

Abstract
Background. Central intermediary metabolism (CIM) in bacteria is defined as a set of metabolic biochemical reactions within a cell, which is essential for the cell to survive in response to environmental perturbations. The genes associated with CIM are commonly found in both pathogenic and non-pathogenic strains. As these genes are involved in vital metabolic processes of bacteria, we explored the efficiency of the genes in genotypic characterization of Burkholderia pseudomallei isolates, compared with the established pulsed-field gel electrophoresis (PFGE) and multilocus sequence typing (MLST) schemes.
Methods. Nine previously sequenced B. pseudomallei isolates from Malaysia were characterized by PFGE, MLST and CIM genes. The isolates were later compared to the other 39 B. pseudomallei strains, retrieved from GenBank using both MLST and sequence analysis of CIM genes. UniFrac and hierachical clustering analyses were performed using the results generated by both MLST and sequence analysis of CIM genes.
Results. Genetic relatedness of nine Malaysian B. pseudomallei isolates and the other 39 strains was investigated. The nine Malaysian isolates were subtyped into six PFGE profiles, four MLST profiles and five sequence types based on CIM genes alignment. All methods demonstrated the clonality of OB and CB as well as CMS and THE. However, PFGE showed less than 70% similarity between a pair of morphology variants, OS and OB. In contrast, OS was identical to the soil isolate, MARAN. To have a better understanding of the genetic diversity of B. pseudomallei worldwide, we further aligned the sequences of genes used in MLST and genes associated with CIM for the nine Malaysian isolates and 39 B. pseudomallei strains from NCBI database. Overall, based on the CIM genes, the strains were subtyped into 33 profiles where majority of the strains from Asian countries were clustered together. On the other hand, MLST resolved the isolates into 31 profiles which formed three clusters. Hierarchical clustering using UniFrac distance suggested that the isolates from Australia were genetically distinct from the Asian isolates. Nevertheless, statistical significant differences were detected between isolates from Malaysia, Thailand and Australia.
Discussion. Overall, PFGE showed higher discriminative power in clustering the nine Malaysian B. pseudomallei isolates and indicated its suitability for localized epidemiological study. Compared to MLST, CIM genes showed higher resolution in distinguishing those non-related strains and better clustering of strains from different geographical regions. A closer genetic relatedness of Malaysian isolates with all Asian strains in comparison to Australian strains was observed. This finding was supported by UniFrac analysis which resulted in geographical segregation between Australia and the Asian countries.
Keywords: Burkholderia pseudomallei, MLST, Central intermediary metabolism, Genetic variants, Geographical distribution
Introduction
Burkholderia pseudomallei is an environmental saprophytic Gram-negative bacterium that causes melioidosis, a potentially fatal bacterial infection. Infection can be acquired through percutanous inoculation, inhalation or ingestion of either contaminated soil or water (Wiersinga, Currie & Peacock, 2012). Southeast Asia and northern Australia are listed as predominantly endemic for melioidosis (White, 2003; Puthucheary, 2009). Sporadic cases have also been reported in non-endemic regions such as Africa, India and United States (Wiersinga, Currie & Peacock, 2012). The clinical presentations of this disease vary largely among patients, but the most common are pneumonia and septicaemia (Cheng & Currie, 2005). Certain clinical presentations may display region-specific predominance. For instance, genitourinary infections are more prevalent in Australia while hepatosplenic abscesses occur more frequently in Thailand (Cheng & Currie, 2005; Limmathurotsakul & Peacock, 2011). However, to date, no correlation has been reported between the clinical presentations and genotypes, although geographical partitioning between Australian and Asian population of B. pseudomallei has previously been reported (Cheng et al., 2004; Koonpaew et al., 2000)
Analyses on the relatedness among bacterial strains using genotyping methods aid to determine source of outbreak or infection (local epidemiology) and understand the diversity and evolution of a microbial population (global epidemiology) (Carrico et al., 2013). Genotyping mainly distinguishes bacterial strains based on genetic variation resulted from mutation or recombination, and selection pressure on the genome (Urwin & Maiden, 2003). The plethora of genotyping methods can be grouped into two categories; DNA pattern typing and DNA sequence typing (Carrico et al., 2013). Pulsed-field gel electrophoresis (PFGE) is an example of DNA pattern typing method, where bacterial genome is cleaved by restriction enzymes and divided into different lengths of DNA fragments. The DNA fragments are subsequently separated on an agarose gel, which form banding patterns known as pulsotype (Sabat et al., 2013). This method is comparatively inexpensive, highly discriminative and has excellent typeability, but it is tedious, time-consuming and limited in portability (Sabat et al., 2013). PFGE has been extensively used in subtyping of B. pseudomallei strains in Malaysia and high level of diversities was observed. However, based on PFGE, the strains from East Malaysia could not be differentiated from those isolated in Peninsular Malaysia (Chua et al., 2010; Chua et al., 2011).
On the other hand, DNA amplification-based typing such as multilocus sequence typing (MLST) utilizes nucleotide sequences to differentiate strains (Maiden et al., 1998). The unambiguous nature of nucleotide sequences allows interlaboratory comparisons to be performed and thus overcome the portability issue of other molecular typings (Maiden et al., 1998; Sabat et al., 2013). The MLST scheme utilizes seven housekeeping genes that are sequenced, combined and assigned to a sequence type (ST) according to the allelic profiles deposited on MLST.net (http://bpseudomallei.mlst.net/) (Godoy et al., 2003). Housekeeping genes are selected as gene markers in MLST since genetic variation accumulates slowly for metabolic function, thus making it useful for long-term epidemiology monitoring (Maiden et al., 1998; Urwin & Maiden, 2003). Assuming that gene mutations of the housekeeping genes are under stabilizing selection, then the clonality of the species is conserved (Maiden et al., 1998; Urwin & Maiden, 2003). However, it is not suitable for local surveillance of some pathogens as it is insufficiently discriminative (Urwin & Maiden, 2003; Sabat et al., 2013). Modifications to the MLST schemes have been proposed to improve the discriminatory or resolution power of the “conventional” MLST scheme. The MLST schemes of Neisseria meningitidis and Salmonella enterica demonstrated better discriminatory power with the addition of fumC gene, and a combination of two housekeeping (gyrB and atpD) and flagellin (fliC and fliB) genes, respectively (Holmes, Urwin & Maiden, 1999; Tankouo-Sandjong et al., 2007).
Central intermediary metabolism (CIM) in bacteria is important as it plays an essential role for the cell to grow, survive and response to environmental perturbations. In living cells, thousands of different biochemical reactions and transport processes are linked together to generate constant supply of energy and to maintain their biological functions (Spector, 2009; Shimizu, 2013). The genes associated with CIM are housekeeping genes that encode metabolic functions, hence CIM genes are considered good molecular markers for the study of phylogenetic relationship at species level (Naharro, Rubio & Luengo, 2011). In this study, we tested the feasibility of the CIM genes as alternative approach for strains characterization. The genetic relatedness of the strains that were previously deposited in the National Center for Biotechnology Information (NCBI) Genbank (http://www.ncbi.nlm.nih.gov/genome/476) was determined based on MLST and the genes related to CIM. This study may provide better insight into the genetic diversity of B. pseudomallei and epidemiological distribution of B. pseudomallei population in Malaysia and in comparison to other countries.
