

Abstract
Rice is a facultative short-day plant (SDP), and the regulatory pathways for flowering time are conserved, but functionally modified, in Arabidopsis and rice. Heading date 1 (Hd1), an ortholog of Arabidopsis CONSTANS (CO), is a key regulator that suppresses flowering under long-day conditions (LDs), but promotes flowering under short-day conditions (SDs) by influencing the expression of the florigen gene Heading date 3a (Hd3a). Another key regulator, Early heading date 1 (Ehd1), is an evolutionarily unique gene with no orthologs in Arabidopsis, which acts as a flowering activator under both SD and LD by promoting the rice florigen genes Hd3a and RICE FLOWERING LOCUST 1 (RFT1). Here, we report the isolation and characterization of the flowering regulator Heading Date Repressor1 (HDR1) in rice. The hdr1 mutant exhibits an early flowering phenotype under natural LD in a paddy field in Beijing, China (39°54'N, 116°23'E), as well as under LD but not SD in a growth chamber, indicating that HDR1 may functionally regulate flowering time via the photoperiod-dependent pathway. HDR1 encodes a nuclear protein that is most active in leaves and floral organs and exhibits a typical diurnal expression pattern. We determined that HDR1 is a novel suppressor of flowering that upregulates Hd1 and downregulates Ehd1, leading to the downregulation of Hd3a and RFT1 under LDs. We have further identified an HDR1-interacting kinase, OsK4, another suppressor of rice flowering under LDs. OsK4 acts similarly to HDR1, suppressing flowering by upregulating Hd1 and downregulating Ehd1 under LDs, and OsK4 can phosphorylate HD1 with HDR1 presents. These results collectively reveal the transcriptional regulators of Hd1 for the day-length-dependent control of flowering time in rice.
Author Summary
In rice, flowering time affects the potential yield, the growing season and regional adaptability. Change in day length is a key seasonal cue for regulating flowering time in rice, a facultative short-day (SD) plant. The photoperiodic pathway of rice contains the evolutionarily conserved Hd1-Hd3a module, which is homologous to the CO-FT module in the long-day (LD) plant Arabidopsis. In this work, we cloned a novel gene, HDR1, that activates Hd1 and represses Ehd1, thereby down-regulating the florigen genes Hd3a and RFT1 to postpone rice flowering. A protein associated with HDR1, OsK4, was also identified, and the resulting complex can interact with HD1 to phosphorylate HD1. We conclude that HDR1 is a novel transcriptional regulator of Hd1 that plays a crucial role in regulating flowering time via the photoperiodic pathway in rice.
Introduction
The ability of plants to reproduce during the appropriate season enables them to adapt to environmental changes in day length and temperature and requires precise monitoring of environmental and endogenous signals [1–5]. These external and internal signals comprise a complex regulatory network that includes the aging, autonomous, vernalization, photoperiod, gibberellin, and ambient temperature pathways [6,7]. This network allows plants to grow at different latitudes and altitudes and during different seasons [3,8,9]. The basis for this complex network is the control of photoperiod-dependent flowering, which involves processes such as day-length measurement in leaves, the generation of mobile signals called florigens, the transport of florigens from leaves to the shoot apex, and the perception of florigens at the shoot apical meristem to initiate floral evocation [10,11].
The molecular mechanisms that regulate plant flowering time via the photoperiodic pathway have been extensively studied in Arabidopsis, a long-day plant (LDP), and in rice, a short-day plant (SDP) [2,6,11–13]. In Arabidopsis, the GIGANTEA (GI)-CONSTANS (CO)-FLOWERING LOCUS T (FT) transcriptional regulatory pathway has been characterized. GI integrates signals from photoreceptors and the circadian clock [14] and, as a regulator of transcription, activates CO, which in turn promotes FT expression. FT functions as a florigen and coordinates with SUPPRESSOR OF OVER-EXPRESSION OF CONSTANT 1 (SOC1) to promote flowering [15,16].
Rice is a facultative SDP. Thus, short-day conditions (SDs) promote flowering in rice plants, and multiple flowering genes prevent flowering under long-day conditions (LDs) [3,13]. Phylogenetic analyses and functional studies have revealed that the rice orthologs of GI, CO, and FT are OsGI, Heading date 1 (Hd1), and Heading date 3a (Hd3a)/ RICE FLOWERING LOCUS T1 (RFT1), respectively. The pathway regulating flowering time is conserved in Arabidopsis and rice, but the functions of specific genes differ [17–22]. Similar to Arabidopsis GI, OsGI upregulates Hd1, which suppresses flowering under LDs but promotes flowering under SD by influencing the expression of Hd3a [18,19,23–25]. Hd3a is also modulated by Early heading date 1 (Ehd1), which encodes a B-type response regulator that functions independent of Hd1 [26]. Ehd1 is an evolutionarily unique gene that does not have any ortholog in Arabidopsis and that functions as a flowering activator under both SDs and LDs by promoting the rice florigensHd3a and RFT1 [20,21,26]. Several flowering regulators involved in this Ehd1 pathway have been identified [27–38]. Conversely, OsMADS51, which is controlled by OsGI, serves as a flowering activator upstream of Ehd1 under SD [27]. OsID1/Ehd2/RID1, hereafter referred to a as Early heading date 2 (Ehd2), and Early heading date 3 (Ehd3) promote flowering under both SDs and LDs by inducing the expression of Ehd1 [28–30]. OsMADS50 and OsMADS56 antagonistically regulate LD-dependent flowering in rice [31,32]. OsLFL1 (Oryza sativa LEC2 and FUSCA3 Like 1), an Ehd1 promoter-binding B3-type transcription factor, inhibits Ehd1 expression to repress rice flowering only under LD [33]. Ehd1 and Hd3a are suppressed by Ghd7 (Grain number, plant height and heading date 7) only under LDs, resulting in the late flowering of rice plants under LD [34]. DTH8 (QTL for days to heading on chromosome 8), a major quantitative trait locus (QTL) that regulates grain productivity, plant height, and heading date in rice, is also a strong repressor of Ehd1 and accounts for late flowering in rice under LDs [35,36]. Ehd3, which encodes a plant homeodomain finger-containing protein, is a critical promoter of rice flowering both under SDs and LDs by regulating the expression of Ehd1 [37]. Early heading date 4 (Ehd4) encodes a novel CCCH-type zinc-finger protein that promotes flowering only under natural LDs (NLD) [38]. Although several regulatory factors have been characterized in Hd1-dependent and Ehd1-dependent pathways in rice, whether there is an uncharacterized regulatory factor involved in these two pathways in rice is unknown.
In this study, we reported the identification of an early flowering rice mutant, and the cloning of HDR1 (Heading Date Repressor 1), a novel gene encoding a 210-amino-acid protein with a molecular weight of ~23 kD. HDR1 encodes a ubiquitous protein in plants, and the moss ortholog PpSKI encodes a putative kinase ligand that is important for the recognition of and adaptation to conditions of limited energy during plant development [39,40]. Expression and functional analyses of HDR1 revealed that it activates Hd1 and represses Ehd1, thereby downregulating the florigen genes Hd3a and RFT1 to postpone rice flowering. An HDR1-interacting kinase protein encoded by OsK4 was also identified, which functions to phosphorylate HD1 to regulate flowering time in rice.
Results
Identification and characterization of the early flowering mutant hdr1
We previously reported the development and characterization of T-DNA-mutagenized rice populations [41–43]. An average of 2.8 copies of T-DNA and 2.1 copies of newly transposed Tos17 were integrated into the rice genome during transformation process. Systematic characterization of the mutant populations led to the identification of early and late flowering mutants, and the sequences flanking the T-DNA/Tos17 insertion sites in the mutants were amplified [41–43]. One mutant, named hdr1, flowered approximately 30 days earlier than wild-type (WT, Oryza sativa var. Nipponbare) plants following growth under NLDs (Fig 1A) in a paddy field in Beijing, China (39°54'N, 116°23'E). The phenotypes of WT and hdr1 plants were assessed under natural-day field conditions (NDs) in Beijing and under LDs (14 h of light/10 h of dark) and SDs (10 h of light/14 h of dark) conditions in a controlled growth chamber. Under NDs and LDs, the hdr1 plants flowered approximately 30 days earlier than the WT plants, whereas no significant difference was observed under SDs (Fig 1B). These data suggested that HDR1 might participate in the regulation of photoperiodic flowering in rice.
Fig 1. Phenotypes of the hdr1 mutant.

(A) The early flowering phenotype of hdr1 mutant in a paddy field (right). The Nipponbare WT (left). Bar = 20 cm. (B) Flowering time of hdr1 and WT plants under different day-length conditions (n = 12). NDs, natural day conditions; SDs, short day conditions; LDs, long day conditions. (C) mRNA expression analysis of HDR1 using real-time PCR in WT and hdr1 mutant. Mean values ±s.d. were obtained from three technical replicates and two biological replicates. (D) to (H) Comparisons of plant height, tilling number, panicle length, 500-grain weight, and yield per plant between WT and hdr1 plants. The values presented are the mean ± s.d. (n = 15). Double asterisks indicate statistically significant differences in the means between WT and hdr1 according to a two-tailed Student’s t test (p<0.01), and n.s. indicates no significant difference. (I) and (J) hdr1 plants had the same leaf-emergence rate as WT under both SDs and LDs. The values presented are the mean ± s.d. (n = 12). s.d.: standard deviations.
