

Abstract
Background
To explore whether plasma fatty acids and SNPs in the fatty acid desaturase (FADS) gene associated with type 2 diabetes (T2D) and coronary artery disease (CAD).
Methods
In this cross-sectional study, we utilized gas chromatography–mass spectrometric analysis and the high-resolution melting method to detect plasma fatty acids and SNPs respectively (rs174537G>T, rs174616C>T, rs174460T>C, and rs174450A>C) in 234 T2D, 200 CAD, 185 T2D&CAD patients, and 253 healthy controls.
Results
We found that T2D&CAD patients had the highest plasma arachidonic acid, dihomo-gamma-linolenic acid and delta-6 desaturase, and the lowest stearic acid, linolenic acid, and saturated fatty acids; plasma eicosapentaenoic acid and docosahexaenoic acid elevated in T2D patients, but significantly reduced in CAD patients. Moreover, T2D patients with rs174537 GG genotype were at risk of developing T2D&CAD (odds ratio (OR) 1.763; 95 % CI 1.143–2.718; p = 0.010), with elevated plasma LDL-cholesterol, arachidonic acid, and delta-6 desaturase.
Conclusions
Our results show that SNPs in FADS gene (particularly rs174537) associate with plasma fatty acids and desaturase levels in patients with both T2D and CAD, which maybe increases the risk of CAD in diabetic patients.
Electronic supplementary material
The online version of this article (doi:10.1186/s12967-016-0834-8) contains supplementary material, which is available to authorized users.
Keywords: Coronary artery disease (CAD), Type 2 diabetes (T2D), Fatty acids, Desaturase, Polymorphism(s)
Background
Type 2 diabetes (T2D) and coronary artery disease (CAD) are global public health concerns [1]. People with diabetes also have a high incidence of CAD [2]; T2D patients with CAD (T2D&CAD) have mortality rates that are about 2 to 4 times higher than those of T2D patients without CAD [3]. Recently, An et al. [4] reported that in Chinese adults, diabetes is associated with a substantially increased risk of cardiovascular-cause mortality. Thus, it is important to explore risk factors of T2D, CAD and T2D&CAD: these factors include obesity, metabolic syndrome, family history of T2D or CAD, impaired glucose tolerance, low physical activity, increased plasma triglycerides (TG), and decreased HDL-cholesterol [5–7]. Of these risk factors, plasma fatty acid composition is of particular interest because of the role of plasma fatty acids in normal and pathophysiologic responses [8].
The desaturase enzymes are of great importance in the chemical structure and functions of fatty acids. The stearoyl coenzyme is definitely required for the conversion of saturated fatty acids into monounsaturated fatty acids, and delta-5 desaturase (D5D) and delta-6 desaturase (D6D) catalyze the rate-limiting steps in the conversion of linoleic acid (LA, C18:2n-6) and gamma-linolenic acid (GLA, C18:3n-6) into long-chain n-6 and n-3 polyunsaturated fatty acids. In fact, the influence of desaturase enzymes on fatty acids has been identified as an essential factor of T2D and CAD [9]. The fatty acid desaturase genes FADS1 and FADS2 code, respectively, for the desaturase enzymes D5D and D6D, which cluster in a head-on-head direction on chromosome 3. The role of FADS3 in fatty acid metabolism is still unclear, although FADS3 is also clustered at the same location as FADS1 and FADS2 [10].
Previous studies have suggested that plasma and tissue concentrations of n-3 and n-6 fatty acids are associated with several single nucleotide polymorphisms (SNPs) in the FADS1 and FADS2 genes [11–13]. Our previous research demonstrated that participants with the rs174460 C allele had a higher risk of CAD than those who had the corresponding T allele, and that the rs174537 T allele is associated with a lower risk of CAD when compare with the carriers of rs174537 G allele [14]. Genome-wide association studies in humans have also highlighted the influence of variations in the FADS1 and FADS2 gene cluster on glucose and lipid metabolisms, such as total cholesterol [15] and LDL-cholesterol, and disease conditions such as metabolic syndrome, myocardial infarction, and dyslipidemia [16, 17]. However, few studies have specifically focused on the association between genetic polymorphisms in the FADS gene cluster and the risk of T2D&CAD. Due to the functions of FADS gene in fatty acids metabolism and homeostasis, we hypothesize that SNPs in FADS gene will influent the desaturase activity, which therefore alters the characteristics of plasma fatty acids and the risk of T2D&CAD. To test our hypothesis, in this study, we explored genetic polymorphisms in the FADS gene cluster and plasma fatty acids in patients with T2D, CAD or T2D&CAD, and healthy controls.
