

Summary
Aging constitutes the key risk factor for age‐related diseases such as cancer and cardiovascular and neurodegenerative disorders. Human longevity and healthy aging are complex phenotypes influenced by both environmental and genetic factors. The fact that genetic contribution to lifespan strongly increases with greater age provides basis for research on which “protective genes” are carried by long‐lived individuals. Studies have consistently revealed FOXO (Forkhead box O) transcription factors as important determinants in aging and longevity. FOXO proteins represent a subfamily of transcription factors conserved from Caenorhabditis elegans to mammals that act as key regulators of longevity downstream of insulin and insulin‐like growth factor signaling. Invertebrate genomes have one FOXO gene, while mammals have four FOXO genes: FOXO1, FOXO3, FOXO4, and FOXO6. In mammals, this subfamily is involved in a wide range of crucial cellular processes regulating stress resistance, metabolism, cell cycle arrest, and apoptosis. Their role in longevity determination is complex and remains to be fully elucidated. Throughout this review, the mechanisms by which FOXO factors contribute to longevity will be discussed in diverse animal models, from Hydra to mammals. Moreover, compelling evidence of FOXOs as contributors for extreme longevity and health span in humans will be addressed.
Keywords: aging, animal models, FOXO transcription factors, insulin and IGF‐1 signaling pathway, longevity
Abbreviations
- 4E‐BP
4E‐binding protein
- ADIPOQ
adiponectin gene (structural homology to complement factor C1q)
- AGE‐1
aging alteration 1
- AFX
acute leukemia fusion gene located in chromosome X
- AKT
human homolog of viral oncogene v‐akt
- AMP
adenosine monophosphate
- AMPK
AMP‐activated protein kinase
- APG
autophagy
- APOE
apolipoprotein E
- aqp‐1
aquaporin 1
- ATP
adenosine triphosphate
- AUT10
autophagocytosis 10
- bec‐1
beclin 1
- CHARGE
cohorts for heart and aging research in genomic epidemiology
- CRM1
chromosome region maintenance 1
- COQ7
coenzyme Q7 homolog
- DAF
dauer formation
- dFoxO
Drosophila FOXO
- Dilps
Drosophila insulin‐like peptides
- DNA
deoxyribonucleic acid
- DR
dietary restriction
- FIRKO
fat‐specific insulin receptor knockout
- FKHR
Forkhead in rhabdomyosarcoma
- FKHRL1
Forkhead in rhabdomyosarcoma like protein 1
- FOXA
Forkhead box transcription factors of the class A
- FOXO
Forkhead box transcription factors of the class O
- FOXS
Forkhead box transcription factors of the class S
- GFP
green fluorescent protein
- GH
growth hormone
- GWAS
genomewide association scan
- HSCs
hematopoietic stem cells
- HOMA
homeostasis model assessment
- HSF‐1
heat‐shock factor 1
- IGF‐1
insulin‐like growth factor 1
- IGF1R
insulin‐like growth factor 1 receptor
- IIS
insulin and IGF‐1 signaling pathway
- InR
insulin‐like receptor
- Ins
insulin‐like peptide
- JNK
Jun N‐terminal kinase
- KRI‐1
Caenorhabditis elegans orthologous to human Krev interaction trapped 1 (KRIT1)
- L1‐L4
larval stages of C. elegans
- MAF
minor allele frequency
- mTOR
mammalian target of rapamycin
- Nrf
nuclear respiratory factor
- PDK‐1
3‐phosphoinositide‐dependent kinase‐1
- PHA‐4
pharynx 4‐cell embryo
- PI3K
phosphoinositide 3‐kinase
- PSMD11
proteasome (PSM), non‐ATPase subunit (D) 11
- PTEN
phosphatase and tensin homolog
- RNA
ribonucleic acid
- S6K1
ribosomal S6 protein kinase 1
- SCL‐1
SCP‐like extracellular protein 1
- SGK
serum‐ and glucocorticoid‐inducible protein kinase
- SIRT1
sirtuin 1
- sir‐2.1
sirtuin 2.1
- SKN‐1
skinhead 1
- SNPs
single nucleotide polymorphisms
- STACs
sirtuin‐activating compounds
- TCER
transcription elongation regulator
- TOR
target of rapamycin
- VPS30
vacuolar protein sorting 30
Introduction
Aging can be generally defined has the functional deterioration of physiological mechanisms which strictly depends on the passage of time. This decline constitutes the key risk factor for age‐related diseases such as cancer and cardiovascular and neurodegenerative disorders. It is therefore not surprising that humankind has put considerable efforts in understanding the processes of aging (and how to delay them) since ancient times.
Human longevity and healthy aging are complex phenotypes influenced by environmental (diet, physical activity, health habits, and psychosocial factors) and genetic factors (Herskind et al., 1996; Christensen et al., 2006; Bishop & Guarente, 2007). Heritability accounts for ≈25% of lifespan in an average‐lived population. The genetic contribution of lifespan increases with greater age, particularly after the age of 60, reaching estimates of 33% in women and 48% in men living to at least 100 (Sebastiani & Perls, 2012; Brooks‐Wilson, 2013). Accordingly, longevity clusters within families as parents and siblings of centenarians have an increased probability of reaching advanced age (Perls et al., 2000, 2002; Atzmon et al., 2004; Willcox et al., 2006).
It has become increasingly evident that lifespan is closely related to health span and that long‐lived individuals develop chronic illnesses (both physical and cognitive) later in life, thereby confirming the compression of morbidity hypothesis proposed by Fries in 1980 (Fries, 1980; Hitt et al., 1999; Andersen et al., 2012). A study performed in centenarians revealed that there is a progressive compression of disability and morbidity such that at the remarkable survival age of 110, subjects presented with age‐related diseases in the last 5.2% of their lives (vs. 17.9% in controls, 9.4% in individuals 100–104, and 8.9% in individuals 105–109) (Andersen et al., 2012). As extreme longevity appears to be a result of genetic factors more than environmental ones, when compared to an average lifespan, significant effort has gone into determining which genetic variants can slow aging and diminish the risk for age‐related diseases. Interestingly, the Leiden Longevity Study showed that, by GWAS, genomes of nonagenarians carry the same number of disease risk alleles for coronary artery disease, cancer and type 2 diabetes as young controls (Beekman et al., 2010). These results hint toward the perception that these long‐lived individuals could carry “protective genes” that may work in general cellular defense mechanisms, for example, against oxidative stress. In fact, several studies have consistently revealed APOE and FOXOs (FOXO1 and FOXO3) as “longevity genes” (Willcox et al., 2006; Anselmi et al., 2009; Flachsbart et al., 2009; Soerensen et al., 2010, 2015; Brooks‐Wilson, 2013; Bao et al., 2014; Broer et al., 2015). Aging has long been considered a process of degradation occurring in a random fashion that would lead to the accumulation of cellular damage in a stochastic fashion and, consequently, tissue decline and death. However, it is now known that aging can be modulated by genetic pathways and biochemical processes which are evolutionarily conserved (Kenyon, 2010b; Lopez‐Otin et al., 2013). According to the quasi‐programmed theory, aging is not programmed, but rather a consequence of genetic programs that determine developmental growth early in life (Blagosklonny, 2013a,b). Lopez‐Otin et al. (2013) in an attempt to define common denominators of aging in different organisms have defined nine cellular and molecular hallmarks of aging: genomic instability, telomere attrition, epigenetic alterations, loss of proteostasis, deregulated nutrient sensing, mitochondrial dysfunction, cellular senescence, stem cell exhaustion, and altered intercellular communication.
Among these hallmarks, the “deregulated nutrient sensing” was the first to be described to influence aging in animals, through the insulin and IGF‐1 signaling pathway (IIS) (Kenyon, 2005). IGF‐1 is produced by several cells types (mainly hepatocytes) in response to GH release from the anterior pituitary. IGF‐1 has been shown to trigger the same intracellular signaling pathways stimulated by insulin. The IIS pathway is the most evolutionarily conserved pathway of aging, shown to modulate lifespan in model organisms across a great evolutionary distance from Caenorhabditis elegans to mice (Kimura et al., 1997; Tatar et al., 2001; Fontana et al., 2010; Kenyon, 2010b; Mercken et al., 2013). Accordingly, genetic polymorphisms/mutations that cause loss of function of GH, IGF‐1 receptor, insulin receptor or its downstream factors, have been implicated in human longevity as in model organisms (Fontana et al., 2010; Kenyon, 2010b; Tazearslan et al., 2011; Barzilai et al., 2012; Milman et al., 2014). Dietary restriction is a well‐known environmental signal shown to expand lifespan in eukaryote species, from yeast to primates (Colman et al., 2009; Fontana et al., 2010; Mattison et al., 2012). The “longevity response” to dietary restriction is regulated by several nutrient‐sensing pathways: the kinase TOR, AMP kinase, sirtuins, and the IIS (Kenyon, 2005).
FOXO transcription factors
FOXO proteins are the most important transcriptional effectors of the IIS (Kenyon et al., 1993; Gottlieb & Ruvkun, 1994; Brunet et al., 1999; Dong et al., 2008). FOXOs represent a subfamily of the Forkhead family of transcription factors. This family is characterized by a conserved DNA‐binding domain (the Forkhead box or FOX) and comprises more than 100 members in humans, from FOXA to FOXS (Zanella et al., 2010; Genin et al., 2014). The FOXO subfamily is conserved from C. elegans to mammals but, while invertebrates have only one FOXO gene, mammals have four FOXO genes: FOXO1 (FKHR), FOXO3 (FKHRL1), FOXO4 (AFX), and FOXO6 (Kaestner et al., 2000; Hannenhalli & Kaestner, 2009). In mammals, this subfamily is involved in a wide range of crucial cellular processes regulating stress resistance, metabolism, cell cycle arrest, and apoptosis, but their role in longevity still remains to be elucidated. FOXO proteins function mainly as transcriptional activators by binding the consensus core recognition motif TTGTTTAC, and their activity is inhibited by the IIS pathway (Biggs et al., 1999; Brunet et al., 1999; Henderson & Johnson, 2001; Lin et al., 2001; Calnan & Brunet, 2008; Zanella et al., 2010; Webb & Brunet, 2014). Briefly, insulin or IGF‐1 triggers an intracellular pathway mediated by PI3K‐AKT, allowing phosphorylation of FOXO factors by the serine/threonine kinase AKT at three conserved residues within the FOXO proteins. AKT‐mediated phosphorylation of FOXO leads to its nuclear exclusion and, in turn, to suppression of FOXO‐dependent transcription of target genes (Guo et al., 1999; Murphy et al., 2003). Conversely, in the absence of growth factor signaling or upon cellular stress, FOXOs translocate into the nucleus and activate FOXO‐dependent gene expression. A diverse set of posttranslational modifications in addition to phosphorylation, such as acetylation/deacetylation, methylation, or ubiquitination has been shown to promote changes of subcellular localization, protein levels, DNA binding, and transcriptional activity of FOXO factors (Calnan & Brunet, 2008; Webb & Brunet, 2014) The combinatorial result of FOXO posttranslational modifications has been proposed to lead to the recruitment of specific FOXO‐binding partners regulating different FOXO‐dependent gene expression programs (Greer et al., 2007b; Calnan & Brunet, 2008; Hill et al., 2014). Several mechanisms of how FOXO proteins promote longevity have been suggested.