A genetic analysis revealed that a newly transposed copy of Tos17 that was inserted into the 3'-untranslated region (UTR; 22 nt after the stop codon) of a rice gene (LOC_Os02 g55080 in the TIGR database [http://rice.plantbiology.msu.edu]) co-segregated with the early flowering phenotype of hdr1. HDR1 expression was analyzed using quantitative real-time PCR (qPCR) in WT and hdr1 plants using two pairs of primers based on the sequences of exon 2 (Q-p1 and Q-p2) and the 3’-UTR (Q-p3 and Q-p4). Primers Q-p1 and Q-p2 indicated that HDR1 expression was dramatically decreased in hdr1, and expression was only very weakly detected using primers Q-p3 and Q-p4, indicating that a low level of HDR1 mRNA was produced in the hdr1 plants (Fig 1C). The mature hdr1 plants were slightly shorter and had similar numbers of tillers (Fig 1D and 1E). No obvious difference in panicle length, 500-grain weight, or yield per plant was detected, indicating that the fertility levels of the mutant and WT plants were similar (Fig 1F–1H). Notably, the leaf-emergence rate of hdr1 was similar to that of WT under both SDs and LDs (Fig 1I and 1J), indicating that the early flowering phenotype was not caused by an increase in growth rate.
HDR1 controls photoperiodic flowering in rice
To verify that the early flowering phenotype we observed was caused by the disruption of HDR1, we performed a complementation experiment using a genomic DNA construct that included the promoter, coding, and terminator regions of HDR1 and a full-length HDR1 cDNA construct driven by the HDR1 promoter. In total, 204 positive T0 lines were obtained and confirmed by PCR (69 lines for the genomic construct and 135 lines for the full-length cDNA construct). Of these, 142 lines (44 for the genomic construct and 98 for the cDNA construct) exhibited a flowering time of 20–28 days later than hdr1, similar to WT (in a paddy field in Beijing, China. From these lines, 30 complemented T1 plants (15 for the genomic construct and 15 for the cDNA construct) were randomly selected to produce homozygote plant for further analysis; these plants exhibited a normal flowering times compared with WT plants. Flowering time was subsequently measured in WT, hdr1, and complemented plants under different photoperiods. The hdr1 plants flowered approximately 4 weeks earlier than the WT and complemented plants under LDs (14 h of light/10 h of dark) during the natural growing season (Beijing, China) (Fig 2A–2D). Slightly early flowering was also observed under SDs (Fig 2D). These data indicate that both constructs complemented the early flowering phenotype of the mutant completely (S3 Table).
Fig 2. HDR1 controls rice flowering time.

(A) to (C) Complementation of hdr1 in a paddy field. A, wild type; B, hdr1 mutant; and C, complemented transgenic plant (CP). Bar = 20 cm. (D) Flowering time in WT and Complemented plant of transgenic lines 1 and 4 under NDs, SDs and LDs. The values presented are the mean ± s.d. (n = 12). (E) The phenotype of HDR1-RNAi plants in a paddy field. WT, left; line 3, middle; and line 8, right. (F) HDR1-RNAi plants exhibited early flowering under both NDs and LDs. The values presented are the mean ± s.d. for each of 12 independent plants of transgenic lines 3 and 8. (G) Real-time PCR analysis of HDR1 mRNA levels in WT and RNAi seedlings; Transgenic lines 3 and 8 were used. Mean values ± s.d. were obtained from three technical replicates and two biological replicates. s.d.: standard deviations.
To further investigate the function of HDR1 in rice flowering time, we generated HDR1 RNA interference (RNAi) and overexpression transgenic plants. The HDR1-RNAi T0 plants exhibited an early flowering phenotype similar to that of the hdr1 mutant; in 118 PCR-confirmed T0 plants, we observed 32 lines that flowered 1 to 4 weeks earlier than WT. Furthermore, two independent T2 transgenic lines (HDR1-RNAi-3 and HDR1-RNAi-8) exhibited early flowering under NDs and LDs, along with a dramatic reduction in HDR1 expression levels (Fig 2E–2G). However, late flowering was not observed in paddy field-grown overexpression lines. Moreover, careful observation of plants under NDs, SDs and LDs revealed similar flowering times among the WT and transgenic overexpression plants (S1A–S1C Fig). Taken together, our data indicated that HDR1 is a negative regulator of flowering time under LDs in rice.
HDR1 encodes a nuclear protein with a diurnal expression pattern
HDR1 in rice encodes an ortholog of PpSKI, a putative kinase ligand in moss [39,40]. HDR1-like proteins are ubiquitous in monocots, dicots, and moss, implying an ancient origin for this protein. Subsequent phylogenetic analysis revealed three corresponding main subgroups in monocots (e.g., rice, maize, sorghum, barley, millet, and wheat), eudicots (e.g., soybean, chickpea, grape, cucumber, tomato, potato, watermelon, almond tree, rape, apple tree, and Arabidopsis), and mosses and ferns (Physcomitrella patens and Selaginellamoellendorffii, respectively). The Amborellatrichopoda homolog of HDR1 formed a single clade consistent with its unique evolutionary line of flowering plants that diverged very early (~130 million years ago) from all other extant species of flowering plants (Fig 3A). We also noted that the C-terminus of the HDR1-like family is highly conserved in different species (S2 Fig).
Fig 3. Characterization of HDR1.

(A) Phylogeny of HDR1 homologs from different plant species. An un-rooted neighbor-joining tree was reconstructed using amino acid sequences. The length of a branch shows the relative protein sequence difference. (B) to (E) HDR1 is a nuclear protein. B, HDR1-GFP; C, Bright-field; D, OsMADS3-mCherry; and E, merged. Bar = 10 μm in (B) to (E). (F) HDR1 transcript levels in various organs. Mean values ± s.d. were obtained from three technical replicates and two biological replicates. (G) Rhythmic expression of HDR1. The rice Ubiquitin-1 (Ubq1) gene was used as an internal control. The values presented are the mean ± s.d. of three independent experiments and two biological replicates. The open and filled bars at the top and bottom represent the light and dark periods, respectively. s.d.: standard deviation. (H) to (K) HDR1 expression pattern in pHDR1::GUS transgenic rice. GUS staining of a transverse section of a leaf (H), a small spikelet (I), a primary root (J), and a bud (K). Bars = 100 μm. s.d.: standard deviations.
Upon HDR1 expression in rice leaf protoplasts, the in-frame HDR1 cDNA::GFP fusion protein was specifically localized in the nucleus (Fig 3B–3E). To investigate the expression profile of HDR1 in rice, we examined HDR1 expression levels in several rice tissues under LDs. Higher HDR1 expression was detected in leaves and floral organs (Fig 3F), in accordance with the previously described expression patterns of several other flowering-time genes [19–38]. Furthermore, genomic HDR1fragments (including 2 kb of the promoter and part of the coding region) were fused in frame to the β-GLUCURONIDASE (GUS) gene to generate rice transgenic plants expressing the fusion protein. Histochemical staining revealed that HDR1 was strongly detected in leaves compared with other tissues (Fig 3H–3K).
Many flowering genes display a diurnal expression pattern [19–38]. We therefore examined the daily rhythmic expression pattern of HDR1 transcripts under SDs and LDs. Under both conditions, HDR1 expression showed a diurnal expression pattern in leaves. The transcript level began to increase after dusk, reaches a peak before dawn, and damps rapidly thereafter under both SD and LD (Fig 3G).
HDR1 acts as an upstream activator of Hd1 and a repressor of Ehd1
The early flowering of hdr1 compared with WT under LDs but not SDs is similar to the phenotypes of a previously described Hd1 knock-down mutant [23–25] and Ehd1-overexpressing plants [26]. Hd1 and Ehd1 are two pivotal parts of the network that controls flowering in rice [13]; therefore, Hd1 and Ehd1 expression were studied in WT and hdr1 plants under LDs and SDs. Hd1 rhythmic expression was significantly decreased, whereas Ehd1 expression was significantly increased, under LDs but not SDs in hdr1 compared with WT plants (Fig 4A and 4B).These results suggested that HDR1 might participates in the Hd1 and Ehd1 pathways to control flowering time in rice under LDs. Previous work showed that Hd1 can repress Ehd1 expression [44]; therefore, it is possible that the down-regulation of Ehd1 is due to altered Hd1 transcription.
Fig 4. The expression patterns of Hd1, Ehd1, Hd3a and RFT1 in hdr1, and HDR1 in flowering-time mutants or their NILs.

(A) The rhythmic expression patterns of Hd1, Ehd1, Hd3a and RFT1 under SDs. The open and filled bars at the bottom represent the light and dark periods, respectively. The rice Ubiquitin-1 (Ubq1) gene was used as an internal control. The values presented are the mean ± s.d. of three independent experiments and two biological replicates. (B) The rhythmic expression patterns of Hd1, Ehd1, Hd3a and RFT1 under LDs. The open and filled bars at the bottom represent the light and dark periods, respectively. The rice Ubiquitin-1 (Ubq1) gene was used as an internal control. The values presented are the mean ± s.d. of three independent experiments and two biological replicates. (C) Real-time PCR analysis of HDR1 expression level in various flowering-time mutants or their NILs (near-isogenic lines) corresponding WTs. Transcriptional levels was analyzed under both SD and LD conditions. The values of folds change presented are the mean ± s.d. of three independent experiments and two biological replicates. s.d.: standard deviation.
The expression of a non-functional allele of Hd1 or low-level expression of Hd1 in plants causes high-level expression of the florigen gene Hd3a, indicating that Hd1 represses Hd3a expression and delays flowering under LDs [18,25], whereas Ehd1 increases Hd3a and RFT1 expression to promote flowering under LDs [26]. To confirm whether the decreased expression of Hd1 and increased expression of Ehd1 in hdr1 are responsible for the up-regulation of Hd3a and RFT1 and the subsequent promotion of flowering in rice, we examined the rhythmic expression patterns of Hd3a and RFT1 in mutant and WT plants. Under SDs, Hd3a and RFT1 expression remained unchanged between hdr1 and WT (Fig 4A). However, Hd3a and RFT1 expression increased significantly compared with the levels in WT in the light and were maintained at low levels in the dark under LDs (Fig 4B). These results were consistent with the conclusion that the two FT-like genes Hd3a and RFT ensure flowering in rice under LDs [13,22].
