

Abstract
This protocol describes a simple and quick method to isolate and characterize mouse antral GV (Germinal Vesicle) oocytes as able (SN, Surrounded Nucleolus) or unable (NSN, Not Surrounded Nucleolus) to develop to the blastocyst stage after in vitro maturation (IVM) and in vitro fertilization (IVF). It makes use of Hoeschst33342 (or any other DNA intercalating dye) able to bind to the heterochromatin of the nucleolus showing a ring in the SN oocytes or not, like in the NSN oocytes. This represents the easiest and quickest way to sort both antral oocytes that can be eventually used for IVM or IVF procedures.
Briefly, the protocol consists of the following steps: hormone injection to stimulate follicular growth; isolation of the oocytes at the GV stage from the antral compartment by puncturing the ovary with a sterile needle; preparation of thin glass pipettes for mouth pipetting of the oocytes; sorting of the oocytes with Hoechst33342 prepared at a supravital concentration; IVM, IVF or any other molecular/cellular analysis.
Unfortunately there are still few evidences to sort SN and NSN oocytes using less invasive techniques. If and once they will be identified, they could be potentially applied to human assisted reproductive technologies, although with several aspects that should be modified. To date, this technique has potential implications to dramatically increase IVM and IVF successful procedures in both endangered and species with economic interest.
Keywords: Developmental Biology, Issue 107, oocyte, nucleolus, chromatin organization, Hoechst33342, oocyte maturation, embryo development, infertility, developmental biology
Introduction
Antral fully grown oocytes (GV, germinal vesicle) isolated from the antral compartment of the ovary are routinely used for different purposes such as, for example, the study of the molecular and cellular mechanisms behind in vitro maturation till Metaphase II stage.
Despite their beautiful appearance most antral oocytes, but not all, are able to complete meiosis and reach the blastocyst stage; in fact, in many mammalian species, including humans1,2, roughly 30% of them arrest their development at the 2-cell stage, if fertilization occurs3-5. To date, few parameters are used to distinguish between oocytes able to sustain the embryonic development from those unable to do so.
For example, Cresyl blue staining (BCB) is a valid method to sort the oocytes based on the activity of glucose-6-phosphate-dehydrogenase although the utility of this sorting method related to BCB+ or BCB- staining is still laboratory dependent6-8.
Another well-established method resides in their chromatin organization: if stained with any DNA intercalating dyes (like Hoechst33342), 70% of antral oocytes show a dye-positive ring around the nucleolus and are called Surrounded Nucleolus (SN) while the remaining 30% show a more spotted positive signals and are called Not Surrounded Nucleolus (NSN)3-5. Recently we showed that the absence of cytoplasmic lattices9 (CPLs, important storage sites for maternally derived mRNA and ribosomes in both mammalian oocytes and embryos)10 and the down-regulation of MATER (essential for CPLs formation11), together with the presence of lipid droplets in their cytoplasms9,12, can be other causes related to the NSN oocytes inability to proceed beyond the 2-cell stage.
Unfortunately, few techniques can be applied to sort antral SN and NSN oocytes and, for this reason, the development of a less-invasive method (without nuclear staining, fixation procedures or manipulation with mouth pipettes) could become very important to increase the rates of both human and animal embryo development.
In this paper, we detail the simple and quick protocol used to isolate ad sort SN and NSN antral oocytes from female mice and it is addressed to all the scientists interested in the study of female gametogenesis, oocyte maturation and embryo development.
Protocol
This protocol is conducted in accordance with the guiding principles of European (EU 63/2010) and Italian (26/2014) laws protecting animals used for scientific research. Cervical dislocation euthanasia is used for ovaries isolation and it is performed by expert and trained individuals listed in the approved protocol covering this study.
1. Animals
Maintain adult 3- to 6-week-old female mice in a room with controlled temperature and humidity (with phases of 12L:12D and fed ad libitum).
Inject mice intraperitoneally with 2.5 IU of Pregnant mare serum gonadotropin (PMSG) to stimulate follicular growth by mimicking the effects of follicle stimulating hormone (FSH). Isolate the ovaries for antral oocytes collection 48hrs later. Euthanize the female mouse previously injected with PMSG, according to your institutional guidelines.
- Quickly remove the ovaries from the body cavity and place them into the petri dish containing M2 medium for subsequent oocytes isolation (see section 3).