FOXO and autophagy
Webb & Brunet (2014) have recently unveiled the role of FOXOs as prolongevity factors through the maintenance of protein homeostasis (proteostasis) (Morley et al., 2002; Hsu et al., 2003). In fact, it has been shown that FOXO factors participate in the regulation of genes responsible for two main mechanisms of intracellular clearance: autophagy and the ubiquitin‐proteasome system (Webb & Brunet, 2014). Defects in autophagy, the process of degradation and recycling of cytoplasmic proteins and organelles in response to starvation have been associated with premature aging and age‐related disorders (Hara et al., 2006; Komatsu et al., 2006; Jung et al., 2008; Pickford et al., 2008; Masiero et al., 2009; Lee et al., 2010a, 2010b). FOXOs affect the expression of genes involved in autophagy and mitophagy (muscle‐specific autophagy) in muscle cells from flies (dFOXO) to mammals (FOXO3), allowing adaptation of the tissues to starvation (Zhao et al., 2007; Sengupta et al., 2009; Demontis & Perrimon, 2010). Additionally, FOXO1 and FOXO3 activate autophagy mechanisms in diverse cell types: neurons, cardiomyocytes, renal tubular cells, and HSCs (Webb & Brunet, 2014). As mentioned, FOXOs are involved in the proteasome system degradation of short‐lived and regulatory cytosolic proteins. Aging is associated with a decreased proteasomal activity, leading to excess of damaged proteins in muscle, liver, and heart (Conconi et al., 1996; Petropoulos et al., 2000; Bulteau et al., 2002; Husom et al., 2004). Moreover, pathogenesis of neurodegenerative disorders such as Parkinson's, Alzheimer's, or Huntington's disease is generally related to an abnormal ubiquitin‐proteasome mechanism as either a primary cause or secondary consequence (Ciechanover & Brundin, 2003; Kikis et al., 2010; Webb & Brunet, 2014). FOXOs act on both the upregulation of ubiquitin ligases and by controlling the composition of the proteasome (Bodine et al., 2001; Sandri et al., 2004, 2006; Stitt et al., 2004; Vilchez et al., 2012). However, the direct effect of FOXO‐mediated proteostasis in mammals remains to be understood.
FOXO and resistance to oxidative stress
One of the most significant functions of FOXO proteins is their role in cellular responses to oxidative stress. As accumulation of damage caused by ROS (reactive oxygen species) was postulated to be causative for aging, it has been hypothesized that FOXO factors influence aging and age‐related diseases by increasing the antioxidant capacity of cells (Kops et al., 2002; Storz, 2011). ROS play an important role as second messengers of cellular signaling and can lead to oxidative stress when cellular detoxification activity is decreased. As high and very low levels of ROS lead to impaired cellular functions, maintaining intracellular ROS homeostasis is essential to prevent pathological processes including cancer and other age‐associated diseases. FOXOs are regulated by oxidative stress via changes in upstream FOXO regulatory pathways or directly sensing the cellular redox status through reversible oxidation and reduction of cystein residues (Essers et al., 2004; Eijkelenboom & Burgering, 2013; Putker et al., 2013). FOXO factors regulate the expression of the key detoxification enzymes MnSOD (manganese superoxide dismutase), catalase, and GADD45 (Kops et al., 2002; Nemoto & Finkel, 2002). Accordingly, inactivation of Foxo factors has been shown to lead to intracellular accumulation of ROS promoting accelerated atherosclerosis, proliferation of transformed cells, and compromising long‐term proliferative potential of normal stem cells (Tothova et al., 2007; Tsuchiya et al., 2013).
FOXO and stem cells
There is accumulating evidence that FOXO factors play an important role in stem cell biology and tissue homeostasis. During aging, the balance of removal and regeneration of cells in tissues becomes disturbed mainly due to a decrease in the regenerative potential of adult stem cells. Conditional deletion of Foxo1/3a/4 in the adult hematopoietic system of mice leads to apoptosis of HSCs preventing the repopulation of these stem cell populations. Similarly, aged mice in which Foxo3a was deleted display reduced regenerative potential (Miyamoto et al., 2007). Foxo‐deficient HSCs in these animals are thought to be driven out of quiescence into cell cycle, resulting in depletion of the stem cell pool (Tothova & Gilliland, 2007). Interestingly, the treatment of Foxo‐deficient mice with the antioxidant N‐acetylcysteine restores the HSC compartment, suggesting that the accumulation of ROS disturbs stem cell function. This observation is in line with the idea that decreased function of adult stem cells involved in the onset of age‐related diseases is secondary to the accumulation of cellular stress (Boyette & Tuan, 2014). The role of FOXO proteins in stem cell biology is not limited to adult stem cells. FOXO1 has been shown to directly control the expression of OCT4 and SOX2 two transcription factors critically involved in stemness. Accordingly, FOXO1 is necessary to maintain pluripotency of human ESC and the ortholog FOXO1 exerts a similar function in mouse ESCs (Zhang et al., 2011a).
In this review, we give an overview of the current evidence that implicates FOXO transcription factors in human longevity. Although the precise mechanism by which FOXO factors influence human aging is not understood, an overwhelming amount of data from several animal models including Hydra vulgaris, C. elegans, Drosophila melanogaster, and mice shed light on the critical functions of FOXO protein family members in aging. We discuss the FOXO activities which might be relevant for human longevity throughout this review organized by species (Fig. 1).
Figure 1.
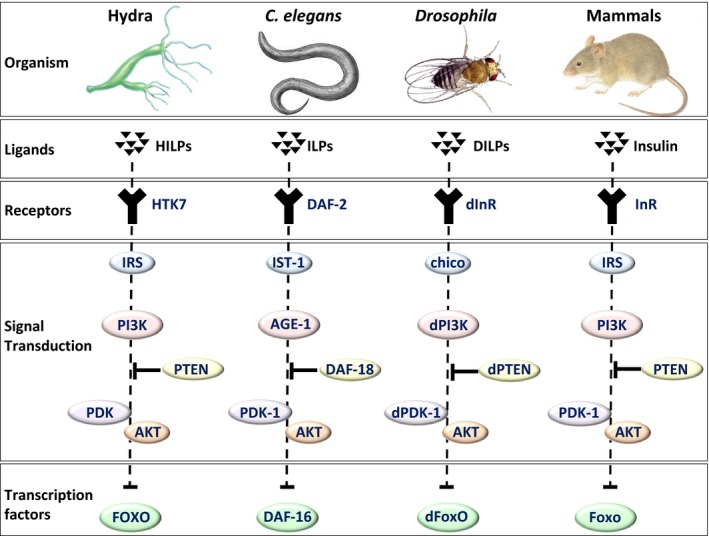
The key components of the PI3K signal transduction pathway are well conserved throughout evolution. The corresponding orthologues for these components in Hydra, Caenorhabditis elegans, Drosophila melanogaster and mammals are illustrated. It is important to note, that there is a single gene for many components, in Hydra, C. elegans and D. melanogaster whereas mammals have several isoforms of these components with the exception of PTEN. AGE‐1, ageing alteration 1; AKT, v‐akt murine thymoma viral oncogene homologue 1; DAF‐2, abnormal dauer formation‐2; FOXO, forkhead family of transcription factor; INR, insulin receptor; IRS, substrate; IST‐1, insulin receptor substrate (IRS)‐like adaptor; PDK, phosphatidylinositide‐dependent protein kinase 1; PTEN; phosphatase and tensin homologue.
Population studies
The understanding that the genetic contribution to lifespan strongly increases with greater age provides basis for research on which genetic differences distinguish centenarians from average‐lived individuals (Herskind et al., 1996; Hjelmborg et al., 2006; Murabito et al., 2012; Sebastiani & Perls, 2012; Brooks‐Wilson, 2013). Accordingly, longevity clusters within families as parents and siblings of centenarians present an increased probability of reaching advanced age (Perls et al., 2000, 2002; Atzmon et al., 2004; Willcox et al., 2006). Alleles which are enriched in centenarians most likely represent genes that are significant for longevity and therefore a significant number of studies have been done in the quest for these alleles/genes.
Willcox et al. (2008) first reported that genetic variation in FOXO3A was strongly associated with human longevity. In a long‐lived population of male Americans of Japanese ancestry (mean attained age = 97.9 vs. control group = 78.5), a nested‐case–control study of five candidate genes (ADIPOQ, FOXO1A, FOXO3A, SIRT1, and COQ7) was performed. Among these, only the rs2764264, rs13217795, and rs2802292 FOXO3A SNPs stood out as they were found to be associated with longevity and healthy aging (Willcox et al., 2008). These long‐lived men had lower prevalence of cancer and cardiovascular disease better self‐reported health as well as high physical and cognitive function, even though they were in average 11 years older than controls. Homozygosity for the G allele of FOXO3A rs2802292 was shown to confer a significant protection considering the prevalence of congestive heart disease. The same allele was associated with markedly lower insulin, log insulin, and HOMA (homeostasis model assessment) score in the control group (Willcox et al., 2008).
Anselmi et al. (2009) validated the association of these FOXO3 polymorphisms with extreme longevity in males from the Southern Italian Centenarian Study. In particular, rs2802288, a proxy of rs2802292, showed the best allelic association‐MAF (minor allele frequency). These results were confirmed in the German population (Flachsbart et al., 2009). An extensive collection of 1762 German centenarians/nonagenarians and younger controls evidenced FOXO3A polymorphisms with the ability to reach exceptional old age. This association was substantially stronger in centenarians than in nonagenarians, highlighting the importance of centenarians in genetic longevity research (Flachsbart et al., 2009). Furthermore, the variation in FOXO3A was replicated by both case–control and longitudinal data in Danish population (the oldest‐old vs. middle‐aged individuals) (Soerensen et al., 2010). Among the fifteen SNPs analyzed, they found association of eight SNPs with longevity: four previously reported (rs13217795, rs2764264, rs479744, and rs9400239) and four novel SNPs (rs12206094, rs13220810, rs7762395, and rs9486902) (Soerensen et al., 2010).
Li et al. (2009) analyzed six SNPs from FOXO1A and FOXO3A genes by comparing 761 centenarians and 1056 younger individuals (control group) of the Han Chinese population. They found two SNPs of FOXO1A to be negatively associated with longevity in women, rs2755209 and rs2755213. On the other hand, all three SNPs studied for FOXO3A (rs2253310, rs2802292, and rs4946936) were positively associated with longevity in both genders (Li et al., 2009). They have concluded that as FOXO1A is more strictly associated with human female longevity, the genetic contribution to longevity trait may be affected by genders (Li et al., 2009). Also in the Han Chinese population, Zeng et al. analyzed the independent and joint effects of the FOXO1A and FOXO3A SNPs on long‐term survival. These authors found substantial gender differences in the independent effects and showed that the positive effects of FOXO3A and negative effects of FOXO1A largely compensate each other, although FOXO3A has a stronger impact (Zeng et al., 2010). Others have also revealed the importance of genetic variations in the IIS pathway in long‐lived individuals from Italian, Japanese, Ashkenazi Jewish, or Dutch ethnicity (Arai et al., 2001; Bonafe et al., 2003; Kojima et al., 2004; Kuningas et al., 2007; Suh et al., 2008; Pawlikowska et al., 2009). A meta‐analysis comprising 11 independent case–control studies and 5241 cases from different ethnic groups revealed five FOXO3A polymorphisms as associated with longevity: rs2802292, rs2764264, rs13217795, rs1935949, and rs2802288 (Bao et al., 2014). Moreover, rs2802292 and rs2764264 polymorphism are male‐specific longevity polymorphisms that may potentially identify long‐lived men. FOXO3A along with APOE has consistently been associated with longevity in multiple independent studies (Wheeler & Kim, 2011; Brooks‐Wilson, 2013). Accordingly, Broer et al. (2015) conducted a meta‐analysis of GWAS with 6036 longevity cases (age ≥ 90 years) and 3757 controls (CHARGE consortium). Among the ~2.5 million SNPs analyzed, only the APOE and FOXO3 variants confirmed a significant association with longevity (Broer et al., 2015). Recent work by Soerensen et al. (2015) compared aging‐related traits (cognitive function, hand grip strength, activity of daily living, and self‐rated health) with 15 FOXO3A SNPs in Danish oldest‐old individuals. Gene‐based testing revealed a significant increase in activity of daily living and reduced bone fracture risk for carriers of the minor alleles of 8 and 10 FOXO3A SNPs, respectively (Soerensen et al., 2015). In summary, gathered evidence from Japanese Americans, Han Chinese, Californians, New Englanders, Ashkenazi Jews, Danish, Germans, and Italians shows that FOXO3A SNPs are associated with exceptional longevity. Although there is compelling evidence that FOXO3 gene sequence variants influence longevity, it remains to be determined how these variations translate into phenotypic characteristics that enable a long lifespan. FOXO3A alleles associated with longevity are intronic and not linked to known coding SNPs (Donlon et al., 2012; Murabito et al., 2012; Brooks‐Wilson, 2013), suggesting that these SNPs will most likely affect FOXO3A expression rather than protein activity.
