

Abstract
Agonism of human peroxisome proliferator-activated nuclear receptor gamma (PPARγ1) was recently observed in 15 of 25 samples of indoor dust extracts at environmentally relevant exposure levels. In this study, an effect-directed analysis approach was used to identify the primary contributors of PPARγ1 activity in the dust extracts. Three dust extracts showing significant PPARγ1 activity were fractionated with normal phase high-performance liquid chromatography (NP-HPLC) and each fraction was tested for PPARγ1 activity. Three dust extracts showed a similar PPARγ1 activity distribution in the NP-HPLC fractions. In most active fractions, fatty acids (FAs), including oleic acid, stearic acid, palmitic acid and myristic acid, were the primary chemicals identified using gas-chromatography mass spectrometry (GC-MS). Chemical measurements of the FAs in house dust extracts revealed a positive and significant correlation with the observed PPARγ1 activity. To test the role of FAs in the activity, a mixture of four FAs was prepared in the ratios measured in the dust samples and tested for activity. The activity of this mixture was 30–50% of the activity observed in the dust extracts, suggesting they were contributing to the observed activity, but also suggesting additional unknown compounds are likely still present in the dust extracts. To tentatively identify sources of FAs in the dust samples, FAs were quantified in human/animal hair, dead skin cells, and cooking oil. FAs were abundant in all samples and our data indicate that all of these may be sources to indoor dust.
Graphical abstract
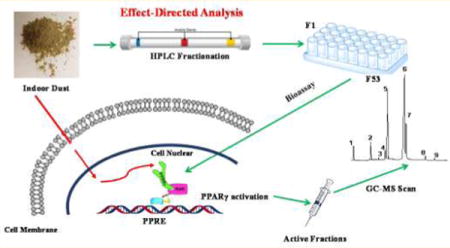
INTRODUCTION
Peroxisome proliferator-activated nuclear receptor gamma (PPARγ) is a master nuclear receptor that regulates lipid metabolism, cell proliferation signal transduction and cell differentiation.1 To date, several environmental contaminants have been shown to significantly bind and activate PPARγ signaling. Several well-established model chemicals include tributyltin (TBT), triphenyltin (TPT), and mono(2-ethylhexyl) phthalate bis(2-ethylhexyl) (MEHP) (a metabolite of the bis(2-ethylhexyl) phthalate or DEHP), which have been shown to upregulate and stimulate several PPARs.2,3 More recently, 2,2′,6,6′-tetrabromo bisphenol (TBBPA), 3,3′,5,5′-tetrachlor-obisphenol A (TCBPA), and triphenyl phosphates (TPP) were identified as partial agonists of PPARγ.4,5 As these chemicals are ubiquitously detected in indoor dust, PPARγ binding and activation of chemicals present in indoor dust extracts was recently investigated.6,7 Using a PPARγ1 (one isoform of PPARγ) reporter assay, we found that 15 of 25 dust extracts tested displayed significant activation, with a clear increase in activation with increasing dose. In some cases, activation was as high as 50% for the most potent dust extracts. The observed PPARγ1 activity could either be driven by some known PPARγ1 agonists, such as phthalates, triphenyl phosphate (TPP), and organotins, or some unknown ligands. Therefore, further research is needed to identify the primary chemical(s) driving the PPARγ1 activity in the indoor dust.
Effect-directed analysis (EDA) is a technique utilizing chemical separation, bioassay evaluation and qualitative chemical analysis to identify the chemical(s) leading to a specific biological response in complex mixtures. EDA usually includes the extraction, enrichment and stepwise fractionation of an environmental mixture with manipulations directed by a specific bioassay response. Chemicals in active fractions can then be identified through chemical analysis (e.g., mass spectrometry). To date, EDA has become a valuable approach to identify chemicals responsible for biological effects in various complex environmental samples (reviewed in8).
In the present study, our goal was to identify the active chemicals leading to PPARγ1 activity in indoor dust extracts. To this end, we first fractionated three dust extracts that showed relatively high PPARγ1 activity, and then subsequently tested the activity in each fraction. The active fractions were scanned using mass spectrometry and possible chemicals were identified. To confirm those possible candidates, the concentration of those chemicals in dust samples was measured and a correlation analysis was conducted with PPARγ1 activity in the dust extracts. The potential sources of those candidate chemicals were also investigated.
MATERIALS AND METHODS
Chemicals
Most of the chemicals used in this study were described in previous studies.6,7 Besides, oleic acid (OA, ≥ 99%), palmitic acid (PA, ≥ 99%), stearic acid (SA, ≥ 98.5%), myristic acid (MA, ≥ 99%), 31-deuterated palmitic acid (31d-PA, 99%), tris(2-carboxyethyl)phosphate (TCEP, 97%), tris (1-chloro-2-propyl) phosphate (TCPP), and tris(1,3-dichloro-2-propyl)phosphate (TDCPP) were purchased from Sigma-Aldrich (St Louis, MO). 4-Monochlorophenol (98%) was supplied from TCI America (Boston, MA). All solvents and other materials were of HPLC grade. All the cell culture reagents were from Life Technology Inc. (Grand Island, NY).
Sample Preparation
The indoor dust extracts were from our previous PPARγ binding and reporter assay.6,7 In brief, the indoor dust samples were collected from the main living areas of homes for Group A, D, and E. Dust samples in Group B and C were collected from gymnastics studios and office environments; respectively. All dust samples were extracted with acetone:hexane (1:1, v/v) using sonication, and then concentrated, filtered, and cleaned by gel permeation chromatography [GPC, Environgel GPC system (Waters, Milford, CA)]. Half of the extracts were reconstituted in dimethyl sulfoxide (DMSO) for bioassay testing and the other half was stored for the EDA. After our preliminary data indicated that fatty acids (FAs) may be the potential PPARγ ligands in the 25 dust extracts, 10 additional dust samples from Group D and E were then prepared and tested without using GPC cleanup to eliminate the concern that chemical composition might be changed after multiple cleanup steps. To identify potential sources of FAs to the indoor dust, animal hair was collected from one dog and one cat, and one sample of human hair was also collected. Furthermore, dandruff (n = 1) was collected by brushing the scalp using a solvent-cleaned wooden comb. A vegetable oil sample (soybean) and one cod fish liver oil (ISI BRANDS INC., American Fork, UT) sample were purchased from a local market. To analyze FAs in these samples, ~0.01 g of hair or dandruff were spiked with 31d-PA and extracted using the same method used for indoor dust samples. Oil samples were diluted with acetone, spiked with 31d-PA, and directly analyzed.
