

Abstract
Large-scale, comparative cognition studies are set to revolutionize the way we investigate and understand the evolution of intelligence. However, the conclusions reached by such work have a key limitation: the cognitive tests themselves. If factors other than cognition can systematically affect the performance of a subset of animals on these tests, we risk drawing the wrong conclusions about how intelligence evolves. Here, we examined whether this is the case for the A-not-B task, recently used by MacLean and co-workers to study self-control among 36 different species. Non-primates performed poorly on this task; possibly because they have difficulty tracking the movements of a human demonstrator, and not because they lack self-control. To test this, we assessed the performance of New Caledonian crows on the A-not-B task before and after two types of training. New Caledonian crows trained to track rewards moved by a human demonstrator were more likely to pass the A-not-B test than birds trained on an unrelated choice task involving inhibitory control. Our findings demonstrate that overlooked task demands can affect performance on a cognitive task, and so bring into question MacLean's conclusion that absolute brain size best predicts self-control.
Keywords: evolution of intelligence, comparative cognition, self-control, corvids, primates, New Caledonian crows
1. Introduction
With developments in phylogenetic analyses, and an increasing ability to coordinate research efforts around the world, large-scale comparisons of cognition across multiple species are set to revolutionize the way we investigate the evolution of intelligence [1]. However, this type of work is only as robust as the comparative tests used. If factors other than variation in cognition can systematically affect the performance of a subset of the animals tested, there is a real danger that faulty conclusions will be drawn.
In 2014, MacLean and co-workers reported on an impressive collaborative study testing 36 species of primates, birds, rodents, carnivores and elephants on two self-control tasks [2]. (The use of the term ‘self-control’ to describe these tasks is debated [3], but for consistency we follow MacLean's usage here.) The first was a ‘cylinder’ task, where subjects had to detour to the side of a transparent cylinder to retrieve food. The second was an ‘A-not-B’ task, where subjects retrieved food from cup ‘A’ three times, then witnessed a human move the food from cup ‘A’ to cup ‘B’. Subjects had to inhibit their desire to select previously rewarded cup ‘A’ to succeed. However, they also had to accurately track the movement of food by a human hand. This additional demand may have selectively disadvantaged non-primates, who frequently fail tasks involving human demonstrators [4–7]. Given this confounding factor, there is a real possibility that the conclusion reached by MacLean et al. [2]—that absolute brain size predicts self-control—is wrong.
Supporting this, while primates performed similarly on the cylinder and A-not-B tasks, six of the eight non-primates given both tasks showed a drop in performance of 41–66% on the A-not-B task. Elephants failed every trial. Pigeons and domestic dogs were the only exceptions, and strikingly, they were the only non-primates trained to retrieve food from containers after witnessing baiting. It is therefore unclear if the (almost unanimous) test failures by non-primates were due to limited self-control or an inability to track food moved by human hands.
Across the discipline of animal cognition, there is a growing impetus for researchers to attend to failures, which have a multitude of causes, and not just problem-solving successes [8,9]. This becomes essential when we intend to draw conclusions from variation in test performance, both across and within species [10]. Lowering task demands, or correlating performance from different tasks, can provide valuable information about why a subject or species failed a particular test [8]. Even more powerful are intervention studies, where subjects are trained to overcome a specific weakness, to determine whether this was a cause of their failure (a practice commonly used to study developmental disorders [11]).
Here, we ran an intervention study with two groups of wild-caught New Caledonian crows to assess their performance on the A-not-B task before and after training. One group was trained to attend to the movement of food by a human demonstrator, while the other received reversal learning: an unrelated choice task, but one that involves inhibitory control [12]. New Caledonian crows have small absolute, but large relative, brain sizes [13]. However, like several species with similarly small brains [14,15], they display various behaviours that involve self-control. New Caledonian crows delay foraging in order to manufacture tools in the wild [16] and perform well on cognitive tasks that involve inhibiting their approach towards food [17] or switching their responses across trials [18,19]. Thus, if this species fails the A-not-B task, it may be because they struggle to attend to the movement of food by a human demonstrator. If this is the case, training crows to track the demonstrator's actions should improve their A-not-B performance.