Animal models
Hydra vulgaris
Hydra vulgaris is a freshwater radial‐symmetric polyp of the phylum Cnidaria, placed at the basal root of the animal life. Bridge et al. (2010) first described the presence of a single FoxO gene in Hydra. These authors found significant parallels in the regulation of FoxO between Hydra and bilaterian animals, showing that FoxO transcriptional activity is negatively modulated by the PI3K/AKT/SGK pathway and, accordingly, nuclear localization of a FoxO‐GFP fusion protein is significantly increased by a PI3K inhibitor (Bridge et al., 2010). Lasi et al. (2010) showed that transient expression of FoxO‐GFP protein induced an apoptosis rate of 20–60% in Hydra epithelial cells. Conversely, co‐expression of one of the Hydra insulin‐like genes with the FoxO‐GFP protein was shown to decrease the rate of apoptosis in these cells. As the IIS acts through the PI3K/AKT/SGK pathway, one can hypothesize that the IIS reduces Hydra′s FoxO activity (Lasi et al., 2010).
Hydra presents itself as unique model to study longevity due to its extraordinary regenerative abilities through the self‐renewal and differentiating capacities of its epithelial and interstitial stem cells. These properties are thought to be related to the fact that these animals routinely reproduce asexually (Wittlieb et al., 2006; Khalturin et al., 2007; Bosch, 2009; Bellantuono et al., 2015). Three stem cell lineages allow this continuous self‐renewal: ectodermal and endodermal epitheliomuscular stem cells and interstitial stem cells (Bosch, 2009; Bosch et al., 2010). The search for transcriptome‐specific signatures enabling the regulation of self‐renewal and differentiation showed that FoxO is highly expressed in the three stem cell lineages of Hydra (Boehm et al., 2012). The overexpression of FoxO in the multipotent interstitial stem cell lineages increases stem cell and progenitor cell proliferation and activates expression of stem cell genes in terminally differentiated somatic cells (Boehm et al., 2012). Conversely, silencing FoxO in epithelial cells increased the number of terminally differentiated cells (Boehm et al., 2012) Altogether, these results suggest a key role for FoxO in Hydra's apparent biological immortality, specifically affecting its continuous self‐renewal capacity (Bellantuono et al., 2015). In a recent essay, Schaible and Sussman hypothesized that whereas FoxO is exclusively devoted to life‐prolonging cell renewal in Hydra, the evolutionary diversification of FoxO functions in multicellular eukaryotes that manifest aging resulted in a dilution of FoxO's rejuvenating capacity (Schaible & Sussman, 2013).
Caenorhabditis elegans
Caenorhabditis elegans is a free‐living unsegmented pseudocoelomate member of the phylum Nematoda that has an average lifespan of about 2–3 weeks. It has two sexes: a self‐fertilizing hermaphrodite and a male. The life cycle of C. elegans is comprised of the embryonic stage, four larval stages (L1–L4), and adulthood. If the environmental conditions are not favorable (absence of food, high temperature, or presence of a pheromone indicating high population density), L1 worms develop into alternative larval form called the dauer larva (Fielenbach & Antebi, 2008). The role of the IIS pathway in aging was first discovered in C. elegans through mutations that extended lifespan. Mutations in DAF‐2 (dauer larvae formation‐2), a hormone receptor similar to insulin and IGF‐1 receptors, doubled the lifespan of the worm (Kenyon et al., 1993; Kimura et al., 1997). This lifespan extension caused by DAF‐2 mutations required the activity of DAF‐16, which encodes the single FoxO homolog in C. elegans (Kenyon et al., 1993; Lin et al., 1997; Ogg et al., 1997). DAF‐16 is inactivated via its nuclear export by the evolutionary conserved signaling pathway downstream of DAF‐2 receptor (Henderson & Johnson, 2001; Lee et al., 2001; Lin et al., 2001). HSF‐1, the C. elegans heat‐shock transcription factor, is also required in DAF‐2 mutants to extend lifespan. HSF‐1, along with DAF‐16, promotes longevity by activating specific longevity genes, including genes that encode small heat‐shock proteins (Garigan et al., 2002; Hsu et al., 2003; Walker & Lithgow, 2003; Morley & Morimoto, 2004). The importance of DAF‐18/PTEN tumor suppressor in longevity and dauer larva formation, through the regulation of the IIS pathway, has been established in the late 1990s by several research groups (Ogg & Ruvkun, 1998; Gil et al., 1999; Mihaylova et al., 1999; Rouault et al., 1999). DAF‐18/PTEN has been shown to regulate L1 arrest in the germ line, and insulin‐like signaling appears to be transduced by AGE‐1/PI3K during L1 arrest (Weinkove et al., 2006; Zhang et al., 2011b).
DAF‐16 functions as the major target of the IIS pathway (Kenyon et al., 1993; Gottlieb & Ruvkun, 1994). DAF‐16/FoxO has been shown to regulate hundreds of genes in C. elegans including those related to stress response, antimicrobial activity, and metabolism, unveiling DAF‐16/FoxO as a central player of a complex network involving multiple upstream pathways and downstream target genes (Lee et al., 2003; McElwee et al., 2003; Murphy et al., 2003). The lifespan expansion effect accomplished by DAF‐16/FOXO is most likely due to two isoform variants DAF‐16a and DAF‐16d/f (Kwon et al., 2010; Chen et al., 2015). Diverse approaches including different dietary regimens or using compounds that mimic dietary restriction have been employed to investigate the effect of dietary restriction on lifespan extension in C. elegans (Kenyon, 2010b). Chronic food limitation increases lifespan by downregulating TOR activity, which increases autophagy and diminishes translation, probably through PHA‐4/FOXA transcription factor and S6 kinase, respectively, in a DAF‐16/FoxO‐independent manner (Kaeberlein et al., 2005; Hansen et al., 2007; Pan et al., 2007; Sheaffer et al., 2008). AMPK (AMP kinase), an energy sensor for cellular AMP/ATP ratio that enables catabolic reactions for energy gain, has been shown to respond to dietary restriction by increasing stress resistance and extending longevity in C. elegans in a DAF‐16/FOXO‐dependent manner (Apfeld et al., 2004; Greer et al., 2007a). AMPK directly phosphorylates and activates DAF‐16/FoxO (Greer et al., 2007a). Dietary restriction with every‐other‐day feeding is likely to promote C. elegans longevity via downregulation of the IIS pathway (Honjoh et al., 2009). In C. elegans, overexpression of sirtuin gene sir‐2.1 activates DAF‐16/FoxO by interacting with 14‐3‐3 proteins (Berdichevsky et al., 2006). The ability of sirtuins to directly deacetylate DAF‐16/FoxO together with the fact that IIS pathway mutants do not need sir‐2.1 to promote longevity, suggests that sirtuins act on DAF‐16/FoxO independently of the IIS pathway (Kenyon, 2010b). Interestingly, whereas high levels of ROS led to premature death, low concentrations caused a prolongation of lifespan and this extension is dependent on both daf‐16 and sir‐2.1 genes (Heidler et al., 2010). Kim et al. (2014) have recently shown that antioxidant treatment can extend the lifespan of C. elegans through the phosphorylation and cytoplasmic retention of DAF‐16/FoxO, involving 3‐phosphoinositide‐dependent kinase‐1 (PDK‐1).
Lee et al. showed that adding 2% glucose to bacterial diet of C. elegans shortened the lifespan by inhibiting the activity of DAF‐16/FoxO and heat‐shock factor HSF‐1. This effect involves the downregulation of an aquaporin glycerol channel, aqp‐1, which is also inhibited by glucose in mammals. Moreover, when 2% glucose was provided to C. elegans insulin/IGF‐1 receptor mutants, their lifespan extension was nearly completed repressed (Lee et al., 2009). The expression of components of the insulin/IGF‐1 longevity pathway in subsets of cells can affect the rate of aging of the entire organism, implying an active coordination of the aging rates between the different tissues to establish homeostasis. Murphy et al. (2007) reveal that in C. elegans, this communication between the tissues is mediated by INS‐7 (an insulin‐like peptide), which is regulated by DAF‐16/FoxO activity in the intestine, and that this regulation allows DAF‐16 activity in the intestine to influence DAF‐16 activity in other tissues. But insulin‐like peptides may not be the only signals that act downstream of DAF‐16/FoxO to influence lifespan. In daf‐16; daf‐2 double mutants, the expression of DAF‐16 specifically in one tissue (intestine/adipose tissue) can increase the worm lifespan by 60% (Libina et al., 2003). As nonintestinal tissues do not contain DAF‐16/FOXO in these double mutant animals, it must influence other downstream longevity signaling pathways in those tissues. The secreted peptide SCL‐1 is a candidate for such a downstream signal (Ookuma et al., 2003).
Several reports indicate that FoxO induces autophagy acting in a non‐cell‐autonomous manner, evidencing that proteostasis is beneficial for longevity. Melendez et al. (2003) showed that autophagy is a cellular pathway essential for dauer development and lifespan extension in C. elegans (Melendez et al., 2003). Using daf‐2 mutants, the authors demonstrate that bec‐1, the C. elegans ortholog of mammalian autophagy gene APG6/VPS30/beclin1, is essential for normal dauer morphogenesis and lifespan extension. Dauer formation is associated with increased autophagy and requires autophagy genes APG1, APG7, APG8, and AUT10 (Melendez et al., 2003). Accordingly, increased autophagy is essential for lifespan extension in dietary restriction conditions or TOR inhibition in C. elegans (Hansen et al., 2008). But, whereas DAF‐2 mutants require both autophagy and the transcription factor DAF‐16/FoxO to promote longevity, autophagy takes place in the absence of DAF‐16/FoxO. This may suggest that autophagy provides raw material for new macromolecular synthesis that requires the action of DAF‐16/FoxO, recycling this material into cell‐protective longevity proteins (Hansen et al., 2008). In Salmonella‐infected worms, inactivation of the autophagy pathway was shown to increase bacterial intracellular replication, reducing animal lifespan, culminating in an apoptotic‐independent death (Jia et al., 2009). Mutation of DAF‐2 or overexpression of the DAF‐16/FoxO conferred pathogen resistance, which is precluded with the genetic knockdown of autophagy genes (Jia et al., 2009).
Lifespan in C. elegans can also be modulated by its reproductive system. The removal of the worm′s germ cells extends its life by 60% (Hsin & Kenyon, 1999; Kenyon, 2010b). Germ line loss stimulates nuclear accumulation of DAF‐16/FoxO and the expression of the transcription elongation/splicing factor homolog TCER‐1 in intestinal cells. Although the precise mechanism of how the information about the reproductive status is transferred to the intestine is unclear, it has been shown that the intestinal adaptor protein KRI‐1 is required to induce DAF‐16/FoxO target gene expression and extend lifespan (Berman & Kenyon, 2006; Ganapathy et al., 2010). This pathway is apparently independent of the IIS and, accordingly, in daf‐2 mutants, the loss of germ line acts additively with the IIS to expand the long lifespan of these mutants. Wang et al. (2008) proposed that this lifespan expansion may be related to lipid metabolism, by showing that C. elegans germ line stem cells actively promote systemic lipolysis via induction of a specific triglyceride lipase, identified as lipl‐4. As DAF‐16/FoxO upregulates this triglyceride lipase in response to germ line removal, it has been suggested it may be involved in the pathway that allows DAF‐16/FoxO activity in the worm's intestine/adipose tissue to influence C. elegans lifespan (Wang et al., 2008; Kenyon, 2010a).