PPARγ1 Reporter Assay
As described in our previous study,7 a PPARγ1 reporter assay (GeneBLAzer PPARγ1 non-DA Assay, Invitrogen) was used to investigate the PPARγ1 activation of indoor dust extracts and their NP-HPLC prepared fractions. An amalar blue assay prepared from resazurin was used for the cell viability test. The PPARγ1 activity in the indoor dust after fractionation was expected to decrease due to the separation of the active chemical mixtures. To ensure that PPARγ activity was observed after fractionation, two higher doses (4200 and 1400 μg dust equivalent quantity (DEQ)/mL in assay medium) were used in the bioassay. In addition, cytotoxicity was found to decrease after the fractionation step, which was probably due to the separation of the toxic compounds in the dust matrix, and thus higher doses could therefore be tested.
Fractionation Procedures
Fractionation of indoor dust extracts was achieved using a normal phase high performance liquid chromatography (NP-HPLC) system consisting of a Waters 1525 Binary HPLC pump (Milford, CA) equipped with a degasser, a 2.5 mL injection loop and a Supelcosil LC-Diol column (250 × 10 mm, 5 μm particle size). To figure out the elution profile and optimize the gradient elution, a mixture including several typical flame retardants or their metabolites, halogenated phenols, phthalate metabolites, and organotin chloride was prepared and fractionated (Table S1 in the Supporting Information (SI)). These chemicals covered a wide range of physicochemical properties (e.g., log Kow) and some of them were known PPARγ agonists including TBBPA, phthalates, TPP, and organotins.2–5 Therefore, the elution profile could also help with a preliminary evaluation of the PPARγ1 activity contribution of several targeted compounds in the dust fractions. During optimization, 1 min fractions of the chemical mixture were collected and analyzed by mass spectrometry as described in the Chemical Analysis section. Finally, the following gradient elution with a total running time of 53 min was used: isocratic elution with hexane for 5 min, followed by gradient elution to 100% dichloromethane (DCM) within 15 min, then maintained at 100% DCM for 2 min, then followed by another gradient elution to 100% acetone within 15 min, and finally a column flush with 100% methanol for 16 min. A flow rate of 5 mL min−1 was used during the entire fractionation. Three dust extracts (i.e., dust I, II, and III), which showed high PPARγ activation in our previous study,7 were evaporated to near dryness under a gentle stream of nitrogen and reconstituted in hexane/DCM (1:1, v/v) to a final volume of 1 mL prior to injection. In Dust I, fractions were collected every 1.5 min, resulting in 30 fractions. After finding that most active fractions were in the first 20 min fractions, 1 min fractions were then collected for Dust II and III (53 fractions) to reduce the time window and increase the separation for each fraction. Each fraction was split for PPARγ1 activity testing and chemical analysis. One quarter of each fraction was evaporated under nitrogen gas and reconstituted in DMSO for PPARγ activity test and the remaining 75% was retained for chemical analysis. Preliminary experiments evaluated semipreparative reverse-phase HPLC (RP-HPLC, Nucleosil 100−5 C18) for the separation of Dust I (data not shown); however, most PPARγ activity was lost after fractionation, but could be mostly recovered in the insoluble fraction attached to the vial bottom (i.e., I–Fhydrophobic). Losses were therefore likely attributed to the high hydrophobicity of the active components, which were not soluble in the initial loading solution of methanol:H2O (1:1, v/v). Therefore, RP-HPLC fractionation was rejected in favor of NP-HPLC fractionation.
Chemical Identification
Fractions with significant PPARγ1 activity were subjected to qualitative mass spectrometric analysis by gas chromatography mass spectrometry (GC-MS, Agilent GC 6890N, MS 5975, Newark, DE) operated in full-scan mode (m/z 50–1050) using the electron ionization (EI) mode (GC/EI-MS). The chemical identification was based on our previous study9 and fully described in Text S1 Chemical Identification (SI). To further investigate how much of the PPARγ1 activity could be explained by the identified chemicals, a chemical mixture containing the equivalent concentrations of the primary compounds identified during the EDA was also prepared and tested for PPARγ1 activity.
Chemical Analysis
To determine the elution profile in the NP-HPLC fractionation, the prepared mixture in each fraction was analyzed using mass spectrometry. Specifically, 2-ethylhexyl-2,3,4,5-tetrabromobenzoate (TBB), bis(2-ethylhexyl)-tetrabromophthalate (TBPH), and polybrominated diphenyl ethers (PBDEs) were analyzed using electron capture negative chemical ionization mass spectrometry (GC/ECNI-MS). Organophosphates (e.g., TDCPP, TCPP, TPP, and TCEP) and phthalates (e.g., DEHP) were analyzed with GC/EI-MS. Both instruments used selected ion monitoring (SIM). Halogenated phenols, bisphenols, tetrabrominated benzoic acid (TBBA), organotins and other phthalate metabolites were analyzed using liquid-chromatography tandem mass spectrometry (LC-MS/MS, Agilent 6410B) in the multiplereaction monitoring (MRM) mode. Most of the SIM and MRM transition are described in our previous study.6 After identifying FAs as the possible source of the PPARγ1 activity in the dust extracts, the concentration of phthalates, several organophosphates, and FAs in 35 dust samples were quantified using GC-MS/EI. 31d-PA was used as the internal standard for all the FAs analyzed. The quantifiers and qualifiers of FAs and phthalates are listed in SI Table S2. To keep chemical concentrations and the PPARγ1 activity comparable, ~5 μL of the dust extracts in the PPARγ dosing stock solution was directly transferred to ~150 μL glass microvolume insert and ~100 μL extra acetone was added.
