

Abstract
Clinical infections caused by Acinetobacter spp. have increasing public health concerns because of their global occurrence and ability to acquire multidrug resistance. Acinetobacter calcoaceticus–Acinetobacter baumannii (ACB) complex encompasses A. calcoaceticus, A. baumannii, A. pittii (formerly genomic species 3), and A nosocomial (formerly genomic species 13TU), which are predominantly responsible for clinical pathogenesis in the Acinetobacter genus.
In our previous study, a putative novel species isolated from 385 non-A. baumannii spp. strains based on the rpoB gene phylogenetic tree was reported. Here, the putative novel species was identified as A. seifertii based on the whole-genome phylogenetic tree. A. seifertii was recognized as a novel member of the ACB complex and close to A. baumannii and A. nosocomials. Furthermore, we studied the characteristics of 10 A. seifertii isolates, which were distributed widely in 6 provinces in China and mainly caused infections in the elderly or children. To define the taxonomic status and characteristics, the biochemical reactions, antimicrobial susceptibility testing, pulsed field gel electrophoresis (PFGE), multilocus sequence typing (MLST), and whole-genome sequence analysis were performed.
The phenotypic characteristics failed to distinguish A. serfertii from other species in the ACB complex. Most of the A. seifertii isolates were susceptible to antibiotics commonly used for nosocomial Acinetobacter spp. infections, but one isolate (strain A362) was resistant to ampicillin/sulbactam, ceftazidime and amikacin. The different patterns of MLST and PFGE suggested that the 10 isolates were not identical and lacked clonal relatedness.
Our study reported for the first time the molecular epidemiological and genomic features of widely disseminated A. seifertii in China. These observations could enrich the knowledge of infections caused by non-A. baumannii and may provide a scientific basis for future clinical treatment.
INTRODUCTION
The genus Acinetobacter is widely distributed in nature and commonly occurs in soil. During the past decades, it has been increasingly recognized as a significant pathogen of nosocomial infections, including ventilator-associated pneumonia, bloodstream infections, skin and soft-tissue infections, meningitis, and urinary tract infections.1–3 Most of the studies of the etiological organisms concentrate on A. baumannii, which is notorious for its multidrug resistance or even pan-drug resistance.2,4,5 The insufficient knowledge in databases and the intrinsic intragenus similarity make current phenotypic tests difficult to distinguish different Acinetobacter species, especially between members of the Acinetobacter calcoaceticus–Acinetobacter baumannii (ACB) complex (including A. baumannii, A. calcoaceticus, A. nosocomials, and A. pittii).6,7A. seifertii was recognized as a novel member of the ACB complex by Nemec et al.8 It was formerly known as gen sp “close to 13TU,”9 which was isolated from human clinical specimens and the environment in different countries and areas. Further, study of A. seifertii is necessarily required.
In this study, we report on the detection of A. seifertii in China, mainly using the whole-genome sequence and molecular typing methods to clarify the phylogenetic relationships with other Acinetobacter species and molecular epidemiology characteristics.
MATERIALS AND METHODS
Bacterial Strains and Phenotypic Characteristics
In our previous study,10 we reported a putative, novel Acinetobacter species: A total of 385 non-A. baumannii isolates were collected from 27 provinces in China from January 2009 to September 2010.11 By 16S rRNA and RNA polymerase β-subunit gene (rpoB) sequencing, we found that the most common species was A. pittii (49.09%).10 Nevertheless, 10 isolates constituted a novel cluster and could not be assigned into any previously known species (GenBank accession numbers: KF982810-KF982820). Here, we chose the 10 isolates to study further. The colonies were observed after 18 to 24 h at 37°C on tryptic soy agar (Oxoid). Utilization tests were evaluated by VITEK 2 system (Sysmex-bioMérieux, Marcy l’Etoile, France).