Methods
Patients
872 unrelated individuals were recruited at Zhongnan Hospital of Wuhan University, including: 234 patients with T2D, 200 patients with CAD, 185 patients with T2D&CAD, and 253 healthy controls. T2D was diagnosed according to 2012 ADA diabetes treatment guidelines [18]. CAD patients were with either coronary angiography or myocardial infarction, which was defined as a ≥50 % stenosis in any major coronary artery [19]. Healthy controls were randomly selected from physical examination population who had normal liver function and kidney function, and fasting blood glucose and lipids were within reference ranges; and exclusion criteria were diabetes, cardiovascular disease or other serious disease or use of any medicine or fish oil supplement. Healthy controls should maintain their usual diet, T2D patients should maintain low-fat and -carbohydrate diet, CAD patients should maintain low-fat and -sodium diet, T2D&CAD patients should maintain low-fat, -carbohydrate and -sodium diet. All patients underwent statins for cholesterol-lowering. Written informed consent was obtained from all participants, and the study protocol was approved by the ethics committee of Zhongnan Hospital of Wuhan University.
Fatty acids analysis
To detect fatty acid methyl esters, we used a previously described method [20] with minor modifications. After filtration, 1μL of sample was injected into the Agilent 7890/5975 gas chromatography–mass spectrometry system (Agilent Technologies, Santa Clara, CA, USA). The HP-INNOWax column (30 m × 0.25 mm × 0.25 μm) (Agilent Technologies) was used for gas chromatography–mass spectrometry analysis. The gas chromatography oven temperature was programmed to increase from 60 °C (2 min) to 160 °C at 20 °C/min and then to 240 °C (6 min) at 10 °C/min, and the flow rate of carrier gas was at 1 mL/min. The interface temperature was 250 °C, the ion-source temperature was 230 °C, and the electron-impact ionization was 70 eV, with a full scan ranging from 20 to 550 m/z and a solvent delay of 3 min. Peak identification of target compounds was based on the retention times and full scan spectra of the standards. The levels of individual fatty acids are expressed as a percentage of total fatty acid methyl esters. D5D activity was estimated as the ratio of AA to DGLA, and D6D activity was estimated as the ratio of GLA to LA. Palmitoleic acid (C16:1) to palmitic acid (C16:0) and OA to SA ratios were used as a surrogate estimation of the activity of D9D-16 and D9D-18, respectively.
SNP selection and genotyping
We used a commercially available DNA isolation kit (TIANamp, Beijing, China) to extract genomic DNA from whole blood (200 μL), according to the protocol. SNPs in FADS gene were identified using the International HapMap Project SNP database and Tag SNPs (tSNPs) were selected with their features in Haploview V4.1 by a minor allele frequency (MAF >5 %) and pairwise tagging (r2 ≥ 0.8), and reference population was CHB. Moreover, we examined linkage disequilibrium (LD) between any two of four selected tSNPs using the SNP Annotation and Proxy Search (SNAP) database. Four SNPs (Flap structure-specific endonuclease-10154 rs174537G>T, FADS2 rs174616C>T, rs174460T>C, and rs174450A>C) were genotyped with the high-resolution melting of a small amplicon, as described previously [14].
Statistical analysis
All statistical analyses were performed with SPSS 19.0 for Windows (IBM, Armonk, NY, USA). Continuous variables are expressed as means ± SDs. Skewed variables are described by the median and interquartile range. K-independent nonparametric analysis was used to compare the fatty acid levels among the T2D, CAD, T2D&CAD, and healthy groups. Ordinal logistic regression analysis (ordinal values were 1 for healthy controls, 2 for patients with T2D or CAD, and 3 for patients with T2D&CAD) were used to evaluate the associations of SNPs with diseases. Hardy–Weinberg equilibrium, genotype and allele frequency distributions were performed using SNPStats (Barcelona, Spain) [21] after adjusting for age and sex. All statistical tests were two-sided, and p values of less than 0.05 or Bonferroni correction-adjusted p values of less than 0.0125 were considered statistically significant.
