

Abstract
Background
The enzymatic degradation of lignocellulosic materials by fungal enzyme systems has been extensively studied due to its effectiveness in the liberation of fermentable sugars for bioethanol production. Recently, variants of the fungus Penicillium echinulatum have been described as a great producer of cellulases and considered a promising strain for the bioethanol industry.
Results
Penicillium echinulatum, wild-type 2HH and its mutant strain S1M29, were grown on four different carbon sources: cellulose, sugar cane bagasse pretreated by steam explosion (SCB), glucose, and glycerol for 120 h. Samples collected at 24, 96, and 120 h were used for enzymatic measurement, and the 96-h one was also used for secretome analysis by 1D-PAGE LC–MS/MS. A total of 165 proteins were identified, and more than one-third of these proteins belong to CAZy families. Glycosyl hydrolases (GH) are the most abundant group, being represented in larger quantities by GH3, 5, 17, 43, and 72. Cellobiohydrolases, endoglucanases, β-glycosidases, xylanases, β-xylosidases, and mannanases were found, and in minor quantities, pectinases, ligninases, and amylases were also found. Swollenin and esterases were also identified.
Conclusions
Our study revealed differences in the two strains of P. echinulatum in several aspects in which the mutation improved the production of enzymes related to lignocellulosic biomass deconstruction. Considering the spectral counting analysis, the mutant strain S1M29 was more efficient in the production of enzymes involved in cellulose and hemicellulose degradation, despite having a nearly identical CAZy enzymatic repertoire. Moreover, S1M29 secretes more quantities of protein on SCB than on cellulose, relevant information when considering the production of cellulases using raw materials at low cost. Glucose, and especially glycerol, were used mainly for the production of amylases and ligninases.
Electronic supplementary material
The online version of this article (doi:10.1186/s13068-016-0476-3) contains supplementary material, which is available to authorized users.
Keywords: Lignocellulosic biomass, Biofuels, Penicillium echinulatum strains, Secretome, CAZymes
Background
One of the global priorities is the development of alternative energy sources other than fossil fuels, and the use of cellulosic biomass has the potential to contribute to meeting the demand for liquid fuel [1]. Lignocellulose is the most abundant natural material present on earth and is composed of the polyphenol lignin and three polysaccharides: cellulose, hemicellulose, and pectin [2, 3]. These components are strongly intermeshed and chemically bonded by noncovalent forces and covalent crosslinkages, forming an intricately linked network that provides strength to the plant cell wall [4].
The manufacturing processes of saccharification and bioproducts from lignocellulose biomass are complex and lengthy ones [5]. One approach to depolymerization of plant cell wall polysaccharides involves using hydrolytic enzymes produced by some species of bacteria and fungi [6]. The degradation of lignocellulosic materials into monomeric sugars is of major importance, since the fermentable sugars can be used as raw materials in many biotechnological processes, including the production of second-generation ethanol [7]. In nature, fungi perform a major role in the degradation of plant biomass, producing an extensive set of a carbohydrate degrading enzyme specifically dedicated to break down plant cell wall polysaccharides. However, these varied sets differ between the species of fungi and the carbon sources employed. For example, while Trichoderma reesei has a set of highly efficient enzymes in the degradation of cellulose [8, 9], Aspergillus species produce many enzymes for degrading pectin [10].
Penicillium species have been reported to produce enzyme systems with higher performance than T. reesei and A. niger [11–14]. Among the species of Penicillium, P. echinulatum has been the focus of attention because of its potential to produce high titers of cellulases and has been considered a promising strain for the industry of second-generation ethanol [15–22]. All the mutant strains used for the production of cellulases are derived from the wild type called 2HH that was isolated from larvae of coleoptera Anobium punctatum. A mutant strain of P. echinulatum 9A02S1 was obtained from the strain 2HH, after several steps of mutagenesis, characterized by being a mutant partially depressed by glucose. This microorganism is deposited in the Deutsche Sammlung von Mikroorganismen und Zellkulturen–DSM 18942. The mutant strain S1M29 of P. echinulatum was obtained from the 9A02S1 strain through employing hydrogen peroxide mutagenesis and a selection of mutants in a medium supplemented with 2-deoxyglucose [15]. This mutant S1M29, due to high production of cellulases, is an industrial strain with many patents pending for approval.
In this context, it is important to study the secretome of fungal strains, since the genetic improvement process of these organisms are unknown. In this paper, we strived to elucidate the mechanism involved in S1M29-enhanced biomass decomposition phenotype on this question through the secretome level, presenting the first study of wild-type 2HH and mutant S1M29 secretomes of P. echinulatum in submerged cultivation using sugar cane bagasse pretreated by steam explosion (SCB), using cellulose, glucose, and glycerol as carbon sources. To supplement this study, an enzymatic analysis using a set of substrates was conducted in order to further validate and describe the enzymatic repertoire available to degrade lignocellulosic biomass by P. echinulatum.
Results
General features of the P. echinulatum genome strains
The fungus P. echinulatum has a genome with a similar size to other Penicillium species described in the literature [23, 24]. The genomic sequencing showed that both strains of P. echinulatum, wild-type 2HH and mutant S1M29, have a genome of the same size (Table 1). There were no significant changes in genome size with respect to the deletions in regions encoding proteins. Thus, changes in enzyme production between the two strains is probably explained at the transcription level and is not due to large scale genomic rearrangements.
Table 1.
General features of Penicillium echinulatum genome
| Category | Wild-type 2HH | Mutant S1M29 |
|---|---|---|
| Genome statistic | ||
| Genome size (MB) | 29.82 | 29.83 |
| Number of bases | 29.821.618 | 29.827.032 |
| Number of ambiguous bases (N) | 92.041 | 82.971 |
| Number of contigs | 1018 | 1124 |
| n50 | 147,886 | 137,718 |
| n90 | 32,016 | 32,016 |
| GC % | 50.5 | 50.5 |
| Number of predicted genes | 8504 | 8552 |
| Average gene size | 1686 | 1684 |
| Median gene size | 1441 | 1442 |
| Average protein size (AA) | 505 | 504 |
| Number of predicted exons | 25.098 | 25.494 |
| Number of predicted introns | 16,638 | 16,997 |
| Average number of exons per gene | 3 | 3 |
| Average number of introns per gene | 2 | 2 |
| Average exon size | 514 | 507 |
| Average intron size | 87 | 87 |
| Median intron size | 73 | 23 |
| Intergenic size | 15,484,048 | 15,422,613 |
| Intragenic size | 14,337,570 | 14,404,419 |
| Number of introns in gene with largest splicing capacity | 23 | 23 |
| Number of putative secreted proteins | 649 | 649 |
| Number of predicted proteins with e value of e 5 or less | 8157 | 8190 |
| Genome read | ||
| Number of reads | 679.730.992 | 545.354.384 |
| Number of read pairs | 339.865.496 | 272.677.192 |
| Number of bases | 33.986.549.600 | 27.264.719.200 |
Electrophoretic profile of P. echinulatum
The analysis of the gels (1-D PAGE) of total proteins secreted by both strains of P. echinulatum grown on SCB, cellulose, glucose, and glycerol, after 96 h of culture, shows the diversity of molecular masses of proteins (Fig. 1). It is possible to see differences in the level of some proteins that appear in greater quantity in the mutant strain S1M29 (Fig. 1b—red squares) than in the wild-type 2HH when grown on cellulose or SCB, which is evidenced by proteomic data. Some of the proteins are correlated to cellulases and hemicellulases found in the range of 50–75 kDa (Tables 2, 3). On the other hand, protein bands of higher molecular weight (~120 kDa) are displayed only in the wild type (Fig. 1a—red squares).
Fig. 1.
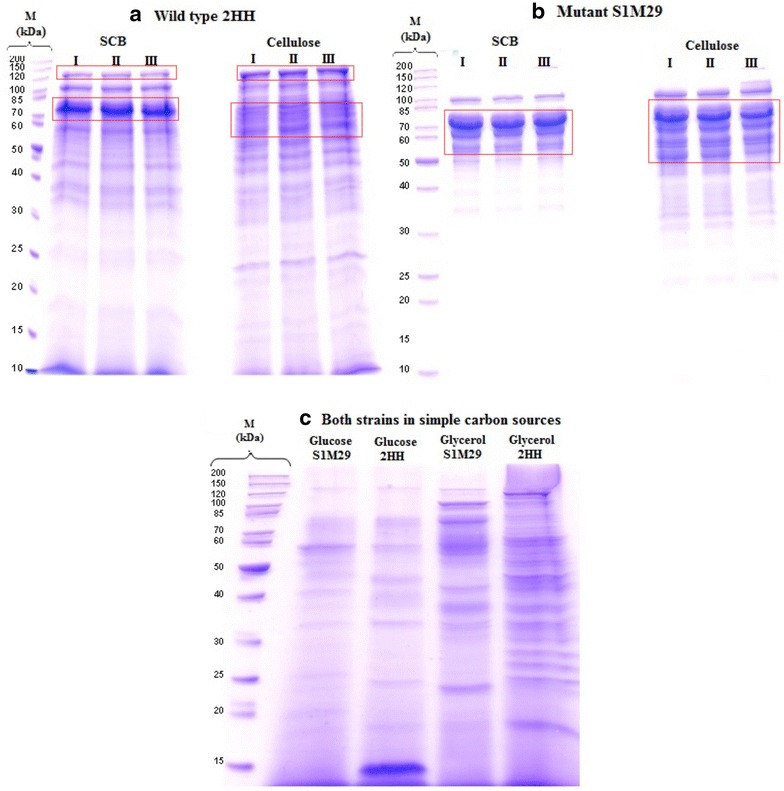
Electrophoretic profile of Penicillium echinulatum grown on SCB and cellulose at 96 h of cultivation. On the left a, 1-D PAGE of wild-type 2HH and on the right b 1-D PAGE of the mutant strain S1M29. I, II and III refer to triplicate. c Replicates for glucose and glycerol were performed, and profiles of wild-type 2HH and mutant S1M29 are shown intercalary in the gel above. M molecular marker, kDa molecular weight. Red squares indicate the differences in the level of some proteins that appear in greater quantity considering both strains and complex carbon sources
Table 2.
Identified cellulose-degradation/modifying enzymes and spectrum counts in different carbon sources by wild-type 2HH and mutant S1M29 of Penicillium echinulatum at 96 h during submerged cultivation
| Accession number | Identified proteins | CAZyme | Organism | MW (kDa) | Secretionb | Spectral counta | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2HH | S1M29 | ||||||||||||
| Glu | Gly | SCB | Cel | Glu | Gly | SCB | Cel | ||||||
| g2773 | Cellobiohydrolase I | GH7 (CBM1) | Penicillium decumbens | 56 | Y | 0 | 0 | 81 | 107 | 0 | 2 | 202 | 80 |
| g8068 | Cellobiohydrolase II | GH6 (CBM1) | Penicillium decumbens | 49 | Y | 0 | 0 | 16 | 4 | 0 | 0 | 46 | 62 |
| g8473 | Endoglucanase 1 | GH5 (CBM1) | Penicillium echinulatum | 44 | Y | 0 | 0 | 19 | 27 | 0 | 1 | 75 | 59 |
| g5809 | Endoglucanase Cel5C | GH5 (CBM1, CBM46) | Penicillium decumbens | 69 | Y | 0 | 1 | 28 | 30 | 0 | 1 | 56 | 58 |
| g2788 | Endoglucanase I | GH7 (CBM1) | Penicillium decumbens | 50 | Y | 0 | 0 | 5 | 4 | 0 | 0 | 14 | 10 |
| g2291 | Endoglucanase II | GH5 (CBM1) | Penicillium decumbens | 44 | Y | 0 | 0 | 4 | 4 | 0 | 0 | 13 | 7 |
| g2659 | Endo-β-1,4-glucanase | GH5 (CBM1) | Penicillium oxalicum | 52 | Y | 27 | 0 | 0 | 3 | 49 | 1 | 0 | 6 |
| g3996 | Endo-β-1,4-glucanase | GH12 | Penicillium oxalicum | 26 | Y | 0 | 0 | 9 | 10 | 0 | 0 | 12 | 8 |
| g7274 | Endo-β-1,4-glucanase | GH5 | Penicillium oxalicum | 55 | N | 0 | 0 | 5 | 1 | 0 | 0 | 0 | 0 |
| g6766 | β-glucosidase I | GH3 | Penicillium decumbens | 93 | Y | 0 | 7 | 2 | 1 | 1 | 17 | 2 | 0 |
| g8479 | Related to β-glucosidase | GH132 | Colletotrichum gloeosporioides | 46 | Y | 0 | 2 | 0 | 22 | 4 | 0 | 0 | 0 |
| g3303 | Cellulose monooxygenase Cel61A | AA9 | Penicillium oxalicum | 26 | Y | 0 | 0 | 3 | 0 | 0 | 0 | 20 | 11 |
| g8342 | Cellulose monooxygenase | AA9 | Penicillium oxalicum | 45 | Y | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 |
| g5167 | Swollenin | CBM1, CBM63 | Penicillium oxalicum | 52 | Y | 0 | 0 | 1 | 4 | 0 | 0 | 9 | 6 |
| g1074 | GMC oxidoreductase | AA3 | Neosartorya fischeri | 70 | Y | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| g3947 | GMC oxidoreductase | AA3 | Penicillium roqueforti | 67 | N | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 0 |
aSecretomic analysis based on spectral counting. A quantitative analysis was conducted for samples grown on cellulose or SCB (mean of triplicates), while a semiquantitative analysis for samples grown on glucose or glycerol (one replicate) was performed. The complete protein reports are given in Additional file 3
bThe secretion of each protein was verified by the softwares SignalP, SecretomeP, and YLoc. When at least two of three softwares give a positive result, the protein was considered secreted
Table 3.
