

Abstract
The SDF-1-receptor CXCR4 has been associated with early metastasis and poorer prognosis in breast cancers, especially the most aggressive triple negative subtype. In line with previous reports, we found that tumoral CXCR4 expression in patients with locally advanced breast cancer was associated with increased metastases and rapid tumor progression. Moreover, high CXCR4 expression identified a group of bone marrow disseminated tumor cells (DTC) negative patients at high risk for metastasis and death. The Protein Epitope Mimetic (PEM) POL5551, a novel CXCR4 antagonist, inhibited binding of SDF-1 to CXCR4, had no direct effects on tumor cell viability, but reduced migration of breast cancer cells in vitro. In two orthotopic models of triple negative breast cancer, POL5551 had little inhibitory effect on primary tumor growth but significantly reduced distant metastasis. When combined with eribulin, a chemotherapeutic microtubule inhibitor, POL5551 additively reduced metastasis and prolonged survival in mice after resection of the primary tumor compared to single-agent eribulin. Hypothesizing that POL5551 may mobilize tumor cells from their microenvironment and sensitize them to chemotherapy, we utilized a “chemotherapy framing” dosing strategy. When administered shortly before and after eribulin treatment, 3 doses of POL5551 with eribulin reduced bone and liver tumor burden more effectively than chemotherapy alone. These data suggest that sequenced administration of CXCR4 antagonists with cytotoxic chemotherapy synergize to reduce distant metastases.
Introduction
The chemokine receptor CXCR4 plays an important role in the trafficking and homing of hematopoietic stem cells (HSCs) and leukocytes. Its ligand SDF-1 (also called CXCL12) is produced by activated osteoblasts, bone marrow and lung stromal cells, and endothelial cells (1). During embryogenesis, the CXCR4-expressing HSCs migrate from fetal liver and home to the bone marrow via chemotactic CXCR4/SDF-1 interactions (2). Mice with targeted genetic deletion of CXCR4 or SDF-1 are embryonic lethal due to impaired cardiovascular development and failure of HSC to home to the bone marrow (3–5). SDF-1 binding to CXCR4 also maintains the adult HSC niche; pharmacological antagonism of CXCR4 causes HSC to rapidly accumulate in the peripheral circulation (6).
Cancer cells, especially subpopulations with stem-like or metastatic properties, often utilize pathways typical of HSC and compete for their niche (7, 8). Accordingly, CXCR4 is overexpressed in a number of cancer types, including breast, lung, colon, glioblastoma and multiple myeloma (9). During metastasis, CXCR4 activation promotes the migration of tumor cells to SDF-1-rich distant organs, where they interact closely with resident stroma and extracellular matrix through both direct cell-to-cell contact (adhesion molecules) and soluble factors (chemokines, cytokines) (10–12). The establishment of these tumor/stromal interactions has been shown to promote tumor cell cycle arrest, survival and resistance to chemotherapy in hematological malignancies, such as MM and CML (particularly with MM (13)), and in solid tumors (14). Similar to CXCR4 inhibition in HSC, CXCR4 antagonists can mobilize tumor cells out of these protective stromal niches into the peripheral blood, which could enhance susceptibility to anoikis, and to conventional chemotherapy. In addition, CXCR4 antagonists may disrupt metastasis of tumors that use CXCR4 to home to bone and other SDF-1 rich distant organs. In contrast to its role in HSC and cancer cell mobilization, POL5551 and other CXCR4 inhibitors have also been shown to blunt mobilization of some other cell types, such as mesenchymal stem cells to sites of injury or inflammation, reducing for example neointimal formation (15, 16).
Breast cancer is the most prevalent cancer in women worldwide. Over 90% of the mortality of breast cancer patients is associated with metastasis and relapse. CXCR4 expression is detectable in breast cancer of various subtypes and has been associated with early metastasis and poorer prognosis, especially with triple negative breast cancer (TNBC), a particularly aggressive subtype (17). The presence of microscopic disseminated tumor cells (DTC) in the bone marrow is also associated with early metastasis and poorer survival (18). As CXCR4 can be expressed by both primary tumors and circulating tumor cells, the interaction between tumor CXCR4 and the niche-attractant SDF1 has been proposed to be an important player of early micrometastasis (19, 20).
Several CXCR4 antagonists have been developed for therapeutic applications including hematopoietic stem cell mobilization and as anti-tumor therapies (21). In preclinical studies small molecule inhibitors of CXCR4 induced prostate cancer cell mobilization in vivo (22) and decreased pulmonary metastases in models of both osteosarcoma (23) and melanoma (24). POL5551 (Polyphor Ltd, Basel Switzerland) is a novel fully synthetic cyclic peptide antagonist of CXCR4 developed using Protein Epitope Mimetic (PEM) technology (25). A representative structure of a PEM molecule incorporating a β-hairpin is shown in Supplemental Figure 1A. Like the FDA approved CXCR4 inhibitor, plerixafor, POL5551 competes with SDF-1 for the extracellular loop binding site of CXCR4 (pharmacology summarized in Supplemental Table 1). POL5551 has a higher affinity for CXCR4 and an increased HSC mobilization activity compared to plerixafor (26). At high doses in mice, POL5551 mobilized hematopoietic stem cells levels similar to that produced by G-CSF, a far greater mobilization than achieved with plerixafor, or that has been reported for other CXCR4 antagonists (26). In mouse models, POL5551 has been demonstrated to inhibit neointima hyperplasia in a model of atherosclerosis (27) and to prolong survival when added to anti-VEGF therapy in a model of glioblastoma (28).
In this study, we found that in stage II/III breast cancer patients that did not have detectable bone marrow DTC, tumoral CXCR4 expression could identify patients at risk for early mortality and metastasis. We hypothesized that antagonism of CXCR4 receptor with POL5551 would reduce metastases and improve survival in CXCR4 expressing breast cancer, and addressed this hypothesis in preclinical models. We found that POL5551 inhibited tumor cell migration and decreased adhesion-independent survival in vitro, although it had no direct effect on tumor cell viability. In agreement, single agent POL5551 reduced tumor cell metastasis to the distant sites outside the chest wall and lung, despite having no effect on the primary tumor growth, which suggests a disruption of the metastatic process rather than a direct anti-tumor effect. When combined with low-dose eribulin chemotherapy, POL5551 further decreased distant metastasis and prolonged survival in an orthotopic breast cancer model. We hypothesized that part of POL5551’s additive effects on chemotherapy could be explained by the displacement of cancer cells from chemoprotective niches in the tumor microenvironment. When administered shortly before and after eribulin, POL5551 treatment further reduced bone tumor burden. Our study suggests that combining CXCR4 antagonism to chemotherapy may be beneficial to treat triple negative breast cancer patients, and provides supporting evidence to a newly-initiated clinical trial with the CXCR4 antagonist POL6326 (ClinicalTrials.gov: NCT01837095).
Materials and Methods
Patient clinical trial information
A total 120 patients with newly diagnosed stage II–III breast cancer were enrolled on an open label randomized single-blind phase 2 trial (NCT00242203) to assess the efficacy of zoledronic acid in reducing disseminated tumor cells (DTC) and improving bone health. 81 patients who had evaluable data for both bone marrow DTC and primary tumor gene expression profiling were chosen for this study (Supplemental Table 3). Exclusion criteria included previous history of malignancy, Easter Cooperative Oncology Group (ECOG) score of 2 or higher, and evidence of distant metastases by CT scan (of pelvis, abdomen, and chest) or 99TC-MDP bone scan. The complete enrollment criteria and results of the study have been previously described (29, 30). Patients were monitored every 3–4 months for the first 2 years after enrollment, then every 6 months until year 5 and then annually thereafter. In the context of this trial, bone marrow aspirates were collected from each anterior iliac crest prior to treatment initiation, and DTC identified by cytokeratin staining as previously described (31). Patients were considered to be positive for DTC (DTCpos) if at least one CK-positive cell was isolated from either BM aspirate (31). Gene expression profiling was performed on the tumor biopsies of 81 of these patients (GEO accession #: GSE71258). A cut-off value for CXCR4 expression was selected utilizing the dedicated Cutoff Finder application (http://molpath.charite.de/cutoff/) (32) to analyze the entire set of 81 patients. Based on the distribution of primary tumor CXCR4 expression data, a cut-off value (10250) was determined: CXCR4 expression signal above 10250 was defined as CXCR4High (10 patients), while below 10250 was defined as CXCR4Low (28 patients). The overall survival by Kaplan-Meier curve and log-rank test was analyzed between CXCR4High and CXCR4Low patients. In order to investigate the association between CXCR4 expression and micrometastases at diagnosis, we then compared frequencies of DTCpos in CXCR4high vs. CXCR4low patients, and tested the association by Fisher’s exact test and odds ratio. In order to analyze the effect of CXCR4 on survival, we considered the gene expression profiling and DTC status of the 38/81 patients that were assigned to the placebo arm of the trial. Of these, 16 patients had bone marrow micrometastases. DTCneg (20 patients) were divided into CXCR4high (n=5) and CXCR4low (n=15) and analyzed for overall survival by Kaplan-Meier curve analysis and log-rank test.