Antimicrobial Susceptibility Testing
The antibiotic susceptibility profile of all isolates to different antibiotics, including ampicillin/sulbactam, ceftazidime, imipenem, colistin, amikacin, tigecycline, tetracycline, ciprofloxacin, aztreonam, and fosfomycin, was determined by the Etest (AB bioMérieux, Solna, Sweden), and the interpretation was according to the CLSI 2015 guidelines.12 The breakpoints for Enterobacteriaceae of the European Committee on Antimicrobial Susceptibility Testing were used for tigecycline and aztreonam. (http://www.eucast.org/). Escherichia coli ATCC 25922 was used as a reference strain for quality control.
Pulsed Field Gel Electrophoresis
Genomic DNA was digested by the restriction enzyme ApaI. The conditions were 22 h at 6 V/cm and 14°C, with a pulse angle of 120 degree, and pulse time from 5 to 35 s with a CHEF-Mapper XA pulsed field gel electrophoresis (PFGE) system (Bio-Rad, Hercules, CA, USA). Salmonella enterica serotype Braenderup H9812 was used as the size marker.13 The restriction patterns were analyzed with BioNumerics 7.0 (Applied Maths BVBA, Sint-Martens-Latem, Belgium). Interpretation was performed according to Tenover's criteria.14
Multilocus Sequence Typing
Multilocus sequence typing (MLST) following the Oxford scheme was performed as described by Bartual et al.15 The internal fragments of seven housekeeping genes, including gltA, gyrB, gdhB, recA, cpn60, gpi, and rpoD, were PCR-amplified. PCR reactions were designed as follows: predenaturation at 94°C for 5 min, followed by 35 cycles of denaturation at 94°C for 1 min, annealing at 55°C for 30 s, and extension at 72°C for 60 s. Sequence types (STs) were assigned using the PubMLST database (http://pubmlst.org/abaumannii/).
Whole-Genome Sequence and Phylogenetic Analysis
We chose 3 isolates (strains A354, A360, and A362) for whole-genome sequencing. Total DNA was extracted and sequenced using next-generation sequencing technology (either Illumina HiSeq2000TM with 2 × 100 bp paired-end reads or Illumina MiSeqTM with 2 × 300 bp paired-end reads). The derived short reads were assembled into contigs using CLC Genomics Workbench 8.0 (CLC bio, Denmark). Acquired resistance genes and virulence genes were screened using the ResFinder 2.1 tool on the CGE server (https://cge.cbs.dtu.dk/services/ResFinder/).
Similarities of protein-coding sequences were determined using the BLASTP program of the NCBI Basic Local Alignment Search Tool (BLAST). For a coding sequence to be considered homologous, the protein identity had to be >80%, e-value smaller than 1e−10, and aligned length >80% of the gene sequence. Phylogenetic reconstruction was performed using the core genes were shared by the genome of the compared Acinetobacter spp. strains with the MEGA 5.0 Maximum-likelihood program and BacWGSTdb platform.16,17 The similarity of protein-encoding genes and average amino acid among A. seifertii, A. baumannii, and A. nosocomials was converted to a Venn diagram using R (http://www.r-project.org/), which shows the number of the genes in the specific strains.
Nucleotide Sequence Accession Numbers
The nucleotide sequence data reported here have been submitted to the GenBank database with the assigned accession number: LFZQ01 (strain A354), LFZR01 (strain A360), and LFZS01 (strain A362).
RESULTS AND DISCUSSION
Whole-Genome Sequence Analyses
According to our previous work, there were 10 isolates clustered in the same branch in the rpoB-based phylogenetic tree among a total of 385 non-A. baumannii isolates.10 The rpoB gene sequence of the 10 isolates were the closest matched with A. seifertii, indicating they probably belong to this species. Thus, the 3 isolates (strains A354, A360, and A362) were chosen for whole-genome sequencing.
Genome comparison revealed that a total of 1941 core genes were shared by the genome of the compared Acinetobacter spp. strains (Figure 1 A). The phylogenetic tree based on the shared core genes showed that the 3 isolates (strain A354, A360, and A362) constituted the same branch with A. seifertii NIPH 973T (formally Acinetobacter gen. sp. “close to 13TU”), which is relatively closer to A. baumannii and A. nosocomials but distant from A. calcoaceticus (Figure 1A). These 3 isolates were clustered with each other and constituted a cohesive group. According to the whole-genome-based phylogenetic tree, we concluded that these isolates were identified as A. seifertii.
