargreaves K, Dubner R, Brown F, Flores C, Joris J. A new and sensitive method for measuring thermal nociception in cutaneous hyperalgesia. Pain. 1988;32:77–88. doi: 10.1016/0304-3959(88)90026-7. [DOI] [PubMed] [Google Scholar]
- 36.Hargraves WA, Hentall ID. Analgesic effects of dietary caloric restriction in adult mice. Pain. 2005;114:455–61. doi: 10.1016/j.pain.2005.01.010. [DOI] [PubMed] [Google Scholar]
- 37.Yarnitsky D, Sprecher E, Zaslansky R, Hemli JA. Heat pain thresholds: Normative data and repeatability. Pain. 1995;60:329–32. doi: 10.1016/0304-3959(94)00132-x. [DOI] [PubMed] [Google Scholar]
- 38.Verdugo R, Ochoa JL. Quantitative somatosensory thermotest: A key method for functional evaluation of small calibre afferent channels. Brain. 1992;115(pt 3):893–913. doi: 10.1093/brain/115.3.893. [DOI] [PubMed] [Google Scholar]
- 39.Le Bars D, Gozariu M, Cadden SW. Animal models of nociception. Pharmacol Rev. 2001;53:597–652. [PubMed] [Google Scholar]
- 40.Hogan Q. Animal pain models. Reg Anesth Pain Med. 2002;27:385–401. doi: 10.1053/rapm.2002.33630. [DOI] [PubMed] [Google Scholar]
- 41.Jourdan D, Ardid D, Chapuy E, Eschalier A, Le Bars D. Audible and ultrasonic vocalization elicited by single electrical nociceptive stimuli to the tail in the rat. Pain. 1995;63:237–49. doi: 10.1016/0304-3959(95)00049-X. [DOI] [PubMed] [Google Scholar]
- 42.Kobayashi H, Kikuchi K, Tsubono Y, Tagami H. Measurement of electrical current perception threshold of sensory nerves for pruritus in atopic dermatitis patients and normal individuals with various degrees of mild damage to the stratum corneum. Dermatology. 2003;206:204–11. doi: 10.1159/000068892. [DOI] [PubMed] [Google Scholar]
- 43.Katims J. Electrodiagnostic functional sensory evaluation of the patient with pain: A review of the neuroselective current perception threshold and pain tolerance threshold. Pain Digest. 1998;8:219–230. [Google Scholar]
- 44.Masson EA, Boulton AJ. The Neurometer: Validation and comparison with conventional tests for diabetic neuropathy. Diabet Med. 1991;8:S63–6. doi: 10.1111/j.1464-5491.1991.tb02159.x. [DOI] [PubMed] [Google Scholar]
- 45.Oishi M, Mochizuki Y, Suzuki Y, Ogawa K, Naganuma T, Nishijo Y, Mizutani T. Current perception threshold and sympathetic skin response in diabetic and alcoholic polyneuropathies. Intern Med. 2002;41:819–22. doi: 10.2169/internalmedicine.41.819. [DOI] [PubMed] [Google Scholar]
- 46.Finkel JC, Yang CI, Yarvitz JL, Patel KM. Neuroselective sensory electrodiagnostic evaluation of 4% liposomal topical lidocaine. Anesth Analg. 2002;94:1259–62. doi: 10.1097/00000539-200205000-00039. [DOI] [PubMed] [Google Scholar]
- 47.Liem EB, Joiner TV, Tsueda K, Sessler DI. Increased sensitivity to thermal pain and reduced subcutaneous lidocaine efficacy in redheads. Anesthesiology. 2005;102:509–14. doi: 10.1097/00000542-200503000-00006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Alkire MT. Functional brain imaging: Some parameters for success. Anesthesiology. 2004;101:817–9. doi: 10.1097/00000542-200410000-00003. [DOI] [PubMed] [Google Scholar]
- 49.Kiso T, Nagakura Y, Toya T, Matsumoto N, Tamura S, Ito H, Okada M, Yamaguchi T. Neurometer measurement of current stimulus threshold in rats. J Pharmacol Exp Ther. 2001;297:352–6. [PubMed] [Google Scholar]
- 50.Oda M, Kitagawa N, Yang BX, Totoki T, Morimoto M. Quantitative and fiber-selective evaluation of dose-dependent nerve blockade by intrathecal lidocaine in rats. J Pharmacol Exp Ther. 2005;312:1132–7. doi: 10.1124/jpet.104.076893. [DOI] [PubMed] [Google Scholar]
- 51.Thompson SW, King AE, Woolf CJ. Activity-dependent changes in rat ventral horn neurons in vitro: Summation of prolonged afferent evoked postsynaptic depolarizations produce a d-2-amino-5-phosphonovaleric acid sensitive windup. Eur J Neurosci. 1990;2:638–49. doi: 10.1111/j.1460-9568.1990.tb00453.x. [DOI] [PubMed] [Google Scholar]
- 52.Sivilotti LG, Thompson SW, Woolf CJ. Rate of rise of the cumulative depolarization evoked by repetitive stimulation of small-caliber afferents is a predictor of action potential windup in rat spinal neurons in vitro. J Neurophysiol. 1993;69:1621–31. doi: 10.1152/jn.1993.69.5.1621. [DOI] [PubMed] [Google Scholar]
- 53.Coderre TJ, Katz J, Vaccarino AL, Melzack R. Contribution of central neuroplasticity to pathological pain: Review of clinical and experimental evidence. Pain. 1993;52:259–85. doi: 10.1016/0304-3959(93)90161-H. [DOI] [PubMed] [Google Scholar]
- 54.Woolf CJ, Chong MS. Preemptive analgesia: Treating postoperative pain by preventing the establishment of central sensitization. Anesth Analg. 1993;77:362–79. doi: 10.1213/00000539-199377020-00026. [DOI] [PubMed] [Google Scholar]
- 55.Thompson SW, Woolf CJ, Sivilotti LG. Small-caliber afferent inputs produce a heterosynaptic facilitation of the synaptic responses evoked by primary afferent A-fibers in the neonatal rat spinal cord in vitro. J Neurophysiol. 1993;69:2116–28. doi: 10.1152/jn.1993.69.6.2116. [DOI] [PubMed] [Google Scholar]
- 56.Tabata H, Ikegami H, Kariya K. A parallel comparison of age-related peripheral nerve changes in three different strains of mice. Exp Anim. 2000;49:295–9. doi: 10.1538/expanim.49.295. [DOI] [PubMed] [Google Scholar]
- 57.Magnusson KR, Nelson SE, Young AB. Age-related changes in the protein expression of subunits of the NMDA receptor. Brain Res Mol Brain Res. 2002;99:40–5. doi: 10.1016/s0169-328x(01)00344-8. [DOI] [PubMed] [Google Scholar]
- 58.Clayton DA, Browning MD. Deficits in the expression of the NR2B subunit in the hippocampus of aged Fisher 344 rats. Neurobiol Aging. 2001;22:165–8. doi: 10.1016/s0197-4580(00)00196-2. [DOI] [PubMed] [Google Scholar]
- 59.Kanda K, Sato H, Kemuriyama T, Iwata K. Temporal facilitation of the flexor reflex induced by C-fiber activity: Comparison between adult and aged rats. Neurosci Lett. 2001;304:49–52. doi: 10.1016/s0304-3940(01)01761-x. [DOI] [PubMed] [Google Scholar]
- 60.Mogil JS, Chanda ML. The case for the inclusion of female subjects in basic science studies of pain. Pain. 2005;117:1–5. doi: 10.1016/j.pain.2005.06.020. [DOI] [PubMed] [Google Scholar]