

ABSTRACT
The molecular regulation of tumor induced T-cell exhaustion remains poorly characterized. Recently, we compared the transcriptome of “exhausted” CD8+ T cells infiltrating melanomas to those of naive and acutely stimulated CD8+ T cells. We demonstrated that MAF is over-expressed and plays a key role in driving the transcriptional program of T-cell exhaustion.
KEYWORDS: CD8+ T cell, exhaustion, maf, melanoma
Numerous studies in humans and in mice clearly established that T cells can recognize tumor cells and control tumor progression. However, despite evidence for spontaneous immune activation in a limited number of cancer patients, the immune system most frequently fails to protect patients with metastatic cancers as tumors develop strategies to escape T-cell-mediated immunity. We know that multiple mechanisms diminish antitumor T-cell responses. These include, for instance, the concomitant accumulation of high numbers of regulatory T cells and myeloid-derived suppressor cells. Further cells and factors contribute to the immunosuppressive tumor micro-environment which inhibits anticancer effector T cells.
A major limitation is the T-cell intrinsic attenuation of tumor-antigen-specific T cells to exert effector functions against tumors. So far, the prevailing explanation for this failure was that tumor-targeting T cells become tolerized or anergized over time. More recently, another T-cell differentiation pathway has been suggested, and different lines argue in favor of the fact that tumor-reactive T cells undergo similar differentiation as antigen-specific T cells in chronic infection, i.e. they undergo “exhaustion” and become functionally attenuated. This state has been thoroughly studied during chronic viral infection, both in mouse and human. It is characterized by the expression of multiple inhibitory receptors (PD-1, CTLA-4, TIM-3…) found to be also expressed on functionally impaired tumor-infiltrated lymphocytes (TILs) (reviewed in1,2). These phenotypical and functional similarities led to the extension of the term exhaustion to TILs. However, scarce details are available on the transcriptional program of TILs and on the transcription factors regulating this state.
In our recent study,3 we took advantage of a mouse model of induced melanoma based on conditional deletion of tumor suppressor genes with concomitant expression of a natural mouse tumor antigen (TiRP mice). In this model, tumor-intrinsic factors control the development of aggressive tumors and their expression of an inflammatory/immunosuppressive program.4 Intra-tumor T cells expressed high levels of inhibitory receptors such as PD-1 and had poor capacity to produce IFNγ upon restimulation, suggesting that they are exhausted. Using this model, we have established the gene expression signature associated with CD8+ T-cell exhaustion during melanoma development. We showed that tumor- and virus-induced exhaustion share many features, with expression of genes encoding molecules such as inhibitory receptors or particular transcription factors. Among the latter, we focused our study on the two transcription factors with the highest fold increase in TILs compared to effector T cells. Nr4a2, encoding an orphan nuclear receptor, was highly expressed in both virus- and tumor-induced exhaustion, whereas maf was highly overexpressed in tumor-exhausted CD8+ T cells and only very weakly during chronic viral infection. We confirmed the overexpression for both genes in Melan-A/MART-1 specific CD8+ T cells isolated from tumor-infiltrated lymph nodes (TILN) from melanoma patients.
Importantly, overexpression of maf by retroviral transduction of tumor-specific CD8+ T cells dampened their intra-tumor accumulation and antitumor activity, while overexpression of nr4a2 did not affect CD8+ T-cell properties. We went on to show that maf expression in antitumor CD8+ T cells contributes to their polarization toward an exhausted phenotype, with high expression of exhaustion associated genes such as il10, bcl6, pdcd1 and others (Fig. 1). We demonstrated in vitro that maf expression could be regulated by IL-6 and/or TGFβ when CD8+ T cells are stimulated. These cytokines are produced at high levels in our TiRP melanoma and appear to be important regulators of exhaustion in this model. In vitro, these cytokines could partially mimic the transcriptional program associated with exhaustion, inducing higher levels of bcl6 or il10 transcripts and dampening the expression of transcripts encoding for ifng or gzmb. However, the expression of some other genes were not affected (nr4a2, nr4a3, rgs1or rgs16) showing that the action of the microenvironment on T cells is much more complex than our in vitro system. Using maf knockout mice in similar experiments, we established that maf−/− CD8+ T cells were much more resistant to the presence of these inhibitory cytokines in terms of proliferation, survival and production of cytokines. In vivo, we also demonstrated that maf−/− tumor specific CD8+ T cells had heightened capacity to restrain tumor growth correlated with a better accumulation inside the tumor, a higher capacity to produce IFNγ upon restimulation and lower surface expression of PD-1 and LAG-3.
Figure 1.