Methods
Genomic sequences and bacterial isolates
Nineteen complete and 29 draft genomes of B. pseudomallei were selected from NCBI Genbank (http://www.ncbi.nlm.nih.gov/genbank/) based upon the availability of background information (e.g., source, year of isolation, country) in order to determine geographical relatedness. Among the 48 genomes selected, nine of them were from Malaysia (Table 1).
Table 1. Background information and MLST analysis of B. pseudomallei genomes used in this study.
| Accession number | Strain | Source | Country | Year | ST | Allele profile | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ace | gltB | gmhD | lepA | lipA | narK | ndh | ||||||
| AGVS01 | 354a | Sputum | Thailand | 1988 | 78 | 1 | 2 | 2 | 1 | 1 | 1 | 1 |
| AHJD01 | 354ea | Sputum | Thailand | 1994 | 78 | 1 | 2 | 2 | 1 | 1 | 1 | 1 |
| AAMM02 | 406e | Toe swab | Thailand | 1988 | 211 | 3 | 1 | 3 | 1 | 1 | 4 | 1 |
| NZCP008777 | 576 | Human | Thailand | 1989 | 501 | 1 | 12 | 6 | 2 | 1 | 2 | 3 |
| AHJA01 | 1026a | Pus | Thailand | 1993 | 102 | 3 | 4 | 12 | 1 | 1 | 4 | 1 |
| NC017851 | 1026b | Blood | Thailand | 1993 | 102 | 3 | 4 | 12 | 1 | 1 | 4 | 1 |
| AAMB02 | 1106b | Pus | Thailand | 1996 | 70 | 3 | 4 | 11 | 3 | 5 | 4 | 6 |
| AHJB01 | 1258a | Sputum | Thailand | 1994 | 221 | 1 | 12 | 2 | 3 | 5 | 29 | 1 |
| AHJC01 | 1258bb | Blood | Thailand | 1995 | 221 | 1 | 12 | 2 | 3 | 5 | 29 | 1 |
| ABBN01 | B7210 | Pus | SE Asia | – | 84 | 3 | 1 | 11 | 4 | 5 | 4 | 6 |
| JNOW01 | BDD | Blood | Australia | – | 956 | 1 | 2 | 13 | 2 | 1 | 45 | 11 |
| JPNW01 | BDE | Sputum | Australia | – | 140 | 4 | 7 | 3 | 4 | 1 | 19 | 1 |
| JPNU01 | BDI | Blood | Australia | – | 252 | 1 | 4 | 3 | 2 | 6 | 19 | 1 |
| NZCP009209 | BDP | Brain | Australia | 1994 | 36 | 1 | 7 | 14 | 7 | 1 | 12 | 11 |
| JOTS01 | BDT | Joint aspirate | Australia | – | 958 | 1 | 2 | 14 | 22 | 1 | 8 | 1 |
| JPNP01 | BEB | Blood | Australia | – | 254 | 1 | 4 | 3 | 2 | 2 | 34 | 1 |
| JPNQ01 | BED | Blood | Australia | – | 24 | 1 | 2 | 13 | 2 | 1 | 8 | 1 |
| BEJ | – | Thailand | – | 15 | 1 | 2 | 2 | 2 | 1 | 3 | 1 | |
| BEM | – | Vietnam | – | 411 | 1 | 4 | 4 | 3 | 1 | 3 | 1 | |
| BEX | – | Thailand | – | 501 | 1 | 12 | 6 | 2 | 1 | 2 | 3 | |
| BFD | Blood | Australia | – | 445 | 8 | 4 | 13 | 2 | 1 | 8 | 1 | |
| BGR | – | Thailand | – | 102 | 3 | 4 | 12 | 1 | 1 | 4 | 1 | |
| NC018527 | BPC006 | Blood | China | 2008 | 70 | 3 | 4 | 11 | 3 | 5 | 4 | 6 |
| APLM01 | CB | Blood | Malaysia | – | 46 | 3 | 1 | 2 | 1 | 1 | 3 | 3 |
| APLH01 | CMS | Blood | Malaysia | – | 54 | 3 | 1 | 3 | 3 | 1 | 2 | 1 |
| APLN01 | CSc | Blood | Malaysia | – | 46 | 3 | 1 | 2 | 1 | 1 | 3 | 3 |
| AVAL01 | HBPUB10134a | Sputum | Thailand | 2010 | 228 | 1 | 2 | 3 | 1 | 1 | 4 | 1 |
| AVAM01 | HBPUB10303a | Sputum | Thailand | 2011 | 48 | 3 | 1 | 2 | 1 | 1 | 4 | 1 |
| APLG01 | LMF | Sputum | Malaysia | – | 289 | 3 | 4 | 11 | 4 | 5 | 4 | 6 |
| NZCP008781 | Mahidol-1106a | – | Thailand | – | 70 | 3 | 4 | 11 | 3 | 5 | 4 | 6 |
| APHN01 | MARAN | Soil | Malaysia | – | 1342 | 4 | 12 | 3 | 2 | 1 | 1 | 3 |
| NZCP004042 | MSHR146 | Goat | Australia | 1992 | 617 | 1 | 16 | 3 | 4 | 2 | 8 | 1 |
| NC021877 | MSHR305 | Brain | Australia | 1994 | 36 | 1 | 7 | 14 | 7 | 1 | 12 | 11 |
| NZCP008764 | MSHR346 | Sputum | Australia | 1995 | 243 | 1 | 2 | 13 | 4 | 15 | 12 | 1 |
| NZCP004023 | MSHR511 | Goat | Australia | 1997 | 617 | 1 | 16 | 3 | 4 | 2 | 8 | 1 |
| NZCP004368 | MSHR520 | Blood | Australia | 1998 | 36 | 1 | 7 | 14 | 7 | 1 | 12 | 11 |
| AVAJ01 | MSHR5848 | Sputum | Australia | 2011 | 553 | 1 | 1 | 3 | 4 | 30 | 2 | 30 |
| JMMV01 | MSHR5855 | Sputum | Australia | 2011 | 553 | 1 | 1 | 3 | 4 | 30 | 2 | 30 |
| AVAK01 | MSHR5858 | Sputum | Australia | 2011 | 562 | 1 | 1 | 4 | 1 | 1 | 29 | 1 |
| AXDS01 | MSHR6137 | Water | Australia | 2012 | 325 | 1 | 1 | 13 | 4 | 15 | 3 | 11 |
| NZCP004003 | NAU20B-16 | Soil | Australia | 2006 | 617 | 1 | 16 | 3 | 4 | 2 | 8 | 1 |
| NZCP004001 | NCTC 13178 | Brain | Australia | – | 286 | 15 | 2 | 13 | 2 | 20 | 19 | 1 |
| NC022659 | NCTC 13179 | Skin ulcer | Australia | – | 613 | 1 | 4 | 43 | 4 | 1 | 48 | 11 |
| APLK01 | OB | Blood | Malaysia | – | 46 | 3 | 1 | 2 | 1 | 1 | 3 | 3 |
| APLL01 | OSd | Blood | Malaysia | – | 46 | 3 | 1 | 2 | 1 | 1 | 3 | 3 |
| ACKA01 | Pakistan 9 | Human | Pakistan | 1988 | 72 | 4 | 1 | 4 | 1 | 1 | 2 | 1 |
| APLJ01 | THE | Spleen | Malaysia | – | 54 | 3 | 1 | 3 | 3 | 1 | 2 | 1 |
| APLI01 | VEL | Sputum | Malaysia | – | 46 | 3 | 1 | 2 | 1 | 1 | 3 | 3 |
Notes.