Identified hemicellulose-degradation enzymes and spectrum count in different carbon sources by wild-type 2HH and mutant S1M29 of Penicillium echinulatum at 96 h during submerged cultivation
| Accession number | Identified proteins | CAZyme | Organism | MW (kDa) | Secretionb | Spectrum counta | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2HH | S1M29 | ||||||||||||
| Glu | Gly | SCB | Cel | Glu | Gly | SCB | Cel | ||||||
| g2645 | Xylanase | GH10 (CBM1) | Penicillium decumbens | 44 | Y | 0 | 0 | 1 | 10 | 0 | 0 | 21 | 27 |
| g1025 | Endo-β-1,4-xylanase | GH10 | Penicillium oxalicum | 40 | Y | 0 | 0 | 12 | 21 | 0 | 0 | 0 | 1 |
| g3831 | Endo-β-1,4-xylanase | GH30 | Penicillium oxalicum | 53 | Y | 0 | 0 | 5 | 11 | 0 | 0 | 0 | 2 |
| g5166 | Endo-β-1,4-xylanase | GH11 (CBM1) | Penicillium oxalicum | 31 | Y | 0 | 0 | 0 | 2 | 1 | 0 | 2 | 5 |
| g3240 | β-xylosidase | GH43 | Penicillium oxalicum | 64 | Y | 0 | 0 | 0 | 8 | 0 | 0 | 0 | 0 |
| g1531 | β-xylosidase | GH43 | Penicillium oxalicum | 38 | Y | 0 | 0 | 8 | 0 | 0 | 0 | 0 | 0 |
| g2200 | β-xylosidase | GH3 | Penicillium oxalicum | 85 | Y | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 15 |
| g5735 | α-mannosidase | GH47 | Penicillium oxalicum | 56 | Y | 0 | 14 | 7 | 3 | 0 | 16 | 0 | 3 |
| g395 | Exo-α-1,6-mannosidase | GH125 | Sphaerulina musiva | 56 | N | 0 | 12 | 0 | 2 | 0 | 7 | 0 | 0 |
| g5829 | β-1,4-mannanase | GH5 (CBM1) | Penicillium oxalicum | 47 | Y | 0 | 0 | 7 | 8 | 0 | 2 | 0 | 12 |
| g8513 | Sorbitol/xylulose reductase | GH99 | Aspergillus clavatus | 28 | N | 0 | 13 | 6 | 22 | 0 | 2 | 1 | 1 |
| g315 | Acetyl xylan esterase | CE1 (CBM1) | Penicillium oxalicum | 41 | Y | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| g2707 | Acetyl xylan esterase | CE5 | Penicillium oxalicum | 24 | Y | 0 | 0 | 0 | 5 | 0 | 0 | 1 | 2 |
aSecretomic analysis based on spectral counting. A quantitative analysis was conducted for samples grown on cellulose or SCB (mean of triplicates), while a semiquantitative analysis for samples grown on glucose or glycerol (one replicate) was performed. The complete protein reports are given in Additional file 3
bThe secretion of each protein was verified by the softwares SignalP, SecretomeP and YLoc. When at least two of three softwares give a positive result the protein was considered secreted
The analysis of electrophoretic profiles of both strains of P. echinulatum grown on glucose or glycerol (Fig. 1c) also reveals a significant number of bands with different molecular weights. However, the bands do not appear so intensely when compared with SCB or medium formulated with cellulose, which clearly indicates that the secretion of proteins changes according to the carbon source.
Figure 2 shows the protein concentration. The amount of proteins present in the media with cellulose or SCB after 48 h of cultivation is statistically higher for the mutant S1M29 compared to the wild-type 2HH. It is worth noting that the SCB medium presents the largest amount of protein. The amount of proteins secreted by the two strains in the media elaborated with glycerol or glucose slightly varied during the time course and corresponds to a smaller amount compared with cellulose and SCB. Still, the quantity of protein in the media formulated with glucose or glycerol did not differ statistically, even at 120 h of cultivation.
Fig. 2.
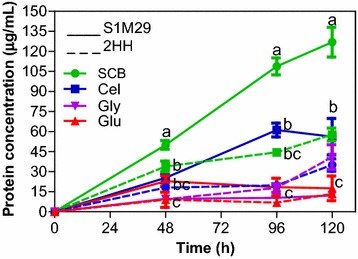
Protein concentration. Levels of protein concentration of Penicillium echinulatum wild-type 2HH and mutant S1M29 grown in four different carbon sources (SCB, cellulose, glucose, and glycerol) for 120 h. Protein concentration was performed according to the method of Bradford and is expressed in µg/mL
Profiling of the P. echinulatum secretome for lignocellulose degradation
The set of proteins found in secretome of both strains have a molecular weight in the range between 11 and 123 kDa. In the secretomic analyses of wild-type 2HH and mutant S1M29 from P. echinulatum, 165 proteins were found. In total, 36 proteins are found exclusively from the wild type, 18 proteins exclusively from the mutant, while 111 are commonly found in both strains (Fig. 3). Using bioinformatics tools to predict secreted proteins, it has been found that 117 proteins (71 %) of the identified proteins are predicted to contain signal peptide and/or are secreted by nonclassical routes.
Fig. 3.
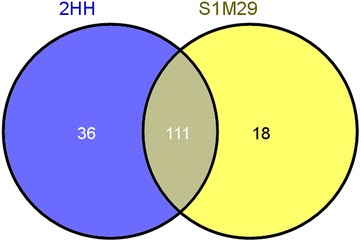
Venn diagram of secretome from Penicillium echinulatum strains. Venn diagram shows the shared secretome (overlap) and specific secretome for wild-type 2HH (blue) and mutant S1M29 (yellow)
The main objective of this work was to find enzymes involved in the degradation of lignocellulosic biomass. Out of the 147 proteins found in the wild type, 63 proteins (~43 %) belonged to families described in the CAZy database [25]. Similarly, 57 proteins (~44 %) from 129 proteins found in the mutant belonged to families in the CAZy database (Fig. 4).
Fig. 4.
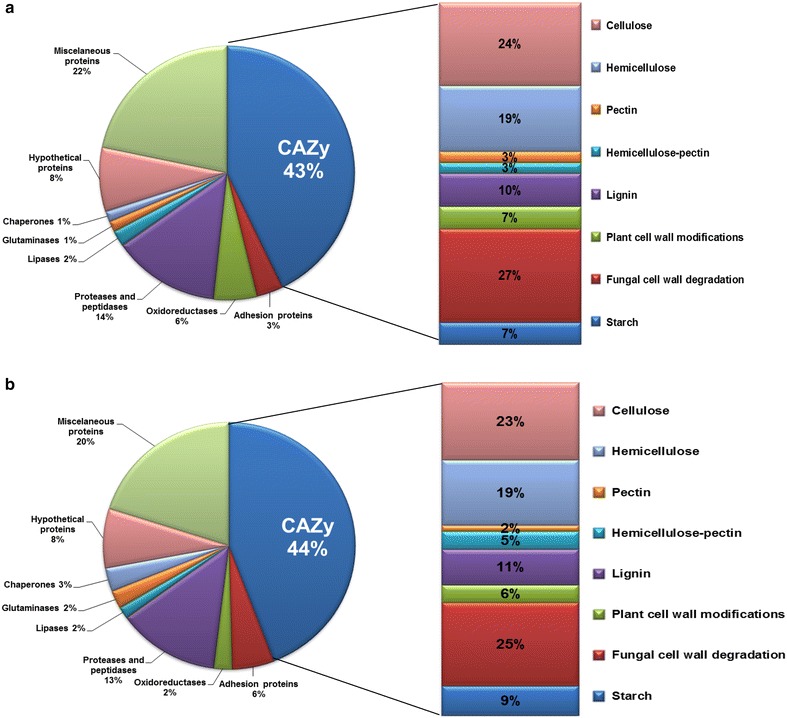
Distribution of proteins identified in secretome of Penicillium echinulatum strains. The proteins were classified according to the CAZy database. a The CAZy families identified in the wild type. b The CAZy families identified in the mutant
Among the proteins found in the secretomes of the wild-type and mutant, cellulases, hemicellulases, and fungal cell wall degradation enzymes were the majority enzymes, accounting for over 50 % of the identified proteins (Fig. 4). However, cellulases correspond to enzymes that were more expressed, according to the spectral counting analysis.
Regarding the secretome profile found in wild-type 2HH of P. echinulatum the largest number of proteins were found in the medium formulated with cellulose (102 proteins, 15 exclusive), followed by glycerol (85 proteins, 14 exclusive) SCB (80 proteins, 10 exclusive) and glucose (41 proteins, 11 exclusive). From the 147 proteins found in the secretome of wild type, 13 were found in all carbon sources evaluated in this work (Fig. 5a).
Fig. 5.
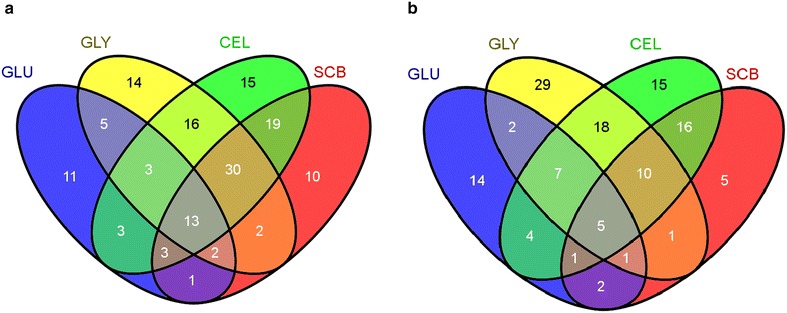
Venn diagram of secretomes from Penicillium echinulatum strains under each condition. a Distribution of 147 proteins identified in the secretome of wild-type 2HH, and b distribution of 129 proteins identified in the secretome of mutant S1M29, exclusive in each carbon source or shared between them. GLU glucose, GLY glycerol, CEL cellulose, and SCB sugar cane bagasse. The venn diagram was prepared by means of the tool, Venny (http://bioinfogp.cnb.csic.es/tools/venny/index.html)
In secretome profile of mutant strain S1M29 of P. echinulatum, the larger number of proteins was found again in the medium formulated with cellulose (76 proteins, 15 exclusive). Glycerol appears in the sequence with 73 proteins identified, and it has the greatest number of exclusive proteins (29). The medium formulated with SCB comes in third place (41 proteins, 5 exclusives), followed by glucose (36 proteins, 14 exclusives). Of the 129 proteins found in the secretome of the mutant strain, five were found in all four different culture media (Fig. 5b).
Among 36 proteins found only in the wild-type secretome, some have glycosyl hydrolase (GH) function, such as exo-β-1,3-glucanase (GH5), 1,3-β-glucanase (GH17), found exclusively in glucose, and chitinase (GH18), found only in glycerol (Table 10). In addition, endo-β-1,4-glucanase (GH5) (Table 2) and β-xylosidase (GH43) were found in cellulose or SCB (Table 3), and exo-α-l-1,5-arabinanase (GH93) was present only in cellulose (Table 4). Carbohydrate acetyl esterase (CE16) (Table 8) and cellulose monooxygenase (AA9) were also detected, and the latter secreted only in SCB (Table 2). As can be viewed in Additional file 1, some hypothetical and adhesion proteins, proteases, lipases, and other proteins were also found in the secretome of P. echinulatum wild type.
Table 10.
Identified fungal cell wall degradation/modifying enzymes and spectrum counts in different carbon sources by wild-type 2HH and mutant S1M29 of Penicillium echinulatum at 96 h during submerged cultivation
| Accession number | Identified proteins | CAZyme | Organism | MW (kDa) | Secretionb | Spectrum counta | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2HH | S1M29 | ||||||||||||
| Glu | Gly | SCB | Cel | Glu | Gly | SCB | Cel | ||||||
| g7097 | Chitin glucanosyltransferase | GH16 | Penicillium oxalicum | 39 | Y | 41 | 11 | 12 | 13 | 32 | 0 | 9 | 10 |
| g1080 | Chitinase1 | GH18 | Penicillium oxalicum | 49 | Y | 1 | 25 | 0 | 1 | 0 | 105 | 0 | 1 |
| g8496 | Chitinase 1, class 5 | CBM50 | Grosmannia clavigera | 37 | Y | 16 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| g1258 | Exo-β-1,3-glucanase | GH17 | Penicillium oxalicum | 47 | Y | 72 | 5 | 4 | 4 | 15 | 10 | 1 | 3 |
| g4052 | β-1,3-glucanosyltransferase | GH17 | Penicillium oxalicum | 33 | Y | 9 | 8 | 3 | 15 | 4 | 9 | 1 | 4 |
| g284 | β-N-acetylhexosaminidase | GH3 | Penicillium oxalicum | 38 | Y | 29 | 19 | 7 | 3 | 0 | 10 | 0 | 0 |
| g244 | β-1,3-glucanosyltransglycosylase | GH72 | Penicillium oxalicum | 49 | Y | 0 | 9 | 7 | 6 | 0 | 11 | 1 | 6 |
| g3495 | Chitosanase | GH75 | Penicillium oxalicum | 37 | Y | 30 | 12 | 3 | 3 | 1 | 2 | 0 | 2 |
| g5309 | Exo-β-1,3-glucanase | GH5 | Penicillium oxalicum | 45 | Y | 0 | 3 | 5 | 8 | 0 | 0 | 0 | 0 |
| g6946 | Chitin glucanosyltransferase | GH16 (CBM 18) | Penicillium oxalicum | 47 | Y | 5 | 6 | 5 | 2 | 0 | 3 | 1 | 2 |
| g980 | β-1,6-N-acetyl glucosaminidase | GH20 | Penicillium oxalicum | 67 | Y | 0 | 4 | 4 | 2 | 0 | 4 | 0 | 1 |
| g2799 | β-1,3-glucanosyl transglycosylase | GH72 | Penicillium oxalicum | 53 | Y | 0 | 0 | 2 | 1 | 0 | 0 | 0 | 1 |
| g1191 | β-1,3-glucanosyl transglycosylase | GH72 (CBM 43) | Penicillium oxalicum | 57 | Y | 2 | 0 | 1 | 0 | 0 | 0 | 0 | 1 |
| g3950 | Exo-β-1,3-glucanase | GH5 | Penicillium oxalicum | 46 | Y | 0 | 1 | 0 | 0 | 0 | 3 | 0 | 0 |
| g6368 | Chitinase | GH18 | Penicillium oxalicum | 47 | Y | 0 | 3 | 0 | 3 | 0 | 0 | 0 | 0 |
| g4641 | β-1,3-glucanase | GH17 | Penicillium oxalicum | 68 | N | 3 | 3 | 0 | 0 | 0 | 0 | 0 | 0 |
| g5953 | β-1,6-glucanase | GH30 | Penicillium oxalicum | 52 | Y | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| g8051 | α-1,3-glucanase | GH71 (CBM24) | Penicillium oxalicum | 114 | Y | 7 | 0 | 0 | 2 | 0 | 0 | 0 | 5 |
aSecretomic analysis based on spectral counting. A quantitative analysis was conducted for samples grown on cellulose or SCB (mean of triplicates), while a semiquantitative analysis for samples grown on glucose or glycerol (one replicate) was performed. The complete protein reports are given in Additional file 3
bThe secretion of each protein was verified by the softwares SignalP, SecretomeP and YLoc. When at least two of three softwares give a positive result the protein was considered secreted
Table 4.