Cell lines and reagents
The 4T1-FL-GFP BALB/c murine breast cancer cell line was generously provided by Dr. David Piwnica-Worms (The University of Texas MD Anderson Cancer Center, TX) and modified to co-express firefly luciferase and green fluorescent protein as previously described (33).
The bone metastatic MDA-MB-231 variant (34) was a kind gift of Dr. T. Guise (Indiana University, Bloomington, IN). CXCL12-CGLuc, NGLuc-CXCR4, and NGLuc-CXCR7 MDA-MB-231 cells were generated as previously described (35).
Cells were cultured at sub-confluence in DMEM + 10% FBS + 0.5% Pen/Strep. Low passage stocks were utilized and regularly tested for mycoplasma and luciferase activity and maintenance of growth characteristics.
POL5551 was supplied by Polyphor. POL5551 has a template-bound β-hairpin peptide structure and belongs to a series of potent CXCR4 antagonists developed by Polyphor employing the proprietary Protein Epitope Mimetics (PEM) Technology (36). The peptide backbone of POL5551 was synthesized using a standard fluorenylmethoxycarbonyl (Fmoc) solid-phase strategy on highly acid labile chlorotrityl chloride resin (100–200 mesh, Novabiochem, 01-64-0114). A disulfide bond was installed on the resin, and macrocyclization was performed in solution. The peptide was purified by preparative reverse phase HPLC. Conversion into acetate salt was performed using an ion exchange resin and the final preparation of POL5551 was recovered by lyophilization. In mice, POL5551 was well tolerated when exposed at intravenous bolus doses up to 120 mg/kg, with little effect on blood pressure, autonomic nervous, hepatic or cardiac tissues and no effect on mortality during subsequent 3 days after dosing was observed (27). The plasma half life of POL5551 in mice post subcutaneous administration is about 30 minutes (26), the exposure profile following subcutaneous administration of a single 30mg/kg dose to mice is provided in the Supplemental Figure 1B. For in vitro experiments POL5551 was dissolved in PBS to desired concentration. For in vivo studies, POL5551 (20 mg/kg) was diluted in saline and administered by subcutaneous injection.
Eribulin (trade name: HALAVEN®) was purchased from Eisai Co (Woodcliff Lake, NJ). Eribulin was dissolved in PBS in vitro to desired concentration. For in vivo studies, eribulin was diluted in saline and administered by intravenous injection once a week at 0.1 mg/kg for primary mammary fat pad therapy and 0.2 mg/kg for metastatic therapy.
Split luciferase assay
For the split-luciferase assay, CXCL12-CGLuc or unfused CGLuc MDA-MB-231 cells (2×104 cells per well in the 96-well-plate) were co-incubated overnight with NGLuc-CXCR4 or NGLuc-CXCR7 MDA-MB-231 cells in DMEM with 0.5% FBS/0.5% Pen/Strep, followed by incubation with indicated concentrations of POL5551 for 6 hours. Bioluminescence from Gaussia luciferase complementation was measured 4 hours later using BioLux® Gaussia Luciferase Assay Kit (New England Biolabs) according to the manufacture’s protocol.
MTT assay
MTT assay was performed as described previously (37).
Scratch wound assay
MDA-MB-231 cells (105 cells per well in 24-well-plate) were seeded to form a confluent monolayer. After overnight serum starvation (0.5% FBS), a wound gap was created by scratch with a pipette tip and POL5551 (0.1–5 μM) was added. Images of cells were taken with a Nikon Eclipse TE300 inverted microscope connected to a Magnafire camera model S99802 (Optronics) as previously described (38). The extent of gap closure was measured after 24 hours using ImageJ (NIH).
Survival assay
To test for survival, MDA-MB-231 cells were plated to 6-well ultra-low attachment plates at a cell density of 5×105 per well in 0.5% FBS DMEM. After 48 hour incubation with SDF-1 (12.5 ng/ml and 50 ng/ml) and in the presence or absence of POL5551 (8 μM), 1:10000 aliquot of the cells were plated to 6-well-plates and grown in 10% FBS DMEM for a week. Cells were fixed in 10% buffered formalin and stained with 0.5% crystal violet dissolved in 1% SDS. Cell density was quantified by measuring the absorbance at 570 and 630 nm by a plate reader (BioTek) (39).
Animal studies
BALB/c and NOD-scid-IL2R gammanull (NSG) mice were obtained from the Jackson Laboratory. Animals were housed under pathogen-free conditions according to the guidelines of the Division of Comparative Medicine, Washington University, St. Louis, MO. All animal experiments were approved by the Washington University Animal Studies Committee.
For xenograft experiments, 6–8 week old female NSG mice were inoculated with 5×105 MDA-MB-231 cells in Matrigel (BD Biosciences) in the #9 mammary fat pad to generate orthotopic breast tumors. As an experimental model of bone metastasis, 1×105 4T1 or MDA-MB-231 cells were injected into the left cardiac ventricle as previously described (40).
In neoadjuvant-adjuvant regimens, POL5551 was administered at the dose of 20 mg/kg subcutaneously twice a day from day 7 as a monotherapy, or from day 10 in combination with eribulin. Eribulin was administered on day 10, 17 and 24 (0.1 mg/kg, i.v.). For the “framing dosing” experiment, on day 10 POL5551 (20 mg/kg) was administered subcutaneously 4 hours before, 4 hours after, and again 18 hours after chemotherapy with erubulin (0.2 mg/kg, i.v.). A simulated exposure profile following administration of POL5551 using this dose regimen is shown in Supplemental Figure 1C. Vehicle-treated controls received saline solution (i.v. or s.c. as appropriate, see individual experiments for details).
Bioluminescence imaging
Bioluminescence imaging was performed using an IVIS 100 device (Caliper Life Sciences) as previously described (40).
Micro–computed tomography
Tibiae and femurs were removed post mortem, scanned in a 17mm holder using micro–computed tomography (μCT-40; Scanco Medical) and evaluated as described previously (38).
Radiography
Osteolytic lesion was imaged by X-Ray imaging system (Faxitron, Buffalo Grove, IL). Quantification of osteolytic lesion area was completed using Image-Pro Plus (MediaCybernetics, Rockville, MD).
Complete blood count
Whole blood was analyzed on a Hemavet Automated Coulter Counter (CBC Tech, Oxford, CT, USA).
Immunohistochemistry
Immunohistochemistry staining was completed by routine ABC method according to protocol (available online). Antigen retrieval was by the microwave heating method with 10 mM sodium citrate buffer (pH=6.0). The anti-human keratin 18 antibody (Spring Bioscience, Pleasanton, CA) was used at 1:200. The anti-F4/80 antibody and anti-CD31 antibody (Abcam, Cambridge, UK) were used at 1:500 and 1:400 respectively. Anti-rabbit (Perkins Elmer, Inc., Waltham, MA) or rat (Jackson ImmunoResearch Laboratories, Inc., West Grove, PA) secondary antibody was used at 1:800 and SA-HRP antibody (Jackson ImmunoResearch Laboratories, Inc., West Grove, PA) was used at 1:1600. Images of histology slides were acquired using a Nanozoomer digital slide scanner (Hamamatsu Photonics, Hamamatsu, Japan) and image data analysis was done using Visiomorph software (Visiopharm, Denmark).
Statistical analysis
Experiments were analyzed using 2-tailed Student t test (2 groups), one-way ANOVA (>2 groups or repeated measures), or two-way ANOVA (two variables, p value refers to the interaction) using Prism5 (GraphPad Software, Inc, San Diego, CA). Results were considered to reach significance at p≤ 0.05 and are indicated with asterisks (*p≤ 0.05, **p≤ 0.01, ***p≤ 0.001). Data are presented as mean values, error bars represent SEM. Mouse weight changes are presented as mean and range.