Drosophila melanogaster
Drosophila melanogaster, or fruit fly, is an invertebrate of the taxonomic order Diptera. It has a lifespan of about 30 days and a four‐stage life cycle: egg, larva, pupa, and adult. Since 2001, it is known that the downregulation of IIS could extend lifespan in the fruit fly D.melanogaster, establishing the evolutionary conserved role of this pathway in aging (Clancy et al., 2001; Tatar et al., 2001). Mutation of Drosophila InR (insulin‐like receptor) homologous to mammalian insulin receptors yields dwarf females with up to an 85% extension of adult longevity and dwarf males with reduced late age‐specific mortality (Clancy et al., 2001; Tatar et al., 2001). Conversely, treating these long‐lived InR‐mutated dwarfs with a juvenile hormone analog restores life expectancy toward that of wild‐type controls (Clancy et al., 2001; Tatar et al., 2001). Drosophila gene chico (named after the small size of the correspondent mutants) encodes an insulin receptor substrate that functions in the IIS pathway. Mutation of this gene extends fruit fly median lifespan by up to 48% in homozygotes and 36% in heterozygotes (Clancy et al., 2001). Importantly, this lifespan extension was not a result of impaired oogenesis in chico females nor was correlated with increased stress resistance (Clancy et al., 2001).
dFoxO, the equivalent of nematode DAF‐16/FoxO and mammalian FOXO3A, has been shown to be a key transcriptional regulator of the insulin pathway that modulates growth and proliferation in D.melanogaster (Puig & Mattila, 2011). Without ligand binding at the insulin‐like receptor, dFoxO remains unphosphorylated and translocated to the nucleus, activating expression of factors that retard cell growth and proliferation (Junger et al., 2003; Puig et al., 2003). On the other hand, insulin treatment leads to dFoxO phosphorylation by dAKT, leading to cytoplasmic retention and inhibition of its transcriptional activity. Mutant dFoxO lacking dAKT phosphorylation sites does not respond to insulin inhibition and is constitutively active in the nucleus (Puig et al., 2003). dFoxO activation induces growth arrest and increases the expression of two key players of the dInR/dPI3K/dAKT pathway: the translational regulator d4E‐BP and the dInR itself. Interestingly, targeted expression of dFoxO in fly tissues regulates organ size by specifying cell number with no effect on cell size (Puig et al., 2003).
As in the worm, the nervous system has been shown to be implicated in the IIS‐mediated extension of lifespan in Drosophila (Fontana et al., 2010). Drosophila has seven genes encoding insulin‐like peptides (dilps), and ablation of the cells expressing only three of the seven dilps in neuroendocrine cells of the brain is sufficient to increase longevity (Broughton et al., 2008). dFoxO has also been shown to regulate D. melanogaster aging when activated specifically in the adult pericerebral fat body. This limited activation of dFoxO reduces expression of the peptide dilp‐2 synthesized in neurons and represses the IIS pathway in peripheral fat body. These findings suggest that autonomous and nonautonomous roles of insulin signaling concomitantly contribute to control aging (Hwangbo et al., 2004).
Dietary restriction has been shown to affect the expansion of lifespan in flies mediated by both TOR and IIS pathways (Kenyon, 2010b). The fall in nutrients leads to a diminished TOR activity, increasing lifespan by two possible mechanisms: by inhibiting general translation and increasing respiration or by enhancing autophagy (Kenyon, 2010b).
Interestingly, life extension through dietary restriction does not require dFoxO (Giannakou et al., 2008). Nevertheless, the overexpression of dFoxO in the adult fat body and gut of Drosophila showed an altered response to dietary restriction, behaving as partially dietary restricted. The authors suggest that, although dFoxO is unnecessary to extend lifespan of flies in response to dietary restriction, the presence of active dFoxO modulates the response to dietary restriction (Giannakou et al., 2008). This likely occurs by changing the expression of dFoxO‐target genes, indicating that dFoxO may mediate the normal response to dietary restriction (Giannakou et al., 2008). Min et al. (2008) showed evidence that the diet‐dependent effects of dFoxO overexpression on fly's lifespan are associated with reduction of dilp2.
The stress‐responsive JNK pathway also requires dFoxO to extend lifespan in Drosophila (Wang et al., 2005). JNK has been shown to antagonize IIS, promoting dFoxO nuclear localization, therefore inducing expression of growth control and stress defense genes. Moreover, the repression of IIS ligands by JNK and dFoxO in neuroendocrine cells systemically downregulates the IIS pathway (Wang et al., 2005).
As in C. elegans, dFoxO can regulate autophagy in Drosophila at an organismal level. Overexpression of dFoxO or the dFoxO‐target 4E‐BP in the muscle reduces protein aggregation in other tissues (brain, adipose tissue, and retina), delaying muscle functional decay and extending lifespan (Demontis & Perrimon, 2010). Moreover, dFoxO/4E‐BP overexpression in muscle decreases feeding behavior and the release of insulin, therefore delaying the age‐related accumulation of protein aggregates in other tissues. These results reveal a non‐cell‐autonomous mechanism mediated by dFoxO/4E‐BP signaling in the coordination of organismal and tissue aging (Demontis & Perrimon, 2010).
Mammals
The family of mammalian FOXO transcription factors comprises four members. FOXO1, FOXO3, and FOXO4 are highly related, share the same DNA‐binding motifs, present similar patterns of expression, and seem to have overlapping functions (Anderson et al., 1998; Biggs et al., 2001; Jacobs et al., 2003; Obsil & Obsilova, 2011). FOXO6 is mainly expressed in the brain and has been shown to be regulated by distinct mechanisms. Interestingly, knockouts of single Foxo genes in mice present with very distinct outcomes: Foxo1 knockout mice die in utero due to defects in vasculature (Furuyama et al., 2004; Hosaka et al., 2004). Female Foxo3 knockout mice were found to be sterile due to global primordial follicle activation with subsequent oocyte exhaustion (Castrillon et al., 2003; Hosaka et al., 2004). In addition, depletion of Foxo3 resulted in deficient development of regulatory T cells with consequent organ inflammation by a mechanism also involving Foxo1 (Harada et al., 2010; Kerdiles et al., 2010). Foxo4 and Foxo6 knockout mice present with only mild phenotypes (Zhu et al., 2011; Salih et al., 2012). Accordingly, conventional genetic analysis did not reveal an overt tumor‐prone or hematopoietic phenotype for the deficiency of any one of the Foxo family members. As closely related members of gene families may mask individual gene functions in single knockout experiments, conditional alleles for Foxo1, Foxo3, and/or Foxo4 have been generated. Paik et al. (2007) showed that triple knockout mice prompted a progressive cancer‐prone condition characterized by thymic lymphomas and hemangiomas which lead to early death. This study established mammalian Foxo proteins as bona fide tumor suppressors and confirming the functional redundancy of Foxo family members. However, it still remains to be determined why these animals did not present neoplastic phenotypes in epithelial tissues. Alternative downstream arms of the PI3K/AKT pathway might play a more prominent trumorigenic role in epithelial compartments. Conditional deletion of Foxo1, Foxo3, and Foxo4 in the adult hematopoietic system resulted in a marked context‐dependent increase in ROS in Foxo‐deficient HSC compared with wild‐type HSC that correlated with changes in expression of genes that regulate ROS. The number and the long‐term repopulating activity of HSC were found to be significantly reduced in these animals (Tothova et al., 2007). These results highlight the importance of mammalian Foxo proteins for the long‐term regenerative potential of HCS (Tothova & Gilliland, 2007). Ablation of the three Foxo proteins resulted in an increase in oxidative stress in bone and osteoblast apoptosis and a decrease in the number of osteoblasts, the rate of bone formation and bone mass (Ambrogini et al., 2010). Moreover, the overexpression of a Foxo3 transgene in mature osteoblasts decreased oxidative stress and osteoblast apoptosis and increased osteoblast number, bone formation rate, and vertebral bone mass. These authors concluded that Foxos provide an oxidative defense mechanism to deal with the aerobic metabolism of osteoblasts, being indispensable for bone mass homeostasis (Ambrogini et al., 2010). Numerous research groups have used the same approach to conditionally delete Foxo1, Foxo3, and Foxo4 in different tissues, but the effect in aged knockout mice has not been examined. There is no doubt that in the near future experiments will shed light on this central aspect of FOXO biology. Intriguingly, recent studies show that the life‐extending effect of dietary restriction requires Foxo3 but not Foxo1 in mice (Yamaza et al., 2010; Shimokawa et al., 2015). The mechanism(s) by which Foxo factors contribute to lifespan remain elusive. As previously mentioned, decreased autophagy has been related to premature aging and age‐related associated disorders (Hara et al., 2006; Komatsu et al., 2006; Jung et al., 2008; Pickford et al., 2008; Masiero et al., 2009; Lee et al., 2010a, 2010b). Foxo factors have been shown to regulate autophagy in mouse muscle, particularly Foxo3, which induces the expression of several autophagy genes and increases autophagosome formation (Mammucari et al., 2007; Zhao et al., 2007; Webb & Brunet, 2014). Foxo1 and Foxo3 overexpression also promotes mitophagy (degradation of mitochondria by the autophagy–lysosomal pathway) by upregulating the expression of the mouse mitochondrial E3 ubiquitin protein ligase 1 (Lokireddy et al., 2012). Remarkably, Foxos have also been implicated in the autophagy and mitophagy of neurons. FOXO3 has been shown to control the accumulation of human α‐synuclein, a protein known to participate in the development of Parkinson's disease (Pino et al., 2014). Mild FOXO3 activity protects nigral neurons against the accumulation of human α‐synuclein by promoting its degradation. These results suggest a determinant role for FOXO3 in Parkinson's disease, via neuronal survival in the substantia nigra (Kume et al., 2010). Furthermore, FoxO1 and FoxO3 factors have also shown to promote autophagy in rat neonatal cardiomyocytes and primary renal proximal tubular cells (Sengupta et al., 2009; Kume et al., 2010).
Warr et al. (2013) demonstrated that mouse HSCs robustly induce autophagy and identified Foxo3A as critically important for rapid induction of autophagy upon starvation. Interestingly, these authors also showed that old HSCs retain an intact Foxo3A‐driven pro‐autophagy gene program and that autophagy is required to maintain energy homeostasis and promote survival of the HSCs (Warr et al., 2013).
Aging is associated with a decreased proteasomal activity, leading to excess of damaged proteins in muscle, liver, and heart (Conconi et al., 1996; Petropoulos et al., 2000; Bulteau et al., 2002; Husom et al., 2004). Foxo3 has been shown to be a transcriptional regulator of muscle‐specific E3 ubiquitin ligases, which are major effectors of protein degradation in muscle (Sandri et al., 2004, 2006; Stitt et al., 2004). But FOXOs can also affect the composition of the proteasome. In particular, FOXO4 has been shown to be required for the expression of the proteasome component PSMD11 in human embryonic stem cells (Vilchez et al., 2012).
Conclusions
An exciting research area on FOXO transcription factors' impacting on longevity has arisen in recent years. Studies have been conducted to address their upstream regulation, their downstream effectors, and respective signaling pathways in various animal models. Consequently, how these FOXO‐mediated programs affect cellular or tissue function and whether there is an effect at an organismal level, has also been scrutinized. Several lines of evidence suggest that FOXOs affect longevity in a pleiotropic fashion, influencing several cell‐regulated activities such as stress resistance, metabolism, cell cycle arrest, and apoptosis.
A myriad of future work can be envisioned at this time. The induction of FOXO‐mediated programs in tissues with distinct metabolic potential such as brain, muscle, or adipose tissue and with different stages of differentiation or metabolic conditions (nutrition, oxidative stress) will enlarge our knowledge of how FOXO factors affect cellular/organismal lifespan. To further comprehend how FOXOs affect longevity, it is of high importance to understand how human FOXO sequence variants (namely FOXO3A) affect protein expression, its structure, or transcriptional activity. In order to see how these variants translate into physiological profiles, future investigations should address how these variants affect the level of FOXO proteins and their downstream effectors in serum. This approach has been used successfully in patients with vitiligo, in which FOXO3A levels were shown to be decreased when compared with the control group (Ozel Turkcu et al., 2014).
Compression of morbidity relates to both extended lifespan and delayed onset of age‐related diseases, such as cancer and cardiovascular disorders. The development of molecules targeting aging mechanisms that underlie a number of age‐related diseases is an exciting field that is nowadays in its first steps. It is noteworthy that clinical trials to test lifespan extension in humans would be challenging and require markers that can detect difference in aging rate across a short time frame. But given the potential of FOXO proteins to impact on numerous disorders such as cancer, diabetes, neurodegeneration, or immune system dysfunction, novel therapeutic modalities based on FOXOs will most likely take place in the near future.