Identified pectin-degradation enzymes and spectrum counts in different carbon sources by wild-type 2HH and mutant S1M29 of Penicillium echinulatum at 96 h during submerged cultivation
| Accession number | Identified proteins | CAZyme | Organism | MW (kDa) | Secretionb | Spectrum counta | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2HH | S1M29 | ||||||||||||
| Glu | Gly | SCB | Cel | Glu | Gly | SCB | Cel | ||||||
| g4880 | Pectin lyase | PL1 | Penicillium oxalicum | 40 | Y | 0 | 0 | 0 | 1 | 12 | 0 | 0 | 0 |
| g3609 | Exo-α-l-1,5-arabinanase | GH93 | Penicillium oxalicum | 44 | Y | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 |
aSecretomic analysis based on spectral counting. A quantitative analysis was conducted for samples grown on cellulose or SCB (mean of triplicates), while a semiquantitative analysis for samples grown on glucose or glycerol (one replicate) was performed. The complete protein reports are given in Additional file 3
bThe secretion of each protein was verified by the softwares SignalP, SecretomeP and YLoc. When at least two of three softwares give a positive result the protein was considered secreted
Table 8.
Identified enzymes involved in plant cell wall modifications and spectrum counts in different carbon sources by wild-type 2HH and mutant S1M29 of Penicillium echinulatum at 96 h during submerged cultivation
| Accession number | Identified proteins | CAZyme | Organism | MW (kDa) | Secretionb | Spectrum counta | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2HH | S1M29 | ||||||||||||
| Glu | Gly | SCB | Cel | Glu | Gly | SCB | Cel | ||||||
| g1590 | Carboxylesterase, type B | CE10 | Penicillium roqueforti | 62 | Y | 9 | 1 | 0 | 2 | 0 | 1 | 0 | 5 |
| g7004 | Carbohydrate acetyl esterase | CE16 | Penicillium oxalicum | 33 | Y | 0 | 0 | 0 | 5 | 0 | 0 | 0 | 0 |
| g5807 | Homoserine acetyltransferase | CE1 | Talaromyces marneffei | 37 | N | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 |
| g1708 | Carboxylesterase, type B | CE10 | Penicillium roqueforti | 61 | Y | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 |
aSecretomic analysis based on spectral counting. A quantitative analysis was conducted for samples grown on cellulose or SCB (mean of triplicates), while a semiquantitative analysis for samples grown on glucose or glycerol (one replicate) was performed. The complete protein reports are given in Additional file 3
bThe secretion of each protein was verified by the softwares SignalP, SecretomeP and YLoc. When at least two of three softwares give a positive result the protein was considered secreted
Concerning the mutant strain secretome, 18 exclusive proteins were identified, among these also enzymes with GH function, such as α-l-arabinofuranosidase (GH62), found only in SCB and cellulose (Table 5) and β-1,6-glucanase (GH30) (Table 10). Esterases, such as acetyl xylan esterase (CE1) (Table 3) and enzymes related to the depolymerization of lignin (Table 6) were identified. Additional data also show the other exclusive proteins found refer to ribosomal, hypothetical, and adhesion proteins, lipases, and proteases/peptidases (Additional file 1).
Table 5.
Identified hemicellulose/pectin-degradation enzymes and spectrum counts in different carbon sources by wild-type 2HH and mutant S1M29 of Penicillium echinulatum at 96 h during submerged cultivation
| Accession number | Identified proteins | CAZyme | Organism | MW (kDa) | Secretionb | Spectrum counta | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2HH | S1M29 | ||||||||||||
| Glu | Gly | SCB | Cel | Glu | Gly | SCB | Cel | ||||||
| g1097 | α-l-arabinofuranosidase | GH62 (CBM1) | Penicillium oxalicum | 41 | Y | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 3 |
| g7308 | α-l-arabinofuranosidase | GH43 | Penicillium oxalicum | 34 | Y | 0 | 2 | 2 | 0 | 0 | 0 | 1 | 0 |
| g1098 | α-l-arabinofuranosidase | GH43 (CBM1) | Penicillium oxalicum | 62 | Y | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 |
aSecretomic analysis based on spectral counting. A quantitative analysis was conducted for samples grown on cellulose or SCB (mean of triplicates), while a semiquantitative analysis for samples grown on glucose or glycerol (one replicate) was performed. The complete protein reports are given in Additional file 3
bThe secretion of each protein was verified by the softwares SignalP, SecretomeP and YLoc. When at least two of three softwares give a positive result the protein was considered secreted
Table 6.
Identified lignin-degradation enzymes and spectrum counts in different carbon sources by wild-type 2HH and mutant S1M29 of Penicillium echinulatum at 96 h during submerged cultivation
| Accession number | Identified proteins | CAZyme | Organism | MW (kDa) | Secretionb | Spectrum counta | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2HH | S1M29 | ||||||||||||
| Glu | Gly | SCB | Cel | Glu | Gly | SCB | Cel | ||||||
| g1557 | Isoamyl alcohol oxidase | AA7 | Talaromyces stipitatus | 66 | Y | 11 | 2 | 6 | 0 | 35 | 31 | 3 | 4 |
| g1100 | Isoamyl alcohol oxidase | AA7 | Arthroderma benhamiae | 58 | Y | 2 | 0 | 0 | 2 | 0 | 0 | 0 | 1 |
| g5086 | Isoamyl alcohol oxidase | AA7 | Aspergillus flavus | 67 | Y | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 2 |
| g6365 | Related to Isoamyl alcohol oxidase | AA7 | Fusarium fujikuro | 69 | Y | 0 | 1 | 3 | 3 | 18 | 11 | 0 | 3 |
| g7414 | FAD dependent oxidoreductase | AA7 | Aspergillus clavatus | 54 | Y | 0 | 2 | 2 | 1 | 0 | 10 | 0 | 0 |
| g5776 | 6-hydroxy-d-nicotine oxidase | AA7 | Aspergillus kawachii | 54 | Y | 3 | 4 | 2 | 2 | 5 | 6 | 0 | 2 |
aSecretomic analysis based on spectral counting. A quantitative analysis was conducted for samples grown on cellulose or SCB (mean of triplicates), while a semiquantitative analysis for samples grown on glucose or glycerol (one replicate) was performed. The complete protein reports are given in Additional file 3
bThe secretion of each protein was verified by the softwares SignalP, SecretomeP and YLoc. When at least two of three softwares give a positive result the protein was considered secreted
Most of the proteins identified as exclusive in the secretome of the wild-type 2HH and the mutant strain S1M29 have a low number of spectral counting, which generates some analysis bias due to differences in the length of protein, the molecular weight, and ionization efficiency [26].
Among the proteins belonging to the CAZy database in the wild-type 2HH, 45 proteins were found grouped in 24 different families of GHs (Fig. 6a), with the largest number of spectra belonging to cellobiohydrolases I and II (GH7 and GH6), endoglucanases (GH5 and GH7) and xylanase (GH10). These enzymes were basically identified only in the media formulated with SCB or cellulose. In the secretome profile of the mutant S1M29, 57 proteins belong to CAZy families, and once again, GHs were the most abundant (41 proteins grouped into 25 families), as can be seen in Fig. 6b.
Fig. 6.
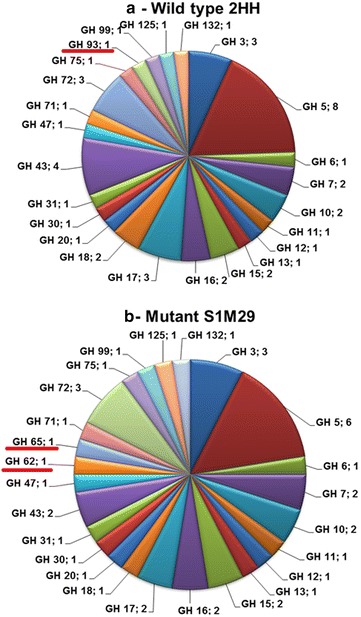
Distribution of GHs families in Penicillium echinulatum secretome. The proteins were classified according to CAZy database. a GHs identified in wild-type 2HH, b GHs identified in mutant S1M29, highlighting the presence of the families GH93 in 2HH (a) and GH62/GH65 in S1M29 (b). They are exclusive to each strain, although the number of spectra for this protein was low
Considering the CAZymes found in mutant and wild type, there were a greater number of spectra identified in the mutant, as can be seen especially in Tables 2 and 3. This happened with cellobiohydrolases I and II (GH7 and GH6), endoglucanases 1 and Cel5C, endoglucanase (GH5), and xylanase (GH10). In addition, these enzymes have been found with a higher number of spectra in SCB and cellulose. β-glucosidase I (GH3) was identified with a greater number of spectra in glycerol medium, with this number also larger in the mutant strain (Table 2). However, another protein related to β-glucosidase was found in the wild type with a considerable amount of spectra in cellulose.
The number of spectra for exo-β-1,3-glucanase (GH17) in both strains cultivated in glucose is relatively larger than has been identified in other carbon sources, especially in the wild type (Table 10), and is related to the degradation of the fungal cell wall. Other GHs found in the wild type with larger spectra in glucose and glycerol than in SCB and cellulose, which are also related to the degradation of the fungal cell wall (Table 10), were chitin glucanosyltransferase (GH16), chitosanase (GH75) and β-N-acetylhexosaminidase (GH3). As shown in additional file the presence of lysophospholipase phospholipase B (Additional file 1) with a greater number of spectra in the wild type, especially in medium prepared with glucose or glycerol, is a further indication of the arsenal of enzymes produced by the wild type for the degradation of fungal cell wall.
Other enzymes with GH function found in both strains with a reduced number of spectra were putative endoxylanase (GH30), α-mannosidase (GH47), with the largest number of spectra in glycerol; β-1,4-mannanase (GH5), found in SCB and cellulose; β-xylosidase (GH3), found only in the media formulated with cellulose, in both strains (Table 3). Distinct endoglucanases (GH5 and GH12) were expressed differently in the culture media—GH5 with more spectra in glucose, but GH12 in SCB and cellulose (Table 2).
The presence of enzymes related to starch degradation were also found in both strains, although with a lower number of spectra, as the case of α-amylase (GH13), α-glucosidase (GH31) and two glucoamylases (GH15), the latter being identified only when glucose or glycerol was used as a carbon source (Table 7).
Table 7.
Identified starch-degradation enzymes and spectrum counts in different carbon sources by wild-type 2HH and mutant S1M29 of Penicillium echinulatum at 96 h during submerged cultivation
| Accession number | Identified proteins | CAZyme | Organism | MW (kDa) | Secretionb | Spectrum counta | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2HH | S1M29 | ||||||||||||
| Glu | Gly | SCB | Cel | Glu | Gly | SCB | Cel | ||||||
| g4201 | α-amylase | GH13 | Penicillium oxalicum | 74 | Y | 0 | 2 | 2 | 1 | 0 | 1 | 0 | 6 |
| g5478 | Glucoamylase Amy15A | GH15 (CBM20) | Penicillium oxalicum | 66 | Y | 1 | 1 | 0 | 0 | 9 | 10 | 0 | 0 |
| g36 | Glucoamylase | GH15 (CBM20) | Penicillium oxalicum | 68 | Y | 1 | 0 | 0 | 0 | 14 | 0 | 0 | 0 |
| g7292 | α-glucosidase | GH31 | Penicillium oxalicum | 108 | Y | 0 | 7 | 0 | 0 | 0 | 1 | 0 | 0 |
| g1651 | α-trehalase | GH65 | Penicillium oxalicum | 119 | Y | 0 | 0 | 0 | 0 | 0 | 23 | 0 | 0 |
aSecretomic analysis based on spectral counting. A quantitative analysis was conducted for samples grown on cellulose or SCB (mean of triplicates), while a semiquantitative analysis for samples grown on glucose or glycerol (one replicate) was performed. The complete protein reports are given in Additional file 3
bThe secretion of each protein was verified by the softwares SignalP, SecretomeP and YLoc. When at least two of three softwares give a positive result the protein was considered secreted
Six carbohydrate esterases were identified in both strains, with a reduced number of spectra, grouped in four different families, including carboxylesterases (CE10), homoserine acetyltransferase (CE1), carbohydrate acetylesterase (CE16) (Table 8) and acetyl xylan esterase (CE1 and CE5) (Table 3).