Results
Clinical characteristics
The demographic and clinical characteristics of T2D, CAD, and T2D&CAD patients and healthy controls are listed in Table 1. We found significant differences in plasma TG, TC, HDL-cholesterol, LDL-cholesterol, and fasting plasma glucose (FPG) levels among four groups. Besides, T2D patients had the highest FPG and HbA1c levels, yet CAD patients and T2D&CAD patients had lower levels of TC and LDL-cholesterol than that in T2D patients.
Table 1.
Clinical characteristics and Differential fatty acid levels and desaturase activities in healthy controls and patients
| Characteristics | Healthy controls (n = 253) | T2D patients (n = 234) | CAD patients (n = 200) | T2D&CAD patients (n = 185) | p a |
|---|---|---|---|---|---|
| Male/Female (%) | 60.5/39.5 | 62/38 | 55.5/44.5 | 63.2/36.8 | 0.412 |
| Age (year) | 59.73 ± 10.06 | 57.74 ± 12.76 | 59.47 ± 10.53 | 60.30 ± 9.73 | 0.282 |
| Systolic blood pressure (mmHg) | 128.19 ± 20.59 | 130.90 ± 17.73 | 129.16 ± 18.94 | 131.87 ± 19.54 | 0.247 |
| Diastolic blood pressure (mmHg) | 78.42 ± 13.75 | 76.64 ± 10.71 | 78.51 ± 12.20 | 78.61 ± 11.78 | 0.513 |
| Total cholesterol (mmol/l) | 4.43 (3.97, 4.91) | 4.68 (3.99, 5.31) | 4.05 (3.33, 4.64) | 4.07 (3.31, 4.99) | <0.0001 |
| Triglyceride (mmol/l) | 1.01 (0.78, 1.32) | 1.82 (1.21, 2.74) | 1.26 (0.93, 1.60) | 1.37 (1.01, 1.98) | <0.0001 |
| HDL-cholesterol (mmol/l) | 1.29 (1.12, 1.49) | 1.05 (0.90, 1.20) | 1.17 (0.99, 1.35) | 1.03 (0.87, 1.29) | <0.0001 |
| LDL-cholesterol (mmol/l) | 2.70 (2.33, 3.05) | 2.70 (2.10, 3.21) | 2.42 (1.72, 2.89) | 2.42 (1.84, 3,12) | <0.0001 |
| Fasting plasma glucose (mmol/l) | 4.92 (4.59, 5.33) | 8.50 (6.45, 11.73) | 5.68 (5.11, 6.29) | 6.93 (6.03, 8.16) | <0.0001 |
| HbA1c (%) | _ | 7.70 (6.50, 9.48) | _ | 6.70 (6.10, 7.30) | <0.0001 |
| HbA1c (mmol/mol) | _ | 60.66 (47.54, 80.11) | _ | 49.73 (43.17, 56.28) | <0.0001 |
| Fatty acids (%) | |||||
| Palmitic acid, C16:0 | 22.41 (21.40, 23.63) | 23.61 (21.78, 25.34) | 23.17 (21.38, 24.64) | 23.40 (21.76, 24.77) | <0.0001 |
| Stearic acid, C18:0 | 9.33 (8.52, 9.98) | 9.14 (7.63, 10.78) | 9.16 (8.37, 9.89) | 8.96 (8.14, 9.85) | 0.048 |
| Total monounsaturated fatty acid | 16.05 (14.14, 18.12) | 19.27 (16.93, 21.71) | 17.40 (15.47, 19.60) | 18.47 (15.99, 20.65) | <0.0001 |
| Palmitoleic acid, C16:1 | 0.70 (0.52, 0.95) | 0.88 (0.42, 1.50) | 0.95 (0.66, 1.23) | 0.90 (0.59, 1.55) | <0.0001 |
| Oleic acid, C18:1n-9 | 14.85 (13.10, 16.74) | 18.29 (16.06, 20.52) | 15.90 (14.29, 17.83) | 17.23 (14.75, 19.65) | <0.0001 |
| Total polyunsaturated n-3 fatty acid | 3.65 (2.93, 4.31) | 7.07 (4.42, 10.05) | 3.40 (2.78, 4.00) | 4.22 (3.20,7.25) | <0.0001 |