HDR1 rhythmic expression in hd1-mutant and ehd1 plants was also analyzed. HDR1 expression did not differ significantly in hd1-mutant plants compared with WT under LDs (S3A Fig). The lack of change in HDR1 expression in the hd1 background and the significant suppression of Hd1 expression in an hdr1 background under LDs demonstrates that HDR1 is an upstream regulator of Hd1. HDR1 expression was not also significantly affected in ehd1 plants, indicating that Ehd1 might encode a downstream component of HDR1 (S3A Fig).
OsGI [18,19,23–25], OsPhyB [45], Ehd2 [28–30], Ehd3 [37], Ehd4 [38], Ghd7 [34], DTH8 [35], OsMADS50 [31], and OsMADS51 [27] are important genes under the control of Hd1 and Ehd1 expression under LDs. We next investigated whether these regulatory genes are down- or up-regulated by HDR1. The expression levels of OsGI, OsphyB, Ehd2, Ehd3, Ehd4, Ghd7, DTH8,OsMADS50, and OsMADS51 in the hdr1 background were similar to WT under LDs (S3C Fig). When we tested the expression levels of HDR1 in osgi, osphyb, ghd7, dth8, osmads50, osmads51, ehd2, ehd3 and ehd4 mutants or nonfunctional NIL, the qPCR assay did not indicate down or up-regulation of these flowering-time regulation genes (Fig 4B). The expression data indicated that HDR1 function as flowering regulator, independent of OsGI, OsphyB, hd2, Ehd3, Ehd4, Ghd7, DTH8, OsMADS50, and OsMADS51. In addition, a yeast two-hybrid assay did not indicate any direct association between HDR1 and these flowering regulators to regulate flowering in rice (S4B Fig). We further carried out a yeast one-hybrid assay to investigate whether HDR1 can bind the promoter regions of Hd1 and Ehd1. However, HDR1 could not bind the promoter regions of Hd1 and Ehd1 (S8 Fig).
HDR1 and OsK4 proteins form nuclear complexes
HDR1 encodes a homolog of the ligand of the SNF1/AMPK/SnRK1 kinase PpSKI in moss [40]. Several kinase genes belonging to the SnRK family have been identified in plants, including Arabidopsis and rice [46–49]. A yeast two-hybrid (Y2H) library, which consisted of different developmental stages leaf and floral tissues, was screened to identify interacting kinase of HDR1 in rice. Using HDR1 as bait, multiple independent fragments for many interacting proteins were identified. Among them, 5 independent clones that fulfilled the criteria of interaction with HDR1 were isolated and sequenced. The different sequences of these 5 clones match to be a gene OsK4 (LOC_Os08 g37800 [TIGR]). A protein with 97.84% similarity to OsK4, named OsK3 (Os03 g0289100 [TIGR]), also exists in the rice genome (S5A Fig). Firstly, we conducted yeast two-hybrid assays to confirm direct interaction of OsK4 and OsK3 with HDR1. Indeed, OsK4 and OsK3 both directly interacted with HDR1 (Fig 5A, S5B Fig). Detailed assays were also performed to clarify the role of the different exons of HDR1 in interacting with OsK4 in the Y2H system, which revealed that exon1 and exon2, but not exon3, strongly interacted with OsK4. Given the evolutionary conservation of the C-terminus of HDR1, these data indicated that the N-terminus of HDR1 is important for interacting with OsK4 (Fig 5B and S2 Fig). In addition, we also noted that hdr1 exhibited late germination, similar to that reported for osk3 and osk4 mutants (S6 Fig and [48]).
Fig 5. HDR1 interacts with OsK4.

(A) HDR1 directly interacted with kinase OsK4 in the Y2H system. (B) Exon1 and Exon2 of HDR1, but not Exon3, directly interacted with OsK4 in yeast cells. (C) BiFC analyses of the interactions of HDR1 with OsK4 in Arabidopsis protoplast. YFP signals resulted from the physical association of HDR1 with OsK4 proteins, not negative pairs, in the nuclei. Scale bar = 10μm. (D) Co-immunoprecipitation of HDR1 with OsK4 in rice plant. Total protein extracted from the leaves expressing FLAG-HDR1 and WT, and then FLAG-HDR1 was immunoprecipitated with anti-FLAG, followed by western blotting to analyze the OsK4 with OsK4 antibody.
To investigate the subcellular localization of the HDR1 and OsK4 interaction in vivo, bimolecular fluorescence complementation (BiFC) was employed to confirm direct interaction between HDR1 and OsK4 using Arabidopsis protoplasts. As expected, YFP fluorescence was only detected in Arabidopsis protoplasts co-transfected with pSAT1-nVenus-C-HDR1 and pSAT1-cCFP-N-OsK4, not two negative controls of pSAT1-nVenus-C-HDR1/pSAT1-cCFP-N and pSAT1-nVenus-C/pSAT1-cCFP-N-OsK4, confirming the physical interaction of HDR1 and OsK4 in vivo (Fig 5C). Next, we conducted co-immunoprecipitation (Co-IP) experiments to verify the interaction between HDR1 and OsK4. For this purpose, we generated transgenic lines expressing FLAG-HDR1 driven by Actin1 promoter could rescue hdr1 phenotype (S10 Fig), and utilized an OsK4 antibody that could specifically recognize the OsK4 protein (S7 Fig). We found that anti-FLAG (recognizing FLAG-HDR1) could efficiently immunoprecipitate OsK4, revealing that OsK4 associated with HDR1 (Fig 5F). Collectively, these data revealed that HDR1 strongly interacts with OSK4.
OsK4 acts similarly to HDR1 to repress flowering
To investigate the function of OsK4 in rice flowering, we generated OsK4- and OsK3-RNAi plants. The RNAi lines were first identified using a GUS activity assay (the vector pTCK309 contains a β-GLUCURONIDASE (GUS) gene derived by the 35S promoter within the T-border, which enables the convenient identification of transgenic plants), and then confirmed by qPCR (Fig 6A). The OsK4-RNAi lines flowered approximately 2 weeks earlier than WT, whereas no early flowering plants were observed among the OsK3-RNAi lines (Fig 6B and 6C). We further constructed the hdr1 OsK4-RNAi double mutant, and found it flowered similarly with hdr1 mutant in LDs (Fig 6D).
Fig 6. OsK4-RNAi plants exhibit early flowering.

(A) The mRNA expression levels of OsK3 and OsK4 in WT and RNAi plants. The values presented are the mean ± s.d. of three independent experiments and two biological replicates. (B) The early flowering phenotype of OsK4-RNAi plants in a paddy field (middle and right). WT Nipponbare (left). (C) Flowering time of OsK3-RNAi, OsK4-RNAi and WT plants under different day-length conditions (n = 12). NDs, natural day conditions; SDs, short day conditions; LDs, long day conditions. (D) Flowering time of hdr1, OsK4-RNAi, hdr1, OsK4-RNAi and WT plants under NDs and LDs (n = 12). s.d.: standard deviations.
The early flowering of the hdr1 and OsK4-RNAi plants coupled with the interaction between HDR1 and OsK4 raised the question of whether a similar regulatory pathway involving HDR1 and OsK4 operates in rice under LD. qPCR analyses were conducted under LDs and SDs, but the upregulation of Ehd1 and downregulation of Hd1 were detected only under LD, confirming that OsK4 exhibited the same pattern of regulation as HDR1. Similarly, the increase in Ehd1 and steep decrease in Hd1 caused the upregulation of the florigensRFT1 and Hd3a was observed only under LD (Fig 7A).
Fig 7. The expression patterns of flowering genes in OsK4-RNAi plants are similar to those in hrd1.

(A) The expression patterns of Hd1, Ehd1, Hd3a and RFT1 under SDs and LDs. The rice Ubiquitin-1 (Ubq1) gene was used as an internal control. The values presented are the mean ± s.d. of three independent experiments and two biological replicates. (B) Real-time PCR analysis of representative flowering-related genes in WT and OsK4-RNAi plants. Nine reported flowering-related genes (Ehd2, Ehd3, Ehd4, OsGI, OsPhyB, OsMADS50, OsMADS51, Ghd7 and DTH8) were analyzed under both SDs and LDs. The values presented are the mean ± s.d. of three independent experiments and two biological replicates. s.d.: standard deviations.
OsGI, OsPhyB, Ehd2, Ehd3, Ehd4, Ghd7, DTH8, OsMADS50 and OsMADS51 were also investigated in OsK4-RNAi plants to check for an alternative regulatory pathway. The two RNAi lines exhibited no obvious differences in expression compared with WT plants (Fig 7B).
OsK4 associated with HDR1 functions to phosphorylate HD1
To clarify the relationship between HDR1 and OsK4, we first analyzed the localization and expression of OsK4 and HDR1 in hdr1 or OsK4-RNAi transgenic plants. In hdr1, the OsK4-GFP fusion protein localized to the nucleus (S9A–S9D Fig), in accordance with a previous report [48]. To investigate whether a lack of HDR1 function affects the OsK4 protein level, we performed western blotting to measure OsK4 protein levels in hdr1 and WT, and found no differences in the levels of OsK4 (Fig 8A and 8B). Two transgenic lines were used to determine the HDR1-GFP fusion protein location. HDR1 was exclusively localized to the nucleus, as in WT (S9E–S9L Fig). Interestingly, HDR1 protein levels were dramatically decreased in OsK4-RNAi plants (Fig 8C and 8D).
Fig 8. The relationship between HDR1 and OsK4.