The same families of auxiliary redox enzymes (AA9), which act together with CAZymes, were found in both strains. Cellulose monooxygenase Cel 61A (AA9/g3303) was one of the most produced enzymes when we consider S1M29 in the medium prepared with SCB and cellulose. For 2HH, the presence of this AA9 is observed only in the medium with SCB, although the presence of another AA9 (putative) is possible in the SCB medium.
The presence of only one polysaccharide lyase, the pectin lyase (PL1) and one exo-arabinanase was verified (Table 4). Arabinofuranosidase was also found in less quantity (Table 4).
The secretome analysis has shown that approximately 30 % of the identified CAZymes have carbohydrate-binding module (CBMs). Eight different CBM families were identified, with the CBM1 as the most abundant. In the case of endoglucanase Cel5C (GH5/g5809), two CBMs were found attached to the same CAZyme: CBM46 and CBM1, and also two CBMs for swollenins: CBM63 and CBM1 (Table 2).
Additional data features the presence of xylose reductase in both strains grown in SCB and cellulose, and they may indicate the potential of these broths for enzymatic production of xylitol and perhaps the potential use as a facilitator for xylose-to-ethanol conversion by other microorganisms (refer to Additional file 1). Xylulose reductase was also found (Table 3), but this enzyme does not participate in the conversion of xylose to xylulose for ethanol production [27].
Some enzymes involved in the degradation of lignin were detected. In the secretome of mutant strain, isoamyl alcohol oxidase (AA7) was found in a large number of spectra in glucose and glycerol (Table 6). GMC oxidoreductase (AA3), an enzyme which acts both in cellulose and lignin—possibly related to cellobiose dehydrogenase [28, 29]—was found in the medium with glycerol (Table 2). Manganese superoxide dismutase and glutathione S-transferase were also identified, as shown in Additional file 1.
In Table 9, the main differences between the secretomes of the wild type and mutant are represented, regarding the identification of key proteins in the lignocellulosic biomass degradation. For most enzymes analyzed, there are statistical differences (t test p < 0.05) between the number of spectra found in SCB or cellulose, when comparing the spectra in wild type with the mutant. That difference is clearly shown, i.e., the expression of these enzymes in SCB by the mutant. The expression of cellobiohydrolases, endoglucanases, and xylanase reveal the mutant S1M29 greater potential for biomass hydrolysis.
Table 9.
Key proteins involved in the lignocellulosic biomass degradation and its significant difference between the spectra produced in SCB and cellulose
| Accession number | Identified proteins | CAZyme | Spectrum count |
T testa
(p < 0.05) |
T testa (p < 0.05) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2HH | S1M29 | |||||||||||
| Glu | Gly | SCB | Cel | Glu | Gly | SCB | Cel | SCB | CEL | |||
| g2773 | Cellobiohydrolase I | GH7 (CBM1) | 0 | 0 | 81 | 107 | 0 | 2 | 202 | 80 | 0.019 | 0.29 |
| g8068 | Cellobiohydrolase II | GH6 (CBM1) | 0 | 0 | 16 | 4 | 0 | 0 | 46 | 62 | 0.027 | 0.0045 |
| g5809 | Endoglucanase Cel5C | GH5 (CBM1, CBM46) | 0 | 1 | 28 | 30 | 0 | 1 | 56 | 58 | 0.0018 | 0.0032 |
| g2788 | Endoglucanase I | GH7 (CBM1) | 0 | 0 | 5 | 4 | 0 | 0 | 14 | 10 | 0.0011 | 0.10 |
| g2291 | Endoglucanase II | GH5 (CBM1) | 0 | 0 | 4 | 4 | 0 | 0 | 13 | 7 | 0.0073 | 0.0072 |
| g3996 | Endo-β-1,4-glucanase | GH12 | 0 | 0 | 9 | 10 | 0 | 0 | 12 | 8 | 0.045 | 0.064 |
| g3303 | Cellulose monooxygenase Cel61A | AA9 | 0 | 0 | 3 | 0 | 0 | 0 | 20 | 11 | 0.0013 | 0.00060 |
| g5167 | Swollenin | CBM1, CBM63 | 0 | 0 | 1 | 4 | 0 | 0 | 9 | 6 | 0.00030 | 0.23 |
| g2645 | Xylanase | GH10 (CBM1) | 0 | 0 | 1 | 10 | 0 | 0 | 21 | 27 | 0.00010 | 0.012 |
| g1025 | Endo-β-1,4-xylanase | GH10 | 0 | 0 | 12 | 21 | 0 | 0 | 0 | 1 | 0.0056 | 0.038 |
| g1531 | β-xylosidase | GH43 | 0 | 0 | 8 | 0 | 0 | 0 | 0 | 0 | 0.0020 | 1.00 |
| g5829 | β-1,4-mannanase | GH5 (CBM1) | 0 | 0 | 7 | 8 | 0 | 2 | 0 | 12 | 0.0046 | 0.42 |
| g5309 | Exo-β-1,3-glucanase | GH5 | 0 | 3 | 5 | 8 | 0 | 0 | 0 | 0 | 0.00010 | 0.010 |
a Statistical approach (T test, p < 0.05) was performed between the strains for SCB and cellulose media. Proteins with statistically significant differences are highlighted in italics
When comparing the total number of spectra found in SCB or cellulose between strains, it is found that 77 % of the spectra found in SCB in the mutant strain correspond to enzymes relating to cellulose and hemicellulose degradation, whereas in the medium with cellulose, 58 % of these spectra are found. In contrast, when the same comparison is made with the wild type, there were a greater number of spectra (~38 %) in cellulose compared to SCB (~31 %). It is suggested that SCB has become a better inducer of cellulolytic and hemicellulolytic enzymes, at least for the mutant strain. Considering either glucose or glycerol as carbon source, the amounts of cellulases and hemicellulases produced corresponded to less than 10 % of the total spectra for both strains.
Taking another approach, Additional file 2 demonstrates that, considering the total number of spectra, when we look at the ten most expressed proteins in each carbon source for each strain, it was again evident that the secretome of mutant strain in the media with SCB or cellulose is more geared for the production of cellulolytic and hemicellulolytic enzymes than the wild type; more than 70 % of these proteins correspond to cellulases and hemicellulases. Meanwhile, the secretome of media containing glucose or glycerol have a protein complex consisting of a majority of proteins related to degradation of fungal cell wall, adhesion proteins, and proteases; only two cellulases (g2659/g6766) and one hemicellulase (g5735) were identified among the ten most expressed proteins in these conditions and only in the mutant strain.
Enzymatic analysis of the P. echinulatum secretome
For further validation of proteomic data, the hydrolytic potential of stocks of the wild-type 2HH and mutant S1M29 strains was assessed using nine different polysaccharide substrates and four p-nitrophenyl (pNP) substrates. Some results in Fig. 7 are shown in another work, in the form of absolute enzymatic activity (unpublished data). However, due to proteomic analysis, some parts of these data are presented here in the form of specific enzymatic activity.
Fig. 7.
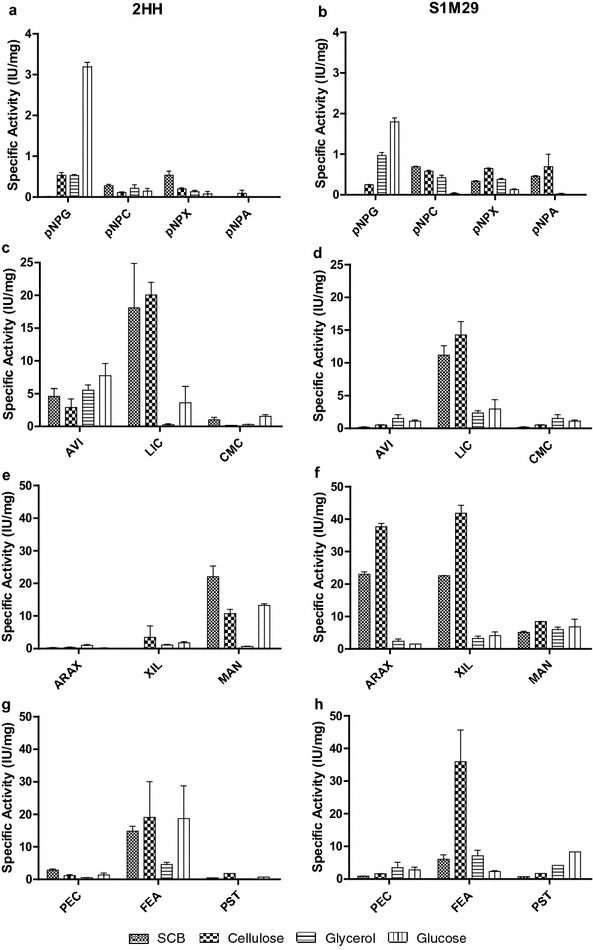
Specific enzymatic activity of Penicillium echinulatum strains in submerged cultures using different carbon sources at 96 h of cultivation. The graphs on the left correspond to the wild-type 2HH, and the graphs on the right correspond to the mutant strain S1M29. The hydrolytic potential of the broths of Penicillium echinulatum strains was tested on different substrates. a, b β-glucosidases. c, d endoglucanases. e, f xylanase, and g, h pectinase. pNPG p-nitrophenyl-β-d-glucopyranoside, pNPC p-nitrophenyl-β-d-cellobioside, pNPX p-nitrophenyl-β-d-xylopyranoside, pNPA p-nitrophenyl-α-d-arabinofuranoside, AVI Avicel®, LIC Lichenan, CMC carboxymethyl cellulose, ARAX rye arabinoxylan, XIL xylan, MAN mannan, PEC pectin, FEA feruloyl acetate, PST potato starch
The highest activity of β-glucosidases (Fig. 7a,b), using the substrate p-nitrophenyl-β-d-glucopyranoside (pNPG), was found for both strains, in the media elaborated with glucose or glycerol, which is according to the proteomic data. The largest number of spectra for cellobiohydrolases, dosed with p-nitrophenyl-β-d-cellobioside (pNPC) (Fig. 7b), was found in the media formulated with SCB or cellulose, mainly in mutant broths (Table 2).
Regarding endoglucanases activities, when the substrate lichenan has been employed, the specific activity was higher in media prepared with cellulose or SCB, for both strains, which agrees with proteomics data (Fig. 7c, d). However, upon checking the endoglucanases found in the Penicillium secretome, some spectra were found in glycerol and glucose media, for both strains (Table 2, protein g2659). It would explain the higher activities of endoglucanases in glycerol and glucose media, when the substrate carboxymethyl cellulose (CMC) was employed (Fig. 7c, d). The higher enzymatic activities on Avicel® by 2HH may be explained due the presence of multienzymatic complexes (data not shown) or higher diversity of proteins bands that can contribute to degradation of microcristaline cellulose (Fig. 7c, d).
The specific activity of xylanase, dosed with xylan from beechwood substrate (Fig. 7e, f), was higher in the medium formulated with cellulose or SCB, especially for mutant strains, which stood out with respect to the wild type, as can be seen in Table 3. The presence of β-xylosidases found in both strains with more spectra in cellulose and SCB also showed specific activities in these media (Fig. 7a, b). Higher specific activities of hemicellulases in media prepared with cellulose can be explained by the characterization of carbon sources employed. The cellulose used in this work consists of 12.19 % xylose, while SCB employed consists of 6.57 % xylose [30].
Pectinase activity was low, which is consistent with a proteome composed predominantly by cellulases (Fig. 7g, h). The low arabinofuranosidase activities, dosed with pNPA (Fig. 7a, b), in both strains are in accord with the low expression of this enzyme in the proteomic analysis. The absence of arabinose in media prepared with cellulose and the low amount of this sugar in the media prepared with SCB (~2 %) corroborate these results.
Different esterases were found in both strains, more spectra in cellulose, such as CE1, CE5, and CE10. The highest specific activity for both strains was also displayed in the medium elaborated with cellulose (Fig. 7g, h). The specific activity in starch from potato was also higher when the mutant strain was used. In both strains, the highest activity was detected in the medium prepared with glucose, followed by the one made with glycerol. However, there were limitations to establishing some relationships between enzyme activities with the results obtained in secretomic broths. This is possibly due to synergistic interactions between enzymes.
Discussion
Microbial cellulase and hemicellulase production is dependent on the carbon source [31]. The substrates used in this study have provided a high heterogeneity of secreted proteins due to the substrate and the protein profile related to each strain. The main objective of this work was achieved; that is, proteins and enzymes related to degradation of biomass, in particular the CAZymes, were found in P. echinulatum and the enzyme control change according to the different carbon source. Also, it is possible to confirm that both strains have the potential for hydrolysis of the cell wall, highlighting the specificities of the mutant strain.
Some proteomic studies involving the more efficient producer of cellulases known, i.e., T. reesei, have also been performed [32–35] and some results of these studies can be comparable to P. echinulatum. For example, in these studies involving the wild-type QM6a and the mutant RUT-C30 of T. reesei grown in media containing cellulose, corn straw, and sawdust, the authors concluded that lignocellulolytic enzyme in the secretome of both strains of T. reesei are dependent of lignocellulosic carbon sources [32]. Moreover, the functional classification of these biological quantified proteins revealed that 31.3 and 17.9 % correspond to cellulases and hemicellulases, respectively, and similar data with the amount of these enzymes, especially hemicellulases, in P. echinulatum secretome (Fig. 4).
Another similar study was conducted with Trichoderma harzianum grown on glucose, carboxymethyl cellulose, xylan, and sugar cane bagasse [33]. The characterization of T. harzianum secretome revealed that sugar cane bagasse induced greater cellulolytic and xylanolytic activities compared to the other substrates. The secretome analysis identified a wide range of proteins, including CAZymes. Although the secretome induced by sugar cane bagasse has the greatest cellulolytic and xylanolytic activities, it does not correspond to a higher complexity of proteins, since the induced secretome by carboxymethyl cellulose was significantly more diverse.