Data Analysis and Statistical Analyses
PPARγ1 activation% was calculated to describe the relative potency/efficacy normalized to the maximal response from rosiglitazone as fully described in our previous study.7 All statistical analyses were conducted using SigmaPlot 12.0 (Systat Software Inc.), testing hypotheses at α = 0.05, and all tests were two-tailed. For the dust extracts and their fractions, a one way ANOVA was conducted and Newman-Keuls posthoc test was used to identify which doses were significantly different from the DMSO control and the procedural control; respectively. To examine the relationship between the PPARγ1 activation and the chemical concentrations in the dust extracts, Pearson correlation and Spearman correlation analyses were conducted with the 25 dust extracts (normally distributed) and 10 raw dust extracts (non-normally distributed); respectively. To identify the possible sources of the FAs in the indoor dust, the contribution (%) of each individual FA to the sum of the FAs in the house dust extracts and several possible sources were calculated and a principle component analysis (PCA, SPSS 12.0.1, IBM) was conducted.
Quality Control
For each dosing level, triplicate analyses were run in 384 well plates. To reduce the variability between batches, similar passages (6th or 7th) of cells were collected and used for the reporter assay. To ensure the dose–response of each dust extract fraction was comparable, all fractions from one dust extract were run at the same time on one plate. A solvent blank was prepared for both fractionation procedures and chemical analyses in the house dust. Due to the large number of fractions, the PPARγ1 reporter assay with the corresponding blank fraction was only run for the active fraction in each dust extract. All the chemical concentrations in the house dust were blank corrected.
RESULTS
Fractionation of Dust Extracts and PPARγ1 Activation in the Fractions
To first ensure that a majority of the targeted analytes were well separated with our gradient elution profile, we first ran a chemical mixture (see Materials and Methods) through the HPLC and examined their elution behavior. As shown in SI Table S1, log Kow strongly influenced the fractionation process, but all chemical classes were well separated. Nonpolar chemicals such as PBDEs, TBB, TBPH, and phthalates were eluted as early as 3–4 min in the hexane fraction, which was followed by the organophosphates flame retardants (i.e., TPP, TDCPP, TCPP, and TCEP). Halogenated phenolic compounds including 2,4,5-tribromophenol (2,4,5-TBP), 2,4,6-TBP, monochlorophenol, and 6-hydroxylated-2,2′,4,4′-tetra-bromodiphenyl ether (6-OH-BDE47) eluted primarily between 17 and 21 min, which was in the DCM fraction. The polar compounds including metabolites of phthalates, TBBA, and organotins were eluted after 40 min during the 100% methanol flush. The elution of organotins on the NP-HPLC column lasted over 8 min, suggesting these compounds have a very strong retention on the normal phase column.
Fractionation was then performed on dust samples I, II, and III and PPARγ1 activity was tested individually for each fraction. In our previous study,7 these three dust extracts showed a significant dose–response relationship and the maximal PPARγ activation was ~53%, 25%, and 30% of the maximal response of rosiglitazone; respectively. As shown in Figure 1, a clear dose–response relationship was observed in several specific fractions in all dust extracts after fractionation. No obvious PPARγ1 activity was observed in the corresponding blanks (data not shown). For Dust I, the active fractions included those from fraction 7 (I–F7) to I–F16, and I–F22. The corresponding RT was 10–23 and 33 min, respectively, since each fraction was collected over 1.5 min in this dust sample. The most active fractions were I–F7 (RT: 9–10.5 min), I–F8 (RT: 10.5–12 min), I–F12 (RT: 16.5–18.0 min) and I–F22 (RT: 31.5–33 min). In Dust sample II, significant PPARγ1 activation was observed in the fractions from II–F11 to II–F18 (i.e., RT: 10–18 min) and II–F32 (RT: 32 min). The most active fractions were II–F11, II–F12, II–F13, and II–F17. In general, the fraction-dependent PPARγ1 activity pattern in Dust II was very similar to that observed in Dust I. However, in Dust III, the most active fraction was III–F4 (3–4 min) with the highest activation of 53% at the dosing concentration of 4,200 μg DEQ/mL. Besides that, other active fractions included III–F11, III–F12, III–F14, III–F16, and III–F32, which were very similar to the pattern observed in both Dust I and II.
Figure 1.
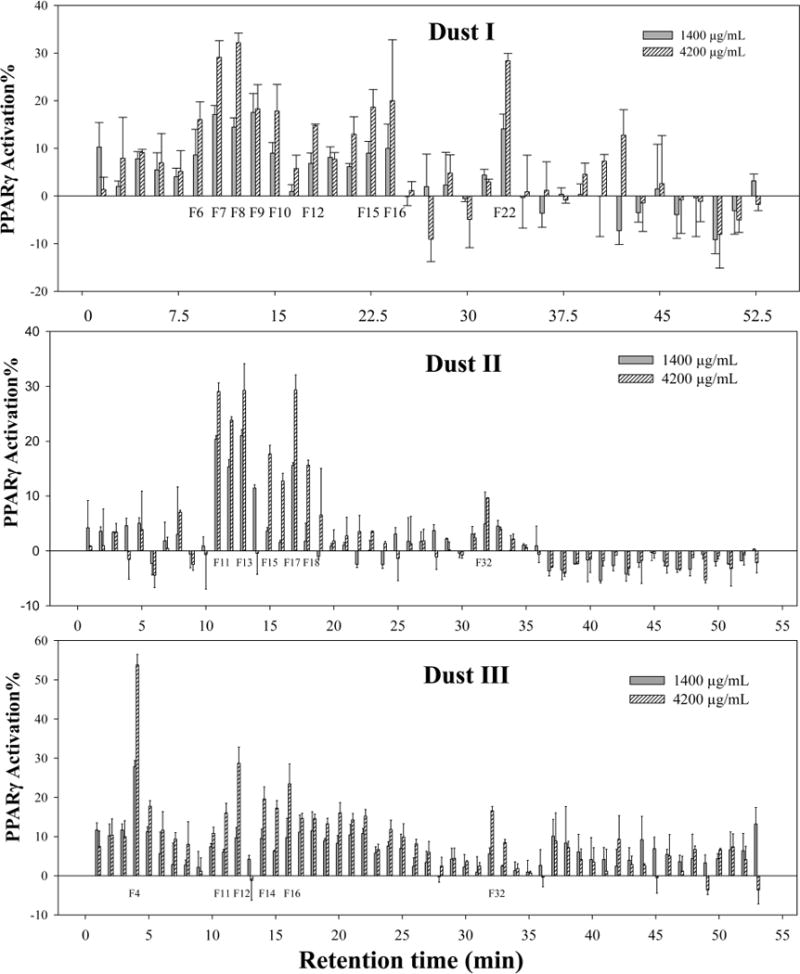
PPARγ1 activation (%) at dosing concentrations of 1400 and 4200 μg DEQ/mL with (a) 35 NP-HPLC fractions from Dust I (1.5 min/fraction); (b) 53 fractions from Dust II (1.0 min/fraction); (c) 53 fractions from Dust III (1.0 min/fraction). Values represent the average of the triplicates, and error bars represent standard deviation.