2. Material and method
(a). Subjects
Subjects were 14 wild-caught New Caledonian crows held in captivity for 10 to 17 weeks before testing began (table 1). All birds had previously taken part in experiments involving tool-use and learning object properties, and some had taken part in a cooperative study with conspecifics (light blue, D3R, D4R, D4B and RWY [20]), but no subjects had ever been required to attend to human actions or the movement of rewards.
Table 1.
Subject information and performance.
| post-training A-not-B tests |
|||||||
|---|---|---|---|---|---|---|---|
| bird | sex | age class | boldness | passed first A-not-B test? | passed training? | correct/5 | order of successes |
| experimental group: hand-tracking | |||||||
| white | F | juvenile | bold | N | Y | 5 | 11111 |
| RWY | M | adult | medium | N | Y | 3 | 01011 |
| D3R | F | juvenile | bold | N | Y | 2 | 01100 |
| Azzuro | M | adult | medium | N | N | — | — |
| Emma | F | adult | bold | N | N | — | — |
| Joe | M | adult | medium | N | N | — | — |
| light blue | F | juvenile | bold | N | N | — | — |
| D4R | F | juvenile | medium | N | N | — | — |
| control group: reverse learning | |||||||
| red | F | adult | medium | N | Y | 1 | 00010 |
| D4G | F | juvenile | medium | N | Y | 1 | 00010 |
| D4B | M | adult | bold | N | Y | 0 | 00000 |
| Anton | M | juvenile | medium | N | Y | 0 | 00000 |
| Stella | F | adult | bold | N | Y | 0 | 00000 |
| blue | M | juvenile | bold | N | Y | 0 | 00000 |
(b). Procedure
(i). Initial A-not-B test
Birds were habituated to all elements of the test set-up and trained to remove cardboard lids from pre-baited cups. In all cases, bait was meat, which these crows do not appear to locate using olfactory cues (electronic supplementary material). They then received an A-not-B test. At the start of each trial, three cups and three lids were placed on a table at the front of a 2.5 × 5 × 3 m cage (figure 1). From outside the cage, the experimenter reached through a flap to bait one cup (A) and lid all three cups. When the bird chose the baited cup (A) three trials in a row, they received an A-not-B trial, where, after baiting A, the experimenter visibly took the reward out of A and moved it to B (electronic supplementary material, movie). To pass, birds had to choose cup B. To be comparable with [2], we followed the procedure for Eurasian jays, except that the experimenter's body was concealed. This was necessary to minimize fear among our wild-caught birds.
Figure 1.
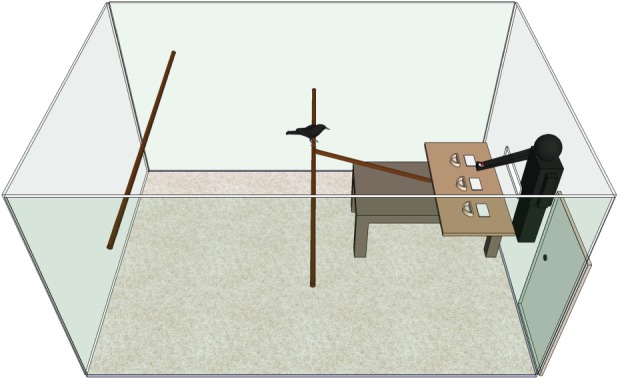
Diagram of the experimental set-up. The experimenter reached through a flap in the cage to bait and lid the cups. All walls were opaque. (Online version in colour.)