| α-linolenic acid, C18:3n-3 | 0.55 (0.35, 0.76) | 0.58 (0.13, 1.08) | 0.55 (0.34, 0.78) | 0.65 (0.34, 0.92) | 0.195 |
| Eicosapentaenoic acid, C20:5n-3 | 0.21 (0.00, 0.46) | 0.87 (0.28, 1.75) | 0.17 (0.00, 0.40) | 0.18 (0.00, 1.17) | <0.0001 |
| Docosahexaenoic acid, C22:6n-3 | 2.72 (2.17, 3.38) | 4.78 (1.95, 7.40) | 2.52 (2.00, 3.12) | 3.22 (2.49, 5.01) | <0.0001 |
| Total polyunsaturated n-6 fatty acid | 46.32 (43.57, 49.04) | 45.66 (42.34, 49.19) | 44.37 (41.37, 47.46) | 45.08 (41.69, 48.27) | <0.0001 |
| Linoleic acid, C18:2n-6 | 35.96 (32.94, 39.49) | 34.63 (30.50, 40.53) | 33.11 (29.66, 36.64) | 32.61 (29.36, 36.47) | <0.0001 |
| γ-linolenic acid, C18:3n-6 | 0.22 (0.05, 0.42) | 0.13 (0.00, 0.37) | 0.32 (0.10, 0.57) | 0.17 (0.00, 0.49) | <0.0001 |
| Dihomo-γ-linolenic acid, C20:3n-6 | 1.35 (1.07, 1.63) | 1.37 (0.16, 2.13) | 1.54 (1.18, 2.03) | 1.63 (1.03, 2.40) | <0.0001 |
| Arachidonic acid, C20:4n-6 | 7.98 (6.68, 9.43) | 7.59 (3.97, 10.82) | 7.92 (6.28, 9.70) | 9.18 (6.67, 11.27) | <0.0001 |
| Desaturase activity | |||||
| C20:4n-6/C20:3n-6 (D5D) | 6.22 (4.61, 7.67) | 5.11 (3.71, 8.26) | 5.23 (3.39, 7.60) | 5.07 (3.47, 7.75) | 0.051 |
| C20:4n-6/C18:2n-6 (D6D) | 0.22 (0.18, 0.27) | 0.22 (0.10, 0.35) | 0.24 (0.19, 0.31) | 0.28 (0.19, 0.36) | <0.0001 |
| C16:1/C16:0 (D9D-16) | 0.03 (0.02, 0.04) | 0.04 (0.02, 0.07) | 0.04 (0.03, 0.05) | 0.04 (0.03, 0.06) | 0.002 |
| C18:1n-9/C18:0(D9D-18) | 1.62 (1.39, 1.90) | 1.97 (1.58, 2.42) | 1.77 (1.49, 2.06) | 1.94 (1.67, 2.29) | <0.0001 |
| n-3/n-6 | 0.08 (0.06, 0.10) | 0.16 (0.09, 0.23) | 0.08 (0.06, 0.09) | 0.10 (0.07, 0.17) | <0.0001 |
a p values derived from K-independent non-parametric analysis
Plasma fatty acids and desaturase activities
As shown in Fig. 1, a total of 34 standard fatty acid methyl esters were distinctly separated on HP-INNOWax column within 20 min. Plasma fatty acids and desaturase activities, except for Alpha-linolenic acid (ALA, 18:3n-3) and D5D,differed among the four groups according to the results of K-independent nonparametric analysis (Table 1). Of the four groups, T2D&CAD patients had the highest levels of arachidonic acid (AA, C20:4n-6), dihomo-gamma-linolenic acid (DGLA, C20:3n-6) and D6D; and the lowest levels of stearic acid (SA, C18:0), LA and saturated fatty acids. Of the four groups, T2D patients had the highest levels of palmitic acid (C16:0), oleic acid (OA, C18:1n-9), EPA, DHA, monounsaturated fatty acids, n-3 fatty acids, D9D-16 (C16:1/C16:0), D9D-18 (C18:1n9/C18:0) and n-3/n-6; and the lowest levels of GLA, AA and D6D (AA/LA). Of the four groups, CAD patients had the highest levels of palmitoleic acid (C16:1) and GLA and the lowest levels of EPA, DHA, and n-3 fatty acids.
Fig. 1.