- Make an incision on the skin covering the abdominal wall followed by an incision in the peritoneum by using sterile dissection scissors. Open the incision to expose the abdomen, move the guts on one side using sterile tweezers and localize the uterine tubes, forming a V-shape starting from the bladder.
- Observe the ovaries located below the kidneys. Hold each tube with sterile tweezer and make a small incision at the bottom of the ovaries to release them. Transfer both ovaries at the bottom of a petri dish containing pre-equilibrated medium.
2. Hormone Preparation
Dilute PMSG (available in different stock concentrations) with sterile isotonic saline solution to obtain a working concentration of 2.5 I.U. hormone per 0.1 ml for an appropriate injection volume. NOTE: Hormone solutions must be aliquoted in 0.5 ml tubes and stored at -20 °C until the day of use. Do not keep diluted aliquots longer than three months at -20 °C. Once thawed, if serial injections are performed, store the aliquots of hormone solutions at +4 °C and use them within 4 days.
3. Antral Oocytes Isolation
For proper oocytes isolation:
Equilibrate M2 medium13 (M2) at 5% CO2 and 37 °C for at least 1 hr before starting the oocytes isolation procedure. Prepare two petri dishes (35 mm x 10 mm), one with 1 ml of M2 for ovary puncturing and the second one with small drops (20 µl) of M2 covered with mineral oil (~1.5 ml) for oocytes transfer after isolation from the ovary (see 3.4-3.6). Keep both petri dishes in the incubator at 5% CO2 and 37 °C until ready to be used.
Use a stereomicroscope during oocytes isolation procedures.
- Prepare thin mouth glass pipettes to collect the oocytes by stretching glass Pasteur pipettes kept horizontally (holding the ends of the pipette with both hands), over a vertical flame coming from the Bunsen burner (Figure 1).
- Once the glass begins to melt, take the pipet out of the flame and perform a quick pulling movement to obtain the desired pipette. Firmly cut the excess glass close to the narrow point of the pipette with fingernails to adjust the length and the diameter of the internal capillary. Ensure that the thin capillary has an internal diameter of ~100 µm, slightly greater than the oocyte's diameter (~80 µm).
- To mouth pipette, connect one end of the aspirator tube to the pulled pipette by using an appropriate tip and place the other end in the mouth, securing it with the teeth.
Collect the ovaries using sterile tweezers and deposit them at the bottom of a cell culture petri dish (35 mm x 10 mm). Cover them with 1 ml of M2 previously equilibrated at 37 °C and 5% CO2 (Figure 2).
Gently puncture the antral follicles of the ovaries with a sterile insulin syringe needle to release antral oocytes in M2 (Figure 3).
Gently mouth pipet the oocytes through a narrow Pasteur pipette to remove follicle cells and any other possible tissue debris (Figure 4A) and then settle them at the bottom of small drops (20 µl) of M2 covered with mineral oil suitable for embryo culture (previously prepared; Figure 4B). NOTE: It is important to remove all the cumulus cells attached to the zona pellucida of the oocytes by continuous pipetting through several and different drops of M2. Perform this step as quick as possible to avoid alterations in the pH of the medium.
4. Antral Oocytes Staining with Hoechst33342
NOTE: Antral oocytes can be distinguished in SN (Surrounded Nucleolus), NSN (Not Surrounded Nucleolus) or in any other intermediate form, based on their chromatin organization after staining with supravital concentration of Hoechst33342 (or any other markers specific for bases A-T)14. In fact, the SN oocytes show a Hoechst-positive ring around the nucleolus while the NSN do not, hence its name.
For proper oocytes staining:
Transfer the oocytes using a mouth pipette into drops of Hoechst33342 (20 µl) diluted in M2 at the supravital concentration of 50 ng/µl for 10 min in dark.
After the incubation with Hoechst33342, transfer each single oocyte with a mouth pipette at the bottom of small drops (5 µl) of M2 covered with mineral oil, for sorting based on chromatin organization (SN or NSN). Use any fluorescence microscope equipped with Hoechst33342 filter (emission wavelength 486 nm) to visualize the chromatin configuration of the oocytes. NOTE: Depending on the type of fluorescence microscope used, it may become imperative to use glass bottom petri dish for image acquisition. The confocal microscopy Olympus Fluoview Fv10i when equipped with a specimen holder for 35 mm dish requires only glass bottom-dishes for image acquisition, with a 0.17 mm standard thickness and a range from 0.13 to 0.21 mm acceptable.