(A) OsK4 protein level remains unchanged in hdr1. The Heat-shock Protein (HP) was used as a loading control. Total protein extracted from the 40 d old seedlings of WT and hdr1, was subjected to precipitates with anti-OsK4 or anti-HP respectively by western blotting analysis. (B) OsK4 protein intensity in WT and hdr1, quantified by ImageJ program. (C) HDR1 protein level is decreased in OsK4-RNAi transgenic lines 7 and 19. The HP was used as a loading control. Total protein extracted from the 40 d old seedlings of WT, OsK4-RNAi transgenic lines 7 and 19, were subjected to precipitates with anti-HDR1 or anti-HP respectively by western blotting analysis. (D) HDR1 protein intensity in WT and OsK4-RNAi transgenic lines 7 and 19, quantified using ImageJ program. (E) The expression rhythm of HDR1 in OsK4-RNAi-7 and OsK4-RNAi-19 plants. (F) Overexpression of HDR1 rescused the phenotype of OsK4-RNAi.
The low HDR1 levels in Osk4-RNAi lines may due to the low HDR1; therefore, we examined the HDR1 gene rhythmic expression pattern in WT and the OsK4-RNAi lines. The expression rhythm of HDR1 was not changed in OsK4-RNAi (Fig 8E). We also found that overexpression of HDR1 could rescue the early flowering phenotype of OsK4-RNAi plants (Fig 8F).
Given our findings that HDR1 and OsK4 did not directly interact with HD1 or EHD1 (S4A Fig), but that HDR1 acted as an upstream regulators of Hd1 and Ehd1 and could form a complex with OsK4, we explored the possibility of a physical interaction between HDR1-OsK4 and HD1 or EHD1. We performed Y3H assays with HD1 or EHD1 fused to the GAL4 activation domain (AD), and OsK4 fused to the GAL4-binding domain (BD) along with HDR1 co-expression. We found that, indeed, HDR1 and OsK4 together could interact with HD1, not EHD1 (Fig 9A). To further verify that HDR1 and OsK4 interacted to HD1, a Co-IP assay was carried out. We found that HDR1 and OsK4 effectively precipitated HD1 when FLAG-HDR1 and OsK4 were input together, indicating that HD1 is part of the HDR1-OsK4 complex (Fig 9B).
Fig 9. OsK4 associated with HDR1 function to phosphorylate HD1.
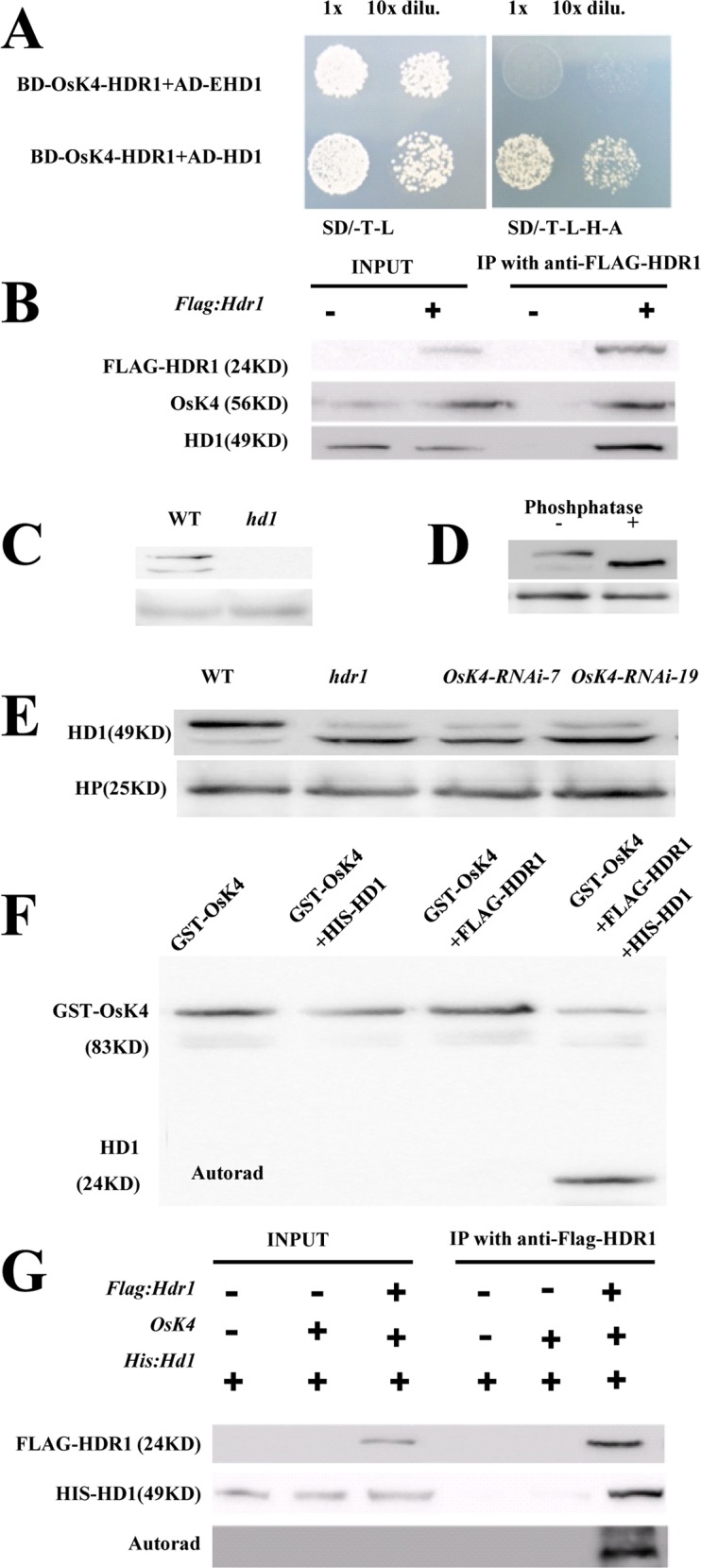
(A). HDR1-OsK4 directly interacted with HD1 in the Y3H system. (B). Co-immunoprecipitation of HDR1-OsK4 with HD1 in rice plant. Total protein extracted from the leaves expressing FLAG-HDR1 and WT, then, FLAG-HDR1 was immunoprecipitated with anti-FLAG, followed by western blotting to analyze the HD1 and OsK4 with specific antibody. (C) Two different forms of HD1 were detected by anti-HD1 antibody in WT.HP protein was used as a loading control. Proteins isolated form 40 d seedlings grown under LDs (14 h light/10 h dark). (D) The slower-migrating form of HD1was susceptible to phosphatase treatment. Nuclear proteins were treated with lambda (λ) phosphatase. Anti-HD1 antibodies were used to detect HD1 protein and HP as a loading control. (E) The different forms of HD1 in WT, hdr1 and Osk4-RNAi plants. The main forms of HD1 are phosphorylated in WT, but in hdr1 and Osk4-RNAi were non-phosphorylated. HP protein was used as a loading control. Proteins isolated form 40 days seedlings grown under LDs (14 h light/10 h dark). (F) In vitro labeling test of HD1 protein with radioactive phosphate. The recombinant purified proteins of OsK4, Hd1 and HDR1 were incubated with radioactive phosphate. (G) OsK4 depended on HDR1 to phosphorylate HD1 in vivo. OsK4-HDR1 complex was immunoprecipitated from transgenic Flag:HDR1 plants. Anti-HD1 antibody was used to detect HD1 protein and HP as a loading control.
It was reported that the HD1 ortholog CO in Arabidopsis could be phosphorylated [50]. We found accordingly that two forms of HD1 protein migrated differently after electrophoresis in WT, but none HD1 protein was detected in hd1 (Fig 9C). To examine whether phosphorylation of HD1 contributes to these different forms, nuclear protein extracts of WT were incubated with λ phosphatase. After incubation of protein phosphatase, the slower-migrating form of HD1 was no longer detected (Fig 9D). The direct interaction of HDR1 and OsK4 with HD1 in vivo raise the question of whether OsK4 phosphorylates HD1, as well as the role of HDR1 in the process of HD1 phosphorylation. Firstly, we performed western blot to verify phosphorylated HD1 level in WT, hdr1 and OsK4-RNAi plants. Two forms of HD1 protein migrated differently after electrophoresis in WT and mutant lines, and less phosphorylated HD1 protein was detected in hd1 and OsK4-RNAi lines (Fig 9E). To verify the phosphorylation of the HD1, we carried out an in vitro phosphorylation assays. Using the recombinant purified proteins GST-OsK4, FLAG-HDR1 and HIS-HD1 incubated with [γ-32P] ATP, autoradiogram indicated only in the presence of HDR1, OsK4 could phosphorylate HD1 (Fig 9F). We further confirmed that OsK4 immunoprecipitated from Flag:HDR1 plants could phosphorylate HD1 in vivo (Fig 9G). All the data gave clues that HDR1was necessary for the phosphorylation of HD1 by OsK4.
Discussion
In this study, we have identified HDR1 interacted with OsK4 kinase protein, and was essential for inhibiting flowering by regulating the expression of Hd1 and Ehd1, leading to the downregulation of Hd3a and RFT1 under LD. We have further determined that OsK4 and HDR1 could phosphorylate HD1. These findings collectively illustrated the important role of the HDR1 and OsK4 complex in the day-length regulation of flowering time via HD1.