FIGURE 1.
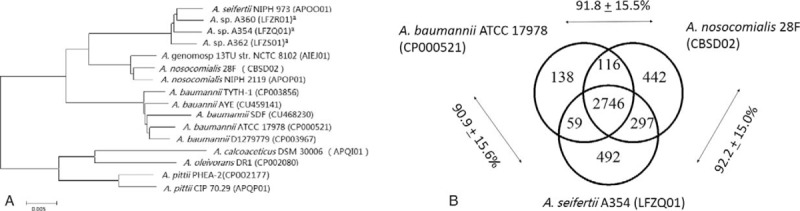
Phylogenetic relationship between 3 strains of putative novel species and other Acinetobacter spp. (A) Whole-genome phylogenetic tree of the 3 strains and other sequenced Acinetobacter spp. genomes. aIndicates the 3 strains that were chosen for whole-genome sequencing in this study. (B) Genome comparison between A. seifertii, A. baumannii, and A. nosocomials. The numbers in the Venn diagram represent the shared genes between the compared strains. Data outside the Venn diagram represent average amino acid identity between the adjacent strains ± standard deviation.
A deeper look inside the ACB complex showed that A. seifertii was highly similar to A. baumannii and A. nosocomials. For example, 83.01% of protein-encoding genes of A. seifertii were shared with A. baumannii and A. nosocomials, with the average amino acid identity over 90% (Figure 1B). The genomes of A354 and A360 isolates both contained only one antimicrobial resistance gene blaADC-25. In contrast, A362 contained much more resistance genes, such as sul2, aph(3’)-Via, sul1, blaPER-1, aacA4, aac(6’)Ib-cr, msr(E), mph(E), aac(3)-IId, floR, and ARR-3. This probably explains why this isolate exhibits a much higher MICs of ampicillin/sulbactam, ceftazidime, amikacin, aztreonam, and fosfomycin than other isolates. Most of these resistance genes were also commonly present in multidrug-resistant A. baumannii. Thus, it is reasonable to hypothesize that A. seifertii and A. baumannii share the common repertoire of resistance genes to survive in the nosocomial environment.18 All 3 isolates carried most of A. baumannii known virulence genes, including ompA, pgaABCD, csu pili, lpsB, pmrB, pbpG, eps, and ptk; so the pathogenicity of A. seifertii is likely equal to A. baumannii.
Epidemiological and Clinical Features
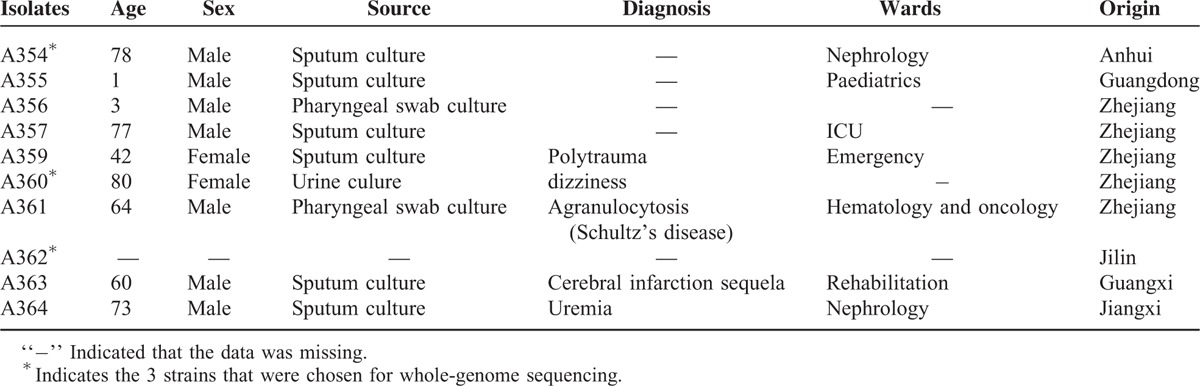
Ten A. seifertii isolates were distributed in 6 provinces in China, which are geographically distant places. The patients were mostly the elderly and children in several wards. The clinical information of 10 A. seifertii isolates is shown in Table 1.