354e is a relapse case of 354a.
1258b is a relapse case of 1258a.
CS is the small colony variant of CB.
OS is the small colony variant of OB.
Pulsed-field gel electrophoresis
The nine Malaysian isolates were characterized by PFGE analysis. PFGE preparation was performed with a few modifications based on the CDC One-Day Standard Laboratory Protocol for Molecular Subtyping (Ribot et al., 2006). In brief, B. pseudomallei isolates were grown overnight on nutrient agar plates at 37 °C. The bacterial cells were suspended in normal saline solution and adjusted to a concentration of OD600 = 0.6–0.8 using Dade Behring Microscan Turbidity Meter (Siemens, Germany). The suspensions were mixed with 1.0% (w/v) agarose gel (Seakem Gold Agarose, Rockland, ME, USA) to prepare plugs. The plugs were lysed overnight at 50 °C before they were digested with SpeI (Promega, Wisconsin, USA) for an additional 2 h (50 °C). The digested DNA fragments were then loaded into a 1.0% (w/v) agarose gel and separated by electrophoresis using the CHEF Mapper apparatus (Bio-Rad Laboratories, Hercules, CA, USA) as previously described (Chua et al., 2011). The gel was visualized under ultraviolet light using the Gel Doc 2000 system (Bio-Rad Laboratories, Hercules, CA, USA). Analysis was performed using the Bionumerics 6.0 software (Applied Maths, Sint-Martens-Latem, Belgium) to construct a Dice coefficient dendrogram based on the unweighted pair group method with arithmetics average (UPGMA) algorithm.
Multilocus sequence typing (MLST)
In silico MLST analyses were performed by aligning B. pseudomallei genomes against the sequences of seven alleles of B. pseudomallei K96243 (accession number BX571965) retrieved from B. pseudomallei MLST database (http://bpseudomallei.mlst.net/) using basic local alignment search tool (BLASTN) (https://blast.ncbi.nlm.nih.gov/).The alignment was downloaded and the sequence for each allele (gene) was submitted to the B. pseudomallei MLST database to assign the allelic numbers (Godoy et al., 2003). A combination of the seven allelic numbers by the order of ace-gltB-gmhD-lepA-lipA-narK-ndh was compared against the B. pseudomallei MLST database to obtain a ST number (Godoy et al., 2003).
Genetic characterization based on central intermediary metabolism (CIM) genes
Seventy CIM gene sequences of K96243 (accession number BX571965) were obtained from the Burkholderia Genome Database (http://beta.burkholderia.com/)(Holden et al., 2004; Winsor et al., 2008). B. pseudomallei genomes were aligned against the CIM gene sequences using basic local alignment search tool (BLASTN) (https://blast.ncbi.nlm.nih.gov/). Twelve CIM genes were finally selected as only these genes showed 100% query coverage across all 48 strains.
Maximum likelihood analysis
Phylogenetic trees for the nine isolates from Malaysia and 48 strains worldwide were constructed using the concatenated sequence of MLST housekeeping genes and CIM genes respectively, based on the maximum likelihood method. The seven genes of MLST and 12 CIM genes were concatenated in order of ace-gltB-gmhD-lepA-lipA-narK-ndh and BPSL0512-BPSL0958–BPSL1410–BPSL1418–BPSL1668–BPSL2657–BPSL2658–BPSL2659–BPSL2662–BPSL2837–BPSS0626–BPSS1635, respectively. The trees were constructed using MEGA6.06 software (Molecular Evolutionary Genetics Analysis software, Arizona State University, Tempe, AZ, USA) with 1,000 bootstrap replications (Tamura & Nei, 1993; Tamura et al., 2013).
Discriminatory index
The discriminatory index (D) of MLST and sequence analysis of CIM genes were calculated using the formula of Simpson’s index of diversity (Hunter & Gaston, 1988). An index of 1.0 indicated that the typing method was able to differentiate each member of a strain population as unique. On the other hand, D value of 0 indicated that all members of a strain population were identical.
Comparison of CIM, MLST and WGS
Phylogenetic relationship of 15 randomly selected B. pseudomallei genomes was established based on Tetra-nucleotide signature correlation index calculated from JSpecies Web Server (http://jspecies.ribohost.com/jspeciesws) (Richter et al., 2015). The server allowed a maximum of fifteen genomes to be compared per session. The correlation matrix was matched with the maximum likelihood distance matrices produced from the corresponding concatenated CIM and MLST genes using Spearman Rank Correlation (ρ). Significant correlation was calculated based on 1,000 permutations.
Unweighted UniFrac analysis
Unweighted Unifrac analysis (Lozupone, Hamady & Knight, 2006) was conducted using Mothur (ver 1.36.1) (Schloss et al., 2009) to identify significant differences in geographical distribution. In brief, the distance matrix (shared distance between strains) was calculated based on individual maximum likelihood trees generated from MLST and CIM genes analyses. The significance was calculated through Monte Carlo permutation with 1000 times resampling. The relationship between these geographical locations was illustrated using hierarchical clustering under Ward’s method. The calculation was conducted using “hcluster” command under R programme.
Results
Selection and diversity of central intermediary metabolism genes
A total of 12 CIM genes (100% coverage) were selected for the analysis and these genes were present in all the 48 B. pseudomallei genomes selected from the NCBI GenBank database (Table 2). The ratio of polymorphic sites in the CIM genes ranged from 0.9% to 2.5%. (Table 2)
Table 2. Properties of the 12 selected CIM genes.