HDR1 transcriptionally regulates Hd1 to control flowering time in rice
Rice is known as a short-day plant, and most rice cultivars may be induced to flower more rapidly under SDs than LDs. There are at least two independent flowering pathways, Hd1 and Ehd1, that regulate floral transition in rice [13,21]. The Hd1 pathway is conserved between rice and Arabidopsis, but the Ehd1 pathway is unique to rice [21]. Our results reveal that HDR1 shows a similar diurnal expression pattern with Hd1 and Ehd1, which accumulates after dusk, reaches a peak before dawn, and damps rapidly thereafter under both SD and LD (Fig 3G). The expression pattern implicated HDR1 in photoperiodic control of flowering through the regulation of Hd1 and Ehd1. Upon the loss of HDR1 function, Hd1 rhythmic expression was significantly decreased under LDs but not SDs, whereas Ehd1 expression was significantly increased (Fig 4A and 4B). Previous studies have indicated that Hd1 may function to repress Ehd1 expression [44]. The results of the present study indicated that Hd1 expression is suppressed, and Ehd1 expression is promoted by HDR1 under LDs (Fig 4B). Thus, HDR1 might integrate different pathways in the regulation of flowering, in which Hd1 is upregulated by HDR1to repress Ehd1 expression, and may also be independently involved in the Ehd1 pathway.
Several genes that control the Hd1 and Ehd1 pathway have been identified and studied, including OsGI, Ehd2, Ehd3, Ehd4, Ghd7, DTH8, OsMADS50 and OsMADS51 [27–38]. In an effort to explore the relationship between HDR1 and these known regulators, we first examined whether the expression levels of these genes were altered in the relevant mutants. The expression levels of HDR1 and these genes remain unchanged in WT and mutants under LD (Fig 4C and S3C Fig). We also observed the nuclear localization of HDR1 which suggests this protein may function to form nuclear complexes with regulatory factors to control flowering time. However, HDR1 did not associate with these flowering regulators (S4B Fig). These results suggested that HDR1 is a novel regulator of flowering time, independent of OsGI, Ehd2, Ehd3, Ehd4, Ghd7, DTH8, OsMADS50, and OsMADS51 (Fig 10). However, HDR1 was unable to bind the Hd1 and Ehd1 promoters (S8 Fig), indicating that there are other components to media the binding of this complex or Hd1 and Ehd1 are likely indirect targets.
Fig 10. A working model of HDR1- and OsK4-mediated flowering pathway in rice.

Rice flowering is mainly regulated by the Hd1-dependent and Ehd1-dependent pathways. HDR1 and OsK4 come together to regulate Hd1 and Ehd1 expression and phosphorylated HD1 in the control of flowering time in rice. The black lines show regulators that have added to this flowering network from our study, and the gray lines indicate previously identified genes. P: indicates Phosphorylation.
HDR1 is associated with OsK4 to control flowering in rice
HDR1 is ubiquitous in plants, including mosses, implying its ancient origin. The only reported homolog is PpSKI [40], which encodes a ligand of the kinase SnRK1 in moss. The deletion of PpSKI increases gametophore formation and reduces protonemal growth under low-light conditions but not under normal light conditions in moss [40]. However, HDR1 functions in floral transition in rice, and disruption of HDR1 shows early flowering. Moreover, the PpSKI-interacting protein SnRK1, which is a Snf1-related protein kinase found in yeast, animals, and plants, is required for necessary metabolic changes in plants during dark hours [39]. In plants, Snf1-related kinases participate in the regulation of many physiological processes, including the cell cycle, meristem development, and pathogen responses [49]. As predicted, HDR1 act as a Snf1-related kinase interactor 2, and associated protein OsK4, a Snf1-related kinase has been identified [39].We postulated that this complex may function to phosphorylate target proteins. To test this hypothesis, we first analyzed the localization and expression of OsK4 and HDR1 in hdr1 or OsK4-RNAi transgenic plants to clarify the relationship between HDR1 and OsK4. In hdr1, the OsK4-GFP fusion protein localized to the nucleus (S9A–S9D Fig), in accordance with a previous report [48]. Interestingly, the HDR1 protein levels were dramatically decreased in OsK4-RNAi plants (S9E–S9L Fig). The fact that disrupting OsK4 function leads to effect steady-state levels of HDR1 expression suggests that OsK4 functions to affect the protein levels of HDR1. These results collectively indicated that the early flowering phenotype in OsK4-RNAi plants might be regulated by a decrease in HDR1 protein levels.
Our study indicated that HDR1 and OsK4 regulate Hd1 expression at transcriptional level. Protein interaction analyses support that HDR1-OsK4 directly interacts with HD1 at protein level. These data suggested that might exist a feedback way to regulate Hd1, The likely regulating model could be found in some important functional gene in plants. Auxin response factors (ARFs), NPH4/ARF7 and ARF19, could bind auxin response promoter elements and mediate gene transcription [51]. Auxin induces the ARF19 gene, and NPH4/ARF7 and ARF19 together are required for its expression, suggesting NPH4/ARF7 and ARF19 may activate ARF19 in a positive feedback loop [51]. Hd1 like ARF19, as above described, is regulated in a positive feedback way needed HDR1-OsK4 functionally.
OsK4 functions depend on HDR1 to phosphorylate HD1
As mentioned above, OsK4 and HDR1 act as a Snf1-related kinase and a Snf1-related kinase interactor 2, respectively. Recently studies indicated that the protein kinase CK2 (Hd6) might control Hd1 activity through the phosphorylation of an unknown interacting factor to control flowering time in rice [50]. We thought that HDR1-OsK4 might also phosphorylate HD1, although the expression level of Hd1 is regulated by HDR1 and OsK4. As expected, HDR1-OsK4 could form nuclear complexes with HD1 in vivo, and phosphorylate HD1 (Fig 9C). The HDR1 is key component of the complex to assist phosphorylation of HD1. Taken together, these findings suggest that HDR1-OsK4 acts to downregulate Hd1/Ehd1 expression and phosphorylate Hd1 to prevent early flowering in LDs, conferring a photoperiodic response.
AtSnRK1, the OsK4 ortholog in Arabidopsis, is an atypical AMPK. AtSnRK1 family members comprise a catalytic α subunit and non-catalytic β and γ subunits, and multiple isoforms of each subunit type exist, giving rise to various isoenzymes [52]. This suggested that HDR1 might functionally act as non-catalytic β or γ subunits in rice. It is reported that the SnRK1 kinase complex are necessary to phosphorylate regulated transcription factors [46,49]. Hence, the phosphorylation of HD1 needs not only catalytic subunit OsK4, but also subunits such as HDR1 presenting. It has not reported that phosphorylated transcription factors prior to combine the kinase complex or not [46,49], whether HDR1 can only interact with phosphorylated Hd1 needed further illuminated later.
Materials and Methods
Plant materials and measurements
The Tos17-tagged mutant hdr1 was identified from our T-DNA insertion rice mutant library (O. sativa L. spp. japonica cv. Nipponbare) [41–43]. Screening for flowering time mutants was carried out by planting the mutant materials under natural LD conditions in the experimental field at the Chinese Academy of Agricultural Sciences in Beijing (39°54'N, 116°23'E), China, from May to October of each year. The agricultural traits of mutant and WT plants were recorded in three growing seasons.
Rice plants were also grown in a growth chamber under LD (14 h light/10 h dark) and SD (10 h light/14 h dark) with a light intensity of 800 μmol•m-2•s-1. To analyze the diurnal expression patterns of flowering genes and flowering time, the materials were first grown under NLD for 30–40 days and then transferred to LD or SD for 10 days.
hdr1 mutant isolation and complementation
Genomic DNA flanking the Tos17 left border was cloned using PCR-based genome walking with nested specific primers according to Peng, with modifications [42]. The modified primers were TB1, TB2, AP1 and AP2. The products were sequenced using TB2 as the sequencing primer. The Tos17 insertion site in HDR1 was identified by BLAST searches of the rice genome (http://blast.ncbi.nlm.nih.gov/Blast.cgi) using the rescued flanking sequences.
Two constructs were used for complementation testing. For the cDNA construct, the full-length HDR1 cDNA (amplified using primer pair HDR1-cp3-F and HDR1-cp3-R, S1 Table) was cloned into the binary vector pCam2300K, which was modified from pCambia2300Actin1 carrying the 1.8-kb HDR1 promoter (amplified using primer pair HDR1-cp1-F and HDR1-cp2-R, S1 Table), to generate pHDR1::HDR1. For the genomic construct, a 5.1-kb fragment containing the promoter, coding, and terminator regions of HDR1 was amplified from genomic DNA (using the primer pair HDR1-cp1-F and HDR1-cp1-R, S1 Table) and then cloned into the vector pCam2300K. The two constructs were introduced into Agrobacterium tumefaciens strain AGL1 by electroporation and then transformed into hdr1 mutant calli. The regenerated plants were grown in a paddy field at the CAAS in Beijing, China [42]. The genotype of each transgenic plant was determined by PCR.
Plasmid construction and generation of p35S::HDR1-RNAi, p35S::OsK4-RNAi, p35S::OsK3-RNAi, and pActin1::Flag:HDR1 transgenic plants
DNA encompassing 113 bp of the 5' UTR of HDR1, 150 bp of the 5' UTR of OsK4 and 160 bp of the 5' UTR of OsK3 were amplified using specific primer pairs (S1 Table) and inserted into pTCK309 using the SacI, SpeI, KpnI, and BamHI sites in inverted orientations. The construct was transferred to A. tumefaciens strain AGL1 by electroporation and then introduced into WT rice calli. The regenerated plants were grown in a paddy field at the CAAS in Beijing. Early flowering RNAi transgenic plants were selected and analyzed. To generate pActin1::Flag:HDR1, the coding region of HDR1 was amplified using a specific primer pair (S1 Table) and inserted into pCamabia1307 using the XbaI and SalI sites. The plasmid containing Flag-HDR1 was digested with XbaI and SalI and then inserted into pCambia2300Actin1 using the XbaI and SalI sites. The construct was introduced into hdr1calli through A. tumefaciens-mediated transformation, and the transgenic plants were grown in a paddy field and analyzed by comparing the flowering time with WT plants.