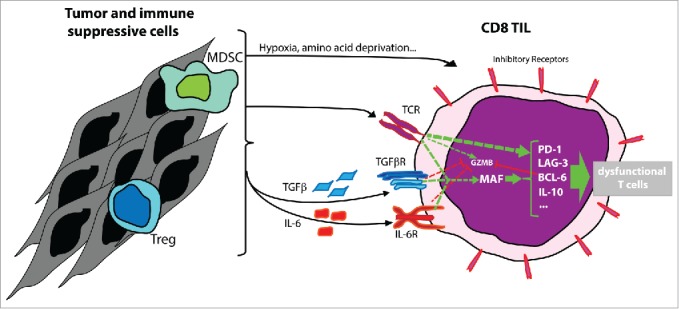
In vivo, tumor infiltrating lymphocytes (TIL) found inside melanoma express high level of MAF. In vitro, overexpression of MAF can be induced by TCR triggering together with IL-6, TGFβ or both. Inside the tumor microenvironment, maf expression could also be promoted by the production of other factors such as hypoxia, amino acid deprivation, other cytokines produced by tumor cells or immunosuppressive cells such as myeloid-derived suppressor cells (MDSCs) or regulatory T cells (Treg). In CD8+ TILs, MAF enhances the expression of pdcd1, lag3, bcl6, il10 and further genes associated with exhaustion, leading to the impairment of T-cell function.
Maf plays central role in several subtypes of CD4+ T cells such as Th2 cells (IL-4 production5), Th17 cells (IL-10 and IL-22 production6,7) or Tfh.8 To our knowledge, our study was the first to show high level of maf in CD8+ T cells in vivo. How general this high expression is in other tumor types still needs to be determined. Nevertheless, it is clear that high level of maf leads to a biased polarization toward an exhausted phenotype, is deleterious to cytotoxic functions of CD8+ T cells, and prevents Th1 differentiation in CD4+ T cells. Therefore, maf represents an interesting target to improve immunotherapy of cancer.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
References
- 1.Wherry EJ. T cell exhaustion. Nat Immunol 2011; 12:492–9; PMID:21739672; http://dx.doi.org/ 10.1038/ni.2035 [DOI] [PubMed] [Google Scholar]
- 2.Verdeil G, Fuertes Marraco SA, Murray T, Speiser DE. From T cell “exhaustion” to anti-cancer immunity. Biochim Biophys Acta 2015; PMID:26123831; http://dx.doi.org/ 10.1016/j.bbcan.2015.06.007 [DOI] [PubMed] [Google Scholar]
- 3.Giordano M, Henin C, Maurizio J, Imbratta C, Bourdely P, Buferne M, Baitsch L, Vanhille L, Sieweke MH, Speiser DE et al.. Molecular profiling of CD8 T cells in autochthonous melanoma identifies maf as driver of exhaustion. EMBO J 2015; 34(15):2042–58; PMID:26139534; http://dx.doi.org/ 10.15252/embj.201490786 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Soudja SM, Wehbe M, Mas A, Chasson L, de Tenbossche CP, Huijbers I, Van den Eynde B, Schmitt-Verhulst AM. Tumor-initiated inflammation overrides protective adaptive immunity in an induced melanoma model in mice. Cancer Res 2010; 70:3515–25; PMID:20406967; http://dx.doi.org/ 10.1158/0008-5472.CAN-09-4354 [DOI] [PubMed] [Google Scholar]
- 5.Ho IC, Lo D, Glimcher LH. c-maf promotes T helper cell type 2 (Th2) and attenuates Th1 differentiation by both interleukin 4-dependent and -independent mechanisms. J Exp Med 1998; 188:1859–66; PMID:9815263; http://dx.doi.org/ 10.1084/jem.188.10.1859 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Rutz S, Noubade R, Eidenschenk C, Ota N, Zeng W, Zheng Y, Hackney J, Ding J, Singh H, Ouyang W. Transcription factor c-Maf mediates the TGF-beta-dependent suppression of IL-22 production in T(H)17 cells. Nat Immunol 2011; 12:1238–45; PMID:22001828; http://dx.doi.org/ 10.1038/ni.2134 [DOI] [PubMed] [Google Scholar]
- 7.Xu J. Yang Y, Qiu G, Lal G, Wu Z, Levy DE, Ochando JC, Bromberg JS, Ding Y. c-Maf regulates IL-10 expression during Th17 polarization. J Immunol 2009; 182:6226–36; PMID:19414776; http://dx.doi.org/ 10.4049/jimmunol.0900123 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Kroenke MA, Eto D, Locci M, Cho M, Davidson T, Haddad EK, Crotty S. Bcl6 and maf cooperate to instruct human follicular helper CD4 T cell differentiation. J Immunol 2012; 188:3734–44; PMID:22427637; http://dx.doi.org/ 10.4049/jimmunol.1103246 [DOI] [PMC free article] [PubMed] [Google Scholar]