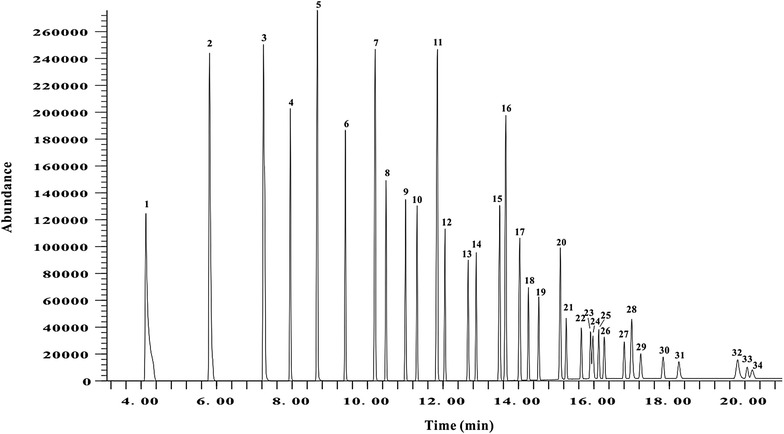
Total-ion chromatogram of standard fatty acid methyl esters separated on HP-INNOWax column. Peaks: 1, C6:0, caproic acid; 2, C8:0, caprylic acid; 3, C10:0, capric acid; 4, C11:0, undecanoic acid; 5, C12:0, lauric acid; 6, C13:0, tridecanoic acid; 7, C14:0, myristic acid; 8, C14:1, myristoleic acid; 9, C15:0, pentadecanoic acid; 10, C15:1, cis-10-pentadecenoic acid; 11, C16:0, palmitic acid; 12, C16:1, palmitoleic acid; 13, C17:0 heptadecanoic acid; 14, C17:1, cis-10-heptadecenoic acid; 15, C18:0, stearic acid; 16, C18:1n-9, oleic acid; 17, C18:2n-6, linoleic acid; 18, C18:3n-6, gamma-linolenic acid; 19, C18:3n-3, alpha-linolenic acid; 20, C20:0, arachidic acid; 21, C20:1, cis-11-eicosenoic acid; 22, C20:2, cis-11,14-eicosadienoic acid; 23, C20:3n-6, dihomo-gamma-linolenic acid; 24, C21:0, heneicosanoic acid; 25, C20:4n-6, arachidonic acid; 26, C20:3n3, cis-11,14,17-eicosatrienoic acid; 27, C20:5n-3, eicosapentaenoic acid; 28, C22:0, behenic acid; 29, C22:1n-9, erucic acid; 30, C22:2, cis-13,16-docosadienoic acid; 31, C23:0, tricosanoic acid; 32, C24:0, lignoceric acid; 33, C22:6n-3, docosahexaenoic acid; 34, C24:1, nervonic acid
Association of SNPs with T2D&CAD risk
Genotype distributions of the four SNPs were in Hardy–Weinberg equilibrium in healthy controls (Table 2). Among the four SNPs, only the genotype distributions of rs174537G>T differed in both additive (p = 0.032) and dominant models (p = 0.027) among all study participants. Ordinal logistic regression analysis was performed in healthy controls, T2D patients and T2D&CAD patients to assess whether rs174537 associated with the risk of T2D&CAD, the results revealed that T2D patients with rs174537 GG genotype were at higher risk of developing T2D&CAD (odds ratio (OR) 1.763; 95 % CI 1.143–2.718; p = 0.010) (Table 3). Besides, similar risk also existed in CAD patients with rs174537 GG genotype (OR, 2.050; 95 % CI 1.292–3.258; p = 0.002) (Additional file1: Table S1).
Table 2.
Distributions of genotype and allele frequency in healthy controls and patients
| SNP | Genotype | Healthy controls (n = 253) |
T2D patients (n = 234) |
CAD patients (n = 200) |
T2D &CAD patients (n = 185) | HWEa | p valueb | Additive model, p valuec | Dominant model, p valuec |
|---|---|---|---|---|---|---|---|---|---|
| rs174537G>T | GG | 65 | 88 | 58 | 63 | 0.56 | 0.009 | 0.032 | 0.027 |
| GT | 131 | 106 | 90 | 91 | |||||
| TT | 57 | 40 | 52 | 31 | |||||
| rs174616C>T | CC | 113 | 104 | 88 | 99 | 0.59 | 0.383 | 0.445 | 0.180 |
| TC | 115 | 109 | 90 | 68 | |||||
| TT | 25 | 21 | 22 | 18 | |||||
| rs174460C>T | TT | 151 | 129 | 106 | 112 | 0.56 | 0.185 | 0.136 | 0.344 |
| TC | 91 | 81 | 78 | 56 | |||||
| CC | 11 | 24 | 16 | 17 | |||||
| rs174450A>C | AA | 101 | 86 | 82 | 80 | 0.46 | 0.550 | 0.743 | 0.589 |
| AC | 122 | 113 | 96 | 80 | |||||
| CC | 30 | 35 | 22 | 25 |
aHardy–Weinberg Equilibrium (HWE) was calculated in healthy controls
b p values derived from the Chi square test of allele frequency
c p values derived from the Chi square test of genotype distribution
Table 3.