(ii). Experimental training (hand-tracking)
Birds were assigned to one of two training conditions before the experiment began, balancing sex, age and boldness (table 1; electronic supplementary material, methods). All birds that failed the initial A-not-B test (which all 14 birds did) then received training. Experimental subjects were trained to attend to the experimenter baiting one of three cups. In each trial, three cups and three lids were placed on the table, then, from outside the cage, the experimenter placed the reward on the table, lidded all three cups, then lifted each lid in turn, placing the reward in one cup. The rewarded cup, the order the lids were lifted and whether the first, second or third cup touched was baited were pseudo-randomized. This ensured birds attended to the movement of the food, not to cues such as ‘which lid was touched last’. Subjects were trained until they achieved 9/10 correct trials with this procedure, or received 250 trials of any kind (including modified trials to overcome side biases or motivational problems).
(iii). Control training (reverse learning)
Control subjects received a reverse colour learning task, conducted in the same location as hand-tracking training, which required inhibitory control, but not attention to human actions or movement of rewards. Subjects had to inhibit choosing a previously rewarded container to succeed; therefore, any differences between experimental and control groups at test were unlikely to stem from experimental birds developing better self-control than the controls during training. In each trial, the experimenter placed two coloured tubes on the table, one containing a reward, then exited the room. Once birds learnt that one colour was rewarded (9/10 correct trials), the rewarded colour was reversed, and birds had to choose the previously unrewarded colour in 9/10 trials. Training continued until birds passed or received 250 reversal trials.
(iv). Post-training five A-not-B tests
Subjects that passed the training (3/8 experimental birds, 6/6 controls) then received five further A-not-B tests (procedure as before). The identity of the A and B cups (left, middle or right cup) changed for each test.
(v). Choices
For the A-not-B tests and hand-tracking training, choices were defined as the bird touching a cup or lid with its beak. In practice, all birds removed the lid of the first cup they touched, making choices unambiguous. For reversal learning, choices were defined as obtaining the reward (if correct) or approaching, within 10 cm, the open end of the empty tube (if incorrect), where the contents would be visible. Any ambiguous choices were recorded as incorrect. Birds were given time to consume rewards if obtained, but were only permitted to make one choice in each trial. This was enforced by the experimenter entering the room to end the trial. This condition was relaxed for some birds during hand-tracking training if they lost motivation, in which case they were allowed to make multiple choices until their motivation increased. Trials were scored live by the experimenter (S.A.J. or assistant) and then from video by S.A.J.
3. Results
All 14 subjects failed the initial A-not-B test. For the hand-tracking training, three out of eight experimental birds passed—taking 100, 150 and 190 trials. A further three birds reached 250 trials, and two had to be stopped at 150 trials owing to motivation and time constraints. All control birds passed colour learning within 10–20 trials, then reverse learning within 40–70 trials (mean: 56.7). The three birds that passed the hand-tracking training also passed the majority of A-not-B tests, scoring 5/5, 3/5 and 2/5 (median: 3/5). One bird passed the first test, and all passed the second test. In contrast, the control birds failed almost every A-not-B test: four scored 0/5 and two scored 1/5, both passing the fourth test only (median: 0/5, table 1). The performances of the two groups were significantly different (Mann–Whitney U-test: U = 0, n1 = 3, n2 = 6, p = 0.02).
4. Discussion
Our results demonstrate that New Caledonian crows are more successful on the A-not-B task if they have been trained to retrieve rewards placed inside cups by a human demonstrator, compared with if they only receive training on an unrelated inhibitory control task. In our small sample, all three birds that learnt to track the demonstrator's actions passed more A-not-B tests than all of the controls. Only one crow passed on the first trial; thus their performance was not perfect. However, if we compare these results with MacLean's (bearing in mind that we conducted multiple tests per subject), New Caledonian crows that learnt to track hands rank joint ninth out of 27 species, equivalent to long-tailed macaques, with 67% success. However, with just 7% success, New Caledonian crows that received reversal learning training ranked second from last, surpassing only elephants [2].