TABLE 1.
The Clinical Features of 10 A. seifertii Isolates
Phenotypic Characteristics
The phenotypic characteristics of A. seifertii were not significantly different from other Acinetobacter species, especially the ACB complex. The colonies were 1 to 1.5 mm in diameter, circular, convex, smooth, and slightly opaque with entire margins. Growth occurred in brain-heart infusion (Oxoid) at temperatures ranging from 15°C to 41°C, and the optimum temperature is 37°C. The optimum pH and NaCl concentration was 5.5 to 9 and 0 to 4%, respectively. The isolates of A. seifertii were Gram-negative, strictly aerobic, oxidase-negative, catalase-positive, and nonmotile coccobacilli. Overall, A. seifertii cannot be reliably distinguished from the ACB complex merely based on phenotypic tests, and therefore it should be a member of the ACB complex; the deposited strain is A354 (=CGMCC 1.15326 = KCTC 42723).
Antimicrobial Susceptibility Testing
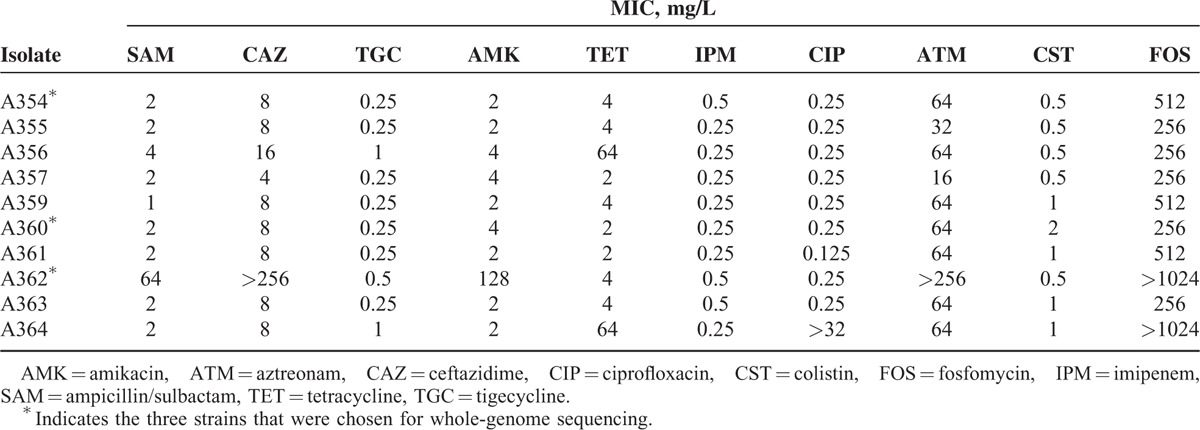
The antibiotic resistance profiles of A. seifertii were determined (Table 2). They were susceptible to ciprofloxacin, imipenem, tigecycline, and colistin. All but one isolate showed resistance to fosfomycin (256 ≥ 1024 mg/mL) and aztreonam (32 ≥ 256 mg/mL). Only one isolate (strain A362) was resistant to ampicillin/sulbactam, ceftazidime, and amikacin, and two isolates were resistant to tetracycline.
TABLE 2.
The Minimal Inhibitory Concentration (MIC) of 10 A. seifertii Isolates
Molecular Epidemiology Characteristics
We further identified intra-species genetic diversity by MLST. The 10 isolates exhibited 10 different allele combinations (Figure 2A), and none of them differed from other isolates by <2 alleles. PFGE was also performed. The PFGE profiles presented that 10 isolates were not identical and lacked clonal relatedness (Figure 2B). These typing results were consistent with the clinical information that the 10 isolates were collected from different areas; it also suggested that A. seifertii was a widely distributed species, rather than through population movement with an identical clone.
FIGURE 2.