Risk estimation of SNPs by ordinal logistic regression analysis in healthy controls, T2D patients, T2D&CAD patientsa
| SNP | Group | Estimate | Wald value | p valueb | OR (95 % CI) |
|---|---|---|---|---|---|
| rs174537G>T | GG | 0.567 | 6.576 | 0.010 | 1.763 (1.143–2.718) |
| GT | 0.299 | 2.145 | 0.143 | 1.349 (0.904–2.014) | |
| rs174616C>T | CC | 0.256 | 0.932 | 0.334 | 1.292 (0.768–2.175) |
| CT | −0.143 | 0.285 | 0.593 | 0.867 (0.513–1.464) | |
| rs174450A>C | AA | 0.054 | 0.052 | 0.820 | 1.055 (0.663–1.680) |
| AC | 0.006 | 0.001 | 0.978 | 1.006 (0.637–1.590) | |
| rs174460C>T | TT | 0.336 | 1.388 | 0.239 | 1.399 (0.800–2.447) |
| TC | −0.162 | 1.006 | 0.316 | 0.850 (0.619–1.168) |
aOrdinal values were 1 for healthy controls, 2 for patients with T2D, and 3 for patients with T2D&CAD
b p values derived from ordinal logistic regression after adjustment for sex, age, TC, TG, HDL-cholesterol, LDL-cholesterol, and FPG
Clinical characteristics, plasma fatty acids and desaturase activities in rs174537 GG genotype participants
Compared with T2D&CAD patients with rs174537 GG genotype, healthy controls with GG genotype had lower levels of TC, TG, LDL-cholesterol, FPG, C16:0, PA, OA, ALA, DHA, DGLA, AA, n-3 fatty acids, n-6 fatty acids, monounsaturated fatty acid, D6D, D9D-18, and n-3/n-6 and had higher levels of HDL-cholesterol, SA and LA. CAD patients with GG genotype had higher levels of HDL-cholesterol and lower levels of FPG, OA, DHA, n-3 fatty acids, D9D-18, and n-3/n-6 than did T2D&CAD patients with GG genotype. T2D patients with GG genotype had higher levels of TC, TG, FPG, OA, EPA, LA, n-3 fatty acids, D5D and n-3/n-6 and had lower levels of LDL-cholesterol, and D6D than did T2D&CAD patients with GG genotype (Table 4). Moreover, we also compared healthy controls, T2D, CAD with T2D&CAD patients with rs174537 GT or TT genotype (Additional file 1: Table S2, Table S3).
Table 4.
Comparisons of clinical parameters, plasma fatty acids and desaturase activities grouping by rs174537 GG genotype
| Characteristics | Healthy controls GG (n = 65) |
T2D patients GG (n = 88) |
CAD patients GG (n = 58) |
T2D&CAD patients GG (n = 63) |
|---|---|---|---|---|
| Total cholesterol (mmol/l) | 4.50 ± 0.72* | 4.79 (4.00, 5.30)* | 4.05 (3.29, 4.60) | 4.52 (3.59, 5.50) |
| Triglyceride (mmol/l) | 0.97 (0.79, 1.22)* | 1.71 (1.21, 2.33)* | 1.31 (1.01, 1.60) | 1.36 (1.05, 1.94) |
| HDL-cholesterol (mmol/l) | 1.37 (1.10, 1.55)* | 1.07 (0.90, 1.21) | 1.10 (0.94, 1.37)* | 1.10 (0.93, 1.35) |
| LDL-cholesterol (mmol/l) | 2.74 (2.27, 3.29)* | 2.81(2.10, 3.17)* | 2.42 (1.83, 2.84) | 2.75 (2.03, 3.53) |
| Fasting plasma glucose (mmol/l) | 4.89 (4.58, 5.54)* | 8.51 (6.56, 12.24)* | 5.47 (5.01, 6.29)* | 7.00 (6.08, 8.24) |
| Total saturated fatty acid | 32.14 (31.21, 33.53) | 32.88 (31.24, 34.39) | 33.07 (31.21, 34.97) | 32.86 (29.45, 34.20) |