Results
High CXCR4 expression associated with high rate of metastases even in patients without detectable bone marrow disseminated tumor cells (DTC)
The presence of microscopic DTC in bone marrow is an independent poor prognostic factor for patients with localized breast cancer (18). As CXCR4 also correlates with metastatic potential (41, 42) we investigated the prognostic implications of CXCR4 expression in the primary tumor and the presence of bone marrow DTC, utilizing gene expression profiling of primary breast tumors from patients enrolled in our previous Phase II trial evaluating the effect of zoledronic acid (ZA) and chemotherapy on DTC in patients with locally advanced breast cancer (29, 31). In this study, 81 patients had evaluable data for both bone marrow DTC and primary tumor gene expression profiling. From this dataset, we determined the cut-off value for high and low populations based on the distribution of CXCR4 expression signal from all patients. We found that DTC were present in 5/10 (50%) CXCR4high and 13/28 (46%) CXCR4low patients, with no significant association between the presence of DTCs and CXCR4 expression (Fisher’s exact test p=0.72, odds ratio 0.6, CI 95% 0.1–2.6) (Supplemental Table 2). Both CXCR4high and CXCR4low populations had comparable frequencies of DTCs relative to the total trial dataset (Supplemental Table 3) (29), and DTC frequencies obtained in other published studies (43).
To analyze the effects of DTC and CXCR4 status on prognosis, we then considered the standard chemotherapy alone arm of the study (38 evaluable patients). The zoledronic acid (ZA) treatment arm was not used because it has been previously reported that treatment of ZA can increase disease-free survival in patients with breast cancer (44–46). 18/38 (47.3%) of these patients had detectable DTCs in the bone marrow at diagnosis. We first evaluated the impact of CXCR4 expression in the primary tumor (Figure 1A, Supplemental Figure 2) on survival, and found that high CXCR4 expression was associated with poor survival, consistent with previous reports (47), and with a higher incidence of metastases (Table 1) to bone, lung and liver (60%), than CXCR4low patients (25%). Importantly, metastases took over five times longer to develop in the CXCR4low expression patients (median of 55 months from diagnosis) compared to in the CXCR4high patients (median of 10 months). We next evaluated CXCR4 expression in the DTC negative population. We found that DTCnegCXCR4high patients had a significantly lower survival (**, p=0.005) (Figure 1B) than DTCnegCXCR4low. These data suggest that that low CXCR4 expression and the absence of bone marrow DTC define a highly favorable prognosis group in patients with locally advanced, stage II and III breast cancer. Since we found that high CXCR4 expression was associated with metastasis and poor prognosis even in patients with no detectable bone marrow DTC, we hypothesized that blocking CXCR4 activity may reduce both visceral and bone metastases.
Figure 1. High CXCR4 expression associated with high rate of metastases even in patients without detectable bone marrow disseminated tumor cells.

A) Overall survival of patients with primary breast tumors that are CXCR4High (n=10) and CXCR4Low (n=28); B) Overall survival of patients that are DTCnegative characterized by CXCR4High (n=5) or CXCR4Low (n=15) expression of the primary tumor
Table 1.
Metastasis profiling of CXCR4High and CXCR4Low patient groups
| % of patient with distant metastasis | First metastasis detected (median months; range) | |
|---|---|---|
| CXCR4High | 60%; 6/10 | **10; (4–35) |
| CXCR4Low | 25%; 7/28 | 55; (8–104) |
p<0.005; Metastases detected in bone, liver, lung and other organs.
CXCR4 selective antagonist, POL5551 decreases breast cancer cell migration and promotes anoikis
We evaluated a novel fully synthetic cyclic peptide CXCR4 antagonist, POL5551, for effects on breast cancer cell viability and migration. Breast cancer cells can express both of the CXCL12/SDF-1 receptors, CXCR4 and the alternative inhibitory receptor CXCR7 (48). To confirm POL5551’s specificity for CXCR4 over CXCR7, we utilized a complementation-imaging model (35), in which the N terminal portion of the Gaussia luciferase (GLuc) is fused to CXCR4 or CXCR7 while the C terminal portion of GLuc is fused to the ligand SDF-1. In the presence of coelenterazine, reconstitution of GLuc and subsequent light production serves as a quantitative measure of receptor-ligand binding and POL5551 bioactivity (Supplemental Figure 3A). POL5551 inhibited luciferase activity at low nanomolar concentration in human MDA-MB-231 breast cancer cells expressing GLuc-engineered CXCR4, but not GLuc - CXCR7, demonstrating specific bioactivity against CXCR4 and not CXCR7 (Figure 2A).
Figure 2. CXCR4 selective antagonist POL5551 decreases breast cancer cell migration and promotes anoikis in vitro.

A) Complementation imaging. Binding of SDF-1-CGLuc to NGLuc-CXCR4 or NGLucCXCR7 reconstitutes GLuc, producing light as a quantitative measure of ligand:receptor binding. SDF-1-CGLuc MDA-MB-231 cells were co-incubated overnight with NGLuc-CXCR4 or NGLuc-CXCR7 MDA-231 cells, followed by incubation with various concentration of POL5551 for 6 hours (***, p<0.001); B) MDA-231 cells were incubated with various concentration of POL5551 for 48 hours and cell viability was assessed by an MTT assay; C) & D) A wound gap was created by scratch in confluent monolayers of SDF-1-secreting MDA-231 cells. Gap closure was measured after 24 hours of treatment with various concentrations of POL5551 (*, p<0.05; ***, p<0.001); E) & F) Cell survival after forced suspension culture with SDF-1 (12.5ng/ml; 50ng/ml) and POL5551 (8uM). Cell colonies were stained with crystal violet. The absorbance of crystal violet dye dissolved with 1% SDS was measured by plate reader at 570–630nm (*, p<0.05; **, p<0.01; ***, p<0.001).
We investigated the bioactivity of POL5551 on breast cancer cell viability and migration, since the activation of CXCR4 receptor with ligand SDF-1 mediates many cellular functions including chemotaxis and survival (1). Under standard culture conditions, neither the chemokine SDF-1 nor the CXCR4 antagonist POL5551 had significant effects on MDA-MB-231 viability (Figure 2B). However, when cultured in anoikis-inducing suspension conditions (0.5% FBS and ultra-low attachment plates), SDF-1 promoted the survival of MDA-MB-231 cells in a concentration-dependent manner, and POL5551 significantly decreased the survival of tumor cells in suspension (Figure 2E&F). Others and our own data have shown that CXCR4 antagonist such as AMD3100 can block SDF-1 mediated migration of breast cancer cells (Supplemental Figure 3B) (49). In a scratch migration assay, POL5551 decreased chemotactic migration of SDF-1-expressing MDA-MB-231 cells in a concentration-dependent manner (Figure 2C&D). These data show that POL5551 had little direct bioactivity on breast cancer cell viability, but inhibited SDF-1-dependent migration and resistance to anoikis.
CXCR4 antagonist POL5551 decreases metastasis but has little effect on primary tumor growth
Because POL5551 decreased CXCR4-mediated migration and tumor cell survival specifically in suspension, we evaluated the effect of POL5551 on local recurrence and distant metastasis. Orthotopic mammary fat pad (MFP) MDA-MB-231 tumors were established in immunocompromised NSG mice. POL5551 monotherapy was initiated when tumors were palpable (day 7) and continued until study termination. Primary MFP tumors were surgically resected by mastectomy on day 21 and mice were followed for the development of local recurrence and distant metastasis by bioluminescent imaging (BLI, Figure 3A). We confirmed that the POL5551 dosing used in this study were sufficient to mobilize hematopoietic cells into the blood [22] and increase WBC counts (Supplemental Figure 4). While POL5551 did not reduce primary tumor growth (Figure 3 B&C and Supplemental Figure 4), distant metastases to chest and leg bones as assessed by BLI, were significantly decreased in POL5551-treated mice (Figure 3 D, E &H). Metastasis to the liver was examined by both ex vivo BLI and histology, however no significant differences were found between the treatment groups (Figure 3I, Supplemental Figure 4 B&C). Micrometastatic tumors in the lung, as quantified histologically by measurement of human cytokeratin-18 (CK-18) positive tumor cells, were significantly decreased in POL5551-treated mice compared to vehicle controls (p=0.001 Figure 3 F&G).
Figure 3. CXCR4 antagonist POL5551 decreases metastasis but has little effect on primary MFP tumor growth in a xenograft NSG model.

A) MDA-MB-231 mammary fat pad orthotopic model schema: MDA-MB-231 tumor cells were injected to the mammary fat pad of mice. Treatment with POL5551 (20mg/kg, s.c., b.i.d, n=3) or saline (n=5) commenced on day 8 when the tumor became palpable and continued until the end of study; B) & C) Primary MFP tumor growth as measured by bioluminescence imaging, representative image showing the primary tumors on Day 21 ; D) & E) Distant metastasis to the chest as measured by bioluminescence imaging, representative image showing the distant metastases on Day 41 (*, p<0.05); F) Human cytokeratin 18 (CK18) staining of MDA-MB-231 tumor cells in lung tissue; G) Quantification of CK18 positive cells in lung tissue; ***, p<0.001; H) Ex vivo BLI of bone metastases (*, p<0.05); I) Ex vivo BLI of liver metastases (p>0.05).