Learning to track hands was difficult. Only three out of eight birds passed this training, which confirms that these crows struggle to follow human demonstrators. Given that the successful birds then passed more A-not-B tests, we should be cautious of the conclusion drawn by MacLean et al. [2], that absolute brain size predicts self-control, particularly in relation to non-primates. We do not know whether subjects could track hands before being given the A-not-B test; therefore, we do not know whether poor performance on this test reflects issues with hand-tracking or issues with self-control. Indeed, several species that performed very poorly on the A-not-B test display impressive self-control in other areas. Elephants wait up to 45 s for a partner in a cooperative task [21] and both Eurasian and Western scrub jays can plan for the future [14,15]. More generally, birds such as corvids, with very small absolute brain sizes, regularly rival apes or monkeys on cognitive tasks that place demands on self-control [9,22].
Our results show that prior experience affects A-not-B task success. However, in addition to learning to track hands, our experimental group also had more experience with the A-not-B apparatus. This is unlikely to have driven our results because both groups were extensively trained to remove lids; however, our findings should be confirmed using identical apparatuses for both training tasks, and, more generally, by conducting studies with different tasks, training and greater sample sizes.
Our findings provide a cautionary tale for future studies comparing cognitive test performances across multiple species. As demonstrated here, if subjects can fail a task for multiple reasons, variation in test performance will not reflect meaningful variation in cognition. We therefore suggest a threefold approach for designing comparative tests. Lowering additional task demands to an absolute minimum is an important starting point [8]. However, it is also critical to level the playing field from the bottom up by using baseline criterion training. Such training ensures that subjects can attend to all additional elements of the task before tests of the desired cognitive ability begin. A third avenue is to adopt a signature-testing approach, where not just problem-solving successes, but also information biases, errors and limits, are compared across species [9]. By reducing task demands, using thorough baseline criterion training, and adopting a signature-testing approach, phylogenetic comparative studies can provide real insights into the evolution of cognition.
Supplementary Material
Supplementary Material
Acknowledgements
Thanks to animal trainer Martina Scheistl, Dakota McCoy for pilot work, and Guido DeFillippo and Yulla Knaus for collecting data.
Ethics
This study was conducted under University of Auckland ethics committee approval (R602). The Province Sud granted us permission to work in New Caledonia.
Data accessibility
The datasets supporting this article have been uploaded as part of the electronic supplementary material.
Authors' contributions
S.A.J. conceived and designed the study, collected data and drafted the manuscript, A.H.T. and R.D.G. contributed to the design and commented on the manuscript. All authors approved the final version and are accountable for its content.
Competing interests
We have no competing interests.
Funding
This research was supported by the NZ Marsden fund (R.D.G.) and a Rutherford Fellowship (A.H.T.).
References
- 1.MacLean EL, et al. 2012. How does cognition evolve? Phylogenetic comparative psychology. Anim. Cogn. 15, 223–238. ( 10.1007/s10071-011-0448-8) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.MacLean EL, et al. 2014. The evolution of self-control. Proc. Natl Acad. Sci. USA 111, E2140–E2148. ( 10.1073/pnas.1323533111) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Beran MJ. 2015. The comparative science of ‘self-control’: what are we talking about? Front. Psychol. 6, 2 ( 10.3389/fpsyg.2015.00051) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Shaw RC, Plotnik JM, Clayton NS. 2013. Exclusion in corvids: the performance of food-caching Eurasian jays (Garrulus glandarius). J. Comp. Psychol. 127, 428–435. ( 10.1037/a0032010) [DOI] [PubMed] [Google Scholar]