Conflict of interest
None declared.
Acknowledgments
GJL and WL were supported with a subcontract from NIH award U19 AG023122.
References
- Ambrogini E, Almeida M, Martin‐Millan M, Paik JH, Depinho RA, Han L, Goellner J, Weinstein RS, Jilka RL, O'Brien CA, Manolagas SC (2010) FoxO‐mediated defense against oxidative stress in osteoblasts is indispensable for skeletal homeostasis in mice. Cell Metab. 11, 136–146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andersen SL, Sebastiani P, Dworkis DA, Feldman L, Perls TT (2012) Health span approximates life span among many supercentenarians: compression of morbidity at the approximate limit of life span. J. Gerontol. A Biol. Sci. Med. Sci. 67, 395–405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson MJ, Viars CS, Czekay S, Cavenee WK, Arden KC (1998) Cloning and characterization of three human forkhead genes that comprise an FKHR‐like gene subfamily. Genomics 47, 187–199. [DOI] [PubMed] [Google Scholar]
- Anselmi CV, Malovini A, Roncarati R, Novelli V, Villa F, Condorelli G, Bellazzi R, Puca AA (2009) Association of the FOXO3A locus with extreme longevity in a southern Italian centenarian study. Rejuvenation Res. 12, 95–104. [DOI] [PubMed] [Google Scholar]
- Apfeld J, O'Connor G, McDonagh T, DiStefano PS, Curtis R (2004) The AMP‐activated protein kinase AAK‐2 links energy levels and insulin‐like signals to lifespan in C. elegans . Genes Dev. 18, 3004–3009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arai Y, Hirose N, Yamamura K, Shimizu K, Takayama M, Ebihara Y, Osono Y (2001) Serum insulin‐like growth factor‐1 in centenarians: implications of IGF‐1 as a rapid turnover protein. J. Gerontol. A Biol. Sci. Med. Sci. 56, M79–M82. [DOI] [PubMed] [Google Scholar]
- Atzmon G, Schechter C, Greiner W, Davidson D, Rennert G, Barzilai N (2004) Clinical phenotype of families with longevity. J. Am. Geriatr. Soc. 52, 274–277. [DOI] [PubMed] [Google Scholar]
- Bao JM, Song XL, Hong YQ, Zhu HL, Li C, Zhang T, Chen W, Zhao SC, Chen Q (2014) Association between FOXO3A gene polymorphisms and human longevity: a meta‐analysis. Asian J. Androl. 16, 446–452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barzilai N, Huffman DM, Muzumdar RH, Bartke A (2012) The critical role of metabolic pathways in aging. Diabetes 61, 1315–1322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beekman M, Nederstigt C, Suchiman HE, Kremer D, van der Breggen R, Lakenberg N, Alemayehu WG, de Craen AJ, Westendorp RG, Boomsma DI, de Geus EJ, Houwing‐Duistermaat JJ, Heijmans BT, Slagboom PE (2010) Genome‐wide association study (GWAS)‐identified disease risk alleles do not compromise human longevity. Proc. Natl Acad. Sci. USA 107, 18046–18049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bellantuono AJ, Bridge D, Martinez DE (2015) Hydra as a tractable, long‐lived model system for senescence. Invertebr. Reprod. Dev. 59, 39–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berdichevsky A, Viswanathan M, Horvitz HR, Guarente L (2006) C. elegans SIR‐2.1 interacts with 14‐3‐3 proteins to activate DAF‐16 and extend life span. Cell 125, 1165–1177. [DOI] [PubMed] [Google Scholar]
- Berman JR, Kenyon C (2006) Germ‐cell loss extends C. elegans life span through regulation of DAF‐16 by kri‐1 and lipophilic‐hormone signaling. Cell 124, 1055–1068. [DOI] [PubMed] [Google Scholar]
- Biggs WH 3rd, Meisenhelder J, Hunter T, Cavenee WK, Arden KC (1999) Protein kinase B/Akt‐mediated phosphorylation promotes nuclear exclusion of the winged helix transcription factor FKHR1. Proc. Natl Acad. Sci. USA 96, 7421–7426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Biggs WH, Cavenee WK, Arden KC (2001) Identification and characterization of members of the FKHR (FOX O) subclass of winged‐helix transcription factors in the mouse. Mamm. Genome 12, 416–425. [DOI] [PubMed] [Google Scholar]
- Bishop NA, Guarente L (2007) Genetic links between diet and lifespan: shared mechanisms from yeast to humans. Nat. Rev. Genet. 8, 835–844. [DOI] [PubMed] [Google Scholar]
- Blagosklonny MV (2013a) Aging is not programmed: genetic pseudo‐program is a shadow of developmental growth. Cell Cycle 12, 3736–3742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blagosklonny MV (2013b) Rapamycin extends life‐ and health span because it slows aging. Aging (Albany NY) 5, 592–598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bodine SC, Latres E, Baumhueter S, Lai VK, Nunez L, Clarke BA, Poueymirou WT, Panaro FJ, Na E, Dharmarajan K, Pan ZQ, Valenzuela DM, DeChiara TM, Stitt TN, Yancopoulos GD, Glass DJ (2001) Identification of ubiquitin ligases required for skeletal muscle atrophy. Science (New York, NY) 294, 1704–1708. [DOI] [PubMed] [Google Scholar]
- Boehm AM, Khalturin K, Anton‐Erxleben F, Hemmrich G, Klostermeier UC, Lopez‐Quintero JA, Oberg HH, Puchert M, Rosenstiel P, Wittlieb J, Bosch TC (2012) FoxO is a critical regulator of stem cell maintenance in immortal Hydra . Proc. Natl Acad. Sci. USA 109, 19697–19702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonafe M, Barbieri M, Marchegiani F, Olivieri F, Ragno E, Giampieri C, Mugianesi E, Centurelli M, Franceschi C, Paolisso G (2003) Polymorphic variants of insulin‐like growth factor I (IGF‐I) receptor and phosphoinositide 3‐kinase genes affect IGF‐I plasma levels and human longevity: cues for an evolutionarily conserved mechanism of life span control. J. Clin. Endocrinol. Metab. 88, 3299–3304. [DOI] [PubMed] [Google Scholar]
- Bosch TCG (2009) Hydra and the evolution of stem cells. BioEssays 31, 478–486. [DOI] [PubMed] [Google Scholar]
- Bosch TC, Anton‐Erxleben F, Hemmrich G, Khalturin K (2010) The Hydra polyp: nothing but an active stem cell community. Dev. Growth Differ. 52, 15–25. [DOI] [PubMed] [Google Scholar]
- Boyette LB, Tuan RS (2014) Adult stem cells and diseases of aging. J. Clin. Med. 3, 88–134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bridge D, Theofiles AG, Holler RL, Marcinkevicius E, Steele RE, Martinez DE (2010) FoxO and stress responses in the cnidarian Hydra vulgaris. PLoS ONE 5, e11686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Broer L, Buchman AS, Deelen J, Evans DS, Faul JD, Lunetta KL, Sebastiani P, Smith JA, Smith AV, Tanaka T, Yu L, Arnold AM, Aspelund T, Benjamin EJ, De Jager PL, Eirkisdottir G, Evans DA, Garcia ME, Hofman A, Kaplan RC, Kardia SL, Kiel DP, Oostra BA, Orwoll ES, Parimi N, Psaty BM, Rivadeneira F, Rotter JI, Seshadri S, Singleton A, Tiemeier H, Uitterlinden AG, Zhao W, Bandinelli S, Bennett DA, Ferrucci L, Gudnason V, Harris TB, Karasik D, Launer LJ, Perls TT, Slagboom PE, Tranah GJ, Weir DR, Newman AB, van Duijn CM, Murabito JM (2015) GWAS of longevity in CHARGE consortium confirms APOE and FOXO3 candidacy. J. Gerontol. A Biol. Sci. Med. Sci. 70, 110–118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brooks‐Wilson AR (2013) Genetics of healthy aging and longevity. Hum. Genet. 132, 1323–1338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Broughton S, Alic N, Slack C, Bass T, Ikeya T, Vinti G, Tommasi AM, Driege Y, Hafen E, Partridge L (2008) Reduction of DILP2 in Drosophila triages a metabolic phenotype from lifespan revealing redundancy and compensation among DILPs. PLoS ONE 3, e3721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brunet A, Bonni A, Zigmond MJ, Lin MZ, Juo P, Hu LS, Anderson MJ, Arden KC, Blenis J, Greenberg ME (1999) Akt promotes cell survival by phosphorylating and inhibiting a Forkhead transcription factor. Cell 96, 857–868. [DOI] [PubMed] [Google Scholar]
- Bulteau A‐L, Szweda LI, Friguet B (2002) Age‐dependent declines in proteasome activity in the heart. Arch. Biochem. Biophys. 397, 298–304. [DOI] [PubMed] [Google Scholar]
- Calnan DR, Brunet A (2008) The FoxO code. Oncogene 27, 2276–2288. [DOI] [PubMed] [Google Scholar]
- Castrillon DH, Miao L, Kollipara R, Horner JW, DePinho RA (2003) Suppression of ovarian follicle activation in mice by the transcription factor Foxo3a. Science 301, 215–218. [DOI] [PubMed] [Google Scholar]
- Chen AT, Guo C, Itani OA, Budaitis BG, Williams TW, Hopkins CE, McEachin RC, Pande M, Grant AR, Yoshina S, Mitani S, Hu PJ (2015) Longevity genes revealed by integrative analysis of isoform‐specific daf‐16/FoxO mutants of Caenorhabditis elegans . Genetics 201, 613–629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Christensen K, Johnson TE, Vaupel JW (2006) The quest for genetic determinants of human longevity: challenges and insights. Nat. Rev. Genet. 7, 436–448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ciechanover A, Brundin P (2003) The ubiquitin proteasome system in neurodegenerative diseases: sometimes the chicken, sometimes the egg. Neuron 40, 427–446. [DOI] [PubMed] [Google Scholar]
- Clancy DJ, Gems D, Harshman LG, Oldham S, Stocker H, Hafen E, Leevers SJ, Partridge L (2001) Extension of life‐span by loss of CHICO, a Drosophila insulin receptor substrate protein. Science (New York, NY) 292, 104–106. [DOI] [PubMed] [Google Scholar]
- Colman RJ, Anderson RM, Johnson SC, Kastman EK, Kosmatka KJ, Beasley TM, Allison DB, Cruzen C, Simmons HA, Kemnitz JW, Weindruch R (2009) Caloric restriction delays disease onset and mortality in rhesus monkeys. Science (New York, NY) 325, 201–204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conconi M, Szweda LI, Levine RL, Stadtman ER, Friguet B (1996) Age‐related decline of rat liver multicatalytic proteinase activity and protection from oxidative inactivation by heat‐shock protein 90. Arch. Biochem. Biophys. 331, 232–240. [DOI] [PubMed] [Google Scholar]
- Demontis F, Perrimon N (2010) FOXO/4E‐BP signaling in Drosophila muscles regulates organism‐wide proteostasis during aging. Cell 143, 813–825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dong XC, Copps KD, Guo S, Li Y, Kollipara R, DePinho RA, White MF (2008) Inactivation of hepatic Foxo1 by insulin signaling is required for adaptive nutrient homeostasis and endocrine growth regulation. Cell Metab. 8, 65–76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Donlon TA, Curb JD, He Q, Grove JS, Masaki KH, Rodriguez B, Elliott A, Willcox DC, Willcox BJ (2012) FOXO3 gene variants and human aging: coding variants may not be key players. J. Gerontol. A Biol. Sci. Med. Sci. 67, 1132–1139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eijkelenboom A, Burgering BM (2013) FOXOs: signalling integrators for homeostasis maintenance. Nat. Rev. Mol. Cell Biol. 14, 83–97. [DOI] [PubMed] [Google Scholar]