To confirm POL5551 inhibition of metastasis in the context of a fully competent immune system and in a metastasis model, 4T1 murine breast cancer cells were inoculated into the left cardiac ventricle of BALB/c mice to establish disseminated metastatic disease. Mice were treated either with POL5551 or saline from day 3 to the end of the study (Figure 4A). Histomorphometric analyses showed that POL5551 treatment significantly decreased tumor metastases into the kidney (p=0.03) (Figure 4 D&E), and micro-CT showed a significant reduction in tumor-associated bone loss (p=0.03) (Figure 4 F&G). These findings were associated with a non-significant trend towards reduction of bone and whole-body tumor burden when measured by BLI (Figure 4 B&C). Together these data show that POL5551 as a single agent had little direct effect on primary tumor burden, but decreased metastases and metastatic tumor burden in 2 breast cancer models.
Figure 4. POL5551 monotherapy decreased distant metastasis in a disseminated tumor model.

A) intracardiac tumor injection model, 4T1-Luc murine tumor cells were injected into the left ventricle of the BALB/c mice. Treatment with POL5551 (20mg/kg, s.c., b.i.d, n=7) or vehicle (saline, n=6) was from day 3 to day 11. Mice were sacrificed on day 11; B) & C) Bioluminescent imaging of 4T1-Luc tumor burden in leg bones (ns, p=0.23) and whole body (ns, p=0.08); D) H&E staining of kidney tissue, 4T1 tumor foci/nodules are outlined in white to increase visibility; E) Quantification of 4T1 tumor foci in kidney per section (n=6/group; **, p<0.01); F) & G) μCT analysis for calculation of trabecular bone to tissue volume ratio (BV/TV) (n=6/group; *, p<0.05).
Combination of eribulin with POL5551 in the neoadjuvant and metastatic settings improves survival compared to eribulin alone
In addition to investigating the effect of POL5551 administered as a monotherapy, we also evaluated the effect of POL5551 in combination with cytotoxic chemotherapy. Eribulin is a microtubule-inhibiting chemotherapeutic agent, with strong activity in patients with metastatic breast cancer that have failed both anthracycline and taxane chemotherapies (50–52). Eribulin decreased MDA-MB-231 viability in vivo (Supplemental Figure 5A–C) and prolonged mice survival in a dose-dependent manner (Supplemental Figure 5D). However, the higher and most effective doses were associated with greater chemotherapy-induced weight loss (Supplemental Figure 5E). Aiming at identifying a combination regimen that would reduce toxicity and reveal synergistic effects with CXCR4 inhibition, we selected a submaximal effective dose of eribulin (0.1 mg/kg) that yielded 50% primary breast tumor reduction when used as single-agent (Supplemental Figure 5F). After orthotopic MFP tumors were established, neoadjuvant treatment was initiated on day 10 and mastectomy was performed when all tumors reached 1 cm3, so that metastasis free survival after surgery could be fairly evaluated for all treatment groups. Due to the anti-tumor effect of eribulin on primary growth, chemotherapy-treated mice required additional time (roughly 5 extra days) to reach primary tumor mass equivalent to the vehicle group. All doses of eribulin were administered before mastectomy, whereas POL5551 administration continued after mastectomy. As expected, eribulin decreased primary tumor growth rate, while POL5551 had no direct or additive effects on primary MDA-MB-231 MFP tumors (Figure 5A–C). In contrast, combination POL5551 plus eribulin treatment significantly decreased distant metastases to the chest and bone as measured by BLI (Figure 5D&E). Finally, POL5551 in combination with eribulin prolonged survival (58 days) compared to eribulin alone (51 days) or vehicle controls (45 days) (Figure 5F).
Figure 5. Combination of eribulin with POL5551 in the neoadjuvant setting improves survival compared to eribulin alone.
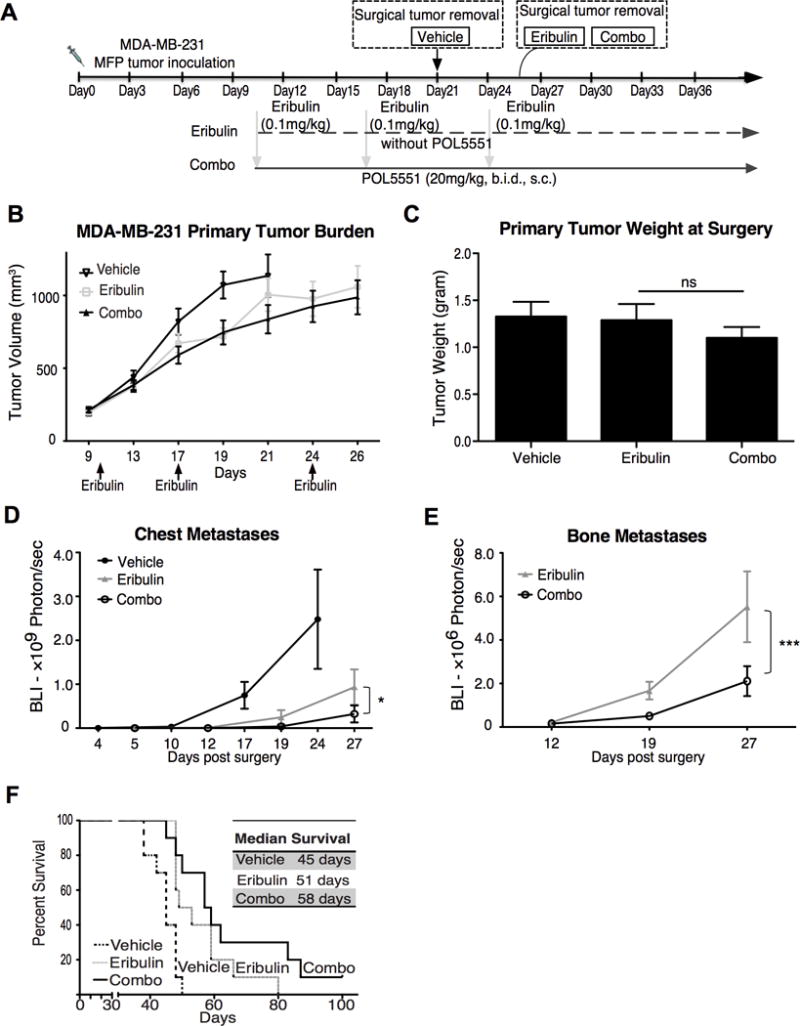
A) Experimental design: MDA-MB-231 cells were mixed with matrigel and implanted s.c. into the right flank of the NSG mice. Ten days after tumor implantation, mice (n=10/group) were treated as follow: a) Vehicle saline (i.v.) on day 10, 17 & 24) plus (s.c., b.i.d) from day 10 to the day of surgery; b) Eribulin alone (0.1mg/kg, i.v) on days 10, 17 and 24; c) Eribulin treatment (days 10, 17 & 24) plus POL5551 (20mg/kg, s.c., b.i.d) administration daily from day 10 until termination; B) Primary tumor size measured by caliper; C) Primary tumor weight at surgery (ns, p>0.05); D) Bioluminescence imaging of distant metastasis to the chest (*, p<0.05); E) Bioluminescence imaging of distant metastasis to the bone in eribulin alone and combo group (***, p<0.001); F) Kaplan Meier survival curve.
Breast cancer cell interactions with the stromal environment can contribute to resistance to cytotoxic chemotherapy (11). We hypothesized that antagonizing CXCR4 with POL5551 for short periods of time around each dose of chemotherapy - a “framing dosing” strategy - could disrupt CXCR4/SDF-1 mediated tumor-stromal cell interactions and potentially enhance chemotherapy efficacy. To test this hypothesis, metastases were established by intracardiac inoculation of MDA-MB-231 cells, and POL5551 was administered 4 hours before eribulin and 4- and 18-hours after eribulin chemotherapy (Figure 6A). Following intracardiac injection, MDA-MB-231 cells preferentially metastasize to the liver and leg bones (Figure 6F). The rate of distant metastasis of mice with successful left ventricle injection was 100%. In the hind leg bones, the metastasizing tumor cells produce severe osteolytic lesions, usually detectable by X-Ray two weeks post inoculation. Compared to vehicle control and eribulin alone, POL5551 in combination with eribulin was superior in decreasing bone tumor burden as measured by BLI (Figure 6C). In the liver, whilst erubulin was very effective in reducing tumor burden (BLI) by 75%, this result was further reduced (86%) in the POL5551+eribulin group (Figure 6B, p>0.05 between Eribulin alone and Combo group). In the bone, POL5551 in combination with eribulin produced a significant reduction in BLI, and fewer tumor-associated osteolytic areas were detectable on X-Ray, compared to the vehicle and eribulin alone group (Figure 6 D&E). Progressive metastatic tumor burden results in weight loss in this model, and the combination of POL5551 with eribulin was superior to eribulin alone in preventing tumor associated weight loss (Figure 6G, P<0.05). Together these data suggest that short, sequenced dosing of POL5551 with eribulin chemotherapy can prevent tumor associated weight loss and decrease tumor burden.