Subcellular localization of HDR1 and OsK4
The full-length coding sequence of Hdr1 or OsK4 was PCR-amplified from cDNA using the primers pairs HDR1-GFP-fw and HDR1-GFP-rv or OsK4-GFP-fw and OsK4-GFP-rv (S1 Table) and cloned into pAN580 to generate p35S:: HDR1:GFP or p35S::OsK4:GFP. The fusion plasmid and pMcherry (p35S::RFP, as a control) were transiently co-expressed in rice leaf protoplasts by PEG (polyethylene glycol) as previously described [38]. The protoplast cell layers were then examined by laser scanning confocal microscopy [Leica TCS SP2; Wetzlar, Germany]. GFP fluorescence was imaged using excitation with the 488-nm argon laser line and a 505–530-nm band-pass emission filter.
Reverse transcription (RT)-PCR and quantitative (q)RT-PCR
Total RNA was extracted using the RNeasy Plant Mini Kit (QIAGEN, Hiden, Germany) from various rice plants (WT, hdr1, hd1, ehd1, OsK4-RNAi, OsK3-RNAi, HDR1-overexpressing and various flowering relating genes [38]). For mutant or near isogenic lines of flowering time genes, penultimate leaves were harvested around the reported peak expression level of each gene during the 24-h photoperiod according to Gao (Penultimate leaves were harvested around the reported peak expression level of each gene during the 24-h photoperiod at dawn for OsPhyB, Ehd1, Ehd2, Hd3a, RFT1 and Ehd4; 3 h after dawn for Ghd7; 8 h after dawn for Ehd3, OsMADS50, OsMADS51 and DTH8; and immediately after dusk for OsGI and Hd1 [38]). Total RNA (2 μg) from each sample was reverse-transcribed with an oligo(dT) primer and Ace RT Enzyme [Toyobo, Osaka, Japan] according to the manufacturer’s instructions. The PCR conditions were as follows: preincubation at 94°C for 2.5 min, followed by 30 cycles of 94°C for 20 s, 52°C for 20 s, and 72°C for 30 s. qRT-PCR was performed on an iQ5 Multicolor Real-Time PCR Detection System [Bio-Rad] with SYBR Green Real-Time PCR Master Mix (Life Technologies). The rice Ubiquitin1 gene was selected as an internal standard. The fold change of the transcript level of a gene of interest to that of UBQ1 is calculated as 2-ΔCt [ΔCT = CT (gene of interest)—CT (UBQ)]. The primer pairs for other gene amplifications are specified in S2 Table.
Histochemical staining for GUS activity
GUS assays were carried out as described previously [43]. Rice tissues were incubated in GUS staining solution (50 mM sodium phosphate, pH 7.0, 10 mM EDTA, 0.1% Triton X-100, 1 mg/ml X-gluc, 0.1 mM potassium ferricyanide, and 10% methanol) at 37°C for 10–12 h, followed by washing with 70% ethanol. The stained tissues were imaged under a zoom stereo microscope [Nikon SMZ1000; Tokyo, Japan].
BiFC assay
The protein-coding regions of HDR1 and OsK4 were amplified using gene-specific primers (S1 Table). Then, the PCR products were ligated into pSAT1-nVenus-C and/or pSAT1-cCFP-N using the EcoRI and XhoI/SalI sites to produce pSAT1-nVenus-C-HDR1, pSAT1-cCFP-N-OsK4, pSAT1-nVenus-C-OsK4, and pSAT1-cCFP-N-HDR1. All constructs were verified by sequencing. Plasmid pairs were transfected into Arabidopsis protoplasts as described previously, using pairs of each construct and the empty vector as negatives control [53]. The cells were then examined by laser scanning confocal microscopy (Leica TCS SP2). YFP fluorescence was imaged by excitation with the 515-nm argon laser line and a 535–565-nm band-pass emission filter.
Yeast two-hybrid analysis
The protein-coding regions of HDR1, OsK3, OsK4, Hd1, Ehd1, Hd3a, RFT1, Ehd2, Ehd3, Ehd4, OsMADS50, OsMADS51, OsGI, OsPhyB, DTH8 and Ghd7 were amplified using gene-specific primers (S1 Table). Then, the PCR products were fused into the activation domain (AD) vector pGADT7 and/or the DNA-binding domain (BD) vector pGBKT7 using the EcoRI and XhoI/SalI sites. All constructs were verified by sequencing. The constructs were then transformed into Saccharomyces cerevisiae strain AH109 [BD Biosciences, Palo Alto, CA, USA] according to the manufacturer’s protocol. No detectable self-activation was observed for each construct on SD selective medium (SD-His-Leu + 5 mM 3-AT or SD-His-Trp + 5 mM 3-AT). Yeast cells were spotted on SD media lacking leucine, tryptophan, histidine and adenine.
Yeast one-hybrid assay
The 2.2-kb and 4.2-kb promoter regions of Hd1 and Ehd1 were amplified (S1 Table), and the fragments were then cloned into the pLacZi vector using EcoRI and XhoI sites to generate pLacZip-Hd1 and pLacZip-Ehd1. The resulting plasmids were used to transform yeast cells, followed by secondary transformation with pGBKT7-HDR1 and selection according to the manufacturer’s manual [PT1031-1, CLONTECH Laboratories Inc., Mountain View, CA]. Full-length of OsLFL1 cDNA was amplified (S1 Table) and then inserted into plasmid pGBKT to generate pGBKT-OsLFL1 as a positive control, as OsLFL1 binds the Ehd1 promoter region. Plasmids were co-transformed into the yeast strain YM4271. Yeast cell were spotted onto SD/His- plates and grown for 2 days. Then, the yeast cells were transferred onto SD/His-/Leu- 45mM 3-AT plates with X-gal (5-bromo-4-chloro-3-indolyl-b-D-galacto-pyranoside) for blue color detection.
Yeast three-hybrid assay
HDR1 was amplified (S1 Table) and digested with EcoRI and BamHI, OsK4 was amplified and digested with BamHI and PstI, and the PCR fragments were then introduced into the pBridge vector using the EcoRI and PstI sites, generating pBridgeHDR1-OsK4. Then, the vector sequence was verified by DNA sequencing, and the constructs were used to transform yeast cells. The resulting strains were co-transformed with the AD fusion vector pGADT7-HD1 and pGADT7-EHD1. Transformants were selected on medium according to the manufacturer’s instructions (PT3212-5; CLONTECH Laboratories Inc., Mountain View, CA, USA). Plasmids were co-transformed into the yeast strain AH109. Yeast cells were spotted and grown on SD medium without leucine, tryptophan, histidine and adenine.
Western blotting
Total proteins were extracted from six-week-old rice seedlings in RIPA buffer (50 mM Tris-HCl, pH 7.2, 150 mM NaCl, 10 mM MgCl2, and 1 mM phenylmethylsulfonyl fluoride, protease inhibitor mixture [Sigma, St. Louis, MO, USA]), and the supernatant was collected. The total protein concentration was determined using a Bradford protein assay kit (Bio-Rad). Proteins were detected using anti-FLAG [Sigma, St. Louis, MO, USA], anti-HDR1, anti-HD1 or anti-OsK4 [kindly supplied by Dr. Liu, BIG, CAS] antibodies at 1:4000 dilution and visualized with an enhanced chemiluminescence (ECL) reagent (GE Healthcare).
The HIS-HDR1 fusion protein were overexpressed in Escherichia coli (strain BL21) and purified to generate multi-clone antibodies. Frozen cells were extracted with 100 μl of extraction buffer (50 mM Tris-HCl. pH 7.6, 15 mM MgCl2, 0.1 M KCl, 0.25 M sucrose, 10% glycerol, 1 mM phenylmethylsulfonyl fluoride, protease inhibitor mixture [Sigma, St. Louis, MO, USA], and 14 mM β-mercaptoethanol). After centrifugation of the sample at 15,000 x g for 10 min, the supernatant was sampled, and the protein concentration was measured. The amount of protein form each extract was measured according to the manufacturer’s instructions [Laboratories Inc., Mountain View, CA, USA].
Co-immunoprecipitation
For Co-IP assays, total protein extracted from FLAG:HDR1 transgenic or none-transgenic and OsK4-RNAi transgenic or none-transgenic was mixed with purified 2mg HIS-HD1 and anti-FLAG antibody. The mixture was incubated overnight with Magna Protein A Magnetic Beads [Millipore, Temecula, California, USA] at 4°C in 1000 μl of binding buffer (20 mM Tris-HCl, pH 7.6, 2.5 mM β-mercaptoethanol, and 0.1 M NaCl). After incubation, the beads were washed five times with washing buffer (20 mM Tris-HCl, pH 7.5, 500 mM NaCl, and 0.5% Triton X-100). After washing, 40 μl of 1× SDS-PAGE sample buffer was added, the mixture was denatured, and the sample was loaded on a 10% SDS-PAGE gel. Proteins were detected using a horseradish peroxidase (HRP)-conjugated anti-FLAG [Sigma, St. Louis, MO, USA], anti-HD1 or anti-HIS antibody, and then visualized with an ECL reagent (GE Healthcare).
Protein phosphorylation assay
Protein phosphorylation experiments were carried out based on previously described [54]. Briefly, for in vitro phosphorylation of rice HD1 by OsK4, purified GST-OsK4, FLAG-HD1 and/or HIS-HDR1 fusion proteins in reaction buffer (50 mM HEPES-KOH (pH 7.5), 10 mM MgCl2, 5 mM MnCl2, 1 mM DTT, 0.1 mM ATP, and 10 mCi of [γ-32P]ATP). The mixture was allowed to react for 30 min at 30°C. The phosphorylation of proteins was taken autoradiogram using a phosphor imager after SDS-PAG electrophoresis.