Chemical Composition of the Toxic Fractions
The most active fractions from the three dust extracts after NP-HPLC fractionation and the insoluble fraction I–Fhydrophobic from Dust I (see the Fractionation Procedures section) were then analyzed by GC-MS/EI in full scan mode. The total ion chromatographs (TIC) as well as our identification of compounds present in these fractions are presented in SI Table S3 and Figures S1–3. Figure 2 provides a specific example of the TIC generated from F8 collected from Dust I. The largest responses in the most active fractions (RT: 10–18 min) from the three dust extracts were identified as FAs and included lauric acid (LA, match factor >90%), myristic acid (MA), stearic acid (SA), oleic acid (OA), and palmitic acid (PA). The latter four compounds were confirmed by comparison to authentic standards. Though some other compounds such as TPP, TDCPP, and tris(2-butoxyethyl)-phosphate (TBOEP) were also identified in the active fractions (e.g., I–F8 (RT: 10.5–11.5 min), II–F11 (RT: 10–11 min), and III–F12 (RT: 11–12 min) in SI Figure S1–3), the relative PPARγ1 activation of these compounds at the measured concentrations in the dust tended to exclude the possibility that they were the active components in the dust extracts. For example, a similar amount of TPP was eluted in II–F10 and II–F11 of Dust II (SI Figure S2); however, much higher activity was observed in II–F11, suggesting other chemicals other than TPP were responsible for the observed activity. TDCPP and TBOEP did not show much PPARγ1 activity in the assay.7 Therefore, these chemicals did not seem likely to be responsible for the observed activity. In III–F4 from Dust III, very large amounts of FAs and phthalates, such as benzyl butyl phthalate (BBzP) and DEHP, were observed. According to SI Table S1, the elution of phthalates in this fraction was expected using NP-HPLC. However, no FAs were detected in the fractions from Dust I and II with similar RT (Data not shown). The early elution of FAs in Dust III suggested that high PPARγ1 activity in this fraction could be caused by poor retention of the chemicals, which might be due to the nonequilibrium of the column when loading the dust extract or a matrix effect. In I–F22 (RT: 32 min) in Dust I and II/III–F32 (RT: 32 min) in Dust II and III, significant PPARγ1 activity was observed. Bis(2-ethylhexyl) fumarate (BEHF), a typical phthalate replacement, was found in I–F22 of Dust I and confirmed with an authentic standard. However, the PPARγ1 reporter assay demonstrated that this chemical did not activate PPARγ1 at 100 μM. Another possible compound, azelaic acid (AzA), was identified in II–F32 of Dust II with a match factor of 60% with the NIST library. However, a pure standard would be needed to further confirm this compound and its PPARγ1 activity. Therefore, the identity of the active components in this fraction remain unidentified after GC/EIMS analysis. As shown in SI Figure S1, the TIC of the fraction I–Fhydrophobic was very similar to the pattern in the most active fractions including I–F7 and I–F8, suggesting that the active compounds in the dust extract were very hydrophobic.
Figure 2.
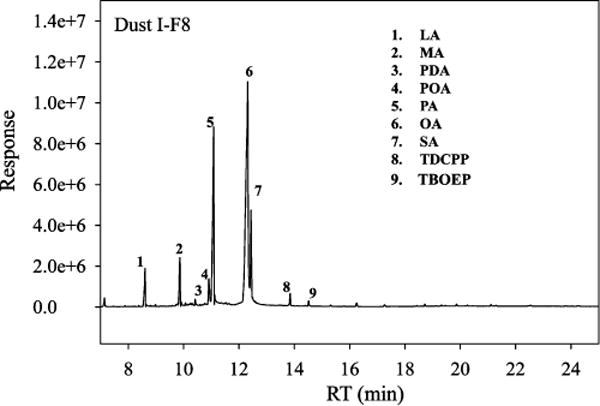
Full scan of F8 collected from the Dust I highlighting the chemicals identified by GC-MS/EI based on comparisons with authentic standards.
PPARγ1 Activity of FAs
To confirm the hypothesis that FAs were the primary contributors of PPARγ1 in the indoor dust samples, the PPARγ1 activation of several identified FAs was investigated. As shown in Figure 3(a), a dose–response relationship was observed for OA, SA, PA, and MA. However, the PPARγ1 activities varied greatly between compounds. The unsaturated OA was the most potent in the activation of PPARγ1, which began to activate PPARγ1 at low micromolar concentrations and the highest activation was close to 40% of the maximal response of rosiglitazone. The saturated MA began to activate PPARγ1 at 11 μM and the maximal activation (~30%) was less than that of the OA. SA and PA showed a much weaker PPARγ activation with the maximal activation less than 15% of the rosiglitazone.
Figure 3.