Figure 6. Reduction of bone tumor burden: POL5551 in combination with eribulin is superior to eribulin alone.

A) Schema of bone metastasis model utilizing “framing dose” strategy: MDA-MB-231 bone metastases were established by intracardiac inoculation in NSG mice, and on day 10 mice were treated as following: a) Vehicle (n=4): saline (s.c. and i.v.); b) Eribulin alone (n=5): eribulin (0.2mg/kg, i.v.) alone and saline (s.c); c) Combination treatment (n=5): POL5551 (20mg/kg, s.c.) was administered 4 hours before eribulin and 4- and 18- hours after eribulin (0.2mg/kg, i.v.) chemotherapy; B) & C) Bioluminescent imaging of tumor burden in liver and bone (*, p<0.05; ***, p<0.001); D) Representative X-Ray images of tumor associated osteolytic lesion in leg bone; E) Quantification of osteolytic lesion area; F) Representative image showing extent of metastatic burden in liver and leg bone on day17 (*, p<0.05; **, p<0.01); G) Mouse weight lost between day 9 and day 17. Statistical analyses are as compared to vehicle treatment (ns, p>0.05; **, p<0.01).
Discussion
Our work shows that high CXCR4 expression in primary breast cancer tissue defined a subgroup of patients that, despite the lack of bone marrow disseminated tumor cells and other favorable characteristics, had high and early incidence of metastasis and poor survival. It has been proposed that CXCR4 antagonism could directly impact tumor cell survival at two stages of metastasis. Early in progression, CXCR4 inhibitors may inhibit tumor cell homing from the primary tumor to metastatic sites expressing high SDF-1. After tumor cells have disseminated to secondary sites and are protected from chemotherapy, CXCR4 antagonists may mobilize tumor cells into the peripheral blood as occurs with hematopoietic stem cells. Once in circulation, tumor cells are no longer protected by interactions with stromal cells in the HSC niche and could become more susceptible to therapeutic agents including chemotherapy, resulting in reduced metastases and tumor burden. We hypothesized that inhibition of CXCR4 could decrease metastasis. We found that the CXCR4 antagonist, POL5551, blocked breast cancer cell migration and resistance to anoikis in vitro, and reduced breast cancer metastasis despite having little effect on primary tumor growth in vivo. When administered in combination with eribulin chemotherapy, POL5551 decreased metastases and improved survival compared with chemotherapy alone. We hypothesized that POL5551 could “mobilize” tumor cells from their stromal and vascular niches and heighten sensitivity to chemotherapy. We found that only 3 doses of POL5551 given just before and just after eribulin further decreased metastatic tumor growth compared to chemotherapy alone. Taken together, our findings suggest that POL5551 is capable of disrupting interactions between cancer cells and their environment that contribute to their metastatic potential and resistance to chemotherapy.
It has been previously reported that POL5551 is a potent mobilization agent, which can mobilize hematopoietic stem and progenitor cells from bone marrow niche with greater efficiency than AMD3100 (plerixafor) (26). In confirmation of this, the POL5551 dose we used in vivo rapidly mobilized leukocytes into the circulation (Supplemental Figure 6). During metastatic dissemination, CXCR4 expressing cancer cells are thought to utilize the HSC-specific pathways to preferentially home to SDF-1 rich sites, such as bone, lymph nodes and lungs. In a mouse metastasis model, Shiozawa et al demonstrated that prostate cancer cells can directly compete with HSC to occupy the HSC niche in the bone marrow through CXCR4/SDF-1 pathway and that administration of CXCR4 antagonist AMD3100 mobilized disseminated tumor cells into the peripheral blood (8). Stromal environments in the bone or lung have been shown to be protective and contribute to chemoresistance to cytotoxic therapies (11); mobilization of tumor cells from the protective microenvironment may therefore increase their susceptibility to cytotoxic chemotherapy.
Two main chemokine receptors capable of SDF-1 binding have been found to play some role in breast cancer: CXCR4 and CXCR7. While CXCR7 may play a role in transendothelial migration (53), Hernandez et al demonstrated that CXCR7 overexpression enhanced tumor growth and vascularization, but reduced invasiveness and metastases (54). Specifically designed to selectively target CXCR4, POL5551 showed no effect on SDF-1 binding to overexpressed CXCR7 by Gaussia luciferase complementation assay in breast cancer cells. Moreover, in agreement with previously published datasets (55), we did not detect CXCR7 expression in MDA-MB-231 cells. We therefore consider it unlikely that CXCR7 might play a major role in the current set of experiments.
Targeting a physiological characteristic of the cell of origin, together with a pro-tumor gain-of-function, a number of clinical trials have used CXCR4 antagonists in hematological malignancies, such as MM, AML and lymphoma (14). In solid tumors, CXCR4 overexpression has been associated with poorer prognosis, and functional studies have shown that CXCR4 plays a critical role in the metastatic dissemination of solid tumor cells (56) (57). In accordance with this hypothesis, we found that, even in patients with no bone marrow micrometastases (DTC negative), high CXCR4 expression in the primary tumor was associated with earlier metastases and poorer prognosis.
In the host, the CXCR4 gene plays an important role in a number of cell types and physiological processes, such as HSCs and leukocytes trafficking. In our previous work, we have shown that targeted gene disruption of host hematopoietic CXCR4 resulted in enhanced osteoclastogenesis and consequent tumor growth in bone, although no differences were found in lung and subcutaneous tumor burden (58). By pharmacologically targeting CXCR4 receptor with an antagonist, especially with an intermittent regimen, CXCR4 expression by normal tissues would not be constantly disrupted, thus potentially having less sustained effects on leukocytosis. Indeed, mice did not show overt hematopoietic toxicity as measured by serial blood counts following a framing dose of POL5551 in combination with eribulin (Supplemental Figure 7). It should be noted that not all HSC and/or DTC are mobilized after a single dose of a CXCR4 antagonist, so repeated dosing may have higher efficacy. In the current study, we found that POL5551 did not induce bone loss and in fact resulted in decreased tumor associated bone loss. This is likely due to decreased tumor burden in the bones, which is consistent with the reduction of metastatic tumor burden in extra-skeletal sites (lungs, kidneys), and with the observed effects on the migration and survival of isolated tumor cells. We also anticipate that the pharmacologic dosing of POL5551 would not affect osteoclasts to the same extent as complete genetic disruption.
Among breast cancers, the triple-negative breast cancer (TNBC) subtype has the most aggressive phenotype with a poorer prognosis (59–61). Unlike ER or HER2 positive breast cancers, which benefit from targeted therapies, the treatment of TNBC patients largely relies on surgery and systemic chemotherapy. The microtubule inhibitor eribulin has shown significant survival benefit in patients with metastatic breast cancer refractory to anthracyclines and taxanes (62) (51). As with most antiproliferative drugs, in addition to the killing effect of chemotherapy on rapidly dividing tumor cells, chemotoxicity often occurs. In a dose ranging experiment, in which we treated mice bearing MDA-MB-231 tumors with eribulin, a high dose of eribulin (>0.5 mg/kg) significantly improved survival, but to the expense of greater treatment associated weight loss due to chemotoxicity. In addition, optimal eribulin dosing reduced tumor burden to a point which was below the level in which our in vivo bioluminescence assays were able to consistently and confidently measure tumor burden, precluding our ability to detect any synergistic effects of POL5551. For these reasons, sub-optimal and less toxic dosing of eribulin (0.1–0.2mg/kg) was utilized for this pre-clinical study. In agreement with previous research that has shown CXCR4 inhibition to sensitize tumor cells to chemotherapy (13, 63), we found that combining POL5551 with low-dose eribulin (0.1–0.2 mg/kg) decreased metastatic tumor burden compared to mice administered eribulin alone. Together with other studies in published literature, our preclinical data suggest that combining CXCR4 antagonism and cytotoxic chemotherapy could decrease metastasis by mobilizing tumor cells or otherwise disrupting their interactions with chemoprotective stromal environments. In the present study POL5551 administered just before and after eribulin was sufficient to decrease tumor burden.