For P. echinulatum, sugar cane bagasse also induced a greater diversity of proteins being cultivated with the mutant strain, while cellulose was best for the wild type. As can be seen in Table 2, the highest amount of cellulases enzymes spectra were even checked in the sugar cane bagasse medium, suggesting that the genes related to degradation of lignocellulosic biomass are potentially transcribed during the cultivation of the fungus, but the relative proportions of the expressed proteins may vary widely depending on the growth medium and the cultivation conditions [34]. For instance, the extracellular cellulolytic system of T. reesei in response to 1 mmol/L sophorose is composed of 70 % cellobiohydrolases, 30 % endoglucanases and only 1 % of β-glucosidase [35]. In our study, however, when P. echinulatum grows in sugar cane bagasse, which proved to be ideal for production of cellulases by the mutant strain, the cellulolytic complex consists of 55 % cellobiohydrolases, 38 % endoglucanases, and 1 % β-glucosidases, being that the mutant secretes approximately two to three times more cellulases compared to the wild type, as has also been suggested for T. reesei [34].
These data lead to note that P. echinulatum presents a cellulolytic complex similar to T. reesei. However, as reported by Martins et al. [36], P. echinulatum shows higher relations between β-glucosidases and filter paper activity than T. reesei. This can be seen when glucose or glycerol are used as carbon sources for β-glucosidases production (Table 2 and Fig. 7a, b).
The genus Aspergillus is also reported in the literature as a good producer of enzymes related to degradation of biomass, and the secretome of different species has been studied [37–39]. A time course analysis of the extracellular proteome of Aspergillus nidulans growing on sorghum stover was also evaluated [38], finding several proteins in the secretome identified in P. echinulatum and additionally, mainly hemicellulases, followed by cellulases, polygalacturonases, chitinases, esterases, and lipases. Other important enzymes to reduce the recalcitrances of the biomass to hydrolysis, cellobiose dehydrogenase, and feruloyl esterase were found in great abundance. Aspergillusniger is also reported as an excellent producer of pectinases [39]. Although pectinases have been identified in P. echinulatum secretome, their diversity and amount are considerably smaller in comparison with other lignocellulolytic enzymes found.
Penicillium species are among many filamentous fungi whose secretome has been described in the last 10 years. Compared with T. reesei, lignocellulolytic enzyme systems produced by many Penicillium species have generally better performances (higher cellulose conversion at equal protein loadings) in lignocellulose hydrolysis [12]. Interestingly, P. decumbens [23] secretome showed a more diversified system of lignocellulolytic enzymes than T. reesei, particularly for cellulose binding domain-containing proteins and hemicellulases. Moreover, the proteomic analysis of P. decumbens revealed that the production of lignocellulolytic enzymes is greater in media containing cellulose and wheat bran than in glucose, which agrees with part of our studies.
Liao et al. [13] verified that a complex substrate composed of cellulose and xylan, simulated an artificially plant biomass, is more efficient than purified cellulose at inducing lignocellulolytic enzyme production in Penicillium oxalicum GZ-2. The addition of xylan to the cellulose culture did not affect fungal growth but significantly increased the activities of cellulase and hemicellulase. However, in T. reesei RUT-C30, cellulose induces a more efficient response.
Relating these results with P. echinulatum, we can conclude that the same relationship exists between P. echinulatum S1M29 and P. oxalicum GZ-2, at least, when we compare the numbers of cellulase and hemicellulase spectra and the protein concentrations in SCB. The data indicate that the mutant S1M29 is more suitable for the production of enzymes to hydrolyze the plant cell wall in comparison with the wild type, especially when it was grown in a culture medium formulated with SCB. This information is relevant for the production of enzyme complexes at lower cost, since there is a greater amount of proteins secreted by the mutant to degrade lignocellulosic wastes that are less costly and more complex.
In the production of second-generation ethanol, the cost of the enzyme is one of the major bottlenecks, and the cost of the substrate for the enzyme production can contribute decisively to this value; a ton of cellulose used for the cellulases production costs around US$1000 per ton, while the steam explosion SCB, is commercialized at US$40 a ton.
However, when we look at the specific activities, P. echinulatum S1M29 has the same behavior as T. reesei RUT-C30, where cellulose induces better enzymatic activities, at least for hemicellulose (Fig. 7f). This can be explained by the fact that the cellulose used in this work has more xylose than the SCB. Moreover, SCB used in our study was pretreated by steam explosion, and this process is characterized by the removal of a part of hemicellulose [40]; because of this, the lowest concentration of xylose in SCB did not favor the increased production of xylanase.
When P. oxalicum was grown in a medium with glycerol or glucose as the sole carbon source, low activities of cellulase and xylanase were detected, suggesting that some components are expressed at basal levels. When lignocellulosic substrates (e.g., cellulose, xylan, xylose, and wheat bran) were used as carbon sources, significantly higher lignocellulolytic enzyme activities were detected. The repressing effect of glucose and inductive effect of cellulose on the expressions of cellulases and xylanases were verified at gene-transcriptional levels [12]. These statements are consistent with the roles of each component in the production of lignocellulolytic enzymes by P. echinulatum, verified in our research.
Few data related to genetics of P. echinulatum have been published until now. Rubini et al. [41] isolated, cloned, and expressed one endoglucanase egl 1 of the mutant P. echinulatum 9A02S1, and found high hydrolytic activity in a medium formulated with CMC. Endoglucanasic activity is a feature of P. echinulatum that remained in S1M29 strain and is according to the cellulolytic activities detected in this strain, as can be seen in Table 2 (protein g8473), showing that it is well expressed.
The use of different carbon sources for the production of cellulases and hemicellulases by P. echinulatum was previously reported by Novello et al. [21] and Schneider et al. [22]. Novello et al. [21] studied the production of endoglucanases, β-glucosidases, and xylanases by strains 2HH, 9A02S1, and S1M29 of P. echinulatum, employing cellulose, glucose, and xylose as carbon sources and reported that xylose acts as an inducer for the production of xylanases and cellulases, in particular, endoglucanases. The authors also found higher enzyme titers in cultures carried out with mutant strains 9A02S1 and S1M29 than those with the wild-type 2HH.
Schneider et al. [22] studied the effect of six different carbon sources (sucrose, glucose, glycerol, cellulose Celuflok®, untreated elephant grass, and SCB) in the production of enzymes by the mutant strain S1M29 of P. echinulatum. Among the six carbon sources, cellulose and SCB were the most suitable for the production of filter paper activity, endoglucanase, xylanases, and β-glucosidases. However, sucrose and glucose showed β-glucosidase activities similar to those obtained with the insoluble sources. The presence of these enzymes, especially β-glucosidase, in the work of Schneider et al. [22], with more activity at 96 h (at the end of the cultivation process), justifies the choice of 96-h duration for proteome analysis in the present study.
Some enzymes detected, such as β-1,3-glucanase (GH17, Table 10), do not have a secretion signal and may be related to a possible indication of cell wall autolysis at 96 h of cultivation, as reported by Vaheri et al. [42] and Fontaine et al. [43]. Many other fungal cell wall degrading enzymes, such as chitinases (GH18, Table 10), exo-β-1,3-glucanase (GH17, Table 10), and others already cited (chitosanase, β-N-acetylhexosaminidase, and lysophospholipase), may also be related to cell lysis. All these enzymes can be induced simply by the same sugars as the other CAZymes. This may be an indication that 2HH responds worse to stress than S1M29, and thus, produces a smaller amount of enzymes, which actually contributes to the degradation of biomass. It is suggested that the two strains deal with stress differently; the mutant is at a less-advanced stage of stress using mainly oxidative enzymes to maintain itself, while the wild type has passed through this stage and is already in a degrading process. From the analysis of the mycelial growth on glucose (data not shown), higher concentration of biomass has been found in wild type in relation to mutant up to 96 h of cultivation. Therefore, the greater growth of wild type on carbon sources of facilitated uptake, such as glucose, may be providing the production of these enzymes related to degradation of fungal cell walls, whereas the mutant makes use of more complex carbon sources for the production of enzymes related to degrading biomass.
The only secretome work on P. echinulatum done so far was conducted by Ribeiro et al. [5], employing the intermediate mutant 9A02S1. In their study, the secretome profiles of P. echinulatum after growing on integral sugar cane bagasse, microcrystalline cellulose, and three types of pretreated sugar cane bagasse were evaluated using shotgun proteomics. The same study revealed that the enzymatic repertoire of P. echinulatum is geared mainly toward producing enzymes from the cellulose complex (endoglucanases, cellobiohydrolases, and β-glucosidases). Glycoside hydrolase family members, important in terms of biomass-to-biofuel-conversion strategies, were identified—including endoglucanases GH5, 6, 7, 12, and 17; β-glycosidase GH3; xylanases GH10 and GH11, as well as debranching hemicellulases from GH43, GH62, and CE2; and pectinases from GH28.
The above results as found in that study agree with the information obtained from the present study, conducted with 2HH and S1M29 strains, where a larger amount of spectra in most enzymes was verified in media formulated with cellulose or SCB (in our study, pretreated by steam explosion), relating to degradation of biomass, such as cellobiohydrolases I and II (GH 6 and 7), endoglucanases 1 and Cel5C (GH5), and xylanases (GH10), demonstrating the potential of producing these enzymes especially by mutant strain S1M29. Although there is not much difference in enzyme diversity between the wild type and the mutants, the mutant S1M29 secretes greater diversity of glycosyl hydrolases compared to the mutant 9A02S1, e.g., GH15, 16, 30, 31, 72, 75, 99, 125, and 132, as well as other CAZy enzymes (Table 11). The specific activity also reveals a change in secretion of some enzymes, when we monitor the genetic improvement of the strain. This can be evidenced by the xylanase activity, which activity increased nearly ten times when comparing the mutants S1M29 (this work) with 9A02S1 [4]. In smaller proportions, the same thing happens with β-glucosidases and cellobiohydrolases activities. In addition, the database employed in our work refers to the sequencing of P. echinulatum strains—another aspect that makes this proteomic analysis to be more consistent.
Table 11.
Comparison between secretomic profiles of different Penicillium echinulatum strains
| Comparative parameters | Penicillium echinulatum strains | ||
|---|---|---|---|
| Mutant 9A02S1 (Ribeiro et al. 2012) |
Wild-type 2HH (This work) |
Mutant S1M29 (This work) |
|
| Proteomic approach | Shotgun LC–MS/MS |
Shotgun 1D-PAGE/LC–MS/MS |
Shotgun 1D-PAGE/LC–MS/MS |
| Carbon sources | Sugar cane bagasse and cellulose | Sugar cane bagasse, cellulose, glucose, and glycerol | Sugar cane bagasse, cellulose, glucose, and glycerol |
| Days of growth | 5 | 4 | 4 |
| Proteins identified | 99 | 147 | 129 |
| Glycoside hydrolases | 16 | 24 | 25 |
| Major proteins found | Cellobiohydrolases Endoglucanases β-glucosidases Xylanases Debranching hemicellulose/pectin Pectinases Swollenin |
Cellobiohydrolases Endoglucanases β-glucosidases Xylanases Debranching hemicellulose/pectin Pectinases Swollenin Cellulose monooxygenases Ligninases |
Cellobiohydrolases Endoglucanases β-glucosidases Xylanases Debranching hemicellulose/pectin Pectinases Swollenin Cellulose monooxygenases Ligninases |
The presence of cellulose monoxygenase Cel61A (Table 2/g3303), an important enzyme responsible for the oxidation of glycan chains on cellulose, found in P. echinulatum S1M29 secretome, is one more difference of this mutant; on the proteomic analysis of mutant strain 9A02S1, this key enzyme was not found. This feature also makes the secretome of P. echinulatum even more similar to P. oxalicum; besides presenting various cellulases and hemicellulases in common, this enzyme is found in higher proportions in P. oxalicum than in T. reesei [12].
The presence of nonhydrolytic proteins like swollenins, proteins similar to plant cell wall expansins, which can disrupt the crystalline parts of the cellulose chains, was observed only in the medium formulated with SCB or cellulose; for both strains, however, more were expressed in the mutant strain. Swollenins contain a carbohydrate-binding domain (CBM) and have been proposed to disrupt cellulose structure via nonhydrolytic mechanisms [44], although the biochemical action of these proteins remains to be fully elucidated [45]. As substrate accessibility is one key issue in plant cell wall degradation, accessory proteins are likely to enhance the efficiency of this process [46]. However, in some cases, the presence of a CBM in an enzyme may also prevent it from acting at different points on a substrate.
The presence of some enzymes involved in the degradation/depolymerization of lignin, such as isoamyl alcohol oxidase [47], manganese superoxide dismutase [48], and glutathione S-transferase [49] showed the first evidence of these enzymes’ production by the fungus P. echinulatum. This is more a characteristic of the genetic evolution of the fungus, since the secretome analysis of mutant 9A02S1 [5], strain before the S1M29 mutant, failed to identify lignin proteins, such as extracellular oxidase.
The lignin peroxidase, manganese-peroxidase, and laccase were generally considered as lignin degrading enzymes. However, according to Blanchette et al. [50] they are too big to penetrate the plant cell wall. Hence, to get access microbes initially activate easily diffusing several oxidases, reactive radical generating enzymes, and quinine reducing enzymes [51, 52]. The spectral counts of isoamyl alcohol oxidase, flavin adenine dinucleotide (FAD) oxidoreductase, and superoxide dismutase indicated that P. echinulatum degrade lignin through oxidases. In addition, this study also quantified expressions of glutathione-S-transferase, another enzyme important to the mechanism of lignin degradation.
The results obtained in this work suggest that in the genetic improvement process of P. echinulatum, performed by mutagenesis employing different mutagens and protoplast fusions [15, 16], numerous changes, possibly at the level of regulation/gene expression, post-translational modifications, and alterations in the capacity to secrete extracellular proteins, occurred with the mutant. These doubts will be elucidated in the future with a conjunct analysis of the secretome, transcriptome, and genome data.