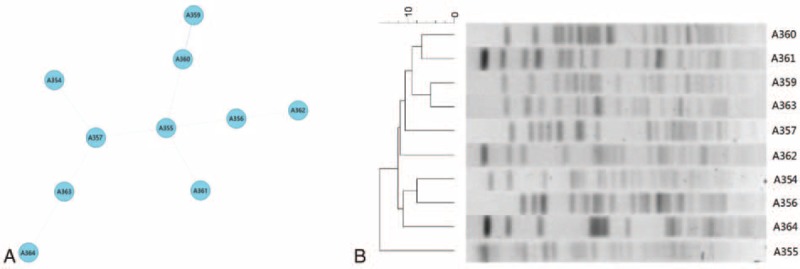
Molecular epidemiology characteristics of 10 A. seifertii isolates. (A) Minimum spanning tree analysis of 10 A. seifertii isolates based on multilocus sequence typing data. Each circle represents independent sequence type (ST). The lines connecting the circles indicate the relationship between different STs. Different types of lines represent a difference in 3 alleles (dashed lines) and ≥4 alleles (dotted lines). (B) Pulsed field gel electrophoresis analysis for the 10 A. seifertii isolates (variation within 3 bands indicates the same clone).
In conclusion, the putative novel species in our previous study was identified as A. seifertii. The whole-genome sequence analyses represented that A. seifertii shares some common resistant genes with A. baumannii to benefit their survival in the nosocomial environment, and the mechanisms of acquiring resistant genes need further study. The clinical information and molecular epidemiology analyses highlighted that A. seifertii was distributed geographically with different clones. The further study of A. seifertii could enrich the knowledge of infection by non-A. baumannii and provide a scientific basis for future clinical treatment. In addition, detailed virulence and epidemiology of A. seifertii require further investigation.
Acknowledgments
We are extremely thankful to Prof Min Wu, Dr Weiyan Zhang, Mr Jing Hu, and Mr Shuaibo Han at College of Life Sciences, Zhejiang University for giving technology assistance and advice. This work was supported by National Natural Science Foundation of China (81230039, 81378158, and 81401698).
Footnotes
Abbreviations: A. baumannii = Acinetobacter baumannii, A. nosocomials = Acinetobacter nosocomials, A. seifertii = Acinetobacter seifertii, ACB = Acinetobacter calcoaceticus–Acinetobacter baumannii, Acinetobacter gen. sp. = Acinetobacter genomic species, AMK = amikacin, ATM = aztreonam, CAZ = ceftazidime, CGMCC = China General Microbiological Culture Collection Center, CIP = ciprofloxacin, CST = colistin, FOS = fosfomycin, IPM = imipenem, KCTC = Korean Collection for Type Cultures, MIC = minimal inhibitory concentration, MLST = multilocus sequence typing, PFGE = pulsed field gel electrophoresis, SAM = ampicillin/sulbactam, TET = tetracycline, TGC = tigecycline.
The authors declare no conflicts of interest.
Author's contributions: YY made substantial contributions to the design of the study, and conducted data generation, analyzed and interpreted the data, prepared the manuscript draft, and approved the final version of the manuscript; JW collected the clinical data in this study, interpreted the clinical data, and approved the final version of the manuscript; YF conceptualized the study, interpreted the data, revised the intermediate version of this manuscript, and approved the final version of the manuscript; ZR analyzed all the data of the work, revised the intermediate version of this manuscript, and approved the final version of the manuscript; YY suggested critical points in the study, analyzed and interpreted the data, and revised and approved the final version of the manuscript.