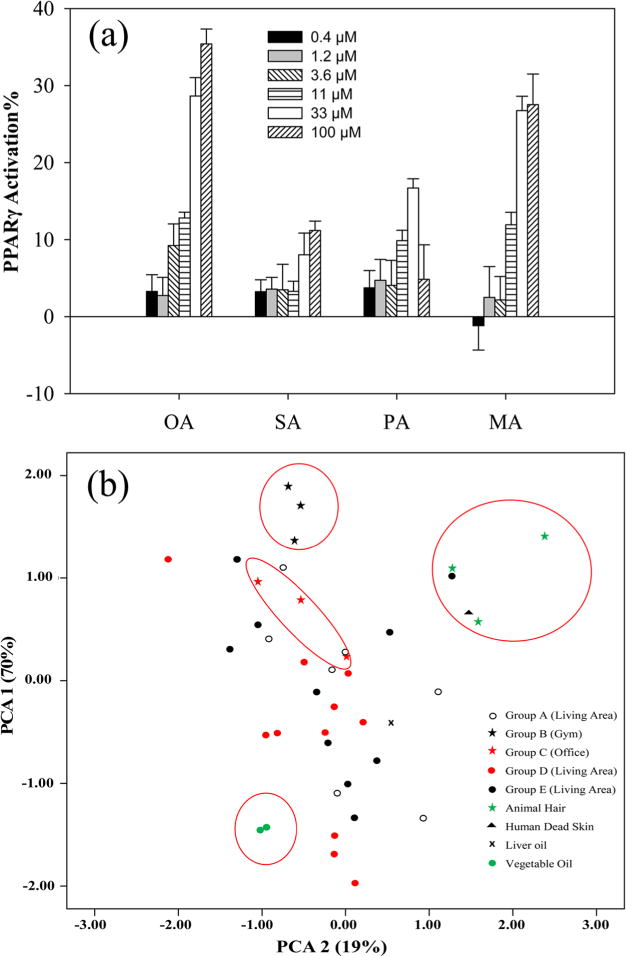
(a): Dose–response relationship of PPARγ activation by four FAs including OA, SA, PA, and MA. Values represent the average of triplicate measurements, and error bars represent standard deviation; (b): Score plot of the contribution (%) of OA, PA, SA, and MA to the sum of the FAs in 35 house dust extracts and several possible sources using principle component analysis (PCA).
Correlation between PPARγ1 Activity and Chemical Concentration
After we observed high abundances of FAs, phthalates and several organophosphates in the PPARγ1 active fractions, we then measured these chemicals in extracts of 25 dust samples. As shown in SI Table S4, the phthalates and FAs were the most abundant components in the dust extracts. DEHP and diisononyl phthalate (DiNP) were the most abundant phthalates with a median concentration of 113.5 and 118.83 μg/g dust; respectively, which were followed by BBzP (median: 16.6 μg/g dust), dibutyl phthalate (DBP, median: 16.06 μg/g dust), and diisobutyl phthalate (DiBP, 8.25 μg/g dust). As for FAs, the median concentrations were 1,104 (range: NA–3,732 μg/g dust), 1,320 (range: 16–4150 μg/g dust), 324 (range: 7.5–1781 μg/g dust), and 244 μg/g dust (range: 3.9–832 μg/g dust) for OA, PA, SA, and MA; respectively. The phthalate concentrations measured in these dust extracts were similar to measures reported in previous studies.10–13 For example, the median concentrations were 13, 29, 480, and 80 μg/g dust for BzBP, DBP, DEHP, and DINP; respectively based on measurements in 278 dust samples in a German study.12
After measuring the concentration of FAs, phthalates and several organophosphates, the correlation between PPARγ1 activity and chemical analyte concentrations were analyzed. As shown in Figure 4 and SI Table S6, a positive and significant correlation was observed between the levels of individual FA and PPARγ1 activation at a dosing level of 200 and 600 DEQ/mL (e.g, r = 0.647 and p < 0.00002 for OA). However, no significant relationship was observed between PPARγ1 activity and phthalates or organophosphates. The correlation of each FA level was also very positive and significant with each other (r > 0.8 and p < 0.0000001), suggesting that many identified FAs might have similar sources. Based on the PPARγ1 activity and abundance of four FAs in the dust, PA, SA, and MA were likely to play less important roles than OA in the observed PPARγ1 activity. Many dust extracts showed clear cyto-toxicity at the highest dose (~1818 DEQ/mL) and the PPARγ1 activity at the lower dose (Level 5 and 6) was not significantly different from the DMSO control. The correlation analysis between FA levels and PPARγ1 activity in an additional 10 dust extracts that were not purified using GPC was also investigated, and again a very significant correlation was observed (SI Figure S4 and Table S7). All these findings suggest that the FAs, especially OA, in the dust are the primary compounds driving the observed PPARγ1 activity. To further investigate how much of the PPARγ1 activity could be explained by the identified FAs, a mock mixture containing the equivalent concentration of four of the major identified FAs (e.g., OA, PA, SA, and MA) in three dust extracts (two were the extracts used for NP-HPLC fractionation and one from 10 fresh dust extracts) was prepared and tested for its PPARγ1 activity. As shown in SI Figure S6, the mixture could explain ~30–50% of the observed activity in the equivalent dust extract at 606 μg DEQ/mL. However, when a dosing solution that was three times the concentration of the mixture in the Dust II and IV samples was tested (SI Figure S6), the observed activity was very similar or even higher than the activity in the dust extracts.
Figure 4.