Conclusions
In this study, among the 165 proteins identified in the secretomes of both strains, approximately 40 % were proteins with CAZy function. The glycoside hydrolases were the most abundant. The secretome of P. echinulatum presents a potential for biomass degradation, mainly of cellulose and hemicellulose. Pectinases were found in minor amounts; however, for the first time, proteins related to lignin degradation were found.
Although cellulose and SCB were the carbon sources resulting in the production of cellulolytic and hemicellulolytic enzymes, glucose and glycerol inducing other proteins as enzymes related to lignin degrading and the presence of β-glucosidase were found more in glycerol than in any other carbon source. Differences in the protein expressions through enzymatic activity and by analyzing the number of spectra generated by proteomics data show the distinction between the secretomes of wild-type 2HH and mutant S1M29, in which the mutant toolbox is more focused on the production of cellulases and hemicellulases than the wild type. These findings allow us to confirm the potential that the strains of P. echinulatum, in particular, the mutant, have for the lignocellulosic biomass degradation.
Methods
Growth and maintenance of Penicillium strains
The wild-type 2HH and mutant S1M29 strains of P. echinulatum were grown and maintained in 100 mL of cellulose agar (agar-C) consisting of 40 mL of swollen cellulose, 10 mL of mineral solution (KH2PO4, 10 g; MgSO4.7H2O, 6 g; CO(NH2)2, 6 g; CaCl2, 6 g; FeSO4.7H2O, 0.1 g; MnSO4.H2O, 0.0312 g); ZnSO4.7H2O, 0.028 g; CoCl2.6H2O, 0.04 g; 1 L distiller water), 0.1 g of proteose peptone (Oxoid L85®), 2 g of agar, and 50 mL of distilled water. The strains were grown in inclined tubes on C-agar for 7 days at 28 °C until the formation of conidia, and then stored at 4 °C, as shown in Dillon et al. [15].
Genome sequencing, assembly, and scaffolding
Genome sequencing
High-molecular-weight genomic DNA was extracted from both the 2HH and S1M29 strains using a protocol for DNA isolation [53] and used to generate libraries for Illumina Sequence by Synthesis (Illumina-SBS) genome sequencing using an unmodified Illumina TruSeq DNA protocol [54]. Post-adaptor ligation size-selected fragments were used for flow-cell cluster generation on the Illumina HiSeq 2000 platform. Illumina 100 bp paired-end chemistry sequencing was conducted using the commercial provider Ambry Genetics (Aliso Viejo, CA, USA). Illumina sequencing yielded 339,864.496 (33.98 GB) quality filtered reads for strain 2HH and 272,677.192 (27.26 GB) for strain S1M29 respectively.
Genome assembly
Illumina quality filtered reads from both the 2HH and S1M29 genome were assembled separately using the De-Bruijn graph short read assembly program Velvet version 1.2.10. The assembly run-time settings used for Velvet were a kmer value of 47, a minimum contig coverage of 7× and a minimum contig length setting of 300 bp. The Velvet assembly produced a total of 29.82 MB, 1018 contigs (n50 147.866 bp/n90 32.016 bp) for the 2HH genome and 29.83 MB, 1124 contigs (n50 137.718/n90 32.016) for the S1M29 genome, respectively. All assembled contigs were used for subsequent gene model creation, functional annotation, comparative studies, and downstream analysis.
Gene calling
All contigs produced from both Velvet assemblies of S1M29 and 2HH were used for genomic analysis. A total of 392,882,306 RNA-seq cDNA reads (39.28 GB) were generated from S1M29 strain using the Illumina TruSeq protocol with poly-A selection were used for gene model training prediction and cDNA alignment using the Eukaryotic gene prediction software Augustus. RNA-seq data were supplied as training files as mentioned by the Augustus RNA-seq protocol which leverages the PASA (Program to Assemble Spliced Assemblies) software [55]. Gene models produced by the Augustus protocol were used for genome annotation, proteomic database creation, and comparative analysis.
Functional annotation and proteome data bank assembly
Bioinformatics tools were used to build a secretome table. To validate the proteomic data the Scaffold 4 Proteomic software (version 4.3.2 20140225) was employed, where the database of proteins from the two strains of P. echinulatum (2HH and S1M29) were crossed. Conditions configured to accept the identification of a protein were: protein probability thresholds greater than 99 %, with a minimum of two different peptides for protein identification, each one with 95 % certainty. The search results showed false discovery rate (FDR) of peptides and proteins equal to zero.
Spectrum count was introduced as a semiquantitative method for the analysis of shotgun mass spectrometry data. Spectrum counts are the total number of tandem spectra assigned to each protein and are commonly used to determine their relative protein abundances, since previous studies have demonstrated a linear correlation between the spectrum count and protein abundance in complex samples [38].
As was used in the analysis performed using the methodology of spectrum count, the FDR was used as one of the parameters for determining the reliability of our experiments. The FDR is defined as the expected percentage of accepted peptide-spectrum match (PSM), with an associated score that is incorrect, where an “accepted PSM” is when it scores above the threshold [56]. Initially, a score is given for each peptide in the primary analysis performed with the Mascot Distiller software v.2.3.2.0, 2009 (Matrix Science Ltd.). This value shows the abundance of each peptide and also the reliability of these data. When the analysis was performed with the Scaffold 4 Proteomic software, the number of spectra was used to measure the abundance and the FDR for reliability. The threshold of ion score was 25 and, as mentioned before, our results showed FDRs of peptides and proteins are equal to zero.
In order to verify if the sequences of proteins annotated from the genomes of 2HH and S1M29 strains with different identification were the same, the alignment of the proteins was performed using the Clustal Omega software (Multiple Sequence Alignment—EMBL-EBI) [57].
To verify the protein sequences, BLAST (basic local alignment search tool) [58] software was employed. The parameters used to make the choice of protein were: e value ≤ e−40; identity ≥ 40 % and query cover ≥ 80 %.
With the aim of finding proteins with activity on carbohydrates (CAZymes), as well as the presence or absence of carbohydrate-binding modules, the software dbCAN (DataBase for Carbohydrate-active enzyme ANotation) was used [59].
The identification of secreted proteins was defined by the interpretation of the results of the following softwares, SignalP (version 4.1, Server-CBS) [60], SecretomeP (version 2.0) [61] and YLoc (interpretable subcellular localization prediction) [62].
Production of enzymes
The culture medium for enzyme production consisted of 0.2 % (w/v) peptone; 0.05 % (w/v) Prodex®; 1 % (w/v) carbon source; 0.1 % (v/v) Tween 80®; 0.002 % (v/v) antibiotic ciprofloxacin (Proflox®, EMS S/A); 5 % (v/v) mineral solution (describe in “Growth and maintenance of Penicillium strains”); and distilled water to complete a final volume of 100 mL.
Erlenmeyer flasks (500 mL), containing 100 mL of the production medium, were inoculated with a suspension of 1 × 105 conidia per mL and kept at 28 °C, in a reciprocal agitation of 180 rpm for 120 h. The experiment was performed in triplicate, and supernatants for 48, 96, and 120 h of cultivation were separated for protein determination, and the 96-h one was also used for proteomic data and enzymatic analysis. The samples were kept under refrigeration and sodium azide for a final concentration of 0.02 % (w/v).
To evaluate the effect of different carbon sources on the production of enzymes by the wild-type and mutant P. echinulatum, the following were employed: cellulose Celuflok E® (Celuflok Comercial Ltda, Cotia, SP, Brazil), sugar cane bagasse pretreated by steam explosion (Usina Vale do Rosário, Morro Agudo, SP, Brazil), glucose (Quimidrol, Joinville, SC, Brazil), and glycerol (Sigma-Aldrich, St. Louis, MO, USA).
Enzyme dosages
The specific enzymatic activity of the collected supernatants at 96 h of cultivation point was tested using a set of 13 different substrates (Megazyme, Wicklow, Ireland; and Sigma-Aldrich, St. Louis, MO, USA) in order to measure the activity of cellulases, hemicellulases, pectinases, esterases, and amylases.
To determine the cellulases, the following substrates were employed: Avicel®, carboxymethyl cellulose (CMC), lichenan (from Cetraria islandica), p-nitrophenyl-β-d-cellobioside (pNPC), and p-nitrophenyl-β-d-glucopyranoside (pNPG). The following substrates were employed to determine the hemicellulases: rye arabinoxylan, xylan from beechwood, p-nitrophenyl-β-d-xylopyranoside (pNPX), and mannan. Pectinases were measured using pectin substrate (from citrus fruits) and p-nitrophenyl-α-d- arabinofuranosidase (pNPA). Esterases were determined with feruloyl acetate and amylases were dosed with potato starch substrate.
All enzymatic activities were performed in triplicate. The methodology employed to determine enzymatic activities was performed according to Cota et al. [63] with modifications, where 50 μL of substrate solution (0.5 % w/v of the substrate diluted in water) were added to 46 μL of enzyme solution and 4 μL of sodium citrate buffer 1 mol/L, pH 4.8. The mixture was incubated at 50 °C for different periods depending on the substrate. The duration of the reaction was 30 min for potato starch, Avicel®, CMC, mannan, pectin, pNPA, pNPX, pNPC, and pNPG (for the latter two, 20 μL of enzyme solution, 30 μL of sodium citrate buffer 1 mol/L dilute to the samples containing cellulose or SCB). The reaction time of 10 min was used for lichenan, for samples containing SCB or cellulose (10 μL enzyme solution, 4 μL sodium citrate buffer, and 36 μL distilled water); 10 min for xylan and rye arabinoxylan for samples of the mutant strain grown in SCB or cellulose (10 μL enzyme solution, 4 μL sodium citrate buffer, and 36 μL distilled water). The reactions with polysaccharides were stopped by adding 300 and 100 μL of dinitrosalicylic acid reagent solution (DNS), respectively [64]. The reactions carried out with pNP substrates were stopped by adding 100 μL of sodium carbonate 10 (w/v) [65].
The activity of esterases was determined according to the methodology described by Koseki et al. [66]. For the samples containing cellulose or SCB, 5 μL of enzyme solution and 20 μL of sodium acetate buffer (SAB) 100 mmol/L, pH 5.5 were used, and 15 μL distilled water was added. For samples containing glucose or glycerol as carbon sources, 20 μL of enzyme solution and 20 μL of SAB buffer were used. Then, 10 μL of substrate (50 mmol/L in dimethylsulfoxide solution, DMSO) was added, incubated at 40 °C in a thermocycler for 30 min. Next, the reaction was terminated by adding 100 μL solution of Fast Garnet reagent (0.1 % w/v and SDS 15 % w/v in DMSO) and kept at room temperature for 10 min.
The units of enzymes with activity on the polysaccharides were defined as the amount of enzyme capable of releasing 1 μmol of reducing sugar per min. Units of enzyme with activity on the p-nitrophenyl substrates were defined as the amount of enzyme capable of liberating 1 μmol of p-nitrophenyl per min. Esterases were defined as the amount of enzyme capable of liberating 1 μmol of α-naphthyl acetate per minute.
Determination of total soluble proteins
For the quantitative determination of soluble proteins in the secretomes of P. echinulatum wild-type and mutant strains, we employed the method of Bradford [67] with Bio-Rad Protein Assay. A calibration curve was constructed using 80 μL bovine serum albumin (BSA) standard solutions with concentrations between 0 and 25 μg/mL and 20 μL of Bradford reagent. The protein quantification of P. echinulatum secretome samples was carried out with 20 μL of Bradford reagent and 80 μL of sample. The reaction was incubated for 10 min at room temperature, and then readings were taken on a spectrophotometer at 595 nm.
SDS-PAGE of total proteins
Samples collected at 96 h of cultivation from P. echinulatum were subjected to analysis of the protein profile. Samples containing cellulose or SCB were made in biological triplicate, whereas the samples containing glucose or glycerol were produced as a replicate. The most relevant comparisons are between the inducing sources, cellulose and SCB. Glucose and glycerol were used as external controls of low production, somehow, serving as a kind of normalization of experiments.
To determine the molecular weight of proteins, electrophoresis was performed in polyacrylamide gels containing 0.1 % (w/v) sodium dodecyl sulfate (SDS-PAGE). The separating gel was prepared in 12 % (w/v) while the stacker gel was prepared 4 % (w/v) according to the methodology described by Laemmli [68]. After standard the samples in 11 μg, it was applied to each slot to electrophoresis running in vertical cube Bio Rad Mini Protean System Cell at 110 V for approximately one and a half hours.
The revelation of bands from the gel was performed incubating the gel for 30 min in a solution of 0.2 % (w/v) Coomassie Brilhant Blue G 250, 50 % (v/v) of ethanol, and 10 % (v/v) of acetic acid. Afterward, the gel was washed with distilled water and immersed in a solution of 50 % (w/v) ethanol and 10 % (w/v) acetic acid for 30 min. The entire process was performed under stirring of 50 rpm until bands were visualized.
Digestion of proteins for analysis by mass spectrometry
The digestion of proteins for analysis by mass spectrometry was performed in two steps, performing them in two successive days, according to Gonçalves et al. [69]. First we cut eight gel bands per lane. SDS was removed with 500 μL of a destain solution during 2 h and the bands were dehydrated for 5 min with 200 μL acetonitrile, reduced for 30 min with 30 μL dithiothreitol (DTT) solution, and alkylated with 30 μL of iodoacetoamide (IAA) solution, also for 30 min. Then the bands were washed with 100 mmol/L ammonium bicarbonate (for 10 min). A new dehydration with acetonitrile and rehydration with sodium bicarbonate was performed. Protein digestion was carried out with 30 μL of trypsin solution (1 mg/mL) in 50 mmol/L ammonium bicarbonate at 37 °C overnight.