| Locus tag | Gene name | Product name | Size for analysis (bp) | No. of alleles | No of variable sites (%) | Genome location (kb) |
|---|---|---|---|---|---|---|
| BPSL 0512 | – | Nitrite reductase | 2445 | 25 | 34 (1.4) | 564 |
| BPSL 0958 | cysH | Phosphoadenosine phosphosulfate reductase | 750 | 15 | 15 (2.0) | 1,116 |
| BPSL 1410 | – | Peptidyl-prolyl cis-trans isomerase | 1935 | 22 | 30 (1.6) | 1,640 |
| BPSL 1418 | – | Exported isomerase | 780 | 12 | 13 (1.7) | 1,654 |
| BPSL 1668 | – | Adenylylsulfate kinase | 852 | 18 | 21 (2.5) | 1,946 |
| BPSL 2657 | ureA | Urease subunit gamma | 303 | 5 | 4 (1.3) | 3,181 |
| BPSL 2658 | ureB | Urease subunit beta | 306 | 9 | 7 (2.3) | 3,181 |
| BPSL 2659 | ureC | Urease subunit alpha | 1707 | 27 | 34 (2.0) | 3,182 |
| BPSL 2662 | ureG | Urease accessory protein | 651 | 14 | 12 (1.8) | 3,184 |
| BPSL 2837 | – | Carbon-nitrogen hydrolase | 828 | 13 | 10 (1.2) | 3,396 |
| BPSS 0626 | – | Luciferase-like monooxygenase | 1095 | 9 | 10 (0.9) | 854 |
| BPSS 1635 | – | Class III aminotransferase | 2787 | 26 | 60 (2.2) | 2,248 |
Relationship between nine Malaysian B. pseudomallei isolates
The relatedness of nine Malaysian B. pseudomallei strains was determined based on PFGE, MLST and sequence analysis of CIM genes (Fig. 1). Based on PFGE analysis (Fig. 1A), these nine Malaysian strains were resolved into six pulsotypes. Two clusters were observed at 75% similarity with the exclusion of the B. pseudomallei K96243. Interestingly, the similarity level between morphology variants, OS/OB and CS/CB was only 67%, and 95%, respectively. In contrast, two strains, OS (clinical strain) and MARAN (soil strain) have an identical pulsotype despite being isolated from different sources. Both of the isolates were found to be distinct from the rest of the strains. In addition, several other isolates were also sharing identical pulsotypes, namely, CB and OB; CMS and THE. On the other hand, four profiles were observed based on MLST analysis and five profiles were generated based on the CIM genes. However, the phylogenetic trees generated by both sequence-based analyses showed high similarity of clustering pattern, where seven isolates were grouped into one cluster while MARAN and LMF were categorized as outgroups. The morphology variant, OS and OB, which shared similar ST were distinguished based on a single nucleotide polymorphism (SNP) observed in one of the CIM genes, BPSL 0512.
Figure 1. Comparison between PFGE, CIM genes and MLST profiles of nine Burkholderia pseudomallei isolates from Malaysia.
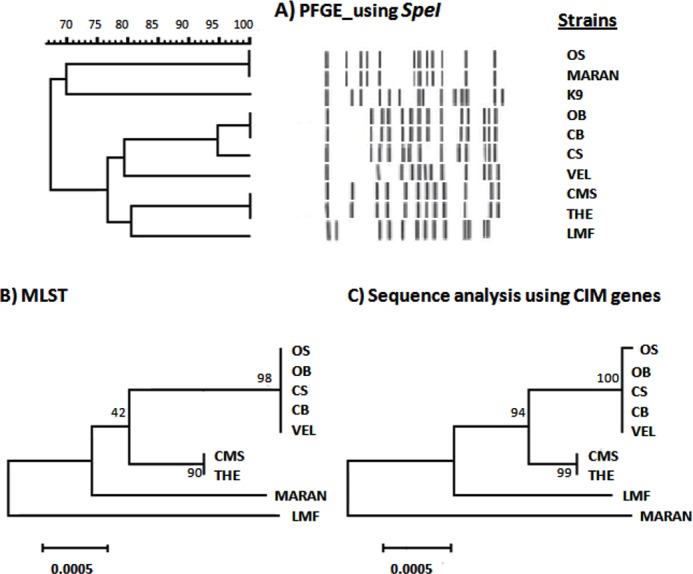
(A) Dendrogram generated using UPGMA based on Dice coefficients showed two clusters at 65% similarity. K9 was the B. pseudomallei reference strain, K96243. (B) Maximum likelihood tree generated using 7 MLST genes showed the close relationship of the colony variants (OB, OS; CB, CS) with the other clinical isolates and LMF and MARAN as outgroups. (C) The relationship of the Malaysian isolates were also depicted using maximum likelihood tree based on sequence analysis of CIM genes. The tree showed the slight difference of OS with its big colony variant, OB. The values on the nodes indicate the bootstrap value with 1,000 times resampling for (B) and (C).
Relationship between B. pseudomallei isolates in Malaysia and other countries
MLST analysis revealed a novel allelic profile of 4-12-3-2-1-1-3 for MARAN and was assigned with ST1342. Based on the maximum likelihood tree of the seven housekeeping genes (Fig. 2), the 48 B. pseudomallei genomes were resolved into 31 profiles and divided into three main clusters (cluster I, II and III). Cluster II and III were mostly comprised by strains from Asian countries, except for MSHR5858 (Australia); while Cluster I was comprised of a mixture of Australian and Asian strains. The eight Malaysian strains were observed in cluster I and majority of the strains were more related to strains from Thailand. Only one strain, LMF was observed in cluster III and was also related to strains from Thailand, China and Southeast Asia. .
Figure 2. Maximum likelihood tree of MLST displaying the relatedness of the 48 B. pseudomallei isolates from endemic countries.
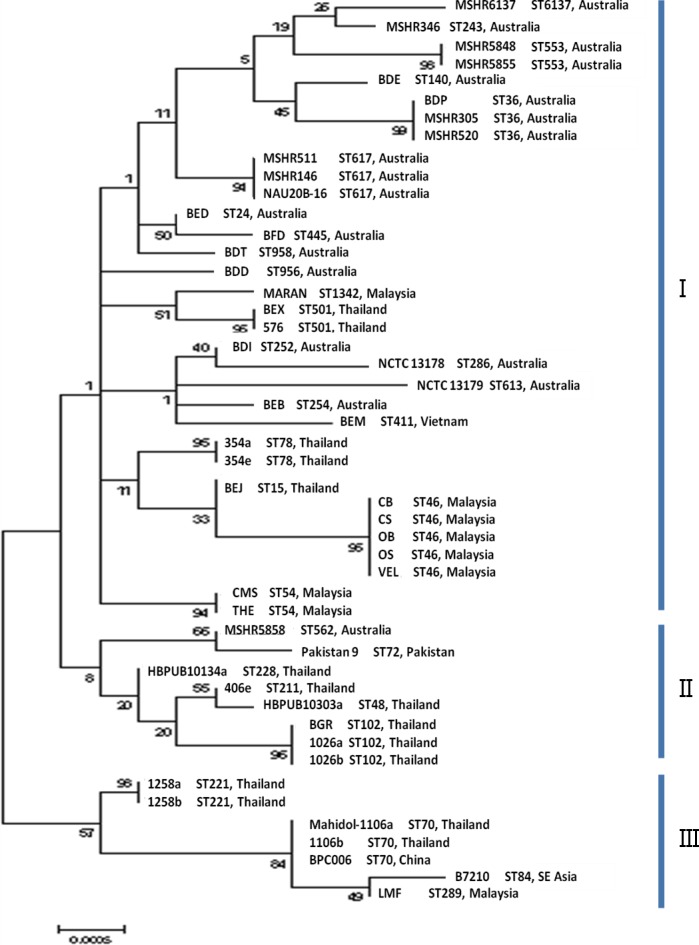
The sequence types and countries were indicated for comparison. Three clusters were identified and labeled as cluster I, II and III. The values on the nodes indicate the bootstrap value with 1,000 times resampling.