Immunoprecipitated proteins from FLAG-HDR1 plant bound to the protein A beads were also subjected to kinase assays. Each reaction contained 10μl of beads in reaction buffer as described above. The mixture was allowed to react for 30 min at 30°C. Radioactive signals were detected using a phosphor imager. An HD1 phosphorylation assay was also conducted using anti-HIS antibodies.
Supporting Information
(A) Phenotype of HDR1 overexpression plants under LDs. (B) The dates of flowering are unchanged for WT and Overexpression plants (OX) under NDs, LDs and SDs. (C) The expression patterns of the key flowering-related genes (Hd1, Ehd1, Hd3a and RFT1) are similar in WT and OX plants, as determined by semi-quantitative RT-PCR.
(TIF)
(TIF)
The rhythmic expression pattern of HDR1 in the hd1 and ehd1 mutants under SD and LD (A), and the expression levels of OsGI, OsPhyB, Ehd2, Ehd3, Ehd4, Ghd7, DTH8, OsMADS50, and OsMADS51 in the WT and hdr1 under LDs (B).
(TIF)
(A) HDR1 or OsK4 had no interaction with HD1 and EHD1. (B) HDR1 had no association with HD3A, RFT1, EHD2, EHD3, EHD4, OsGI, OsPHYB, OsMADS50, OsMADS51 and DTH8.
(TIF)
(A) Sequence alignment of OsK3 and OsK4. (B) OsK3 physically interacted with HDR1 in yeast.
(TIF)
(TIF)
Proteins were exacted from 6-week-old WT, OsK3-RNAi and OsK4-RNAi leaves.
(TIF)
The flowering regulator OsLFL1 was used as a positive control.
(TIF)
(A)-(D) The location of OsK4 was not changed in hrd1. A, HDR1-GFP; B, Bright field; C, OsMADS3-mCherry; and D, Merged. (E) to (H) The location of HDR1 was not changed in the OsK4-RNAi-7 transgenic line. E, HDR1-GFP; F, Bright field; G, OsMADS3-mCherry; and H, Merged. (I) to (L) The location of HDR1 was not changed in the OsK4-RNAi-19 transgenic line. I, HDR1-GFP; J, Bright field; K, OsMADS3-mCherry; and L, Merged. Bar = 10μm.
(TIF)
(TIF)
(DOC)
(DOC)
(DOC)
Acknowledgments
We thank Dr. Chengcai Chu (IDBG, CAS) for providing the binary vector pCAMBIA2300Actin1. We thank Dr. Kang Chong (IB, CAS) for providing the binary vector pTCK309. We thank Dr. Jianmin Wan (ICS, CAAS) for providing the vectors pAN580 and pMcherry, we also thanks Dr. Wan for providing rice mutants of flowering time. We also thank Dr. GuoZhen Liu (BIG, CAS) for providing antibodies for HP, HDR1, HD1, OsK3 and OsK4, which greatly supported this work.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was supported in part by the National Natural Science Foundation of China (grant no. 31271697), the Ministry of Science and Technology of China (grant no. AA10A300), and Recruitment program of Global Youth Expert of China (to XG). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Thomas B, Vince-Prue D. Photoperiodism in plants 2nd ed. San Diego: Academic Press; 1997. [Google Scholar]
- 2.Liu C, Xi W, Shen L, Tan C, Yu H. Regulation of floral patterning by flowering time genes. Dev Cell. 2009;16: 711–722. 10.1016/j.devcel.2009.03.011 [DOI] [PubMed] [Google Scholar]
- 3.Izawa T. Adaptation of flowering-time by natural and artificial selection in Arabidopsis and rice. J Exp Bot. 2007;58: 3091–3097. [DOI] [PubMed] [Google Scholar]
- 4.Distelfeld A, Li C, Dubcovsky J. Regulation of flowering in temperate cereals. CurrOpin Plant Biol. 2009;12: 178–184. [DOI] [PubMed] [Google Scholar]
- 5.Greenup A, Peacock WJ, Dennis ES, Trevaskis B. The molecular biology of seasonal flowering-responses in Arabidopsis and the cereals. Ann Bot. 2009;103: 1165–1172. 10.1093/aob/mcp063 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Fornara F, de Montaigu A, Coupland G. SnapShot: control of flowering in Arabidopsis. Cell. 2010;141: 550.e1–2. [DOI] [PubMed] [Google Scholar]
- 7.Li F, Sun J, Wang D, Bai S, Clarke AK, Holm M. The B-Box family gene STO (BBX24) in Arabidopsis thaliana regulates flowering time in different pathways. PLOS ONE. 2014;9: e87544 10.1371/journal.pone.0087544 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Takahashi Y, Teshima KM, Yokoi S, Innan H, Shimamoto K. Variations in Hd1 proteins, Hd3a promoters, and Ehd1 expression levels contribute to diversity of flowering time in cultivated rice. Proc Natl AcadSci U S A. 2009;106: 4555–4560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Imaizumi T, Kay SA. Photoperiodic control of flowering: not only by coincidence. Trends Plant Sci. 2006;11: 550–558. [DOI] [PubMed] [Google Scholar]
- 10.Kobayashi Y, Weigel D. Move on up, it’s time for change—mobile signals controlling photoperiod-dependent flowering. Genes Dev. 2007;21: 2371–2384. [DOI] [PubMed] [Google Scholar]
- 11.Turck F, Fornara F, Coupland G. Regulation and identity of florigen: FLOWERING LOCUS T moves center stage. Annu Rev Plant Biol. 2008;59: 573–594. 10.1146/annurev.arplant.59.032607.092755 [DOI] [PubMed] [Google Scholar]
- 12.Nonoue Y, Fujino K, Hirayama Y, Yamanouchi U, Lin SY, Yano M. Detection of quantitative trait loci controlling extremely early heading in rice. TheorAppl Genet. 2008;116: 715–722. [DOI] [PubMed] [Google Scholar]
- 13.Tsuji H, Taoka K, Shimamoto K. Regulation of flowering in rice: two florigen genes, a complex gene network, and natural variation. CurrOpin Plant Biol. 2011;14: 45–52. [DOI] [PubMed] [Google Scholar]
- 14.Simpson GG. Evolution of flowering in response to day length: flipping the CONSTANS switch. BioEssays. 2003;25: 829–832. [DOI] [PubMed] [Google Scholar]
- 15.Samach A, Onouchi H, Gold SE, Ditta GS, Schwarz-Sommer Z, Yanofsky MF, et al. Distinct roles of CONSTANS target genes in reproductive development of Arabidopsis. Science. 2000;288: 1613–1616. [DOI] [PubMed] [Google Scholar]
- 16.Suárez-López P, Wheatley K, Robson F, Onouchi H, Valverde F, Coupland G. CONSTANS mediates between the circadian clock and the control of flowering in Arabidopsis. Nature. 2001;410: 1116–1120. [DOI] [PubMed] [Google Scholar]
- 17.Kojima S, Takahashi Y, Kobayashi Y, Monna L, Sasaki T, Araki T, et al. Hd3a, a rice ortholog of the Arabidopsis FT gene, promotes transition to flowering downstream of Hd1 under short-day conditions. Plant Cell Physiol. 2002;43: 1096–1105. [DOI] [PubMed] [Google Scholar]
- 18.Hayama R, Yokoi S, Tamaki S, Yano M, Shimamoto K. Adaptation of photoperiodic control pathways produces short-day flowering in rice. Nature. 2003;422: 719–722. [DOI] [PubMed] [Google Scholar]
- 19.Tamaki S, Matsuo S, Wong HL, Yokoi S, Shimamoto K. Hd3a protein is a mobile flowering signal in rice. Science. 2007;316: 1033–1036. [DOI] [PubMed] [Google Scholar]
- 20.Komiya R, Ikegami A, Tamaki S, Yokoi S, Shimamoto K. Hd3a and RFT1 are essential for flowering in rice. Development. 2008;135: 767–774. 10.1242/dev.008631 [DOI] [PubMed] [Google Scholar]
- 21.Komiya R, Yokoi S, Shimamoto K. A gene network for long-day flowering activates RFT1 encoding a mobile flowering signal in rice. Development. 2009;136: 3443–3450. 10.1242/dev.040170 [DOI] [PubMed] [Google Scholar]
- 22.Itoh H, Nonoue Y, Yano M, Izawa T. A pair of floral regulators sets critical day length for Hd3aflorigen expression in rice. Nat Genet. 2010;42: 635–638. 10.1038/ng.606 [DOI] [PubMed] [Google Scholar]
- 23.Yano M, Katayose Y, Ashikari M, Yamanouchi U, Monna L, Fuse T, et al. Hd1, a major photoperiod sensitivity quantitative trait locus in rice, is closely related to the Arabidopsis flowering time gene CONSTANS. Plant Cell. 2000;12: 2473–2483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Yano M, Kojima S, Takahashi Y, Lin H, Sasaki T. Genetic control of flowering time in rice, a short-day plant. Plant Physiol. 2001;127: 1425–1429. [PMC free article] [PubMed] [Google Scholar]
- 25.Izawa T, Oikawa T, Sugiyama N, Tanisaka T, Yano M, Shimamoto K. Phytochrome mediates the external light signal to repress FT orthologs in photoperiodic flowering of rice. Genes Dev. 2002;16: 2006–2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Doi K, Izawa T, Fuse T, Yamanouchi U, Kubo T, Shimatani Z, et al. Ehd1, a B-type response regulator in rice, confers short-day promotion of flowering and controls FT-like gene expression independently of Hd1. Genes Dev. 2004;18: 926–936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kim SL, Lee S, Kim HJ, Nam HG, An G. OsMADS51 is a short-day flowering promoter that functions upstream of Ehd1, OsMADS14, and Hd3a. Plant Physiol. 2007;145: 1484–1494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Park SJ, Kim SL, Lee S, Je BI, Piao HL, Park SH, et al. Rice indeterminate 1 (OsId1) is necessary for the expression of Ehd1 (early heading date 1) regardless of photoperiod. Plant J. 2008;56: 1018–1029. 10.1111/j.1365-313X.2008.03667.x [DOI] [PubMed] [Google Scholar]