- 5.Mikolasch S, Kotrschal K, Schloegl C. 2012. Is caching the key to exclusion in corvids? The case of carrion crows (Corvus corone corone). Anim. Cogn. 15, 73–82. ( 10.1007/s10071-011-0434-1) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Erdőhegyi Á, Topál J, Virányi Z, Miklósi Á. 2007. Dog-logic: inferential reasoning in a two-way choice task and its restricted use. Anim. Behav. 74, 725–737. ( 10.1016/j.anbehav.2007.03.004) [DOI] [Google Scholar]
- 7.Nawroth C, von Borell E, Langbein J. 2014. Exclusion performance in dwarf goats (Capra aegagrus hircus) and sheep (Ovis orientalis aries). PLoS ONE 9, e93534 ( 10.1371/journal.pone.0093534) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Seed A, Seddon E, Greene B, Call J. 2012. Chimpanzee ‘folk physics’: bringing failures into focus. Phil. Trans. R. Soc. B 367, 2743–2752. ( 10.1098/rstb.2012.0222) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Taylor AH. 2014. Corvid cognition. Wiley Interdiscip. Rev. Cogn. Sci. 5, 361–372. ( 10.1002/wcs.1286) [DOI] [PubMed] [Google Scholar]
- 10.Rowe C, Healy SD. 2014. Measuring variation in cognition. Behav. Ecol. 25, 1287–1292. ( 10.1093/beheco/aru090) [DOI] [Google Scholar]
- 11.Schneider W, Roth E, Ennemoser M. 2000. Training phonological skills and letter knowledge in children at risk for dyslexia: a comparison of three kindergarten intervention programs. J. Educ. Psychol. 92, 284–295. ( 10.1037/0022-0663.92.2.284) [DOI] [Google Scholar]
- 12.Dias R, Robbins T, Roberts A. 1996. Dissociation in prefrontal cortex of affective and attentional shifts. Nature 380, 69–72. ( 10.1038/380069a0) [DOI] [PubMed] [Google Scholar]
- 13.Cnotka J, Güntürkün O, Rehkämper G, Gray RD, Hunt GR. 2008. Extraordinary large brains in tool-using New Caledonian crows (Corvus moneduloides). Neurosci. Lett. 433, 241–245. ( 10.1016/j.neulet.2008.01.026) [DOI] [PubMed] [Google Scholar]
- 14.Raby CR, Alexis DM, Dickinson A, Clayton NS. 2007. Planning for the future by western scrub-jays. Nature 445, 919–921. ( 10.1038/nature05575) [DOI] [PubMed] [Google Scholar]
- 15.Cheke LG, Clayton NS. 2012. Eurasian jays (Garrulus glandarius) overcome their current desires to anticipate two distinct future needs and plan for them appropriately. Biol. Lett. 8, 171–175. ( 10.1098/rsbl.2011.0909) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Hunt GR. 1996. Manufacture and use of hook-tools by New Caledonian crows. Nature 379, 249–251. ( 10.1038/379249a0) [DOI] [Google Scholar]
- 17.Taylor AH, Hunt GR, Holzhaider JC, Gray RD. 2007. Spontaneous metatool use by New Caledonian crows. Curr. Biol. 17, 1504–1507. ( 10.1016/j.cub.2007.07.057) [DOI] [PubMed] [Google Scholar]
- 18.Jelbert SA, Taylor AH, Gray RD. 2015. Reasoning by exclusion in New Caledonian crows (Corvus moneduloides) cannot be explained by avoidance of empty containers. J. Comp. Psychol. 129, 283–290. ( 10.1037/a0039313) [DOI] [PubMed] [Google Scholar]
- 19.Taylor AH, Hunt GR, Medina FS, Gray RD. 2009. Do New Caledonian crows solve physical problems through causal reasoning? Proc. R. Soc. B 276, 247–254. ( 10.1098/rspb.2008.1107) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Jelbert SA, Singh PJ, Gray RD, Taylor AH. 2015. New Caledonian crows rapidly solve a collaborative problem without cooperative cognition. PLoS ONE 10, e0133253 ( 10.1371/journal.pone.0133253) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Plotnik JM, Lair R, Suphachoksahakun W, de Waal FB. 2011. Elephants know when they need a helping trunk in a cooperative task. Proc. Natl Acad. Sci. USA 108, 5116–5121. ( 10.1073/pnas.1101765108) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Emery NJ, Clayton NS. 2004. The mentality of crows: convergent evolution of intelligence in corvids and apes. Science 306, 1903–1907. ( 10.1126/science.1098410) [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The datasets supporting this article have been uploaded as part of the electronic supplementary material.