| Palmitic acid, C16:0 | 22.17 (21.44, 23.58)* | 23.62 (21.80, 25.37) | 22.46 (21.27, 24.41) | 23.96 (21.00, 25.09) |
| Stearic acid, C18:0 | 9.19 (8.51, 9.87)* | 9.02 (7.62, 10.55) | 9.24 (8.56, 10.19) | 8.45 (6.86, 9.75) |
| Total monounsaturated fatty acid | 16.13 (14.40, 18.27)* | 18.93 (16.80, 21.12) | 16.58 (15.04, 19.52) | 18.21 (15.41, 20.39) |
| Palmitoleic acid, C16:1 | 0.73 (0.57, 0.96)* | 0.65 (0.33, 1.30) | 0.88 (0.55, 1.26) | 1.00 (0.55, 1.67) |
| Oleic acid, C18:1n-9 | 15.09 (13.33, 16.89)* | 18.00 (16.16, 20.07)* | 15.38 (13.91, 17.65)* | 16.83 (13.76, 18.70) |
| Total polyunsaturated n-3 fatty acid | 3.92 (3.24, 4.56)* | 6.74 (1.99, 10.07)* | 3.40 (2.81, 4.09)* | 5.37 (3.45, 8.51) |
| Alpha-linolenic acid, C18:3n-3 | 0.66 (0.47, 0.86)* | 0.56 (0.12, 1.13) | 0.59 (0.36, 0.79) | 0.67 (0.33, 0.95) |
| Eicosapentaenoic acid, C20:5n-3 | 0.31 (0.10, 0.56) | 0.75 (0.21, 1.52)* | 0.28 (0.00, 0.47) | 0.27 (0.00, 1.80) |
| Docosahexaenoic acid, C22:6n-3 | 2.94 (2.33, 3.36)* | 4.32 (1.16, 6.91) | 2.50 (1.98, 3.12)* | 3.66 (2.72, 5.92) |
| Total polyunsaturated n-6 fatty acid | 45.70 (43.71, 48.00)* | 46.51 (43.40, 50.40) | 43.94 (41.11, 48.44) | 45.85 (41.87, 48.23) |
| Linoleic acid, C18:2n-6 | 36.21 (33.33, 39.64)* | 35.13 (30.12, 45.38)* | 32.59 (29.13, 35.74) | 32.52 (27.44, 36.37) |
| Gamma-linolenic acid, C18:3n-6 | 0.24 (0.09, 0.32) | 0.12 (0.01, 0.34) | 0.38 (0.11, 0.58) | 0.12 (0.00, 0.54) |
| Dihomo-gamma-linolenic acid, C20:3n-6 | 1.39 (1.11, 1.77)* | 1.29 (0.10, 2.12) | 1.67 (1.19, 2.13) | 1.64 (0.93, 2.58) |
| Arachidonic acid, C20:4n-6 | 7.66 (6.24, 9.19)* | 8.04 (3.97, 11.93) | 8.13 (6.33, 10.43) | 9.21 (6.50, 11.48) |
| Desaturase activity | ||||
| C20:4n-6/C20:3n-6 (D5D) | 5.54 (4.39, 7.30) | 5.92 (4.33, 8.41)* | 5.09 (3.47, 7.78) | 4.20 (2.66, 6.85) |
| C20:4n-6/C18:2n-6 (D6D) | 0.22 (0.16, 0.26)* | 0.23 (0.09, 0.39)* | 0.24 (0.19, 0.34) | 0.29 (0.19, 0.38) |
| C16:1/C16:0 (D9D-16) | 0.03 (0.02, 0.05) | 0.03 (0.01, 0.06) | 0.04 (0.03, 0.06) | 0.04 (0.03, 0.07) |
| C18:1n-9/C18:0(D9D-18)1 | 1.64 (1.39, 2.00)* | 1.86 (1.56, 2.43) | 1.73 (1.44, 2.06)* | 1.95 (1.69, 2.32) |
| n-3/n-6 | 0.08 (0.07, 0.10)* | 0.15 (0.04, 0.23)* | 0.08 (0.06, 0.09)* | 0.11 (0.07, 0.21) |
* p < 0.0125 derived from two-independent nonparametric analysis with Bonferroni correction (T2D&CAD patients vs healthy controls or T2D patients or CAD patients)
Discussion
In this study, we found that either plasma fatty acid levels or the estimated desaturase activities significantly differed in T2D, CAD, T2D&CAD patients and healthy controls; T2D patients with rs174537 GG genotype were at risk of developing T2D&CAD. Additionally, T2D&CAD patients with the rs174537 GG genotype had elevated plasma levels of TC, LDL-cholesterol, FPG, GLA, DGLA, and AA.