- 29.Matsubara K, Yamanouchi U, Wang ZX, Minobe Y, Izawa T, Yano M. Ehd2, a rice ortholog of the maize INDETERMINATE1 gene, promotes flowering by up-regulating Ehd1. Plant Physiol. 2008;148: 1425–1435. 10.1104/pp.108.125542 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Wu C, You C, Li C, Long T, Chen G, Byrne ME, et al. RID1, encoding a Cys2/His2-type zinc finger transcription factor, acts as a master switch from vegetative to floral development in rice. Proc Natl AcadSci U S A. 2008;105: 12915–12920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Lee S, Kim J, Han JJ, Han MJ, An G. Functional analyses of the flowering time gene OsMADS50, the putative SUPPRESSOROFOVEREXPRESSIONofCO1/AGAMOUS-LIKE 20 (SOC1/AGL20) ortholog in rice. Plant J. 2004;38: 754–764. [DOI] [PubMed] [Google Scholar]
- 32.Ryu CH, Lee S, Cho LH, Kim SL, Lee YS, Choi SC, et al. OsMADS50 and OsMADS56 function antagonistically in regulating long day (LD)-dependent flowering in rice. Plant Cell Environ. 2009;32: 1412–1427. 10.1111/j.1365-3040.2009.02008.x [DOI] [PubMed] [Google Scholar]
- 33.Peng LT, Shi ZY, Li L, Shen GZ, Zhang JL. Ectopic expression of OsLFL1 in rice represses Ehd1 by binding on its promoter. BiochemBiophys Res Commun. 2007;360: 251–256. [DOI] [PubMed] [Google Scholar]
- 34.Xue W, Xing Y, Weng X, Zhao Y, Tang W, Wang L, et al. Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice. Nat Genet. 2008;40: 761–767. 10.1038/ng.143 [DOI] [PubMed] [Google Scholar]
- 35.Wei X, Xu J, Guo H, Jiang L, Chen S, Yu C, et al. DTH8 suppresses flowering in rice, influencing plant height and yield potential simultaneously. Plant Physiol. 2010;153: 1747–1758. 10.1104/pp.110.156943 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Yan W, Wang P, Chen H, Zhou H, Li Q, Wang C, et al. A major QTL, Ghd8, plays pleiotropic roles in regulating grain productivity, plant height, and heading date in rice. Mol Plant. 2011;4: 319–330. 10.1093/mp/ssq070 [DOI] [PubMed] [Google Scholar]
- 37.Matsubara K, Yamanouchi U, Nonoue Y, Sugimoto K, Wang ZX, Minobe Y, et al. Ehd3, encoding a plant homeodomain finger-containing protein, is a critical promoter of rice flowering. Plant J. 2011; 66: 603–612. 10.1111/j.1365-313X.2011.04517.x [DOI] [PubMed] [Google Scholar]
- 38.Gao H, Zheng XM, Fei G, Chen J, Jin M, Ren Y, et al. Ehd4 encodes a novel and oryza-genus-specific regulator of photoperiodic flowering in Rice. PLOS Genet. 2013;9: e1003281 10.1371/journal.pgen.1003281 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Thelander M, Olsson T, Ronne H. Snf1-related protein kinase 1 is needed for growth in a normal day-night light cycle. EMBO J. 2004;23: 1900–1910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Thelander M, Nilsson A, Olsson T, Johansson M, Girod PA, Schaefer DG, et al. The moss genes PpSKI1 and PpSKI2 encode nuclear SnRK1 interacting proteins with homologues in vascular plants. Plant Mol Biol. 2007;64: 559–573. [DOI] [PubMed] [Google Scholar]
- 41.Yang Y, Peng H, Huang H, Wu J, Jia S, Huang D, et al. Large-scale production of enhancer trapping lines for rice functional genomics. Plant Sci. 2004;167: 281–288. [Google Scholar]
- 42.Peng H, Huang H, Yang Y, Zhai Y, Wu J, Huang D, et al. Functional analysis of GUS expression patterns and T-DNA integration characteristics in rice enhancer trap lines. Plant Sci. 2005;168: 1571–1579. [Google Scholar]
- 43.Wan S, Wu J, Zhang Z, Sun X, Lv Y, Gao C, et al. Activation tagging, an efficient tool for functional analysis of the rice genome. Plant Mol Biol. 2009;69: 69–80. 10.1007/s11103-008-9406-5 [DOI] [PubMed] [Google Scholar]
- 44.Gómez-Ariza J, Galbiati F, Goretti D, Brambilla V, Shrestha R, Pappolla A, eral.Loss of floral repressor function adapts rice to higher latitudes in Europe. J Exp Bot. 2015;66: 2027–2039. 10.1093/jxb/erv004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Izawa T, Oikawa T, Sugiyama N, Tanisaka T, Yano M, et al. Phytochrome mediates the external light signal to repress FT orthologs in photoperiodic flowering of rice. Genes Dev 2005;16: 2006–2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Halford NG, Hardie DG. SNF1-related protein kinases: global regulators of carbon metabolism in plants? Plant Mol Biol. 1998;37: 735–748. [DOI] [PubMed] [Google Scholar]
- 47.Halford NG, Hey S, Jhurreea D, Laurie S, McKibbin RS, Paul M, et al. Metabolic signallingand carbon partitioning: role of Snf1-related (SnRK1) protein kinase. J Exp Bot. 2003;54: 467–475. [DOI] [PubMed] [Google Scholar]
- 48.Lu CA, Lin CC, Lee KW, Chen JL, Huang LF, Ho SL,et al. The SnRK1A protein kinase plays a key role in sugar signaling during germination and seedling growth of rice. Plant Cell. 2007;19: 2484–2499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Halford NG, Hey SJ. Snf1-related protein kinases (SnRKs) act within an intricate network that links metabolic and stress signalling in plants. Biochem J. 2009;419: 247–259. 10.1042/BJ20082408 [DOI] [PubMed] [Google Scholar]
- 50.Sarid-Krebs L, Panigrahi KC, Fornara F, Takahashi Y, Hayama R, et al. Phosphorylation of CONSTANS and its COP1-dependent degradation during photoperiodic flowering of Arabidopsis. Plant J. 2015;84: 451–63. 10.1111/tpj.13022 [DOI] [PubMed] [Google Scholar]
- 51.Wilmoth JC, Wang S, Tiwari SB, Joshi AD, Hagen G et al. NPH4/ARF7 and ARF19 promote leaf expansion and auxin-induced lateral root formation. Plant J. 2005;43: 118–30. [DOI] [PubMed] [Google Scholar]
- 52.Emanuelle S, Hossain MI, Moller IE, Pedersen HL, van de Meene AM, et al. SnRK1 from Arabidopsis thaliana is an atypical AMPK. Plant J. 2015;82:183–92. 10.1111/tpj.12813 [DOI] [PubMed] [Google Scholar]
- 53.Citovsky V, Lee LY, Vyas S, Glick E, Chen MH, Vainstein A, et al. Subcellular localization of interacting proteins by bimolecular fluorescence complementation inplanta. J Mol Biol. 2006;362: 1120–1131. [DOI] [PubMed] [Google Scholar]
- 54.Li CH, Wang G, Zhao JL, Li-Qing Zhang LQ, Ai LF et al. The Receptor-Like Kinase SIT1 Mediates Salt Sensitivity by Activating MAPK3/6 and Regulating Ethylene Homeostasis in Rice. The Plant Cell. 2014;26: 2538–2553. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(A) Phenotype of HDR1 overexpression plants under LDs. (B) The dates of flowering are unchanged for WT and Overexpression plants (OX) under NDs, LDs and SDs. (C) The expression patterns of the key flowering-related genes (Hd1, Ehd1, Hd3a and RFT1) are similar in WT and OX plants, as determined by semi-quantitative RT-PCR.
(TIF)
(TIF)
The rhythmic expression pattern of HDR1 in the hd1 and ehd1 mutants under SD and LD (A), and the expression levels of OsGI, OsPhyB, Ehd2, Ehd3, Ehd4, Ghd7, DTH8, OsMADS50, and OsMADS51 in the WT and hdr1 under LDs (B).
(TIF)
(A) HDR1 or OsK4 had no interaction with HD1 and EHD1. (B) HDR1 had no association with HD3A, RFT1, EHD2, EHD3, EHD4, OsGI, OsPHYB, OsMADS50, OsMADS51 and DTH8.
(TIF)
(A) Sequence alignment of OsK3 and OsK4. (B) OsK3 physically interacted with HDR1 in yeast.
(TIF)
(TIF)
Proteins were exacted from 6-week-old WT, OsK3-RNAi and OsK4-RNAi leaves.
(TIF)
The flowering regulator OsLFL1 was used as a positive control.
(TIF)
(A)-(D) The location of OsK4 was not changed in hrd1. A, HDR1-GFP; B, Bright field; C, OsMADS3-mCherry; and D, Merged. (E) to (H) The location of HDR1 was not changed in the OsK4-RNAi-7 transgenic line. E, HDR1-GFP; F, Bright field; G, OsMADS3-mCherry; and H, Merged. (I) to (L) The location of HDR1 was not changed in the OsK4-RNAi-19 transgenic line. I, HDR1-GFP; J, Bright field; K, OsMADS3-mCherry; and L, Merged. Bar = 10μm.
(TIF)
(TIF)
(DOC)
(DOC)
(DOC)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.