Few clinical trials have so far targeted CXCR4 in solid tumors, despite encouraging preclinical studies and associations between expression and prognosis. A Phase I study using a peptide CXCR4 antagonist, LY2510924, in patients with advanced cancer was recently completed in the US (64). However, no dedicated breast cancer trial has so far been designed to evaluate CXCR4 antagonism. This study has provided important pre-clinical rationale and proof-of-principle data to examine the combination of CXCR4 antagonism and eribulin chemotherapy in metastatic breast cancer patients. To translate these findings from bench to bedside, a multicenter Phase I clinical trial (“Dose escalation of POL6326 in combination with eribulin in patients with metastatic breast cancer”, ClinicalTrials.gov Identifier: NCT01837095, sponsored by Polyphor), has been initiated. This study will evaluate POL6326, a close analogue of POL5551, which differs at only a single amino acid, has nearly identical bioactivity profiles, but has slightly more favorable pharmacokinetic properties than POL5551. Here, POL6326 will be evaluated in combination with eribulin in a framing dose sequence similar to what we employed in the preclinical studies presented in Figure 6. This is the first clinical trial in breast cancer patients with a CXCR4 antagonist in combination with chemotherapy. In addition, POL6326 is currently in clinical studies for hematopoietic stem cell mobilization.
While the overall magnitude of survival extension in this preclinical study was modest (as compared to vehicle, median survival was increased 12% with eribulin alone and 22% with erubulin+POL555), it is important to note that a suboptimal dose of eribulin was utilized for this study. This was necessary to permit a level of tumor growth sufficient for detection by in vivo bioluminescent assays. Thus, when optimal doses of eribulin are utilized, as in the ongoing Phase I clinical trial above, additional benefit may be seen. Our preclinical data suggest that CXCR4/SDF-1 interactions play a critical role during breast cancer metastasis, and that CXCR4 antagonism administered in sequence with chemotherapy can disrupt tumor cell spread. Future studies to evaluate the impact of CXCR4 inhibition on tumor cell dormancy, tumor microenvironment and immune modulation will be important to better define which patients to treat and how to combine CXCR4 antagonist with ongoing cancer therapies.
Supplementary Material
Acknowledgments
Grant Support:
The authors wish to thank Michael Tomasson, John DiPersio, David DeNardo, Melissa Meyer, Yu Zhu, and Michael Ross for their valuable expert suggestions and criticism. We thank Yalin Xu, Lynne Collins, and Julie Prior for their technical assistance. We thank Dr. Mark Watson for the supervision of the gene array data generation. This research was supported by grants from NIH CA097250 (K.N. Weilbaecher, M.A. Hurchla, J. Xiang, X. Su, F. Fontana), NIH CA100730 (K.N. Weilbaecher, J. Xiang, A.K. Esser), NIH CA154737 (K.N. Weilbaecher, A.K. Esser), NIH research fellowship 1F31CA174096–01A1 (S.R Amend), NIH 1K12CA167540 (F.O. Ademuyiwa), NIH R01CA172623 (R.L. Aft, C. Mudalagiriyappa), NIH CA170198, CA142750, CA182333 (G.D. Luker, K.E Luker). Targeted funding to purchase NSG mice for a subset of experiments and POL5551 compound was provided by Polyphor Ltd. We thank the Musculoskeletal Research Center for microCT (NIH P30 AR057235), the P50 small animal imaging core for bioluminescence studies (CA94056), DDRCC Morphology core (Grant # P30 DK52574), The Barnes-Jewish Foundation, The St. Louis Men’s Group Against Cancer, Hope Center Alafi Neuroimaging Lab (NIH Shared Instrumentation Grant # S10 RR027552).
Footnotes
Disclosure of Potential Conflicts of Interest:
POL5551 compound was provided by Polyphor Ltd. Targeted funding to purchase NSG mice for a subset of experiments was provided by Polyphor Ltd. GD, BR, GT, EC, MB, JZ and KD are all employed by Polyphor and own shares in the company.
References
- 1.Teicher BA, Fricker SP. CXCL12 (SDF-1)/CXCR4 pathway in cancer. Clinical cancer research: an official journal of the American Association for Cancer Research. 2010;16:2927–31. doi: 10.1158/1078-0432.CCR-09-2329. [DOI] [PubMed] [Google Scholar]
- 2.Dar A, Kollet O, Lapidot T. Mutual, reciprocal SDF-1/CXCR4 interactions between hematopoietic and bone marrow stromal cells regulate human stem cell migration and development in NOD/SCID chimeric mice. Experimental hematology. 2006;34:967–75. doi: 10.1016/j.exphem.2006.04.002. [DOI] [PubMed] [Google Scholar]
- 3.Ma Q, Jones D, Borghesani PR, Segal RA, Nagasawa T, Kishimoto T, et al. Impaired B-lymphopoiesis, myelopoiesis, and derailed cerebellar neuron migration in CXCR4- and SDF-1-deficient mice. Proceedings of the National Academy of Sciences of the United States of America. 1998;95:9448–53. doi: 10.1073/pnas.95.16.9448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Nagasawa T, Hirota S, Tachibana K, Takakura N, Nishikawa S, Kitamura Y, et al. Defects of B-cell lymphopoiesis and bone-marrow myelopoiesis in mice lacking the CXC chemokine PBSF/SDF-1. Nature. 1996;382:635–8. doi: 10.1038/382635a0. [DOI] [PubMed] [Google Scholar]
- 5.Ara T, Tokoyoda K, Sugiyama T, Egawa T, Kawabata K, Nagasawa T. Long-term hematopoietic stem cells require stromal cell-derived factor-1 for colonizing bone marrow during ontogeny. Immunity. 2003;19:257–67. doi: 10.1016/s1074-7613(03)00201-2. [DOI] [PubMed] [Google Scholar]
- 6.Liles WC, Broxmeyer HE, Rodger E, Wood B, Hubel K, Cooper S, et al. Mobilization of hematopoietic progenitor cells in healthy volunteers by AMD3100, a CXCR4 antagonist. Blood. 2003;102:2728–30. doi: 10.1182/blood-2003-02-0663. [DOI] [PubMed] [Google Scholar]
- 7.Li L, Neaves WB. Normal stem cells and cancer stem cells: the niche matters. Cancer research. 2006;66:4553–7. doi: 10.1158/0008-5472.CAN-05-3986. [DOI] [PubMed] [Google Scholar]
- 8.Shiozawa Y, Pedersen EA, Havens AM, Jung Y, Mishra A, Joseph J, et al. Human prostate cancer metastases target the hematopoietic stem cell niche to establish footholds in mouse bone marrow. The Journal of clinical investigation. 2011;121:1298–312. doi: 10.1172/JCI43414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Chatterjee S, Behnam Azad B, Nimmagadda S. The intricate role of CXCR4 in cancer. Advances in cancer research. 2014;124:31–82. doi: 10.1016/B978-0-12-411638-2.00002-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Meads MB, Hazlehurst LA, Dalton WS. The bone marrow microenvironment as a tumor sanctuary and contributor to drug resistance. Clinical cancer research: an official journal of the American Association for Cancer Research. 2008;14:2519–26. doi: 10.1158/1078-0432.CCR-07-2223. [DOI] [PubMed] [Google Scholar]
- 11.Castells M, Thibault B, Delord JP, Couderc B. Implication of tumor microenvironment in chemoresistance: tumor-associated stromal cells protect tumor cells from cell death. International journal of molecular sciences. 2012;13:9545–71. doi: 10.3390/ijms13089545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Zhang XH, Jin X, Malladi S, Zou Y, Wen YH, Brogi E, et al. Selection of bone metastasis seeds by mesenchymal signals in the primary tumor stroma. Cell. 2013;154:1060–73. doi: 10.1016/j.cell.2013.07.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Azab AK, Runnels JM, Pitsillides C, Moreau AS, Azab F, Leleu X, et al. CXCR4 inhibitor AMD3100 disrupts the interaction of multiple myeloma cells with the bone marrow microenvironment and enhances their sensitivity to therapy. Blood. 2009;113:4341–51. doi: 10.1182/blood-2008-10-186668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Domanska UM, Kruizinga RC, Nagengast WB, Timmer-Bosscha H, Huls G, de Vries EG, et al. A review on CXCR4/CXCL12 axis in oncology: no place to hide. European journal of cancer. 2013;49:219–30. doi: 10.1016/j.ejca.2012.05.005. [DOI] [PubMed] [Google Scholar]