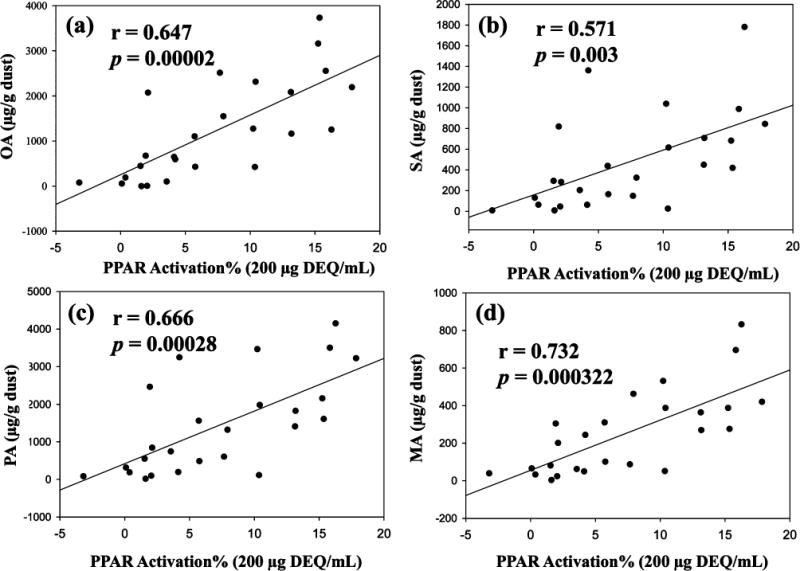
Linear regression between the PPARγ activation (%) at s dose of 200 μg DEQ/mL and the concentration of (a) oleic acid OA, (b) PA, (c) SA and (d) MA in 25 dust extracts.
Possible Sources of FAs in the Indoor Dust
To the authors’ knowledge, there is no information available on the sources and levels of FAs in indoor dust. Possible sources could be evaporation during cooking processes with oil and deposition into dust, or debris accumulating in dust from animal hair or dead skin cells. To provide more information on possible sources of FAs in indoor dust, the concentration of four FAs were measured in hair samples collected from a cat, dog and human, and in a sample of human skin debris. Samples were also taken of cooking oil and fish liver oil. As shown in SI Table S8, very high level of FAs were detected in all samples and the highest was observed in the human skin debris and oil samples with concentrations as high as 14,868 μg/g sample. To potentially identify the possible sources for FAs in dust, the congener profiles of FAs were compared in different samples and analyzed by PCA (see SI Figure S5 and Figure 3(b)). In the cooking oil, the level of OA was more than 50% of the total four FAs, which was followed by the fish liver oil. In the hair samples and skin debris, the contribution of OA was less than 20% and dominated by PA. The OA contribution also varied greatly between different dust samples. Overall, OAs contribution in the dust from Groups A, D, and E (collected from main living areas) (Average ± SD: 43.1 ± 19.7%, n=28) was significantly higher than that from dust collected from gym and office sources (Group B and C: 13.6 ± 11.8%, n = 6) using the student t-test (p < 0.01). In the PCA score plot (Figure 3(b)), dust samples from Group B and C were closer to the animal hairs and dead skins, while many of the samples from the indoor living area were closer to the vegetable cooking oils.
DISCUSSION
PPARγ1 is a master nuclear receptor that regulates lipid metabolism, cell proliferation, and differentiation. In our previous study, several organic contaminants, or their metabolites, present in indoor dust extracts displayed significant PPARγ1 activity.7 The present study was a follow-up investigation on the PPARγ1 binding and activation by organic contaminants and the chemical mixtures in the indoor dust. Using an EDA, FAs in indoor dust were found to outcompete with other possible PPARγ ligands such as phthalates, TPP and organotins, and were identified as the primary chemicals resulting in the observed PPARγ1 activities. Overall, this study is the first report of FAs in house dust and demonstrates that they are primary PPARγ1 ligands.
Some chemicals such as organotins, phthalates and TPP were expected to play an important role in the observed PPARγ1 activity in dust extracts. Organotins could activate PPARγ at the nanomolar level using the PPARγ1 reporter assay7 and have been identified as strong to moderate PPARγ ligands both in vitro and in vivo.3 However, during the NP-HPLC fractionation, very little activity was observed in the late eluting fractions, suggesting organotins are likely not important contributors to the observed PPARγ1 activity in the indoor dust extracts since the RT of TBT and TPT ranged from 42 to 50 min. In a previous study, TBT was detected with a median concentration of 22 ng/g dust (range: <2 ng/g to 300 ng/g) and TPT was not detected.14 In a preliminary experiment using spiked dust samples we found that our current extraction method could recover >80% of any organotins present in the dust samples (data not shown). If 1000 μg DEQ/mL dosing concentration and the maximal concentration of TBT (i.e., 300 ng/g dust) was used in the reporter assay, the calculated TBT concentration was approximately 0.92 nM, which was just above the nonobserved adverse effect level (NOAEL: 0.50 nM) for TBT.7 Therefore, organotins present in indoor dust samples are likely not causing much PPARγ activity. Another possible group of chemicals contributing to activity are the phthalates with concentrations often reaching the mg/g range in indoor dust. In our previous study, BzBP itself showed significant PPARγ1 activation at the micromolar level.7 In the current study, BzBP was measured with a median concentration of 16.1 μg/g dust (range: 0.36–181.1 μg/g dust). A similar calculation was conducted for BzBP and the dosing concentration could reach 0.6 μM in the reporter assay if the maximal concentration of BzBP in the dust was used. The NOAEL was 0.13 μM for BzBP in the PPARγ1 reporter assay and again this chemical might not be a potent contributor for the observed PPARγ1 activity either. In contrast, the level of FAs such as OA has a median concentration of 1104 μg/g dust with a maximal concentration of 3,732 μg/g dust. Similarly, the maximal calculated OA from dust extract in the reporter assay was 13 μM at 1000 μg DEQ/mL, which could lead to a PPARγ1 activation of approximately 15% in the reporter assay. However, it should be noted that some other unidentified FAs may be present in the indoor dust that were not detected by the GC-MS full scan. The observed PPARγ1 activity could likely be a result of contributions from several of the FAs identified in the mixture. The fact that the mixture of four FAs could explain ~30–50% of the observed activity in the dust extracts suggests that some unidentified PPARγ1 ligands are still present in the dust extracts. Further research is needed to identify these active compounds.