Based on the maximum likelihood tree generated using CIM genes (Fig. 3), the 48 strains were grouped into 33 profiles and four clusters (I, II, III and IV). All strains in cluster I were from the Asian countries and all the strains with identical profiles were also found to be of the same STs. Among the nine Malaysian strains, MARAN was the most distinctive as it was clustered in cluster II with three strains from Australia (MSHR305, MSHR520 and BDP) and 2 strains from Thailand (BEX and 576). Strains isolated from Australia showed higher diversity as they were distributed across cluster II, III and IV. Overall, discriminatory (D) index of sequence analysis of CIM genes (D = 0.9814) is slightly higher than MLST (D = 0.9761).
Figure 3. Maximum likelihood tree of sequence analysis of CIM genes displaying the relatedness of the 48 B. pseudomallei isolates from endemic countries.
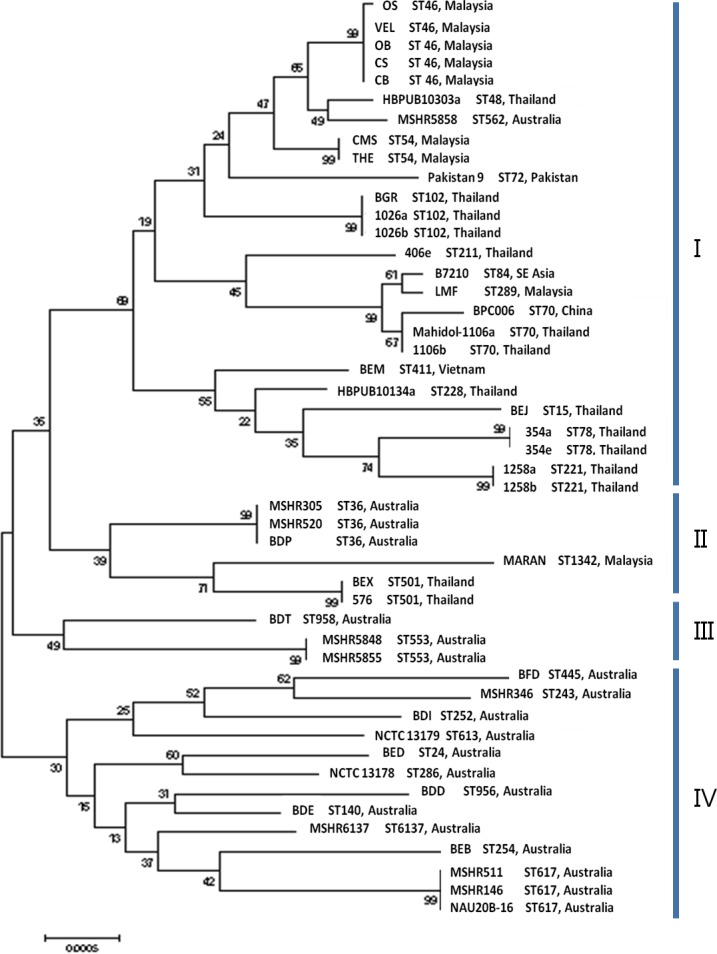
The sequence types and countries were indicated for comparison. Cluster A is divided into two subclusters (AI and AII) and mostly occupied by isolates from Asia. Cluster B and C comprised of mostly isolates from Australia. The values on the nodes indicate the bootstrap value with 1000 times resampling.
Comparing the phylogeny inferred from CIM and MLST with WGS
Pairwise genetic distance of fifteen randomly selected isolates inferred from both CIM and MLST was compared with tetra correlation index of WGS using Spearman Rank Correlation. A moderate but significant correlation was established between CIM and MLST (ρ = 0.42, P = 0.002). However, distance matrices of both CIM and MLST showed no correlation to the phylogeny established with WGS (ρ = − 0.06–0.04, P > 0.25).
Geographical relationship among B. pseudomallei variants
UniFrac analysis was performed to determine the geographical relationship between the studied strains. Based on both MLST and sequence analysis of CIM genes, significant (P ≤ 0.01) geographical clustering was observed (Table 3). For instance, B. pseudomallei strains from Australia were found to be distinct from the strains obtained from the Asian countries (P ≤ 0.05). Single linkage hierarchical clustering based on Unifrac distances of MLST and CIM analyses produced similar “geographical structure.” However, CIM exhibited an overall higher resolution in discerning isolates from different region in comparison to MLST (Fig. 4). For instance, the distance between Malaysian and Thailand strains is approximately 0.6 in MLST but >0.7 in CIM.
Table 3. Pairwise comparison of UniFrac distances based on (A) CIMT and (B) MLST.
Statistical significance was calculated using Monte Carlo permutation with 1,000 iterations. Signifance results (P ≤ 0.05) are red-color-coded.
| A | Australia | Malaysia | Others | Thailand |
|---|---|---|---|---|
| Australia | 0.002 | 0.103 | <0.001 | |
| Malaysia | 0.204 | 0.014 | ||
| Others | 0.173 | |||
| Thailand |
Figure 4. Single linkage hierachical clustering generated based on Unifrac distances of (A) MLST and (B) CIM genes to associate the relatedness according to countries.
Genetical distances of B. pseudomallei isolates inferred from both MLST and CIM produced similar “geographical structure.” However, CIM exhibited an overall higher resolution in discerning isolates from different region in comparison to MLST.
Discussions
CIM is important in bacteria as it plays an essential role for the cell to grow, survive and response to environment perturbation. The genes associated with CIM are commonly found in both pathogenic and non-pathogenic strains. The genes are under stabilizing selection and usually species-specific (Spector, 2009). In this study, a higher D index for the alignment of CIM gene was observed compared to MLST. The slightly higher D index of the sequence analysis of CIM genes is not significant due to the small sample size (N < 100) and might be attributed to the use of 12 full-length genes (303–2787 bp) as opposed to the 7 internal fragments of housekeeping genes in MLST (300–500 bp). Increased numbers of sequence lengths have shown an increase in discriminatory power (Maiden et al., 1998; Godoy et al., 2003; Urwin & Maiden, 2003). Nonetheless, the finding in this study may facilitate further study to assess the variable regions within each CIM sequence and translate them into viable genotyping method. Although WGS is gradually being used as a tool for strains identification and characterization, it is still unlikely to be cost effective for developing countries. Moreover, the laboratory personnel must acquire skills to handle new generation sequencing technology and interpret cumbersome WGS data (Aliyu, 2014).