Excite the oocytes for no longer than 5-10 sec, likely enough time to visualize the presence or the absence of a ring around the nucleolus (Figure 5).
After sorting, culture the antral SN and NSN oocytes in M2 at 5% CO2 and 37 °C to induce in vitro maturation or keep them in appropriate buffers for molecular or cellular analyses.
Representative Results
The following pictures show the method used to isolate antral oocytes from mouse ovaries, starting from pipettes preparation to the oocytes' sorting by means of Hoechst33342. This method is quite simple and can be performed in any laboratory equipped with a CO2 incubator, a stereomicroscope and a fluorescence microscope equipped with the correct filter for the observation of the Hoechst33342. Figure 1 shows the initial step of pipette preparation: as the glass begins to melt and become soft, both ends of the pipette must be pulled apart and, at the same time, the pipette must be taken out of the flame. Alternatively, glass capillary can be pulled in the same way or using a micropipette puller, if properly calibrated. This is a crucial step because it is very important to have the right mouth pipette with the right capillary diameter: if it is too thin, oocytes may be fragmented inside the small capillary and die in few minutes, if it is too big, a proper suction of the medium containing the oocytes can be lost when moving the oocytes, for example, from one drop to another. Figures 2 and 3 show the mechanical isolation of antral oocytes from antral follicles using a sterile insulin needle. It is quite easy to recognize these follicles because they are characterized by a cavity or antrum containing a fluid, that is the medium in which the granulosa cells and oocyte are found and through which different molecules can be exchanged to and from the external environment. Once isolated, antral oocytes must be transferred from one drop to another of M2 medium to remove somatic cells surrounding the oocytes, as shown in Figures 4A and B. Once "denuded", antral oocytes are ready for classification. Figure 5 shows the last step of this protocol, that is the staining of the isolated antral oocytes with Hoechst33342 for a quick sorting based on their chromatin organization. This marker stains living cells much more rapidly than DAPI, for example, and must be used at supravital concentration (50 ng/µl) to preserve cell viability. Cell staining for chromatin organization is probably the most critical step of this protocol: if the oocytes are excited too long they can be damaged and become unusable for future essays. Overall, if applied to mouse oocytes, this is a simple method that do not require specific precautions (except for those indicated in the method section). All the steps can be performed at RT.
 Figure 1.
Preparation of a mouth pipette using glass Pasteur pipettes and a Bunsen burner.
Please click here to view a larger version of this figure.
Figure 1.
Preparation of a mouth pipette using glass Pasteur pipettes and a Bunsen burner.
Please click here to view a larger version of this figure.
 Figure 2. Intact ovaries in petri dish filled with pre-warmed and equilibrated M2 medium.
Please click here to view a larger version of this figure.
Figure 2. Intact ovaries in petri dish filled with pre-warmed and equilibrated M2 medium.
Please click here to view a larger version of this figure.
 Figure 3. Initial isolation of antral oocytes from antral follicles by means of a sterile needle.
Please click here to view a larger version of this figure.
Figure 3. Initial isolation of antral oocytes from antral follicles by means of a sterile needle.
Please click here to view a larger version of this figure.
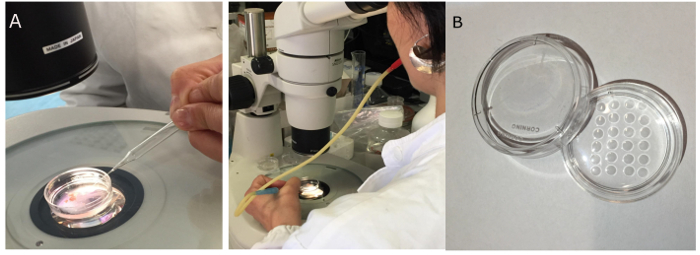 Figure 4. Transfer of the freshly isolated oocytes with a mouth pipette from the petri dish (A) to a different petri dish containing small drops of pre-warmed and equilibrated M2medium (B) Please click here to view a larger version of this figure.