REFERENCES
- 1.Peleg AY, Seifert H, Paterson DL. Acinetobacter baumannii: emergence of a successful pathogen. Clin Microbiol Rev 2008; 21:538–582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Dijkshoorn L, Nemec A, Seifert H. An increasing threat in hospitals: multidrug-resistant Acinetobacter baumannii. Nat Rev Microbiol 2007; 5:939–951. [DOI] [PubMed] [Google Scholar]
- 3.Bergogne-Berezin E, Towner KJ. Acinetobacter spp. as nosocomial pathogens: microbiological, clinical, and epidemiological features. Clin Microbiol Rev 1996; 9:148–165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Biedenbach DJ, Bouchillon SK, Hoban DJ, et al. Antimicrobial susceptibility and extended-spectrum beta-lactamase rates in aerobic gram-negative bacteria causing intra-abdominal infections in Vietnam: report from the Study for Monitoring Antimicrobial Resistance Trends (SMART 2009-2011). Diagnost Microbiol Infect Dis 2014; 79:463–467. [DOI] [PubMed] [Google Scholar]
- 5.Kuo SC, Chang SC, Wang HY, et al. Emergence of extensively drug-resistant Acinetobacter baumannii complex over 10 years: nationwide data from the Taiwan Surveillance of Antimicrobial Resistance (TSAR) program. BMC Infect Dis 2012; 12:200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Nemec A, Krizova L, Maixnerova M, et al. Genotypic and phenotypic characterization of the Acinetobacter calcoaceticus-Acinetobacter baumannii complex with the proposal of Acinetobacter pittii sp. nov. (formerly Acinetobacter genomic species 3) and Acinetobacter nosocomialis sp. nov. (formerly Acinetobacter genomic species 13TU). Res Microbiol 2011; 162:393–404. [DOI] [PubMed] [Google Scholar]
- 7.Lee MJ, Jang SJ, Li XM, et al. Comparison of rpoB gene sequencing, 16S rRNA gene sequencing, gyrB multiplex PCR, and the VITEK2 system for identification of Acinetobacter clinical isolates. Diagnost Microbiol Infect Dis 2014; 78:29–34. [DOI] [PubMed] [Google Scholar]
- 8.Nemec A, Krizova L, Maixnerova M, et al. Acinetobacter seifertii sp. nov., a member of the Acinetobacter calcoaceticus-Acinetobacter baumannii complex isolated from human clinical specimens. Int J SystEvol Microbiol 2015; 65 (Pt 3):934–942. [DOI] [PubMed] [Google Scholar]
- 9.Gerner-Smidt P, Tjernberg I. Acinetobacter in Denmark: II. Molecular studies of the Acinetobacter calcoaceticus-Acinetobacter baumannii complex. APMIS 1993; 101:826–832. [PubMed] [Google Scholar]
- 10.Wang J, Ruan Z, Feng Y, et al. Species distribution of clinical Acinetobacter isolates revealed by different identification techniques. PLoS One 2014; 9:e104882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ruan Z, Chen Y, Jiang Y, et al. Wide distribution of CC92 carbapenem-resistant and OXA-23-producing Acinetobacter baumannii in multiple provinces of China. Int J Antimicrob Agent 2013; 42:322–328. [DOI] [PubMed] [Google Scholar]
- 12.Tan SY, Chua SL, Liu Y, et al. Comparative genomic analysis of rapid evolution of an extreme-drug-resistant Acinetobacter baumannii clone. Genome Biol Evol 2013; 5:807–818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hunter SB, Vauterin P, Lambert-Fair MA, et al. Establishment of a universal size standard strain for use with the PulseNet standardized pulsed-field gel electrophoresis protocols: converting the national databases to the new size standard. J Clin Microbiol 2005; 43:1045–1050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Tenover FC, Arbeit RD, Goering RV, et al. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J Clin Microbiol 1995; 33:2233–2239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Bartual SG, Seifert H, Hippler C, et al. Development of a multilocus sequence typing scheme for characterization of clinical isolates of Acinetobacter baumannii. J Clin Microbiol 2005; 43:4382–4390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ruan Z, Feng Y. BacWGSTdb, a database for genotyping and source tracking bacterial pathogens. Nucleic Acid Res 2016; 44 (D1):D682–D687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Tamura K, Peterson D, Peterson N, et al. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol 2011; 28:2731–2739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Poirel L, Bonnin RA, Nordmann P. Genetic basis of antibiotic resistance in pathogenic Acinetobacter species. IUBMB Life 2011; 63:1061–1067. [DOI] [PubMed] [Google Scholar]