A direct comparison of the three typing methods for characterization of the Malaysian strains showed that the gel-based PFGE method was more discriminative compared to the other two sequence-based methods (MLST and sequence analysis of CIM genes). This finding concurred with previous studies that reported different PFGE banding patterns can be observed from a single ST (Godoy et al., 2003; Cheng et al., 2004; Chantratita et al., 2008). In general, MLST and CIM genes analyses involved essential metabolic genes that are highly conserved. Thus, these methods are unaffected by genomic rearrangement, whereas PFGE that cleave bacterial genome at restriction sites would produce a different banding pattern if there is any large insertion or deletion along the genomic sequence (Chantratita et al., 2008; Cooper & Feil, 2004). Another possible reason would be the longer history of mutation accumulation and history of lateral gene transfer in B. pseudomallei, which leads to homoplasy and apparent homoplasy, respectively, in unrelated isolates (Pearson et al., 2009). For instance, substantial diversity within strains from Australia and Cambodia that shared same STs due to homoplasy had been reported by De Smet et al. (2015). This also indicates the limitation of MLST for characterizing highly recombinogenic species. Overall, our study suggested that PFGE is more suitable for localized epidemiology as it is affected by genomic rearrangements while the sequence-based analyses are more suitable for cross geographical or long-term epidemiological study as previously indicated by Maiden et al. (1998).
In this study, a novel allelic profile (ST1342) was assigned to MARAN, the only soil isolate from Malaysia. Interestingly, PFGE showed that this strain was identical to OS, a clinical isolate and the morphology variant of OB. According to MLST analysis, OS was identified as ST46 and was genetically similar to the other strains of ST46 such as OB, CB, CS and VEL. However, CIM genes analysis revealed a SNP different between OS and other strains of ST46 and indicated that there was a lack of relatedness between OS and the morphology variant, OB as demonstrated by PFGE. To investigate the genetic relatedness of OS, OB and MARAN, further analysis such as WGS will be performed.
On the other hand, both MLST and CIM genes analyses managed to identify the isolates associated with two relapse cases (354a & 354e; 1258a & 1258b), despite the 330 Kb deletion on chromosome 2 of 1258a and 800 Kb inversion on chromosome 2 of 354e that were previously reported by Hayden et al. (2012). In this case, MLST was not affected by the mutations mostly because all the MLST loci are located along chromosome 1. Sequence analysis comprised of two CIM genes on chromosome 2 (BPSS0626 and BPSS1635) yet the two pair isolates were resolved as clonal.
To determine whether the CIM markers are better at recapitulating the whole genome phylogeny compared to the MLST markers, the phylogenetic relationship of 15 randomly selected B. pseudomallei genomes has been determined using the online server. Based on the 15 selected B. pseudomallei strains, distance matrices of both CIM and MLST showed no correlation to the phylogeny established with WGS but a significant correlation was established between CIM and MLST. Overall, CIM genes analysis showed higher resolution in depicting the relationship of the related and non-related strains compared to MLST. For instance, MLST identified two strains from Thailand and one strain from China as ST46; however CIM genes analysis managed to distinguish the strain from China from the other two strains from Thailand. Based on MLST analysis, the strains from Australia and Asian countries are grouped in one big cluster but clustering analysis using CIM genes showed that the strains from Asian were genetically related as they were clustered in cluster I, whereas the strains from Australia were more diversified as they were distributed across clusters II, III and IV (Cheng et al., 2008; Pearson et al., 2009).
UniFrac analysis revealed strong geographical segregation between strains from Australia and the Asian countries from both sequence-based analyses. This finding concurred with previous studies which reported that STs from Australia were distinctly different from those from Southeast Asian countries (Cheng et al., 2004; Chantratita et al., 2008). Such a distribution might be due to the geographical isolation of Australia from the continent of Asia (Torpdahl et al., 2005; Hayden et al., 2012). Nevertheless, a few STs were found in both Australia and Southeast Asia since the increased number of human migrations between the two regions has been observed (McCombie, Finkelstein & Woods, 2006). Melioidosis cases have been reported on military personnel and travelers who traveled to endemic countries such as Thailand, Vietnam and Australia (Limmathurotsakul & Peacock, 2011; Wiersinga, Currie & Peacock, 2012). The migration of animals across a land bridge connecting Australia and Southeast Asia during the Miocene period has also resulted in common genotype shared by both regions (Cheng et al., 2004).
In addition, extensive studies have been conducted to support the phylogeographic partitioning between Australian and Asian B. pseudomallei isolates. Pearson et al. (2009) postulated that B. pseudomallei was introduced to Southeast Asia following collision of these two continents in the vicinity of Wallace’s Line after the late Miocene period. Another possible dispersal route is through Papua New Guinea since the island is located between the two endemic regions of Northern Australia and Southeast Asia and the strains from the island showed shared characteristics of both regions (Baker et al., 2011). Based on the study of Tuanyok et al. (2007), Australian and Asian B. pseudomallei populations can be genetically differentiated by a Yersinia-like fimbrial (YLF) gene cluster, predominantly found in the Asian population, while a B. thailandensis-like flagellum and chemotaxis (BTFC) gene cluster is mainly found in Australian population. However, comparison based on SNPs from WGS inferred that the isolates from the two continents are not separated as a result of uneven geographic sampling. Nevertheless, homoplasy should be taken into consideration when phylogenetic relatedness of B. pseudomallei strains from different regions is being studied based on MLST as it could be due to recombination and lateral gene transfer that resulted in similar characteristics of unrelated samples (Pearson et al., 2009).
Conclusions
Genetic relatedness of strains isolated from Malaysia and other countries have been determined and a significant geographical segregation of B. pseudomallei genomes between Australia and Asian countries were observed. Overall, the sequence analysis of CIM genes is comparable to MLST and may be useful to be developed into genetic markers.
Acknowledgments
We thank Ms. Soo Tein Ngoi from the Institute of Biological Sciences, Faculty of Science, University of Malaya, Kuala Lumpur, Malaysia, for technical assistance in PFGE.
Funding Statement
This research was supported by the University of Malaya Research Grant (UMRG: RP013B-13HTM), University of Malaya Postgraduate Research Grant (PPP: PG165-2015A) and University of Malaya-Ministry of Education (UM-MoE) High Impact Research (HIR) Grant (UM.C/625/1/HIR/MoE/CHAN/02: H-50001-A000013). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Additional Information and Declarations
Competing Interests
The authors declare there are no competing interests.
Author Contributions
Noorfatin Jihan Zulkefli conceived and designed the experiments, performed the experiments, analyzed the data, wrote the paper, prepared figures and/or tables, reviewed drafts of the paper.
Vanitha Mariappan and Kumutha Malar Vellasamy performed the experiments, contributed reagents/materials/analysis tools, wrote the paper, reviewed drafts of the paper.
Chun Wie Chong performed the experiments, analyzed the data, contributed reagents/materials/analysis tools, reviewed drafts of the paper.
Kwai Lin Thong analyzed the data, contributed reagents/materials/analysis tools, reviewed drafts of the paper.
Sasheela Ponnampalavanar analyzed the data, reviewed drafts of the paper.
Jamuna Vadivelu contributed reagents/materials/analysis tools, reviewed drafts of the paper.
Cindy Shuan Ju Teh conceived and designed the experiments, contributed reagents/materials/analysis tools, wrote the paper, prepared figures and/or tables, reviewed drafts of the paper.
DNA Deposition
The following information was supplied regarding the deposition of DNA sequences:
Data Availability
The following information was supplied regarding data availability:
The research in this article did not generate any raw data.