Figure 4. Transfer of the freshly isolated oocytes with a mouth pipette from the petri dish (A) to a different petri dish containing small drops of pre-warmed and equilibrated M2medium (B) Please click here to view a larger version of this figure.
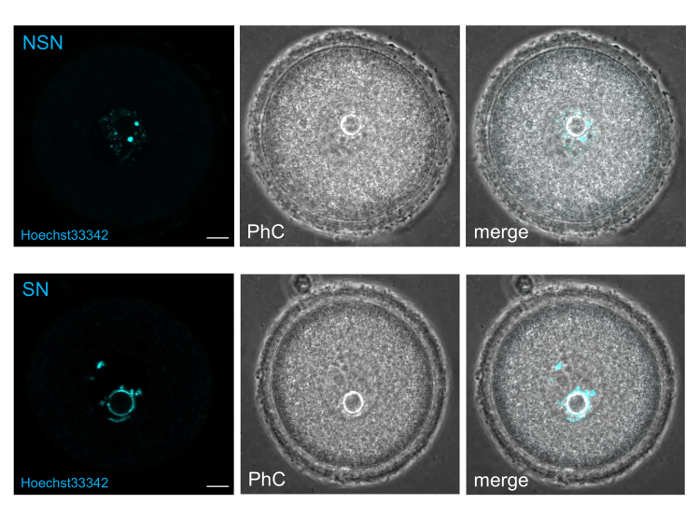 Figure 5. Oocytes stained and sorted with Hoechst33342. Single images of NSN and SN oocytes taken at the confocal microscope Olympus Fluoview Fv10i (magnification 60X; bar: 10 µm; PhC: phase contrast). Please click here to view a larger version of this figure.
Figure 5. Oocytes stained and sorted with Hoechst33342. Single images of NSN and SN oocytes taken at the confocal microscope Olympus Fluoview Fv10i (magnification 60X; bar: 10 µm; PhC: phase contrast). Please click here to view a larger version of this figure.
Discussion
This protocol describes the method used to isolate and classify mouse antral GV oocytes. There are no particular critical steps to underline, with few exceptions. First: this procedure should be performed as quickly as possible to: a) maintain the vitality of the oocytes and b) avoid pH changes in the medium used. Second: oocytes should be briefly excited (5-10 sec) to Hoechst33342 to avoid chromatin damage and the subsequent death of the oocytes, especially if IVM and IVF are the following steps. To date, this is the quickest way to sort SN and NSN mouse antral oocytes.
Hochest33342 has been proven to be an efficient way to characterize SN and NSN oocytes in female mice (and other mammalian laboratory models) but it can't be, unfortunately, used to sort antral oocytes in humans for fertility purposes.
However, since in vitro maturation of immature oocytes is routinely used in humans too, it would be extremely important to find a non-invasive way to distinguish the "good" oocytes (i.e., SN) from the "bad" ones (i.e., NSN) to increase the success of in vitro fertilization and subsequent prospective embryos development. Besides, the isolation and culture of SN oocytes from ovarian tissue could be used for fertility preservation in women undergoing cancer treatments or suffering with ovarian pathologies.
By exploiting this natural phenomenon, important data could be provided for the understanding of naturally occurring embryo preimplantation losses which speaks not only to those interested in the basic biology of oocyte development but also to human and animal IVF clinicians.
The implications of this technique for human health, biopolitics of embryos and animal reproduction are significant15. Being able to identify and eliminate "life-incompatible" oocytes would greatly raise the effective level of IVF techniques, leading to cutting-edge solutions for important unmet clinical needs.
In fact, the international scientific community of gamete biologists could contribute to seek the critical improvements needed to apply this protocol to human IVF procedures. Even though the protocol can't be applied to humans (for clear safety reasons), it still keeps a tremendous capacity to improve the knowledge of the link between chromatin organization (i.e., genome architecture) and gene expression during the final stages of antral maturation, one of the hot topics in human reproduction.
Disclosures
The authors have nothing to disclose
Acknowledgments
M.M. and C.A.R. acknowledge the financial support of the Ministero della Salute, Ricerca Finalizzata/giovani ricercatori anno 2009 of the Fondazione IRCCS Ospedale San Matteo, Pavia (I). M.M and C.A.R. wish to thank Marianna Longo for helping with the images preparation.