- Essers MA, Weijzen S, de Vries‐Smits AM, Saarloos I, de Ruiter ND, Bos JL, Burgering BM (2004) FOXO transcription factor activation by oxidative stress mediated by the small GTPase Ral and JNK. EMBO J. 23, 4802–4812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fielenbach N, Antebi A (2008) C. elegans dauer formation and the molecular basis of plasticity. Genes Dev. 22, 2149–2165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flachsbart F, Caliebe A, Kleindorp R, Blanché H, von Eller‐Eberstein H, Nikolaus S, Schreiber S, Nebel A (2009) Association of FOXO3A variation with human longevity confirmed in German centenarians. Proc. Natl Acad. Sci. USA 106, 2700–2705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fontana L, Partridge L, Longo VD (2010) Extending healthy life span – from yeast to humans. Science 328, 321–326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fries JF (1980) Aging, natural death, and the compression of morbidity. N. Engl. J. Med. 303, 130–135. [DOI] [PubMed] [Google Scholar]
- Furuyama T, Kitayama K, Shimoda Y, Ogawa M, Sone K, Yoshida‐Araki K, Hisatsune H, Nishikawa S, Nakayama K, Ikeda K, Motoyama N, Mori N (2004) Abnormal angiogenesis in Foxo1 (Fkhr)‐deficient mice. J. Biol. Chem. 279, 34741–34749. [DOI] [PubMed] [Google Scholar]
- Ganapathy S, Chen Q, Singh KP, Shankar S, Srivastava RK (2010) Resveratrol enhances antitumor activity of TRAIL in prostate cancer xenografts through activation of FOXO transcription factor. PLoS ONE 5, e15627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garigan D, Hsu A‐L, Fraser AG, Kamath RS, Ahringer J, Kenyon C (2002) Genetic analysis of tissue aging in Caenorhabditis elegans: a role for heat‐shock factor and bacterial proliferation. Genetics 161, 1101–1112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Genin EC, Caron N, Vandenbosch R, Nguyen L, Malgrange B (2014) Concise review: forkhead pathway in the control of adult neurogenesis. Stem Cells 32, 1398–1407. [DOI] [PubMed] [Google Scholar]
- Giannakou ME, Goss M, Partridge L (2008) Role of dFOXO in lifespan extension by dietary restriction in Drosophila melanogaster: not required, but its activity modulates the response. Aging Cell 7, 187–198. [DOI] [PubMed] [Google Scholar]
- Gil EB, Malone Link E, Liu LX, Johnson CD, Lees JA (1999) Regulation of the insulin‐like developmental pathway of Caenorhabditis elegans by a homolog of the PTEN tumor suppressor gene. Proc. Natl Acad. Sci. USA 96, 2925–2930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gottlieb S, Ruvkun G (1994) daf‐2, daf‐16 and daf‐23: genetically interacting genes controlling Dauer formation in Caenorhabditis elegans . Genetics 137, 107–120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greer EL, Dowlatshahi D, Banko MR, Villen J, Hoang K, Blanchard D, Gygi SP, Brunet A (2007a) An AMPK‐FOXO pathway mediates longevity induced by a novel method of dietary restriction in C. elegans . Curr. Biol. 17, 1646–1656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greer EL, Oskoui PR, Banko MR, Maniar JM, Gygi MP, Gygi SP, Brunet A (2007b) The energy sensor AMP‐activated protein kinase directly regulates the mammalian FOXO3 transcription factor. J. Biol. Chem. 282, 30107–30119. [DOI] [PubMed] [Google Scholar]
- Guo S, Rena G, Cichy S, He X, Cohen P, Unterman T (1999) Phosphorylation of serine 256 by protein kinase B disrupts transactivation by FKHR and mediates effects of insulin on insulin‐like growth factor‐binding protein‐1 promoter activity through a conserved insulin response sequence. J. Biol. Chem. 274, 17184–17192. [DOI] [PubMed] [Google Scholar]
- Hannenhalli S, Kaestner KH (2009) The evolution of Fox genes and their role in development and disease. Nat. Rev. Genet. 10, 233–240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hansen M, Taubert S, Crawford D, Libina N, Lee S‐J, Kenyon C (2007) Lifespan extension by conditions that inhibit translation in Caenorhabditis elegans . Aging Cell 6, 95–110. [DOI] [PubMed] [Google Scholar]
- Hansen M, Chandra A, Mitic LL, Onken B, Driscoll M, Kenyon C (2008) A role for autophagy in the extension of lifespan by dietary restriction in C. elegans . PLoS Genet. 4, e24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hara T, Nakamura K, Matsui M, Yamamoto A, Nakahara Y, Suzuki‐Migishima R, Yokoyama M, Mishima K, Saito I, Okano H, Mizushima N (2006) Suppression of basal autophagy in neural cells causes neurodegenerative disease in mice. Nature 441, 885–889. [DOI] [PubMed] [Google Scholar]
- Harada Y, Elly C, Ying G, Paik JH, DePinho RA, Liu YC (2010) Transcription factors Foxo3a and Foxo1 couple the E3 ligase Cbl‐b to the induction of Foxp3 expression in induced regulatory T cells. J. Exp. Med. 207, 1381–1391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heidler T, Hartwig K, Daniel H, Wenzel U (2010) Caenorhabditis elegans lifespan extension caused by treatment with an orally active ROS‐generator is dependent on DAF‐16 and SIR‐2.1. Biogerontology 11, 183–195. [DOI] [PubMed] [Google Scholar]
- Henderson ST, Johnson TE (2001) daf‐16 integrates developmental and environmental inputs to mediate aging in the nematode Caenorhabditis elegans . Curr. Biol. 11, 1975–1980. [DOI] [PubMed] [Google Scholar]
- Herskind AM, McGue M, Holm NV, Sörensen TI, Harvald B, Vaupel JW (1996) The heritability of human longevity: a population‐based study of 2872 Danish twin pairs born 1870‐1900. Hum. Genet. 97, 319–323. [DOI] [PubMed] [Google Scholar]
- Hill R, Kalathur RK, Callejas S, Colaco L, Brandao R, Serelde B, Cebria A, Blanco‐Aparicio C, Pastor J, Futschik M, Dopazo A, Link W (2014) A novel phosphatidylinositol 3‐kinase (PI3K) inhibitor directs a potent FOXO‐dependent, p53‐independent cell cycle arrest phenotype characterized by the differential induction of a subset of FOXO‐regulated genes. Breast Cancer Res. 16, 482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hitt R, Young‐Xu Y, Silver M, Perls T (1999) Centenarians: the older you get, the healthier you have been. Lancet 354, 652. [DOI] [PubMed] [Google Scholar]
- Hjelmborg JV, Iachine I, Skytthe A, Vaupel JW, McGue M, Koskenvuo M, Kaprio J, Pedersen NL, Christensen K (2006) Genetic influence on human lifespan and longevity. Hum. Genet. 119, 312–321. [DOI] [PubMed] [Google Scholar]
- Honjoh S, Yamamoto T, Uno M, Nishida E (2009) Signalling through RHEB‐1 mediates intermittent fasting‐induced longevity in C. elegans . Nature 457, 726–730. [DOI] [PubMed] [Google Scholar]
- Hosaka T, Biggs WH 3rd, Tieu D, Boyer AD, Varki NM, Cavenee WK, Arden KC (2004) Disruption of forkhead transcription factor (FOXO) family members in mice reveals their functional diversification. Proc. Natl Acad. Sci. USA 101, 2975–2980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsin H, Kenyon C (1999) Signals from the reproductive system regulate the lifespan of C. elegans . Nature 399, 362–366. [DOI] [PubMed] [Google Scholar]
- Hsu A‐L, Murphy CT, Kenyon C (2003) Regulation of aging and age‐related disease by DAF‐16 and heat‐shock factor. Science (New York, NY) 300, 1142–1145. [DOI] [PubMed] [Google Scholar]
- Husom AD, Peters EA, Kolling EA, Fugere NA, Thompson LV, Ferrington DA (2004) Altered proteasome function and subunit composition in aged muscle. Arch. Biochem. Biophys. 421, 67–76. [DOI] [PubMed] [Google Scholar]
- Hwangbo DS, Gershman B, Gersham B, Tu M‐P, Palmer M, Tatar M (2004) Drosophila dFOXO controls lifespan and regulates insulin signalling in brain and fat body. Nature 429, 562–566. [DOI] [PubMed] [Google Scholar]
- Jacobs FMJ, van der Heide LP, Wijchers PJEC, Burbach JPH, Hoekman MFM, Smidt MP (2003) FoxO6, a novel member of the FoxO class of transcription factors with distinct shuttling dynamics. J. Biol. Chem. 278, 35959–35967. [DOI] [PubMed] [Google Scholar]
- Jia K, Thomas C, Akbar M, Sun Q, Adams‐Huet B, Gilpin C, Levine B (2009) Autophagy genes protect against Salmonella typhimurium infection and mediate insulin signaling‐regulated pathogen resistance. Proc. Natl Acad. Sci. USA 106, 14564–14569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jung HS, Chung KW, Won Kim J, Kim J, Komatsu M, Tanaka K, Nguyen YH, Kang TM, Yoon K‐H, Kim J‐W, Jeong YT, Han MS, Lee M‐K, Kim K‐W, Shin J, Lee M‐S (2008) Loss of autophagy diminishes pancreatic beta cell mass and function with resultant hyperglycemia. Cell Metab. 8, 318–324. [DOI] [PubMed] [Google Scholar]
- Junger MA, Rintelen F, Stocker H, Wasserman JD, Vegh M, Radimerski T, Greenberg ME, Hafen E (2003) The Drosophila forkhead transcription factor FOXO mediates the reduction in cell number associated with reduced insulin signaling. J. Biol. 2, 20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaeberlein M, Powers RW, Steffen KK, Westman EA, Hu D, Dang N, Kerr EO, Kirkland KT, Fields S, Kennedy BK (2005) Regulation of yeast replicative life span by TOR and Sch9 in response to nutrients. Science (New York, NY) 310, 1193–1196. [DOI] [PubMed] [Google Scholar]
- Kaestner KH, Knochel W, Martinez DE (2000) Unified nomenclature for the winged helix/forkhead transcription factors. Genes Dev. 14, 142–146. [PubMed] [Google Scholar]
- Kenyon C (2005) The plasticity of aging: insights from long‐lived mutants. Cell 120, 449–460. [DOI] [PubMed] [Google Scholar]
- Kenyon C (2010a) A pathway that links reproductive status to lifespan in Caenorhabditis elegans . Ann. N. Y. Acad. Sci. 1204, 156–162. [DOI] [PubMed] [Google Scholar]
- Kenyon CJ (2010b) The genetics of ageing. Nature 464, 504–512. [DOI] [PubMed] [Google Scholar]
- Kenyon C, Chang J, Gensch E, Rudner A, Tabtiang R (1993) A C. elegans mutant that lives twice as long as wild type. Nature 366, 461–464. [DOI] [PubMed] [Google Scholar]
- Kerdiles YM, Stone EL, Beisner DR, McGargill MA, Ch'en IL, Stockmann C, Katayama CD, Hedrick SM (2010) Foxo transcription factors control regulatory T cell development and function. Immunity 33, 890–904. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khalturin K, Anton‐Erxleben F, Milde S, Plötz C, Jr Wittlieb, Hemmrich G, Bosch TCG (2007) Transgenic stem cells in Hydra reveal an early evolutionary origin for key elements controlling self‐renewal and differentiation. Dev. Biol. 309, 32–44. [DOI] [PubMed] [Google Scholar]
- Kikis EA, Gidalevitz T, Morimoto RI (2010) Protein homeostasis in models of aging and age‐related conformational disease. Adv. Exp. Med. Biol. 694, 138–159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim J, Ishihara N, Lee TR (2014) A DAF‐16/FoxO3a‐dependent longevity signal is initiated by antioxidants. BioFactors 40, 247–257. [DOI] [PubMed] [Google Scholar]
- Kimura KD, Tissenbaum HA, Liu Y, Ruvkun G (1997) daf‐2, an insulin receptor‐like gene that regulates longevity and diapause in Caenorhabditis elegans . Science (New York, NY) 277, 942–946. [DOI] [PubMed] [Google Scholar]