References
- Aliyu (2014).Aliyu S. Bacterial whole genome sequencing: the future of clinical bacteriology. Annals of Nigerian Medicine. 2014;8(2):51–57. doi: 10.4103/0331-3131.153352. [DOI] [Google Scholar]
- Baker et al. (2011).Baker A, Pearson T, Price EP, Dale J, Keim P, Hornstra H, Greenhill A, Padilla G, Warner J. Molecular phylogeny of Burkholderia pseudomallei from a remote region of Papua New Guinea. PLoS ONE. 2011;6:e1802. doi: 10.1371/journal.pone.0018343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carrico et al. (2013).Carrico JA, Sabat AJ, Friedrich AW, Ramirez M, Markers ESGfE. Bioinformatics in bacterial molecular epidemiology and public health: databases, tools and the next-generation sequencing revolution. Eurosurveillance. 2013;18:20382–20391. doi: 10.2807/ese.18.04.20382-en. [DOI] [PubMed] [Google Scholar]
- Chantratita et al. (2008).Chantratita N, Wuthiekanun V, Limmathurotsakul D, Vesaratchavest M, Thanwisai A, Amornchai P, Tumapa S, Feil EJ, Day NP, Peacock SJ. Genetic diversity and microevolution of Burkholderia pseudomallei in the environment. PLoS Neglected Tropical Diseases. 2008;2:e1802. doi: 10.1371/journal.pntd.0000182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheng & Currie (2005).Cheng AC, Currie BJ. Melioidosis: epidemiology, pathophysiology, and management. Clinical Microbiology Reviews. 2005;18:383–416. doi: 10.1128/CMR.18.2.383-416.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheng et al. (2004).Cheng AC, Godoy D, Mayo M, Gal D, Spratt BG, Currie BJ. Isolates of Burkholderia pseudomallei from Northern Australia are distinct by multilocus sequence typing, but strain types do not correlate with clinical presentation. Journal of Clinical Microbiology. 2004;42:5477–5483. doi: 10.1128/JCM.42.12.5477-5483.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheng et al. (2008).Cheng AC, Ward L, Godoy D, Norton R, Mayo M, Gal D, Spratt BG, Currie BJ. Genetic diversity of Burkholderia pseudomallei isolates in Australia. Journal of Clinical Microbiology. 2008;46:249–254. doi: 10.1128/JCM.01725-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chua et al. (2010).Chua KH, See KH, Thong KL, Puthucheary SD. DNA fingerprinting of human isolates of Burkholderia pseudomallei from different geographical regions of Malaysia. Tropical Biomedicine. 2010;27:517–524. [PubMed] [Google Scholar]
- Chua et al. (2011).Chua KH, See KH, Thong KL, Puthucheary SD. SpeI restriction enzyme displays greater discriminatory power than XbaI enzyme does in a pulsed-field gel electrophoresis study on 146 clinical Burkholderia pseudomallei isolates. Japanese Journal of Infectious Diseases. 2011;64:228–233. [PubMed] [Google Scholar]
- Cooper & Feil (2004).Cooper JE, Feil EJ. Multilocus sequence typing–what is resolved? Trends in Microbiology. 2004;12:373–377. doi: 10.1016/j.tim.2004.06.003. [DOI] [PubMed] [Google Scholar]
- De Smet et al. (2015).De Smet B, Sarovich DS, Price EP, Mayo M, Theobald V, Kham C, Heng S, Thong P, Holden MT, Parkhill J, Peacock SJ, Spratt BG, Jacobs JA, Vandamme P, Currie BJ. Whole-genome sequencing confirms that Burkholderia pseudomallei multilocus sequence types common to both Cambodia and Australia are due to homoplasy. Journal of Clinical Microbiology. 2015;53:323–326. doi: 10.1128/JCM.02574-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Godoy et al. (2003).Godoy D, Randle G, Simpson AJ, Aanensen DM, Pitt TL, Kinoshita R, Spratt BG. Multilocus sequence typing and evolutionary relationships among the causative agents of melioidosis and glanders, Burkholderia pseudomallei and Burkholderia mallei. Journal of Clinical Microbiology. 2003;41:2068–2079. doi: 10.1128/JCM.41.5.2068-2079.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayden et al. (2012).Hayden HS, Lim R, Brittnacher MJ, Sims EH, Ramage ER, Fong C, Wu Z, Crist E, Chang J, Zhou Y, Radey M, Rohmer L, Haugen E, Gillett W, Wuthiekanun V, Peacock SJ, Kaul R, Miller SI, Manoil C, Jacobs MA. Evolution of Burkholderia pseudomallei in recurrent melioidosis. PLoS ONE. 2012;7:e1802. doi: 10.1371/journal.pone.0036507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holden et al. (2004).Holden MT, Titball RW, Peacock SJ, Cerdeno-Tarraga AM, Atkins T, Crossman LC, Pitt T, Churcher C, Mungall K, Bentley SD, Sebaihia M, Thomson NR, Bason N, Beacham IR, Brooks K, Brown KA, Brown NF, Challis GL, Cherevach I, Chillingworth T, Cronin A, Crossett B, Davis P, DeShazer D, Feltwell T, Fraser A, Hance Z, Hauser H, Holroyd S, Jagels K, Keith KE, Maddison M, Moule S, Price C, Quail MA, Rabbinowitsch E, Rutherford K, Sanders M, Simmonds M, Songsivilai S, Stevens K, Tumapa S, Vesaratchavest M, Whitehead S, Yeats C, Barrell BG, Oyston PC, Parkhill J. Genomic plasticity of the causative agent of melioidosis, Burkholderia pseudomallei. Proceedings of the National Academy of Sciences of the United States of America. 2004;101:14240–14245. doi: 10.1073/pnas.0403302101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holmes, Urwin & Maiden (1999).Holmes EC, Urwin R, Maiden MC. The influence of recombination on the population structure and evolution of the human pathogen Neisseria meningitidis. Molecular Biology and Evolution. 1999;16:741–749. doi: 10.1093/oxfordjournals.molbev.a026159. [DOI] [PubMed] [Google Scholar]
- Hunter & Gaston (1988).Hunter PR, Gaston MA. Numerical index of the discriminatory ability of typing systems: an application of Simpson’s index of diversity. Journal of Clinical Microbiology. 1988;26:2465–2466. doi: 10.1128/jcm.26.11.2465-2466.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koonpaew et al. (2000).Koonpaew S, Ubol MN, Sirisinha S, White NJ, Chaiyaroj SC. Genome fingerprinting by pulsed-field gel electrophoresis of isolates of Burkholderia pseudomallei from patients with melioidosis in Thailand. ACTA Tropica. 2000;74:187–191. doi: 10.1016/S0001-706X(99)00069-8. [DOI] [PubMed] [Google Scholar]
- Limmathurotsakul & Peacock (2011).Limmathurotsakul D, Peacock SJ. Melioidosis: a clinical overview. British Medical Bulletin. 2011;99:125–139. doi: 10.1093/bmb/ldr007. [DOI] [PubMed] [Google Scholar]