On the second day, 10–30 μL of extraction solution 1 (5 % (v/v) of formic acid) was added to each microtube (depending on the size of the gel), incubated for 10 min at room temperature, given a quick spin in a centrifuge, and then the supernatant was collected and transferred to another microtube. Then, 12 μL of the second extraction solution (5 % (v/v) formic acid in 50 % (v/v) acetonitrile) was added to each microtube, and after 10 min, the supernatant was collected and transferred to a tube which was previously kept separated and had already contained the extract from the previous step. This step was repeated once more. Finally, the samples were evaporated in the speed vac, and approximately 1 μL of sample was left. The samples were stored at −20 °C until being transferred to the mass spectrometer subsequently.
Liquid chromatography-tandem mass spectrometry
For the application of trypsinized fractions in the LC–MS/MS, each sample was resuspended with 12 μL of 0.1 % (v/v) formic acid and aliquot of 4.5 μL of the peptides mixture was injected into the chromatograph RP-nanoUPLC (nanoAcquity, Waters). Chromatography was performed on C18 (100 × 100 mm) equilibrated with 0.1 % (v/v) formic acid column buffer. The elution gradient was 2 % (v/v) to 90 % (v/v) acetonitrile in 0.1 % (v/v) formic acid. The whole system operated at the speed of 0.6 µL/min. As the peptides were eluted from the column, they were injected into the spectrometer quadrupole-time of flight Q-Tof (Ultima Mass Spectrometer Waters) with a source of ionization electron spray, for 60 min. The instrument was operated in the “top three–MS and MS/MS mode”, where each MS spectrum acquired, its three peptides not monoloaded (precursors) most abundant were selected for further fragmentation (generating y and b sets) and sequenced generating a MS/MS spectrum for each peptide.
The spectra were acquired using MassLynx v.4.1 software (Waters, Milford, MA, USA), and the raw data were converted to the format “peak list format (mgf)” by Mascot Distiller software v.2.3.2.0, 2009 (Matrix Science Ltd.). These results were processed by Mascot v.2.3.01 engine (Matrix Science Ltd.) software against the genome-sequencing database of P. echinulatum strains (2HH: 8504 sequences, 4286814 residues; M29: 8552 sequences, 4298942 residues) for digestion and theoretical spectral generation of MS/MS. The following parameters were used in this process: carbamidomethylation as a fixed modification, oxidation of methionine as a variable modification, one error of trypsin cleavage, and maximum allowable error in the peptide mass of 0.1 Da. Only peptides with at least five amino acids and a score within probability to be not a random event p < 0.05 were selected as a peptide cleavage product (i.e., they are part of a protein). The peptide was considered only when it differed by at least one amino acid for another or when it differed in covalent modifications (including elongations of N- or C-terminal).
Analysis of results
The analyses of data regarding the protein concentration and enzymatic activities were performed in PrismGraphPad (version 5.0.1.334) software. The statistical analysis of protein concentration was performed by analysis of variance and Tukey’s post-test for a p < 0.05, and the statisticSpecific enzymatic activity of Penicilliumal analysis of spectra was performed by t test for a p < 0.05.
Authors’ contributions
WDHS carried out the enzymes production and the proteomic studies, and wrote the manuscript. TAG carried out the proteomic studies and collaborated in writing the manuscript. CAU carried out the proteomic studies. MBC carried out the genomics studies and the proteomic data. RP carried out the genomics studies and the proteomic data. FMS advised and carried out the proteomic and genomics studies. AJPD participated in its design, coordination, and helped in drafting the manuscript. MC participated in its the design, coordination, and helped in the enzyme production and in drafting the manuscript. All authors read and approved the final manuscript.
Acknowledgements
The authors thank the Coordination of Improvement of Higher Education Personnel (CAPES), the National Laboratory of Science and Technology of Bioethanol (CTBE), the Laboratory of Mass Spectrometry of Brazilian Biosciences National Laboratory (LNBio), and the University of Caxias do Sul (UCS). We are grateful to FAPESP (The State of São Paulo Research Foundation) for the post-doctoral scholarship (2012/19040-0 to CAU) and financial support (2008/58037-9 and 2014/06923-6). We are grateful to the National Council for Scientific and Technological Development (CNPq) for the PhD scholarship (140796/2013-4 to TAG) and financial support (310186/2014-5, 442333/2014-5).
Competing interests
The authors declare that they have no competing interests.
Abbreviations
- 1D-PAGE
one dimensional polyacrylamide electrophoresis
- LC–MS/MS
liquid chromatography-tandem mass spectrometry
- CAZy/CAZymes
carbohydrate active enzymes
- SCB
sugar cane bagasse pretreated by steam explosion
- CEL
cellulose
- GLY
glycerol
- GLU
glucose
- GHs
glicoside hydrolases
- CEs
carbohydrate esterases
- PLs
polysaccharide lyases
- AAs
auxiliary activities
- CBMs
carbohydrate-binding module
- pNP
p-nitrophenyl
- pNPC
p-nitrophenyl-β-d-cellobioside
- pNPG
p-nitrophenyl-β-d-glucopyranoside
- pNPX
p-nitrophenyl-β-d-xylopyranoside
- pNPA
p-nitrophenyl-α-d-arabinofuranoside
- AVI
Avicel®
- LIC
lichenan
- CMC
carboxymethyl cellulose
- ARAX
rye arabinoxylan
- XIL
xylan
- MAN
mannan
- PEC
pectin
- FEA
feruloyl acetate
- PST
potato starch
- DNS
dinitrosalicylic acid
- SAB
sodium acetate buffer
- DMSO
dimethyl sulfoxide
- BSA
bovine serum albumin
- DTT
dithiothreitol
- IAA
iodoacetoamide
- SDS
sodium dodecyl sulfate
- UPLC
ultra performance liquid chromatography
- Q-Tof
quadrople time of fligth
- FDR
false discovery rate
- PSM
peptide-spectrum match
- BLAST
basic local alignment search tool
- dbCAN
database for carbohydrate-active enzyme annotation
- YLoc
interpretable subcellular localization prediction
Additional files
10.1186/s13068-016-0476-3 Other groups of proteins identified and spectrum count in different carbon sources by wild type 2HH and mutant S1M29 of Penicillium echinulatum at 96 h during submerged cultivation. This table shows other groups of proteins found in Penicillium echinulatum secretome, as adhesion proteins, oxidoreductases, proteases and peptidases, lipases, glutaminases, chaperones, hypothetical proteins and miscelaneous proteins.
10.1186/s13068-016-0476-3 Comparative analysis of the most expressed proteins in each condition for each Penicillium echinulatum strain. This comparative analysis was based on the list, in order, of ten proteins with more spectra found in each carbon source in the wild type 2HH and mutant S1M29 and also the most expressed proteins when analyzing both strains.
10.1186/s13068-016-0476-3 Complete protein reports of Penicillium echinulatum strains in each condition. This table shows important data of proteomic analysis: number of peptides used for the identification and sequence coverage (%).
Footnotes
Willian Daniel Hahn Schneider and Thiago Augusto Gonçalves contributed equally to this work
Contributor Information
Willian Daniel Hahn Schneider, Email: willianschneiderwhs@yahoo.com.br.
Thiago Augusto Gonçalves, Email: thiago.bioetanol@gmail.com.
Cristiane Akemi Uchima, Email: cris.uchima@gmail.com.
Matthew Brian Couger, Email: mbcouger@gmail.com.
Rolf Prade, Email: rolf.prade@okstate.edu.
Fabio Marcio Squina, Email: fabio.squina@bioetanol.org.br.
Aldo José Pinheiro Dillon, Email: ajpdillo@ucs.br.
Marli Camassola, Phone: +55 54 3218 2681, Email: mcamassola@gmail.com.
References
- 1.Rubin EM. Genomics of cellulosic biofuels. Nature. 2008;454:841–845. doi: 10.1038/nature07190. [DOI] [PubMed] [Google Scholar]
- 2.Glass NL, Schmoll M, Cate JHD, Coradetti S. Plant cell wall deconstruction by ascomycete fungi. Annu Rev Microbiol. 2013;67:477–498. doi: 10.1146/annurev-micro-092611-150044. [DOI] [PubMed] [Google Scholar]
- 3.Popper ZA, Michel G, Herve C, Domozych DS, Willats WG. Evolution and diversity of plant cell walls: from algae to flowering plants. Annu Rev Plant Biol. 2011;62:567–590. doi: 10.1146/annurev-arplant-042110-103809. [DOI] [PubMed] [Google Scholar]
- 4.Pérez J, Muñoz-Dorado J, De-la-Rubia T, Martínez J. Biodegradation and biological treatments of cellulose, hemicellulose and lignin: an overview. Int Microbiol. 2002;5:53–63. doi: 10.1007/s10123-002-0062-3. [DOI] [PubMed] [Google Scholar]
- 5.Ribeiro DA, Cota J, Alvarez TM, Bruchli F, Bragato J, Pereira BMP, Pauletti BA, Jackson G, Pimenta MTB, Murakami MT, Camassola M, Ruller R, Dillon AJP, Pradella JG, Paes Leme AF, Squina FM. The Penicillium echinulatum secretome on sugar cane bagasse. PLoS One. 2012;7:e50571. doi: 10.1371/journal.pone.0050571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Tian C, Beeson WT, Iavarone AT, Sun J, Marletta MA, Cate JHD, Glass NL. Systems analysis of plant cell wall degradation by the model filamentous fungus Neurospora crassa. Proc Nat Acad Sci. 2009;106(52):22157–22162. doi: 10.1073/pnas.0906810106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Lynd LR, Weimer PJ, van Zyl WH, Pretorius IS. Microbial cellulose utilization: fundamentals and biotechnology. Microbiol Mol Biol Rev. 2002;66:506–577. doi: 10.1128/MMBR.66.3.506-577.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Martinez D, Berka RM, Henrissat B, Saloheimo M, Arvas M, Baker SE, Chapman J, Chertkov O, Coutinho PM, Cullen D, Danchin EG, Grigoriev IV, Harris P, Jackson M, Kubicek CP, Han CS, Ho I, Larrondo LF, de Leon AL, Magnuson JK, Merino S, Misra M, Nelson B, Putnam N, Robbertse B, Salamov AA, Schmoll M, Terry A, Thayer N, Westerholm-Parvinen A, Schoch CL, Yao J, Barabote R, Nelson MA, Detter C, Bruce D, Kuske CR, Xie G, Richardson P, Rokhsar DS, Lucas SM, Rubin EM, Dunn-Coleman N, Ward M, Brettin TS. Genome sequencing and analysis of the biomass-degrading fungus Trichoderma reesei (syn. Hypocrea jecorina) Nat Biotechnol. 2008;26:553–560. doi: 10.1038/nbt1403. [DOI] [PubMed] [Google Scholar]
- 9.Schuster A, Schmoll M. Biology and biotechnology of Trichoderma. Appl Microbiol Biotechnol. 2010;87(3):787–799. doi: 10.1007/s00253-010-2632-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Martens-Uzunova ES, Schaap PJ. Assessment of the pectin degrading enzyme network of Aspergillus niger by functional genomics. Fungal Genet Biol. 2009;46:S170–S179. doi: 10.1016/j.fgb.2008.07.021. [DOI] [PubMed] [Google Scholar]
- 11.Gusakov AV, Sinitsyn AP. Cellulases from Penicillium species for producing fuels from biomass. Biofuels. 2012;3:463–477. doi: 10.4155/bfs.12.41. [DOI] [Google Scholar]
- 12.Liu G, Qin Y, Li Z, Qu Y. Improving lignocellulolytic enzyme production with Penicillium: from strain screening to systems biology. Biofuels. 2013;4(5):523–534. doi: 10.4155/bfs.13.38. [DOI] [Google Scholar]
- 13.Liao H, Li S, Wei Z, Shen Q, Xu Y. Insights into high-efficiency lignocellulolytic enzyme production by Penicillium oxalicum GZ-2 induced by a complex substrate. Biotechnol Biofuels. 2014;7:162. doi: 10.1186/s13068-014-0162-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Yao G, Li Z, Gao L, Wu R, Kan Q, Liu G, Qu Y. Redesigning the regulatory pathway to enhance cellulose production in Penicillium oxalicum. Biotechnol Biofuels. 2015;8:71. doi: 10.1186/s13068-015-0253-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Dillon AJP, Zorgi C, Camassola M, Henriques JAP. Use of 2-deoxyglucose in liquid media for the selection of mutant strains of Penicillium echinulatum producing increased cellulase and β-glucosidase activities. App Microbiol Biotechnol. 2006;70:470–476. doi: 10.1007/s00253-005-0122-7. [DOI] [PubMed] [Google Scholar]
- 16.Dillon AJ, Bettio M, Pozzan FG, Andrighetti T, Camassola M. A new Penicillium echinulatum strain with faster cellulase secretion obtained using hydrogen peroxide mutagenesis and screening with 2-deoxyglucose. J Appl Microbiol. 2011;111(1):48–53. doi: 10.1111/j.1365-2672.2011.05026.x. [DOI] [PubMed] [Google Scholar]
- 17.Camassola M, Dillon AJP. Cellulases and xylanases production by Penicillium echinulatum grown on sugar cane bagasse in solid-state fermentation. Appl Biochem Biotechnol. 2010;162(7):1889–1900. doi: 10.1007/s12010-010-8967-3. [DOI] [PubMed] [Google Scholar]
- 18.Zampieri D, Guerra L, Camassola M, Dillon AJP. Secretion of endoglucanases and β-glucosidases by Penicillium echinulatum 9A02S1 in presence of different carbon sources. Ind Crops Prod. 2013;50:882–886. doi: 10.1016/j.indcrop.2013.08.045. [DOI] [Google Scholar]
- 19.Menegol D, Scholl AL, Fontana RC, Dillon AJP, Camassola M. Potential of a Penicillium echinulatum enzymatic complex produced in either submerged or solid-state cultures for enzymatic hydrolysis of elephant grass. Fuel. 2014;133:232–240. doi: 10.1016/j.fuel.2014.05.003. [DOI] [Google Scholar]
- 20.Reis L, Schneider WDH, Fontana R, Camassola M, Dillon AJP. Cellulase and xylanase expression in response to different pH levels of Penicillium echinulatum S1M29 medium. BioEnerg Res. 2014;7:60–67. doi: 10.1007/s12155-013-9345-0. [DOI] [Google Scholar]
- 21.Novello M, Vilasboa J, Schneider WDH, Reis L, Fontana RC, Camassola M. Enzymes for second generation ethanol: exploring new strategies for the use of xylose. RSC Adv. 2014;4(41):21361–21368. doi: 10.1039/c4ra00909f. [DOI] [Google Scholar]
- 22.Schneider WDH, Reis L, Camassola M, Dillon AJP. Morphogenesis and production of enzymes by Penicillium echinulatum in response to different carbon sources. Biomed Res Int. 2014;2014:254863. doi: 10.1155/2014/254863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Liu G, Zhang L, Wei X, Zou G, Qin Y, Ma L, Li J, Zheng H, Wang S, Wang C, Xun L, Zhao GP, Zhou Z, Qu Y. Genomic and Secretomic analyses reveal unique features of the lignocellulolytic enzyme system of Penicillium decumbens. PLoS One. 2013;8(2):e55185. doi: 10.1371/journal.pone.0055185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Marcet-Houben M, Ballester AR, de la Fuente B, Harries E, Marcos JF, Gonzalez-Candelas L, Gabaldon T. Genome sequence of the necrotrophic fungus Penicillium digitatum, the main postharvest pathogen of citrus. BMC Genomics. 2012;21:13–646. doi: 10.1186/1471-2164-13-646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Cantarel BL, Coutinho, PM, Rancurel C, Bernard T, Lombard V, Henrissat B. The carbohydrate-active enzymes database (CAZy): an expert resource for glycogenomics. Nucleic Acids Res. 2009;37:233–38. CAZy database. http://www.cazy.org/. [DOI] [PMC free article] [PubMed]
- 26.Rudnick PA, Wang X, Yan X, Sedransk N, Stein SE. Improved normalization of systematic biases affecting ion current measurements in label-free proteomics data. Mol Cell Proteomics. 2014;13(5):1341–1351. doi: 10.1074/mcp.M113.030593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Jeffries TW. Utilization of xylose by bacteria, yeasts, and fungi. Adv Biochem Eng/Biotechnol. 1983;27:1–32. doi: 10.1007/BFb0009101. [DOI] [PubMed] [Google Scholar]
- 28.Henriksson G, Johansson G, Pettersson G. A critical review of cellobiose dehydrogenases. J Biotechnol. 2000;78(2):93–113. doi: 10.1016/S0168-1656(00)00206-6. [DOI] [PubMed] [Google Scholar]
- 29.Zámocký M, Hallberg M, Ludwig R, Divne C, Haltrich D. Ancestral gene fusion in cellobiose dehydrogenases reflects a specific of GMC oxidoreductases in fungi. Gene. 2004;338(1):1–14. doi: 10.1016/j.gene.2004.04.025. [DOI] [PubMed] [Google Scholar]
- 30.Zampieri D. Gene expression and activities of cellulases, β-glucosidases, xylanases and swollenins of Penicillium echinulatum S1M29. PhD thesis. University of Caxias do Sul, Biotechnology Institute; 2015.