- Kojima T, Kamei H, Aizu T, Arai Y, Takayama M, Nakazawa S, Ebihara Y, Inagaki H, Masui Y, Gondo Y, Sakaki Y, Hirose N (2004) Association analysis between longevity in the Japanese population and polymorphic variants of genes involved in insulin and insulin‐like growth factor 1 signaling pathways. Exp. Gerontol. 39, 1595–1598. [DOI] [PubMed] [Google Scholar]
- Komatsu M, Waguri S, Chiba T, Murata S, Iwata J‐I, Tanida I, Ueno T, Koike M, Uchiyama Y, Kominami E, Tanaka K (2006) Loss of autophagy in the central nervous system causes neurodegeneration in mice. Nature 441, 880–884. [DOI] [PubMed] [Google Scholar]
- Kops GJ, Dansen TB, Polderman PE, Saarloos I, Wirtz KW, Coffer PJ, Huang TT, Bos JL, Medema RH, Burgering BM (2002) Forkhead transcription factor FOXO3a protects quiescent cells from oxidative stress. Nature 419, 316–321. [DOI] [PubMed] [Google Scholar]
- Kume S, Uzu T, Horiike K, Chin‐Kanasaki M, Isshiki K, Araki S, Sugimoto T, Haneda M, Kashiwagi A, Koya D (2010) Calorie restriction enhances cell adaptation to hypoxia through Sirt1‐dependent mitochondrial autophagy in mouse aged kidney. J. Clin. Invest. 120, 1043–1055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuningas M, Putters ML, Westendorp RGJ, Slagboom PE, van Heemst D (2007) SIRT1 gene, age‐related diseases, and mortality: the Leiden 85‐plus study. J. Gerontol. A Biol. Sci. Med. Sci. 62, 960–965. [DOI] [PubMed] [Google Scholar]
- Kwon ES, Narasimhan SD, Yen K, Tissenbaum HA (2010) A new DAF‐16 isoform regulates longevity. Nature 466, 498–502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lasi M, David CN, Bottger A (2010) Apoptosis in pre‐Bilaterians: Hydra as a model. Apoptosis 15, 269–278. [DOI] [PubMed] [Google Scholar]
- Lee RY, Hench J, Ruvkun G (2001) Regulation of C. elegans DAF‐16 and its human ortholog FKHRL1 by the daf‐2 insulin‐like signaling pathway. Curr. Biol. 11, 1950–1957. [DOI] [PubMed] [Google Scholar]
- Lee SS, Kennedy S, Tolonen AC, Ruvkun G (2003) DAF‐16 target genes that control C. elegans life‐span and metabolism. Science (New York, NY) 300, 644–647. [DOI] [PubMed] [Google Scholar]
- Lee S‐J, Murphy CT, Kenyon C (2009) Glucose shortens the life span of C. elegans by downregulating DAF‐16/FOXO activity and aquaporin gene expression. Cell Metab. 10, 379–391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee JH, Budanov AV, Park EJ, Birse R, Kim TE, Perkins GA, Ocorr K, Ellisman MH, Bodmer R, Bier E, Karin M (2010a) Sestrin as a feedback inhibitor of TOR that prevents age‐related pathologies. Science (New York, NY) 327, 1223–1228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee JH, Budanov AV, Park EJ, Birse R, Kim TE, Perkins GA, Ocorr K, Ellisman MH, Bodmer R, Bier E, Karin M (2010b) Sestrin as a feedback inhibitor of TOR that prevents age‐related pathologies. Science 327, 1223–1228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li Y, Wang W‐JJ, Cao H, Lu J, Wu C, Hu F‐YY, Guo J, Zhao L, Yang F, Zhang Y‐XX, Li W, Zheng G‐YY, Cui H, Chen X, Zhu Z, He H, Dong B, Mo X, Zeng Y, Tian X‐LL (2009) Genetic association of FOXO1A and FOXO3A with longevity trait in Han Chinese populations. Hum. Mol. Genet. 18, 4897–4904. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Libina N, Berman JR, Kenyon C (2003) Tissue‐specific activities of C. elegans DAF‐16 in the regulation of lifespan. Cell 115, 489–502. [DOI] [PubMed] [Google Scholar]
- Lin K, Dorman JB, Rodan A, Kenyon C (1997) daf‐16: an HNF‐3/forkhead family member that can function to double the life‐span of Caenorhabditis elegans . Science (New York, NY) 278, 1319–1322. [DOI] [PubMed] [Google Scholar]
- Lin K, Hsin H, Libina N, Kenyon C (2001) Regulation of the Caenorhabditis elegans longevity protein DAF‐16 by insulin/IGF‐1 and germline signaling. Nat. Genet. 28, 139–145. [DOI] [PubMed] [Google Scholar]
- Lokireddy S, Wijesoma IW, Teng S, Bonala S, Gluckman PD, McFarlane C, Sharma M, Kambadur R (2012) The ubiquitin ligase Mul1 induces mitophagy in skeletal muscle in response to muscle‐wasting stimuli. Cell Metab. 16, 613–624. [DOI] [PubMed] [Google Scholar]
- Lopez‐Otin C, Blasco MA, Partridge L, Serrano M, Kroemer G (2013) The hallmarks of aging. Cell 153, 1194–1217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mammucari C, Milan G, Romanello V, Masiero E, Rudolf R, Del Piccolo P, Burden SJ, Di Lisi R, Sandri C, Zhao J, Goldberg AL, Schiaffino S, Sandri M (2007) FoxO3 controls autophagy in skeletal muscle in vivo. Cell Metab. 6, 458–471. [DOI] [PubMed] [Google Scholar]
- Masiero E, Agatea L, Mammucari C, Blaauw B, Loro E, Komatsu M, Metzger D, Reggiani C, Schiaffino S, Sandri M (2009) Autophagy is required to maintain muscle mass. Cell Metab. 10, 507–515. [DOI] [PubMed] [Google Scholar]
- Mattison JA, Roth GS, Beasley TM, Tilmont EM, Handy AM, Herbert RL, Longo DL, Allison DB, Young JE, Bryant M, Barnard D, Ward WF, Qi W, Ingram DK, de Cabo R (2012) Impact of caloric restriction on health and survival in rhesus monkeys from the NIA study. Nature 489, 318–321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McElwee J, Bubb K, Thomas JH (2003) Transcriptional outputs of the Caenorhabditis elegans forkhead protein DAF‐16. Aging Cell 2, 111–121. [DOI] [PubMed] [Google Scholar]
- Melendez A, Talloczy Z, Seaman M, Eskelinen EL, Hall DH, Levine B (2003) Autophagy genes are essential for dauer development and life‐span extension in C. elegans . Science 301, 1387–1391. [DOI] [PubMed] [Google Scholar]
- Mercken EM, Crosby SD, Lamming DW, JeBailey L, Krzysik‐Walker S, Villareal DT, Capri M, Franceschi C, Zhang Y, Becker K, Sabatini DM, de Cabo R, Fontana L (2013) Calorie restriction in humans inhibits the PI3K/AKT pathway and induces a younger transcription profile. Aging Cell 12, 645–651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mihaylova VT, Borland CZ, Manjarrez L, Stern MJ, Sun H (1999) The PTEN tumor suppressor homolog in Caenorhabditis elegans regulates longevity and dauer formation in an insulin receptor‐like signaling pathway. Proc. Natl Acad. Sci. USA 96, 7427–7432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Milman S, Atzmon G, Huffman DM, Wan J, Crandall JP, Cohen P, Barzilai N (2014) Low insulin‐like growth factor‐1 level predicts survival in humans with exceptional longevity. Aging Cell 13, 769–771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Min K‐JJ, Yamamoto R, Buch S, Pankratz M, Tatar M (2008) Drosophila lifespan control by dietary restriction independent of insulin‐like signaling. Aging Cell 7, 199–206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miyamoto K, Araki KY, Naka K, Arai F, Takubo K, Yamazaki S, Matsuoka S, Miyamoto T, Ito K, Ohmura M, Chen C, Hosokawa K, Nakauchi H, Nakayama K, Nakayama KI, Harada M, Motoyama N, Suda T, Hirao A (2007) Foxo3a is essential for maintenance of the hematopoietic stem cell pool. Cell Stem Cell 1, 101–112. [DOI] [PubMed] [Google Scholar]
- Morley JF, Morimoto RI (2004) Regulation of longevity in Caenorhabditis elegans by heat shock factor and molecular chaperones. Mol. Biol. Cell 15, 657–664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morley JF, Brignull HR, Weyers JJ, Morimoto RI (2002) The threshold for polyglutamine‐expansion protein aggregation and cellular toxicity is dynamic and influenced by aging in Caenorhabditis elegans . Proc. Natl Acad. Sci. USA 99, 10417–10422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murabito JM, Yuan R, Lunetta KL (2012) The search for longevity and healthy aging genes: insights from epidemiological studies and samples of long‐lived individuals. J. Gerontol. A Biol. Sci. Med. Sci. 67, 470–479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murphy CT, McCarroll SA, Bargmann CI, Fraser A, Kamath RS, Ahringer J, Li H, Kenyon C (2003) Genes that act downstream of DAF‐16 to influence the lifespan of Caenorhabditis elegans . Nature 424, 277–283. [DOI] [PubMed] [Google Scholar]
- Murphy CT, Lee S‐J, Kenyon C (2007) Tissue entrainment by feedback regulation of insulin gene expression in the endoderm of Caenorhabditis elegans . Proc. Natl Acad. Sci. USA 104, 19046–19050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nemoto S, Finkel T (2002) Redox regulation of forkhead proteins through a p66shc‐dependent signaling pathway. Science 295, 2450–2452. [DOI] [PubMed] [Google Scholar]
- Obsil T, Obsilova V (2011) Structural basis for DNA recognition by FOXO proteins. Biochim. Biophys. Acta 1813, 1946–1953. [DOI] [PubMed] [Google Scholar]
- Ogg S, Ruvkun G (1998) The C. elegans PTEN homolog, DAF‐18, acts in the insulin receptor‐like metabolic signaling pathway. Mol. Cell 2, 887–893. [DOI] [PubMed] [Google Scholar]
- Ogg S, Paradis S, Gottlieb S, Patterson GI, Lee L, Tissenbaum HA, Ruvkun G (1997) The Fork head transcription factor DAF‐16 transduces insulin‐like metabolic and longevity signals in C. elegans . Nature 389, 994–999. [DOI] [PubMed] [Google Scholar]
- Ookuma S, Fukuda M, Nishida E (2003) Identification of a DAF‐16 transcriptional target gene, scl‐1, that regulates longevity and stress resistance in Caenorhabditis elegans . Curr. Biol. 13, 427–431. [DOI] [PubMed] [Google Scholar]
- Ozel Turkcu U, Solak Tekin N, Gokdogan Edgunlu T, Karakas Celik S, Oner S (2014) The association of FOXO3A gene polymorphisms with serum FOXO3A levels and oxidative stress markers in vitiligo patients. Gene 536, 129–134. [DOI] [PubMed] [Google Scholar]
- Paik JH, Kollipara R, Chu G, Ji H, Xiao Y, Ding Z, Miao L, Tothova Z, Horner JW, Carrasco DR, Jiang S, Gilliland DG, Chin L, Wong WH, Castrillon DH, DePinho R (2007) FoxOs are lineage‐restricted redundant tumor suppressors and regulate endothelial cell homeostasis. Cell 128, 309–323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pan KZ, Palter JE, Rogers AN, Olsen A, Chen D, Lithgow GJ, Kapahi P (2007) Inhibition of mRNA translation extends lifespan in Caenorhabditis elegans . Aging Cell 6, 111–119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pawlikowska L, Hu D, Huntsman S, Sung A, Chu C, Chen J, Joyner AH, Schork NJ, Hsueh W‐CC, Reiner AP, Psaty BM, Atzmon G, Barzilai N, Cummings SR, Browner WS, Kwok P‐YY, Ziv E (2009) Association of common genetic variation in the insulin/IGF1 signaling pathway with human longevity. Aging Cell 8, 460–472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perls T, Shea‐Drinkwater M, Bowen‐Flynn J, Ridge SB, Kang S, Joyce E, Daly M, Brewster SJ, Kunkel L, Puca AA (2000) Exceptional familial clustering for extreme longevity in humans. J. Am. Geriatr. Soc. 48, 1483–1485. [PubMed] [Google Scholar]