- 15.Karshovska E, Zagorac D, Zernecke A, Weber C, Schober A. A small molecule CXCR4 antagonist inhibits neointima formation and smooth muscle progenitor cell mobilization after arterial injury. Journal of thrombosis and haemostasis: JTH. 2008;6:1812–5. doi: 10.1111/j.1538-7836.2008.03086.x. [DOI] [PubMed] [Google Scholar]
- 16.Hu C, Yong X, Li C, Lu M, Liu D, Chen L, et al. CXCL12/CXCR4 axis promotes mesenchymal stem cell mobilization to burn wounds and contributes to wound repair. The Journal of surgical research. 2013;183:427–34. doi: 10.1016/j.jss.2013.01.019. [DOI] [PubMed] [Google Scholar]
- 17.Chu QD, Panu L, Holm NT, Li BD, Johnson LW, Zhang S. High chemokine receptor CXCR4 level in triple negative breast cancer specimens predicts poor clinical outcome. The Journal of surgical research. 2010;159:689–95. doi: 10.1016/j.jss.2008.09.020. [DOI] [PubMed] [Google Scholar]
- 18.Braun S, Vogl FD, Naume B, Janni W, Osborne MP, Coombes RC, et al. A pooled analysis of bone marrow micrometastasis in breast cancer. N Engl J Med. 2005;353:793–802. doi: 10.1056/NEJMoa050434. [DOI] [PubMed] [Google Scholar]
- 19.Chen HW, Du CW, Wei XL, Khoo US, Zhang GJ. Cytoplasmic CXCR4 high-expression exhibits distinct poor clinicopathological characteristics and predicts poor prognosis in triple-negative breast cancer. Current molecular medicine. 2013;13:410–6. [PubMed] [Google Scholar]
- 20.Zhang M, Liu HX, Teng XD, Wang HB, Cui J, Jia SS, et al. The differences in CXCR4 protein expression are significant for the five molecular subtypes of breast cancer. Ultrastructural pathology. 2012;36:381–6. doi: 10.3109/01913123.2012.728687. [DOI] [PubMed] [Google Scholar]
- 21.Peled A, Wald O, Burger J. Development of novel CXCR4-based therapeutics. Expert opinion on investigational drugs. 2012;21:341–53. doi: 10.1517/13543784.2012.656197. [DOI] [PubMed] [Google Scholar]
- 22.Domanska UM, Boer JC, Timmer-Bosscha H, van Vugt MA, Hoving HD, Kliphuis NM, et al. CXCR4 inhibition enhances radiosensitivity, while inducing cancer cell mobilization in a prostate cancer mouse model. Clinical & experimental metastasis. 2014;31:829–39. doi: 10.1007/s10585-014-9673-2. [DOI] [PubMed] [Google Scholar]
- 23.Kim SY, Lee CH, Midura BV, Yeung C, Mendoza A, Hong SH, et al. Inhibition of the CXCR4/CXCL12 chemokine pathway reduces the development of murine pulmonary metastases. Clinical & experimental metastasis. 2008;25:201–11. doi: 10.1007/s10585-007-9133-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.D’Alterio C, Barbieri A, Portella L, Palma G, Polimeno M, Riccio A, et al. Inhibition of stromal CXCR4 impairs development of lung metastases. Cancer immunology, immunotherapy: CII. 2012;61:1713–20. doi: 10.1007/s00262-012-1223-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Robinson JA, Demarco S, Gombert F, Moehle K, Obrecht D. The design, structures and therapeutic potential of protein epitope mimetics. Drug discovery today. 2008;13:944–51. doi: 10.1016/j.drudis.2008.07.008. [DOI] [PubMed] [Google Scholar]
- 26.Karpova D, Dauber K, Spohn G, Chudziak D, Wiercinska E, Schulz M, et al. The novel CXCR4 antagonist POL5551 mobilizes hematopoietic stem and progenitor cells with greater efficiency than Plerixafor. Leukemia. 2013;27:2322–31. doi: 10.1038/leu.2013.266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hamesch K, Subramanian P, Li X, Dembowsky K, Chevalier E, Weber C, et al. The CXCR4 antagonist POL5551 is equally effective as sirolimus in reducing neointima formation without impairing re-endothelialisation. Thrombosis and haemostasis. 2012;107:356–68. doi: 10.1160/TH11-07-0453. [DOI] [PubMed] [Google Scholar]
- 28.Barone A, Sengupta R, Warrington NM, Smith E, Wen PY, Brekken RA, et al. Combined VEGF and CXCR4 antagonism targets the GBM stem cell population and synergistically improves survival in an intracranial mouse model of glioblastoma. Oncotarget. 2014;5:9811–22. doi: 10.18632/oncotarget.2443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Aft R, Naughton M, Trinkaus K, Watson M, Ylagan L, Chavez-MacGregor M, et al. Effect of zoledronic acid on disseminated tumour cells in women with locally advanced breast cancer: an open label, randomised, phase 2 trial. The Lancet Oncology. 2010;11:421–8. doi: 10.1016/S1470-2045(10)70054-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Lin Y, Lin S, Watson M, Trinkaus KM, Kuo S, Naughton MJ, et al. A gene expression signature that predicts the therapeutic response of the basal-like breast cancer to neoadjuvant chemotherapy. Breast cancer research and treatment. 2010;123:691–9. doi: 10.1007/s10549-009-0664-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Watson MA, Ylagan LR, Trinkaus KM, Gillanders WE, Naughton MJ, Weilbaecher KN, et al. Isolation and molecular profiling of bone marrow micrometastases identifies TWIST1 as a marker of early tumor relapse in breast cancer patients. Clinical cancer research: an official journal of the American Association for Cancer Research. 2007;13:5001–9. doi: 10.1158/1078-0432.CCR-07-0024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Budczies J, Klauschen F, Sinn BV, Gyorffy B, Schmitt WD, Darb-Esfahani S, et al. Cutoff Finder: a comprehensive and straightforward Web application enabling rapid biomarker cutoff optimization. PloS one. 2012;7:e51862. doi: 10.1371/journal.pone.0051862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Smith MC, Luker KE, Garbow JR, Prior JL, Jackson E, Piwnica-Worms D, et al. CXCR4 regulates growth of both primary and metastatic breast cancer. Cancer research. 2004;64:8604–12. doi: 10.1158/0008-5472.CAN-04-1844. [DOI] [PubMed] [Google Scholar]
- 34.Guise TA, Yin JJ, Taylor SD, Kumagai Y, Dallas M, Boyce BF, et al. Evidence for a causal role of parathyroid hormone-related protein in the pathogenesis of human breast cancer-mediated osteolysis. The Journal of clinical investigation. 1996;98:1544–9. doi: 10.1172/JCI118947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Luker KE, Mihalko LA, Schmidt BT, Lewin SA, Ray P, Shcherbo D, et al. In vivo imaging of ligand receptor binding with Gaussia luciferase complementation. Nature medicine. 2012;18:172–7. doi: 10.1038/nm.2590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.DeMarco SJ, Henze H, Lederer A, Moehle K, Mukherjee R, Romagnoli B, et al. Discovery of novel, highly potent and selective beta-hairpin mimetic CXCR4 inhibitors with excellent anti-HIV activity and pharmacokinetic profiles. Bioorganic & medicinal chemistry. 2006;14:8396–404. doi: 10.1016/j.bmc.2006.09.003. [DOI] [PubMed] [Google Scholar]
- 37.Heller E, Hurchla MA, Xiang J, Su X, Chen S, Schneider J, et al. Hedgehog signaling inhibition blocks growth of resistant tumors through effects on tumor microenvironment. Cancer research. 2012;72:897–907. doi: 10.1158/0008-5472.CAN-11-2681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Su X, Floyd DH, Hughes A, Xiang J, Schneider JG, Uluckan O, et al. The ADP receptor P2RY12 regulates osteoclast function and pathologic bone remodeling. The Journal of clinical investigation. 2012;122:3579–92. doi: 10.1172/JCI38576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Kochetkova M, Kumar S, McColl SR. Chemokine receptors CXCR4 and CCR7 promote metastasis by preventing anoikis in cancer cells. Cell death and differentiation. 2009;16:664–73. doi: 10.1038/cdd.2008.190. [DOI] [PubMed] [Google Scholar]