- Lozupone, Hamady & Knight (2006).Lozupone C, Hamady M, Knight R. UniFrac–an online tool for comparing microbial community diversity in a phylogenetic context. BMC Bioinformatics. 2006;7:371. doi: 10.1186/1471-2105-7-371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maiden et al. (1998).Maiden MC, Bygraves JA, Feil E, Morelli G, Russell JE, Urwin R, Zhang Q, Zhou J, Zurth K, Caugant DA, Feavers IM, Achtman M, Spratt BG. Multilocus sequence typing: a portable approach to the identification of clones within populations of pathogenic microorganisms. Proceedings of the National Academy of Sciences of the United States of America. 1998;95:3140–3145. doi: 10.1073/pnas.95.6.3140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCombie, Finkelstein & Woods (2006).McCombie RL, Finkelstein RA, Woods DE. Multilocus sequence typing of historical Burkholderia pseudomallei isolates collected in Southeast Asia from 1964 to 1967 provides insight into the epidemiology of melioidosis. Journal of Clinical Microbiology. 2006;44:2951–2962. doi: 10.1128/JCM.00725-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Naharro, Rubio & Luengo (2011).Naharro G, Rubio P, Luengo JM. Aeromonas. In: Liu D, editor. Molecular detection of human bacterial pathogens. Boca Raton: Taylor & Francis/CRC Press; 2011. pp. 789–799. [Google Scholar]
- Pearson et al. (2009).Pearson T, Giffard P, Beckstrom-Sternberg S, Auerbach R, Hornstra H, Tuanyok A, Price EP, Glass MB, Leadem B, Beckstrom-Sternberg JS, Allan GJ, Foster JT, Wagner DM, Okinaka RT, Sim SH, Pearson O, Wu Z, Chang J, Kaul R, Hoffmaster AR, Brettin TS, Robison RA, Mayo M, Gee JE, Tan P, Currie BJ, Keim P. Phylogeographic reconstruction of a bacterial species with high levels of lateral gene transfer. BMC Biology. 2009;7:78. doi: 10.1186/1741-7007-7-78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Puthucheary (2009).Puthucheary SD. Melioidosis in Malaysia. Medical Journal of Malaysia. 2009;64(4):266–274. [PubMed] [Google Scholar]
- Ribot et al. (2006).Ribot EM, Fair MA, Gautom R, Cameron DN, Hunter SB, Swaminathan B, Barrett TJ. Standardization of pulsed-field gel electrophoresis protocols for the subtyping of Escherichia coli O157:H7, Salmonella, and Shigella for PulseNet. Foodborne Pathogens and Disease. 2006;3:59–67. doi: 10.1089/fpd.2006.3.59. [DOI] [PubMed] [Google Scholar]
- Richter et al. (2015).Richter M, Rossello-Mora R, Oliver Glockner F, Peplies J. JSpeciesWS: a web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics. 2015 doi: 10.1093/bioinformatics/btv681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sabat et al. (2013).Sabat AJ, Budimir A, Nashev D, Sa-Leao R, Van Dijl J, Laurent F, Grundmann H, Friedrich AW, Markers ESGoE. Overview of molecular typing methods for outbreak detection and epidemiological surveillance. Euro Surveillance. 2013;18:20380–20394. doi: 10.2807/ese.18.04.20380-en. [DOI] [PubMed] [Google Scholar]
- Schloss et al. (2009).Schloss PD, Westcott SL, Ryabin T, Hall JR, Hartmann M, Hollister EB, Lesniewski RA, Oakley BB, Parks DH, Robinson CJ, Sahl JW, Stres B, Thallinger GG, Van Horn DJ, Weber CF. Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Applied and Environmental Microbiology. 2009;75:7537–7541. doi: 10.1128/AEM.01541-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shimizu (2013).Shimizu K. Metabolic regulation of a bacterial cell system with emphasis on Escherichia coli metabolism. ISRN Biochemistry. 2013;2013:645983. doi: 10.1155/2013/645983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spector (2009).Spector MP. Physiology. In: Schaechter M, editor. Encyclopaedia of microbiology. Philadelphia: Elsevier; 2009. pp. 242–264. [Google Scholar]
- Tamura & Nei (1993).Tamura K, Nei M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Molecular Biology and Evolution. 1993;10:512–526. doi: 10.1093/oxfordjournals.molbev.a040023. [DOI] [PubMed] [Google Scholar]
- Tamura et al. (2013).Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: molecular evolutionary genetics analysis version 6.0. Molecular Biology and Evolution. 2013;30:2725–2729. doi: 10.1093/molbev/mst197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tankouo-Sandjong et al. (2007).Tankouo-Sandjong B, Sessitsch A, Liebana E, Kornschober C, Allerberger F, Hachler H, Bodrossy L. MLST-v, multilocus sequence typing based on virulence genes, for molecular typing of Salmonella enterica subsp. enterica serovars. Journal of Microbiological Methods. 2007;69:23–36. doi: 10.1016/j.mimet.2006.11.013. [DOI] [PubMed] [Google Scholar]
- Torpdahl et al. (2005).Torpdahl M, Skov MN, Sandvang D, Baggesen DL. Genotypic characterization of Salmonella by multilocus sequence typing, pulsed-field gel electrophoresis and amplified fragment length polymorphism. Journal of Microbiological Methods. 2005;63:173–184. doi: 10.1016/j.mimet.2005.03.006. [DOI] [PubMed] [Google Scholar]
- Tuanyok et al. (2007).Tuanyok A, Auerbach RK, Brettin TS, Bruce DC, Munk AC, Detter JC, Pearson T, Hornstra H, Sermswan RW, Wuthiekanun V, Peacock SJ, Currie BJ, Keim P, Wagner DM. A horizontal gene transfer event defines two distinct groups within Burkholderia pseudomallei that have dissimilar geographic distributions. Journal of Bacteriology. 2007;189:9044–9049. doi: 10.1128/JB.01264-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Urwin & Maiden (2003).Urwin R, Maiden MC. Multi-locus sequence typing: a tool for global epidemiology. Trends in Microbiology. 2003;11:479–487. doi: 10.1016/j.tim.2003.08.006. 8. [DOI] [PubMed] [Google Scholar]
- White (2003).White NJ. Melioidosis. Lancet. 2003;361:1715–1722. doi: 10.1016/S0140-6736(03)13374-0. [DOI] [PubMed] [Google Scholar]
- Wiersinga, Currie & Peacock (2012).Wiersinga WJ, Currie BJ, Peacock SJ. Melioidosis. New England Journal of Medicine. 2012;367:1035–1044. doi: 10.1056/NEJMra1204699. [DOI] [PubMed] [Google Scholar]
- Winsor et al. (2008).Winsor GL, Khaira B, Van Rossum T, Lo R, Whiteside MD, Brinkman FS. The Burkholderia Genome Database: facilitating flexible queries and comparative analyses. Bioinformatics. 2008;24:2803–2804. doi: 10.1093/bioinformatics/btn524. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The following information was supplied regarding data availability:
The research in this article did not generate any raw data.