- Perls TT, Wilmoth J, Levenson R, Drinkwater M, Cohen M, Bogan H, Joyce E, Brewster S, Kunkel L, Puca A (2002) Life‐long sustained mortality advantage of siblings of centenarians. Proc. Natl Acad. Sci. USA 99, 8442–8447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petropoulos I, Conconi M, Wang X, Hoenel B, Brégégère F, Milner Y, Friguet B (2000) Increase of oxidatively modified protein is associated with a decrease of proteasome activity and content in aging epidermal cells. J. Gerontol. A Biol. Sci. Med. Sci. 55, B220–B227. [DOI] [PubMed] [Google Scholar]
- Pickford F, Masliah E, Britschgi M, Lucin K, Narasimhan R, Jaeger PA, Small S, Spencer B, Rockenstein E, Levine B, Wyss‐Coray T (2008) The autophagy‐related protein beclin 1 shows reduced expression in early Alzheimer disease and regulates amyloid beta accumulation in mice. J. Clin. Invest. 118, 2190–2199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pino E, Amamoto R, Zheng L, Cacquevel M, Sarria JC, Knott GW, Schneider BL (2014) FOXO3 determines the accumulation of alpha‐synuclein and controls the fate of dopaminergic neurons in the substantia nigra. Hum. Mol. Genet. 23, 1435–1452. [DOI] [PubMed] [Google Scholar]
- Puig O, Mattila J (2011) Understanding Forkhead box class O function: lessons from Drosophila melanogaster. Antioxid. Redox Signal. 14, 635–647. [DOI] [PubMed] [Google Scholar]
- Puig O, Marr MT, Ruhf ML, Tjian R (2003) Control of cell number by Drosophila FOXO: downstream and feedback regulation of the insulin receptor pathway. Genes Dev. 17, 2006–2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Putker M, Madl T, Vos HR, de Ruiter H, Visscher M, van den Berg MC, Kaplan M, Korswagen HC, Boelens R, Vermeulen M, Burgering BM, Dansen TB (2013) Redox‐dependent control of FOXO/DAF‐16 by transportin‐1. Mol. Cell 49, 730–742. [DOI] [PubMed] [Google Scholar]
- Rouault JP, Kuwabara PE, Sinilnikova OM, Duret L, Thierry‐Mieg D, Billaud M (1999) Regulation of dauer larva development in Caenorhabditis elegans by daf‐18, a homologue of the tumour suppressor PTEN. Curr. Biol. 9, 329–332. [DOI] [PubMed] [Google Scholar]
- Salih DA, Rashid AJ, Colas D, de la Torre‐Ubieta L, Zhu RP, Morgan AA, Santo EE, Ucar D, Devarajan K, Cole CJ, Madison DV, Shamloo M, Butte AJ, Bonni A, Josselyn SA, Brunet A (2012) FoxO6 regulates memory consolidation and synaptic function. Genes Dev. 26, 2780–2801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sandri M, Sandri C, Gilbert A, Skurk C, Calabria E, Picard A, Walsh K, Schiaffino S, Lecker SH, Goldberg AL (2004) Foxo transcription factors induce the atrophy‐related ubiquitin ligase atrogin‐1 and cause skeletal muscle atrophy. Cell 117, 399–412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sandri M, Lin J, Handschin C, Yang W, Arany ZP, Lecker SH, Goldberg AL, Spiegelman BM (2006) PGC‐1alpha protects skeletal muscle from atrophy by suppressing FoxO3 action and atrophy‐specific gene transcription. Proc. Natl Acad. Sci. USA 103, 16260–16265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schaible R, Sussman M (2013) FOXO in aging: did evolutionary diversification of FOXO function distract it from prolonging life? BioEssays 35, 1101–1110. [DOI] [PubMed] [Google Scholar]
- Sebastiani P, Perls TT (2012) The genetics of extreme longevity: lessons from the new England centenarian study. Front Genet. 3, 277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sengupta A, Molkentin JD, Yutzey KE (2009) FoxO transcription factors promote autophagy in cardiomyocytes. J. Biol. Chem. 284, 28319–28331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sheaffer KL, Updike DL, Mango SE (2008) The Target of Rapamycin pathway antagonizes pha‐4/FoxA to control development and aging. Curr. Biol. 18, 1355–1364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shimokawa I, Komatsu T, Hayashi N, Kim SE, Kawata T, Park S, Hayashi H, Yamaza H, Chiba T, Mori R (2015) The life‐extending effect of dietary restriction requires Foxo3 in mice. Aging Cell 14, 707–709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soerensen M, Dato S, Christensen K, McGue M, Stevnsner T, Bohr VA, Christiansen L (2010) Replication of an association of variation in the FOXO3A gene with human longevity using both case‐control and longitudinal data. Aging Cell 9, 1010–1017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soerensen M, Nygaard M, Dato S, Stevnsner T, Bohr VA, Christensen K, Christiansen L (2015) Association study of FOXO3A SNPs and aging phenotypes in Danish oldest‐old individuals. Aging Cell 14, 60–66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stitt TN, Drujan D, Clarke BA, Panaro F, Timofeyva Y, Kline WO, Gonzalez M, Yancopoulos GD, Glass DJ (2004) The IGF‐1/PI3K/Akt pathway prevents expression of muscle atrophy‐induced ubiquitin ligases by inhibiting FOXO transcription factors. Mol. Cell 14, 395–403. [DOI] [PubMed] [Google Scholar]
- Storz P (2011) Forkhead homeobox type O transcription factors in the responses to oxidative stress. Antioxid. Redox Signal. 14, 593–605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suh Y, Atzmon G, Cho M‐O, Hwang D, Liu B, Leahy DJ, Barzilai N, Cohen P (2008) Functionally significant insulin‐like growth factor I receptor mutations in centenarians. Proc. Natl Acad. Sci. USA 105, 3438–3442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tatar M, Kopelman A, Epstein D, Tu MP, Yin CM, Garofalo RS (2001) A mutant Drosophila insulin receptor homolog that extends life‐span and impairs neuroendocrine function. Science (New York, NY) 292, 107–110. [DOI] [PubMed] [Google Scholar]
- Tazearslan C, Huang J, Barzilai N, Suh Y (2011) Impaired IGF1R signaling in cells expressing longevity‐associated human IGF1R alleles. Aging Cell 10, 551–554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tothova Z, Gilliland DG (2007) FoxO transcription factors and stem cell homeostasis: insights from the hematopoietic system. Cell Stem Cell 1, 140–152. [DOI] [PubMed] [Google Scholar]
- Tothova Z, Kollipara R, Huntly BJ, Lee BH, Castrillon DH, Cullen DE, McDowell EP, Lazo‐Kallanian S, Williams IR, Sears C, Armstrong SA, Passegue E, DePinho RA, Gilliland DG (2007) FoxOs are critical mediators of hematopoietic stem cell resistance to physiologic oxidative stress. Cell 128, 325–339. [DOI] [PubMed] [Google Scholar]
- Tsuchiya K, Westerterp M, Murphy AJ, Subramanian V, Ferrante AW Jr, Tall AR, Accili D (2013) Expanded granulocyte/monocyte compartment in myeloid‐specific triple FoxO knockout increases oxidative stress and accelerates atherosclerosis in mice. Circ. Res. 112, 992–1003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vilchez D, Boyer L, Morantte I, Lutz M, Merkwirth C, Joyce D, Spencer B, Page L, Masliah E, Berggren WT, Gage FH, Dillin A (2012) Increased proteasome activity in human embryonic stem cells is regulated by PSMD11. Nature 489, 304–308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walker GA, Lithgow GJ (2003) Lifespan extension in C. elegans by a molecular chaperone dependent upon insulin‐like signals. Aging Cell 2, 131–139. [DOI] [PubMed] [Google Scholar]
- Wang MC, Bohmann D, Jasper H (2005) JNK extends life span and limits growth by antagonizing cellular and organism‐wide responses to insulin signaling. Cell 121, 115–125. [DOI] [PubMed] [Google Scholar]
- Wang MC, O'Rourke EJ, Ruvkun G (2008) Fat metabolism links germline stem cells and longevity in C. elegans . Science (New York, NY) 322, 957–960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Warr MR, Binnewies M, Flach J, Reynaud D, Garg T, Malhotra R, Debnath J, Passegue E (2013) FOXO3A directs a protective autophagy program in haematopoietic stem cells. Nature 494, 323–327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Webb AE, Brunet A (2014) FOXO transcription factors: key regulators of cellular quality control. Trends Biochem. Sci. 39, 159–169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weinkove D, Halstead JR, Gems D, Divecha N (2006) Long‐term starvation and ageing induce AGE‐1/PI 3‐kinase‐dependent translocation of DAF‐16/FOXO to the cytoplasm. BMC Biol. 4, 1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wheeler HE, Kim SK (2011) Genetics and genomics of human ageing. Philos. Trans. R. Soc. Lond. B Biol. Sci. 366, 43–50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Willcox BJ, Willcox DC, He Q, Curb JD, Suzuki M (2006) Siblings of Okinawan centenarians share lifelong mortality advantages. J. Gerontol. A Biol. Sci. Med. Sci. 61, 345–354. [DOI] [PubMed] [Google Scholar]
- Willcox BJ, Donlon TA, He Q, Chen R, Grove JS, Yano K, Masaki KH, Willcox DC, Rodriguez B, Curb JD (2008) FOXO3A genotype is strongly associated with human longevity. Proc. Natl Acad. Sci. USA 105, 13987–13992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wittlieb JR, Khalturin K, Lohmann JU, Anton‐Erxleben F, Bosch TCG (2006) Transgenic Hydra allow in vivo tracking of individual stem cells during morphogenesis. Proc. Natl Acad. Sci. USA 103, 6208–6211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamaza H, Komatsu T, Wakita S, Kijogi C, Park S, Hayashi H, Chiba T, Mori R, Furuyama T, Mori N, Shimokawa I (2010) FoxO1 is involved in the antineoplastic effect of calorie restriction. Aging Cell 9, 372–382. [DOI] [PubMed] [Google Scholar]
- Zanella F, Link W, Carnero A (2010) Understanding FOXO, new views on old transcription factors. Curr. Cancer Drug Targets 10, 135–146. [DOI] [PubMed] [Google Scholar]
- Zeng Y, Cheng L, Chen H, Cao H, Hauser ER, Liu Y, Xiao Z, Tan Q, Tian XL, Vaupel JW (2010) Effects of FOXO genotypes on longevity: a biodemographic analysis. J. Gerontol. A Biol. Sci. Med. Sci. 65, 1285–1299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang X, Yalcin S, Lee DF, Yeh TY, Lee SM, Su J, Mungamuri SK, Rimmele P, Kennedy M, Sellers R, Landthaler M, Tuschl T, Chi NW, Lemischka I, Keller G, Ghaffari S (2011a) FOXO1 is an essential regulator of pluripotency in human embryonic stem cells. Nat. Cell Biol. 13, 1092–1099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang X, Zabinsky R, Teng Y, Cui M, Han M (2011b) microRNAs play critical roles in the survival and recovery of Caenorhabditis elegans from starvation‐induced L1 diapause. Proc. Natl Acad. Sci. USA 108, 17997–18002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao J, Brault JJ, Schild A, Cao P, Sandri M, Schiaffino S, Lecker SH, Goldberg AL (2007) FoxO3 coordinately activates protein degradation by the autophagic/lysosomal and proteasomal pathways in atrophying muscle cells. Cell Metab. 6, 472–483. [DOI] [PubMed] [Google Scholar]
- Zhu M, Zhang QJ, Wang L, Li H, Liu ZP (2011) FoxO4 inhibits atherosclerosis through its function in bone marrow derived cells. Atherosclerosis 219, 492–498. [DOI] [PMC free article] [PubMed] [Google Scholar]