- 31.Juhasz T, Szengyel Z, Reczey K, Siika-Aho M, Viikari L. Characterization of cellulases and hemicellulases produced by Trichoderma reesei on various carbon sources. Process Biochem. 2005;40:3519–3525. doi: 10.1016/j.procbio.2005.03.057. [DOI] [Google Scholar]
- 32.Adav SS, Chao LT, Sze SK. Quantitative secretomic analysis of Trichoderma reesei strains reveals enzymatic composition for lignocellulosic biomass degradation. Mol Cell Proteomics. 2012;11(M111):012419. doi: 10.1074/mcp.M111.012419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Gómez-Mendoza DP, Junqueira M, do Vale FLH, Domont GB, Filho EXF, Sousa MV, Ricart CAO. Secretomic survey of Trichoderma harzianum grown on plant biomass substrates. J Proteome Res. 2014;13(4):1810–1822. doi: 10.1021/pr400971e. [DOI] [PubMed] [Google Scholar]
- 34.Allen AL, Roche CD. Effects of strain and fermentation conditions on production of cellulase by Trichoderma reesei. Biotechnol Bioeng. 1989;33:650–656. doi: 10.1002/bit.260330519. [DOI] [PubMed] [Google Scholar]
- 35.Gritzali MJ, Brown RD. The cellulase system of Trichoderma. The relationship between purified extracellular enzymes from induced or cellulose grown cells. Adv Chem Ser. 1979;181:237–260. doi: 10.1021/ba-1979-0181.ch012. [DOI] [Google Scholar]
- 36.Martins LF, Kolling D, Camassola M, Dillon AJP, Ramos LP. Comparison of Penicillium echinulatum and Trichoderma reesei cellulases in relation to their activity against various cellulosic substrates. Biores Technol. 2007;99:1417–1424. doi: 10.1016/j.biortech.2007.01.060. [DOI] [PubMed] [Google Scholar]
- 37.Sharma M, Roni R, Asiya N, Harinder SO, Bhupinder SC. Evaluation of glycosyl hydrolases in the secretome of Aspergillus fumigatus and saccharification of alkali-treated rice straw. Appl Biochem Biotechnol. 2011;163:557–591. doi: 10.1007/s12010-010-9064-3. [DOI] [PubMed] [Google Scholar]
- 38.Saykhedkar S, Ray A, Ayoubi-Canaan P, Hartson SD, Prade R, Mort AJ. A time course analysis of the extracellular proteome of Aspergillus nidulans growing on sorghum stover. Biotechnol Biofuels. 2012;5:52. doi: 10.1186/1754-6834-5-52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Tsang A, Butler G, Powlowski J, Panisko EA, Baker SE. Analytical and computational approaches to define the Aspergillus niger secretome. Fungal Genet Biol. 2009;46:S153–S160. doi: 10.1016/j.fgb.2008.07.014. [DOI] [PubMed] [Google Scholar]
- 40.Ramos LP, da Silva L, Ballem AC, Pitarelo AP, Chiarello LM, Silveira MHL. Enzymatic hydrolysis of steam-exploded sugarcane bagasse using high total solids and low enzyme loadings. Bioresour Technol. 2015;175:195–202. doi: 10.1016/j.biortech.2014.10.087. [DOI] [PubMed] [Google Scholar]
- 41.Rubini MR, Dillon AJP, Kyaw CM, Faria FP, Poças-Fonseca MJ, Silva-Pereira I. Cloning, characterization and heterologous expression of the first Penicillium echinulatum cellulose gene. J Appl Microbiol. 2009;108:1187–1198. doi: 10.1111/j.1365-2672.2009.04528.x. [DOI] [PubMed] [Google Scholar]
- 42.Vaheri MP, Vaheri MEO, Kauppinen VS. Formation and release of cellulolytic enzymes during growth of Trichoderma reesei on cellobiose and glycerol. Eur J Appl Microbiol Biotechnol. 1979;8:73–80. doi: 10.1007/BF00510268. [DOI] [Google Scholar]
- 43.Fontaine T, Hartlan RP, Diaquin M, Siemel C, Latgé JP. Differential patterns of activity displayed by two exo-beta-1,3-glucanases associated with the Aspergillus fumigatus cell wall. J Bacteriol. 1997;179(10):3154–3163. doi: 10.1128/jb.179.10.3154-3163.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Saloheimo M, Paloheimo M, Hakola S, Pere J, Swanson B. Swollenin, a Trichoderma reesei protein with sequence similarity to the plant expansins, exhibits disruption activity on cellulosic materials. Eur J Biochem. 2002;269:4202–4211. doi: 10.1046/j.1432-1033.2002.03095.x. [DOI] [PubMed] [Google Scholar]
- 45.Jager G, Girfoglio M, Dollo F, Rinaldi R, Bongard H. How recombinant swollenin from Kluyveromyces lactis affects cellulosic substrates and accelerates their hydrolysis. Biotechnol Biofuels. 2011;4:33. doi: 10.1186/1754-6834-4-33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Eijsink VG, Vaaje-Kolstad G, Varum KM, Horn SJ. Towards new enzymes for biofuels: lessons from chitinase research. Trends Biotechnol. 2008;26:228–235. doi: 10.1016/j.tibtech.2008.02.004. [DOI] [PubMed] [Google Scholar]
- 47.Ferreira P, Hernandez-Ortega A, Herguedas B, Martínez ÁT, Medina M. Aryl-alcohol oxidase involved in lignin degradation. J Bio Chem. 2009;284:24840–24847. doi: 10.1074/jbc.M109.011593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Barr DP, Aust SD. Effect of superoxide and superoxide dismutase on lignin peroxidase-catalyzed veratryl alcohol oxidation. Arch Biochem Biophys. 1994;311:378–382. doi: 10.1006/abbi.1994.1251. [DOI] [PubMed] [Google Scholar]
- 49.Adav SS, Ravindran A, Sze SK. Quantitative proteomic analysis of lignocellulolytic enzymes by Phanerochaete chrysosporium on different lignocellulosic biomass. J Proteomics. 2012;75(5):1493–1504. doi: 10.1016/j.jprot.2011.11.020. [DOI] [PubMed] [Google Scholar]
- 50.Blanchette AR, Crueler WE, Height DE, Akhtar M, Akin DE. Cell wall alterations in loblolly pine wood decayed by the white rot fungus Ceriporiopsis subvermispora. J Biotechnol. 1997;53:203–213. doi: 10.1016/S0168-1656(97)01674-X. [DOI] [Google Scholar]
- 51.Henriksson G, Zhang LM, Li JB, Ljungquist P, Reitberger T, Pettersson G. Is cellobiose dehydrogenase from Phanerochaete chrysosporium a lignin degrading enzyme? Biochim Biophys Acta Prot Struct Mol Enzym. 2000;1480:83–91. doi: 10.1016/S0167-4838(00)00096-0. [DOI] [PubMed] [Google Scholar]
- 52.Ander P, Marzullo L. Sugar oxidoreductases and veratryl alcohol oxidase as related to lignin degradation. J Biotechnol. 1997;53:115–131. doi: 10.1016/S0168-1656(97)01680-5. [DOI] [PubMed] [Google Scholar]
- 53.Green MR, Sambrook J. Molecular cloning—a laboratory manual. New York: Cold Spring Harbor Laboratory Press; 2012. [Google Scholar]
- 54.http://support.illumina.com/content/dam/illumina-support/documents/myillumina/f5f619d3-2c4c-489b-80a3 e0414baa4e89/truseq_dna_sampleprep_guide_15026486_c.pdf.
- 55.http://bioinf.uni-greifswald.de/webaugustus/trainingtutorial.gsp.
- 56.Käll L, Storey JD, MacCoss MJ, Noble WS. Assigning significance to peptides identified by tandem mass spectrometry using decoy databases. J Proteome Res. 2008;7:29–34. doi: 10.1021/pr700600n. [DOI] [PubMed] [Google Scholar]
- 57.http://www.ebi.ac.uk/Tools/msa/clustalo/.
- 58.http://blast.ncbi.nlm.nih.gov/Blast.cgi.
- 59.http://csbl.bmb.uga.edu/dbCAN/.
- 60.http://www.cbs.dtu.dk/services/SignalP/.
- 61.http://www.cbs.dtu.dk/services/SecretomeP/.
- 62.http://abi.inf.uni-tuebingen.de/Services/YLoc/webloc.cgi.
- 63.Cota J, Alvarez TM, Citadini AP, Santos CR, Neto MO, Oliveira RR, Pastore GM, Ruller R, Prade RA, Murakami MT, Squina FM. Mode of operation and low-resolution structure of a multi-domain and hyperthermophilic endo-b-1,3-glucanase from Thermotoga petrophila. Biochem Biophys Res Commun. 2011;406:590–594. doi: 10.1016/j.bbrc.2011.02.098. [DOI] [PubMed] [Google Scholar]
- 64.Miller GL. Use of dinitrosalicilic acid reagent for determination of reducing sugar. Anal Chemis. 1959;31:426–428. doi: 10.1021/ac60147a030. [DOI] [Google Scholar]
- 65.Daroit DJ, Simonetti A, Hertz PF, Brandelli A. Purification and characterization of extracellular β-glucosidase from Monascus purpureus. J Microbiol Biotechnol. 2008;18:933–941. [PubMed] [Google Scholar]
- 66.Koseki T, Takahashi K, Handa T, Yamane Y, Fushinobu S, Hashizume K. N-linked oligosaccharides of Aspergillus awamori feruloyl esterase are important for thermostability and catalysis. Biosci Biotechnol Biochem. 2006;70:2476–2480. doi: 10.1271/bbb.60207. [DOI] [PubMed] [Google Scholar]
- 67.Bradford MM. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- 68.Laemmli UK. Cleavage of structural proteins during the assembly of the head bacteriophage T4. Nature. 1970;227:680–685. doi: 10.1038/227680a0. [DOI] [PubMed] [Google Scholar]
- 69.Gonçalves TA, Damásio ARL, Segato F, Alvarez TM, Bragatto J, Brenelli LB, Citadini APS, Murakami MT, Ruller R, Paes Leme AF, Prade RA, Squina FM. Functional characterization and synergic action of fungal xylanase and arabinofuranosidase for production of xylooligosaccharides. Bioresource Technol. 2012;119:293–299. doi: 10.1016/j.biortech.2012.05.062. [DOI] [PubMed] [Google Scholar]