- 40.Uluckan O, Becker SN, Deng H, Zou W, Prior JL, Piwnica-Worms D, et al. CD47 regulates bone mass and tumor metastasis to bone. Cancer research. 2009;69:3196–204. doi: 10.1158/0008-5472.CAN-08-3358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Andre F, Xia W, Conforti R, Wei Y, Boulet T, Tomasic G, et al. CXCR4 expression in early breast cancer and risk of distant recurrence. The oncologist. 2009;14:1182–8. doi: 10.1634/theoncologist.2009-0161. [DOI] [PubMed] [Google Scholar]
- 42.Hung CS, Su HY, Liang HH, Lai CW, Chang YC, Ho YS, et al. High-level expression of CXCR4 in breast cancer is associated with early distant and bone metastases. Tumour biology: the journal of the International Society for Oncodevelopmental Biology and Medicine. 2014;35:1581–8. doi: 10.1007/s13277-013-1218-9. [DOI] [PubMed] [Google Scholar]
- 43.Fehm T, Braun S, Muller V, Janni W, Gebauer G, Marth C, et al. A concept for the standardized detection of disseminated tumor cells in bone marrow from patients with primary breast cancer and its clinical implementation. Cancer. 2006;107:885–92. doi: 10.1002/cncr.22076. [DOI] [PubMed] [Google Scholar]
- 44.Gnant M, Mlineritsch B, Stoeger H, Luschin-Ebengreuth G, Heck D, Menzel C, et al. Adjuvant endocrine therapy plus zoledronic acid in premenopausal women with early-stage breast cancer: 62-month follow-up from the ABCSG-12 randomised trial. The Lancet Oncology. 2011;12:631–41. doi: 10.1016/S1470-2045(11)70122-X. [DOI] [PubMed] [Google Scholar]
- 45.Ben-Aharon I, Vidal L, Rizel S, Yerushalmi R, Shpilberg O, Sulkes A, et al. Bisphosphonates in the adjuvant setting of breast cancer therapy–effect on survival: a systematic review and meta-analysis. PloS one. 2013;8:e70044. doi: 10.1371/journal.pone.0070044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Gnant M, Mlineritsch B, Schippinger W, Luschin-Ebengreuth G, Postlberger S, Menzel C, et al. Endocrine therapy plus zoledronic acid in premenopausal breast cancer. N Engl J Med. 2009;360:679–91. doi: 10.1056/NEJMoa0806285. [DOI] [PubMed] [Google Scholar]
- 47.Rhodes LV, Short SP, Neel NF, Salvo VA, Zhu Y, Elliott S, et al. Cytokine receptor CXCR4 mediates estrogen-independent tumorigenesis, metastasis, and resistance to endocrine therapy in human breast cancer. Cancer research. 2011;71:603–13. doi: 10.1158/0008-5472.CAN-10-3185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Burns JM, Summers BC, Wang Y, Melikian A, Berahovich R, Miao Z, et al. A novel chemokine receptor for SDF-1 and I-TAC involved in cell survival, cell adhesion, and tumor development. The Journal of experimental medicine. 2006;203:2201–13. doi: 10.1084/jem.20052144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Kirui JK, Xie Y, Wolff DW, Jiang H, Abel PW, Tu Y. Gbetagamma signaling promotes breast cancer cell migration and invasion. The Journal of pharmacology and experimental therapeutics. 2010;333:393–403. doi: 10.1124/jpet.109.164814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Twelves C, Cortes J, Vahdat L, Olivo M, He Y, Kaufman PA, et al. Efficacy of eribulin in women with metastatic breast cancer: a pooled analysis of two phase 3 studies. Breast cancer research and treatment. 2014;148:553–61. doi: 10.1007/s10549-014-3144-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Cortes J, O’Shaughnessy J, Loesch D, Blum JL, Vahdat LT, Petrakova K, et al. Eribulin monotherapy versus treatment of physician’s choice in patients with metastatic breast cancer (EMBRACE): a phase 3 open-label randomised study. Lancet. 2011;377:914–23. doi: 10.1016/S0140-6736(11)60070-6. [DOI] [PubMed] [Google Scholar]
- 52.Cortes J, Vahdat L, Blum JL, Twelves C, Campone M, Roche H, et al. Phase II study of the halichondrin B analog eribulin mesylate in patients with locally advanced or metastatic breast cancer previously treated with an anthracycline, a taxane, and capecitabine. Journal of clinical oncology: official journal of the American Society of Clinical Oncology. 2010;28:3922–8. doi: 10.1200/JCO.2009.25.8467. [DOI] [PubMed] [Google Scholar]
- 53.Salazar N, Munoz D, Kallifatidis G, Singh RK, Jorda M, Lokeshwar BL. The chemokine receptor CXCR7 interacts with EGFR to promote breast cancer cell proliferation. Molecular cancer. 2014;13:198. doi: 10.1186/1476-4598-13-198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Hernandez L, Magalhaes MA, Coniglio SJ, Condeelis JS, Segall JE. Opposing roles of CXCR4 and CXCR7 in breast cancer metastasis. Breast cancer research: BCR. 2011;13:R128. doi: 10.1186/bcr3074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Hattermann K, Held-Feindt J, Lucius R, Muerkoster SS, Penfold ME, Schall TJ, et al. The chemokine receptor CXCR7 is highly expressed in human glioma cells and mediates antiapoptotic effects. Cancer research. 2010;70:3299–308. doi: 10.1158/0008-5472.CAN-09-3642. [DOI] [PubMed] [Google Scholar]
- 56.Muller A, Homey B, Soto H, Ge N, Catron D, Buchanan ME, et al. Involvement of chemokine receptors in breast cancer metastasis. Nature. 2001;410:50–6. doi: 10.1038/35065016. [DOI] [PubMed] [Google Scholar]
- 57.Kang Y, Siegel PM, Shu W, Drobnjak M, Kakonen SM, Cordon-Cardo C, et al. A multigenic program mediating breast cancer metastasis to bone. Cancer cell. 2003;3:537–49. doi: 10.1016/s1535-6108(03)00132-6. [DOI] [PubMed] [Google Scholar]
- 58.Hirbe AC, Rubin J, Uluckan O, Morgan EA, Eagleton MC, Prior JL, et al. Disruption of CXCR4 enhances osteoclastogenesis and tumor growth in bone. Proceedings of the National Academy of Sciences of the United States of America. 2007;104:14062–7. doi: 10.1073/pnas.0705203104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Bauer KR, Brown M, Cress RD, Parise CA, Caggiano V. Descriptive analysis of estrogen receptor (ER)-negative, progesterone receptor (PR)-negative, and HER2-negative invasive breast cancer, the so-called triple-negative phenotype: a population-based study from the California cancer Registry. Cancer. 2007;109:1721–8. doi: 10.1002/cncr.22618. [DOI] [PubMed] [Google Scholar]
- 60.Haffty BG, Yang Q, Reiss M, Kearney T, Higgins SA, Weidhaas J, et al. Locoregional relapse and distant metastasis in conservatively managed triple negative early-stage breast cancer. Journal of clinical oncology: official journal of the American Society of Clinical Oncology. 2006;24:5652–7. doi: 10.1200/JCO.2006.06.5664. [DOI] [PubMed] [Google Scholar]
- 61.Liedtke C, Mazouni C, Hess KR, Andre F, Tordai A, Mejia JA, et al. Response to neoadjuvant therapy and long-term survival in patients with triple-negative breast cancer. Journal of clinical oncology: official journal of the American Society of Clinical Oncology. 2008;26:1275–81. doi: 10.1200/JCO.2007.14.4147. [DOI] [PubMed] [Google Scholar]
- 62.Donoghue M, Lemery SJ, Yuan W, He K, Sridhara R, Shord S, et al. Eribulin mesylate for the treatment of patients with refractory metastatic breast cancer: use of a “physician’s choice” control arm in a randomized approval trial. Clinical cancer research: an official journal of the American Association for Cancer Research. 2012;18:1496–505. doi: 10.1158/1078-0432.CCR-11-2149. [DOI] [PubMed] [Google Scholar]
- 63.Domanska UM, Timmer-Bosscha H, Nagengast WB, Oude Munnink TH, Kruizinga RC, Ananias HJ, et al. CXCR4 inhibition with AMD3100 sensitizes prostate cancer to docetaxel chemotherapy. Neoplasia. 2012;14:709–18. doi: 10.1593/neo.12324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Galsky MD, Vogelzang NJ, Conkling P, Raddad E, Polzer J, Roberson S, et al. A phase I trial of LY2510924, a CXCR4 peptide antagonist, in patients with advanced cancer. Clinical cancer research: an official journal of the American Association for Cancer Research. 2014;20:3581–8. doi: 10.1158/1078-0432.CCR-13-2686. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.