PPAR is thought to be an orphan nuclear receptor, implying no specific endogenous ligands have been identified. Despite intensive research efforts, it remains to be determined whether PPAR has a highly specific natural ligand or whether it is activated by the combined concentration of weakly activating ligands. A variety of chemicals have been suggested to be natural PPAR ligands, including eicosanoids,15 components of oxidized low-density lipoproteins,16 and oxidized alkyl phospholipids including lysophosphatidic acid17 and nitrolinoleic acid.18 The prostaglandin J2 derivative, 15d-PGJ2, does not naturally exist at sufficient concentrations to activate PPARγ1 in mammalian cells.19 FAs are major components of biological cell membranes that play important roles in intracellular signaling and as precursors for ligands that bind to nuclear receptors. Six FAs including myristic, palmitic, palmitoleic, stearic, oleic, and linoleic acid constituted the majority of the FA mixtures in every tissue sample. Among them, OA and PA identified in this study were the most abundant FAs in human tissues. The mean percentage of OA ranged from 47.3% to 52.0% of the total fatty acids in adipose tissue depending on anatomical site and age-race group.20 According to a previous study, the concentration of free fatty acids in serum can be as high as several hundred micromolar.21 Consistent with the PPARγ1 activity observed for FAs in this study, cells treated with OA displayed PPARγ gene activation in three hepatocytic cell lines.22 Therefore, it is possible the FAs themselves might be important activators for PPARγ in vivo.
To date, no measurements of FAs in dust have been reported, and the sources of FAs remain unidentified. Though it is not the main focus of the current study, it is still interesting to tentatively identify the potential sources of FAs in dust. In this study, several possible sources of FAs including hair, dead skin cells and cooking oils were investigated. High levels of FAs were detected in all those samples, especially in the cooking oil and dead skin cells. However, the congener pattern differed greatly between samples. Vegetable oil has a higher level of unsaturated OA than other sources. In the animal hair samples, saturated PA is the most dominant FA, accounting for more than 50%. This difference could be potentially used to identify the contribution of FAs in the dust extracts. For example, dust samples from Group A, D and E, tended to have a higher level of OA, compared with Groups B and C. The former groups were collected from the main living rooms where cooking activity may occur frequently. In contrast, dust samples from Groups B and C were collected from gymnasiums and offices, where the primary source should be from human dead skin debris or hair. Also, the level of FAs in dust from Group B (n = 3) was the lowest in all samples. Therefore, cooking activities may be an important source for the elevated levels of OAs in indoor dust. However, it should be noted that very few samples have been analyzed in this study, which might increase the uncertainties of these results. Further studies should be conducted to investigate the relationship between the level of FAs in dust and cooking activities. Also, it should be noted that FA exposure from house dust ingestion is not expected to be the primary exposure source, and other pathways such as food intake could be more important sources of human FA exposure.
This study also showed that the natural ligands could outcompete with synthetic compounds in environmentally relevant mixtures to activate PPARγ. The coexposure of natural ligands and environmental contaminants should be considered in future studies, which is highlighted in the field of exposomics, focuses on an individual’s entire chemical exposure. Environmentally relevant samples such as house dust which contain both natural ligands and synthetic ligands could be a very important matrix to evaluate for health effects from realistic exposures. Combined with an EDA approach, house dust could serve as a possible surrogate to estimate health effects from realistic SVOC indoor exposures and help prioritize chemicals for future studies. However, it should be noted that this study failed to consider the effect of bioactivation of each chemical. For example, the in vivo half-lives of most phthalates are estimated to be less than 3 h, and the in vivo bioactivation of those chemicals can be effective in increasing activity.23 The HEK 293H cell used in the reporter assay is not expected to have a significant xenobiotic metabolic activity. The bioactivated phthalate (e.g., MEHP) could be more potent than its parent compound DEHP in activating PPARγ1.7 Also, the potency of the phthalates metabolites (e.g., MEHP, EC20: 2 μM) can be more potent than the FAs (e.g., OA, EC20: 20 μM) in activating PPARγ1. In this study, the levels of phthalates, such as DEHP, were found to be similar in magnitude to the FAs. Therefore, bioactivation might change the contribution of these ligands to total activity and should be investigated in the future. Another potential limitation is that the PPARγ-GAL4 assay used in this study did not include the ligand-binding domain of the RXR nuclear receptor heterodimerized with PPARγ. Therefore, it might underestimate the effect of dual ligands such as organotins (TBT).
Supplementary Material
Acknowledgments
Thanks to Laura Macaulay and Dr. Erin Kollitz from Duke University for supplying the animal hair. This study was funded by grants from the National Institute of Environmental Health Sciences (R01ESO16099 and R01 ES015829).
Footnotes
Supporting Information
Supporting Information is provided as additional information as noted in text. The Supporting Information is available free of charge on the ACS Publications website at DOI: 10.1021/acs.est.5b01524.
Notes
The authors declare no competing financial interest.