References
- Parfenov V, Potchukalina G, Dudina L, Kostyuchek D, Gruzova M. Human antral follicles: oocyte nucleus and the karyosphere formation (electron microscopic and autoradiographic data) Gamete Res. 1989;22(2):219–231. doi: 10.1002/mrd.1120220209. [DOI] [PubMed] [Google Scholar]
- Anderiesz C, Ferraretti AP, Magli C, Fiorentino A, Fortini D, Gianaroli L, Jones GM, Trounson AO. Effect of recombinant human gonadotropins on human, bovine and murine oocyte meiosis, fertilization and embryonic development in vitro. Human Reprod. 2000;15(5):1140–1148. doi: 10.1093/humrep/15.5.1140. [DOI] [PubMed] [Google Scholar]
- Mattson BA, Albertini D. Oogenesis: Chromatin and microtubule dynamics during meiotic prophase. Mol. Reprod. Dev. 1990;25(4):374–383. doi: 10.1002/mrd.1080250411. [DOI] [PubMed] [Google Scholar]
- Debey P, Szollosi MS, Szollosi D, Vautier D, Girousse A, Besomber D. Competent mouse oocytes isolated from antral follicles exhibit different chromatin organization and follow different maturation dynamics. Mol. Reprod. Dev. 1993;36(1):59–74. doi: 10.1002/mrd.1080360110. [DOI] [PubMed] [Google Scholar]
- Zuccotti M, Piccinelli A, Giorgi Rossi P, Garagna S, Redi CA. Chromatin organisation during mouse oocyte growth. Mol. Reprod. Dev. 1995;41(1-2):479–485. doi: 10.1002/mrd.1080410410. [DOI] [PubMed] [Google Scholar]
- Wu YG, et al. Selection of oocytes for in vitro maturation by brilliant cresyl blue staining: a study using the mouse model. Cell Res. 2007;17:722–731. doi: 10.1038/cr.2007.66. [DOI] [PubMed] [Google Scholar]
- Opiela Y, Kątska-Książkiewicz L. The utility of Brilliant Cresyl Blue (BCB) staining of mammalian oocytes used for in vitro embryo production (IVP) Reproductive Biol. 2013;13:177–183. doi: 10.1016/j.repbio.2013.07.004. [DOI] [PubMed] [Google Scholar]
- Labrecque R, Sirard MA. The study of mammalian oocyte competence by transcriptome analysis: progress and challenges. Mol. Human Reprod. 2014;20(2):103–116. doi: 10.1093/molehr/gat082. [DOI] [PubMed] [Google Scholar]
- Monti M, Zanoni M, Calligaro A, Ko MS, Mauri PL, Redi CA. Developmental arrest and mouse antral Not-Surrouded Nucleolus oocyte. Biol. Reprod. 2013;88(1):1–7. doi: 10.1095/biolreprod.112.103887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yurttas P, et al. Role for PADI6 and the cytoplasmic lattices in ribosomal storage in oocytes and translational control in the early mouse embryo. Development. 2008;135(15):2627–2636. doi: 10.1242/dev.016329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li L, Baibakov B, Dean J. A subcortical maternal complex essential for preimplantation mouse embryogenesis. Dev. Cell. 2008;15(3):416–425. doi: 10.1016/j.devcel.2008.07.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ami D, et al. FT-IR spectra signatures of mouse antral oocytes: molecular markers of oocyte maturation and developmental competence. Biochim. Biophys. Acta. 2011;1813(6):1220–1222. doi: 10.1016/j.bbamcr.2011.03.009. [DOI] [PubMed] [Google Scholar]
- Fulton BP, Whittingam DG. Activation of mammalian oocytes by intercellular injection of calcium. Nature. 1978;273(5658):149–151. doi: 10.1038/273149a0. [DOI] [PubMed] [Google Scholar]
- Luttmer SJ, Longo FJ. Examination of living and fixed gametes and early embryos stained with supravital fluorochromes (Hoechst 33342 and 3,3′-dihexyloxacarbocyanine iodide) Gamete Res. 1986;15(3):267–283. [Google Scholar]
- Monti M, Redi CA. The biopolitics of frozen embryos. Int. J. Dev. Biol. 2011;55:243–247. doi: 10.1387/ijdb.103281mm. [DOI] [PubMed] [Google Scholar]