References
- 1.Lehrke M, Lazar MA. The many faces of PPARgamma. Cell. 2005;123:993–9. doi: 10.1016/j.cell.2005.11.026. [DOI] [PubMed] [Google Scholar]
- 2.Feige JN, Gelman L, Rossi D, Zoete V, Metivier R, Tudor C, Anghel SI, Grosdidier A, Lathion C, Engelborghs Y, Michielin O, Wahli W, Desvergne B. The endocrine disruptor monoethyl-hexyl-phthalate is a selective peroxisome proliferator-activated receptor gamma modulator that promotes adipogenesis. J Biol Chem. 2007;282:19152–19166. doi: 10.1074/jbc.M702724200. [DOI] [PubMed] [Google Scholar]
- 3.Grün F, Watanabe H, Zamanian Z, Maeda L, Arima K, Chubacha R, Gardiner DM, Kanno J, Iguchi T, Blumberg B. Endocrine-disrupting organotin compounds are potent inducers of adipogenesis in vertebrates. Mol Endocrinol. 2006;20:2141–2155. doi: 10.1210/me.2005-0367. [DOI] [PubMed] [Google Scholar]
- 4.Pillai HK, Fang M, Beglov D, Kozakov D, Vajda S, Stapleton HM, Webster TF, Schlezinger JJ. Ligand Binding Activation of PPARgamma by Firemaster(R) 550: Effects on Adipogenesis Osteogenesis. Environ Health Perspect. 2014;122:1225–1232. doi: 10.1289/ehp.1408111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Riu A, Grimaldi M, le Maire A, Bey G, Phillips K, Boulahtouf A, Perdu E, Zalko D, Bourguet W, Balaguer P. Peroxisome Proliferator-Activated Receptor gamma Is a Target for Halogenated Analogs of Bisphenol A. Environ Health Perspect. 2011;119:1227–1232. doi: 10.1289/ehp.1003328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Fang M, Webster TF, Ferguson PL, Stapleton HM. Characterizing the Peroxisome Proliferator-Activated Receptor (PPARgamma) Ligand Binding Potential of Several Major Flame Retardants, Their Metabolites, Chemical Mixtures in House Dust. Environ Health Perspect. 2014;123:166–72. doi: 10.1289/ehp.1408522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Fang M, Webster TF, Stapleton HM. Activation of Human Peroxisome Proliferator-Activated Nuclear Receptors (PPARγ) by Semi-Volatile Compounds (SVOCs) and Chemical Mixtures in Indoor Dust. Environ Sci Technol. 2015 doi: 10.1021/acs.est.5b01523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Brack W, Klamer HJC, de Ada ML, Barcelo D. Effect-directed analysis of key toxicants in European river basins - A review. Environ Sci Pollut Res. 2007;14:30–38. doi: 10.1065/espr2006.08.329. [DOI] [PubMed] [Google Scholar]
- 9.Fang M, Getzinger GJ, Cooper EM, Clark BW, Garner LV, Di Giulio RT, Ferguson PL, Stapleton HM. Effect-directed analysis of Elizabeth River porewater: Developmental toxicity in zebrafish (Danio rerio) Environ Toxicol Chem. 2014;33:2767–74. doi: 10.1002/etc.2738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Butte W, Heinzow B. Pollutants in house dust as indicators of indoor contamination. Rev Environ Contam T. 2002;175:1–46. [PubMed] [Google Scholar]
- 11.Fromme H, Lahrz T, Piloty M, Gebhart H, Oddoy A, Ruden H. Occurrence of phthalates musk fragrances in indoor air dust from apartments kindergartens in Berlin (Germany) Indoor Air. 2004;14:188–95. doi: 10.1111/j.1600-0668.2004.00223.x. [DOI] [PubMed] [Google Scholar]
- 12.Nagorka R, Scheller C, Ullrich D. Plasticizer in house dust. Gefahrst Reinhalt L. 2005;65:99–105. [Google Scholar]
- 13.Rudel RA, Camann DE, Spengler JD, Korn LR, Brody JG. Phthalates, alkylphenols, pesticides polybrominated diphenyl ethers and other endocrine-disrupting compounds in indoor air dust. Environ Sci Technol. 2003;37:4543–53. doi: 10.1021/es0264596. [DOI] [PubMed] [Google Scholar]
- 14.Kannan K, Takahashi S, Fujiwara N, Mizukawa H, Tanabe S. Organotin Compounds, Including Butyltins and Octyltins in House Dust from Albany New York USA. Arch Environ Contam Toxicol. 2010;58:901–907. doi: 10.1007/s00244-010-9513-6. [DOI] [PubMed] [Google Scholar]
- 15.Desvergne B, Wahli W. Peroxisome proliferator-activated receptors: nuclear control of metabolism. Endocr Rev. 1999;20:649–88. doi: 10.1210/edrv.20.5.0380. [DOI] [PubMed] [Google Scholar]
- 16.Nagy L, Tontonoz P, Alvarez JG, Chen H, Evans RM. Oxidized LDL regulates macrophage gene expression through ligand activation of PPARgamma. Cell. 1998;93:229–40. doi: 10.1016/s0092-8674(00)81574-3. [DOI] [PubMed] [Google Scholar]
- 17.McIntyre TM, Pontsler AV, Silva AR, St Hilaire A, Xu Y, Hinshaw JC, Zimmerman GA, Hama K, Aoki J, Arai H, Prestwich GD. Identification of an intracellular receptor for lysophosphatidic acid (LPA): LPA is a transcellular PPARgamma agonist. Proc Natl Acad Sci USA. 2003;100:131–136. doi: 10.1073/pnas.0135855100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Schopfer FJ, Lin Y, Baker PR, Cui T, Garcia-Barrio M, Zhang J, Chen K, Chen YE, Freeman BA. Nitrolinoleic acid: an endogenous peroxisome proliferator-activated receptor gamma ligand. Proc Natl Acad Sci USA. 2005;102:2340–5. doi: 10.1073/pnas.0408384102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Bell-Parikh LC, Ide T, Lawson JA, McNamara P, Reilly M, FitzGerald GA. Biosynthesis of 15-deoxy-delta12,14-PGJ2 and the ligation of PPARgamma. J Clin Invest. 2003;112:945–55. doi: 10.1172/JCI18012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kokatnur MG, Oalmann MC, Johnson WD, Malcom GT, Strong JP. Fatty acid composition of human adipose tissue from two anatomical sites in a biracial community. Am J Clin Nutr. 1979;32:2198–205. doi: 10.1093/ajcn/32.11.2198. [DOI] [PubMed] [Google Scholar]
- 21.Zhang JW, Zhao Y, Xu CF, Hong YN, Lu HL, Wu JP, Chen Y. Association between serum free fatty acid levels and nonalcoholic fatty liver disease: a cross-sectional study. Sci Rep. 2014;4:5832. doi: 10.1038/srep05832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Ricchi M, Odoardi MR, Carulli L, Anzivino C, Ballestri S, Pinetti A, Fantoni LI, Marra F, Bertolotti M, Banni S, Lonardo A, Carulli N, Loria P. Differential effect of oleic and palmitic acid on lipid accumulation and apoptosis in cultured hepatocytes. J Gastroenterol Hepatol. 2009;24:830–40. doi: 10.1111/j.1440-1746.2008.05733.x. [DOI] [PubMed] [Google Scholar]
- 23.Hoppin JA, Brock JW, Davis BJ, Baird DD. Reproducibility of urinary phthalate metabolites in first morning urine samples. Environ Health Perspect. 2002;110:515–8. doi: 10.1289/ehp.02110515. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.