Interestingly, T2D patients had the highest EPA and DHA concentrations, whereas CAD patients had the lowest levels of plasma EPA and DHA. T2D&CAD patients showed increased D6D (AA/LA), D9D-18 (OA/SA), and D9D-16 (C16:1/C16:0) activity.
Our findings, to some extent, suggest that elevated levels of EPA and DHA might protect T2D patients from CAD. Several large-scale studies of fish oil supplements that contain high concentrations of EPA and DHA have also confirmed the beneficial effects of n-3 fatty acids on cardiovascular events [22, 23]. Other studies showed that fish oil supplements reduced the number of deaths and episodes of chronic cardiac failure [24] and patients treated with EPA had a 22 % lower risk of CAD than those not treated with EPA [2], which confirmed that high blood EPA levels,compare to DHA, can reduce cardiovascular events [25].
Our data further support these conclusions that EPA can prevent coronary events. Moreover, one study found that EPA suppresses TG synthesis in the liver and lowers serum TG and decreases atherogenic lipoproteins such as remnants and small, dense LDL-cholesterol and concluded that EPA was effective in reducing the incidence of CAD events for patients with dyslipidemia [26]. However, another study found that daily n-3 fatty acid supplements (containing EPA and DHA) did not reduce the rate of death from cardiovascular causes or other outcomes in patients at high risk for cardiovascular events [27]. We also found that CAD patients with or without T2D had lower levels of TC and LDL-cholesterol than did T2D patients. Clinical studies [28, 29] have shown that lipid-lowering therapy, such as statins, plays an extremely important role in preventing and treating CAD, and statins can competitively inhibit 3-hydroxy-3-methylglutaryl-coenzyme A reductase activity and can reduce hepatic cholesterol synthesis, circulating LDL-cholesterol levels, and other apolipoprotein B-containing lipoproteins [30, 31].
We found that T2D patients had the highest D9D-16 and D9D-18 levels, followed by T2D&CAD patients. Moreover, T2D&CAD patients had the lowest D5D activity and the highest D6D activity. Increased D9D activity is associated with insulin resistance, fatty liver disease, and metabolic syndrome, and D9D is considered to be a promising target for treating insulin resistance [32]. Kröger et al. [33] discovered that the estimated D5D activity was negatively correlated with diabetes risk, whereas the estimated D6D activity was strongly and positively correlated with diabetes risk. Several studies have suggested that participants with a high estimated D5D activity have a 50 % lower risk of diabetes compared with those with a low D5D activity; these findings were based on analysis of extreme quintiles [34, 35] or tertiles [36]. CAD patients with elevated D6D activity had a higher risk of diabetes [37]. Although the mechanism by which insulin resistance increases cardiovascular risk is still unclear, D5D and D6D, which are encoded by FADS1 and FADS2, may influence glucose metabolism [38]. Several studies have reported that SNPs in the FADS gene cluster are correlated with desaturase activity and that this correlation differed among ethnic groups [15, 39].
As we all know, cardiac and vascular diseases are often accompanied by diabetes, however, in some patients who had had a CAD but who had not previously been diagnosed with diabetes. Obviously, the severity of healthy control, T2D, and T2D&CAD is gradually increased, which is a typical ordinal event of T2D progress. Thus, we use ordinal logistic regression to find out whether T2D patients with rs174537 GG genotype were at risk of developing T2D&CAD. Our results demonstrate that the genotype distributions of rs174537G>T influence desaturase activity and are associated with the risk of both T2D and CAD, which is consistent with previous reports [11, 40]. Besides, in order to exclude the impact of CAD, we set up a CAD group, therefore the ordinal logistic regression was also applied to the healthy control, CAD and CAD&T2D and the results are listed in Additional file 1: Table S1. We also found that the minor allele T of rs174537 was significantly associated with n-3 fatty acid concentrations in the three patient groups. Numerous SNPs in the FADS gene cluster were reported to be significantly associated with fatty acid alterations in serum and red blood cell membranes [16, 30, 41]. Genome-wide association studies analysis also confirmed that the rs174537 SNP is significantly associated with the AA level in patients with CAD and/or T2D [16]. However, in our study, we observed this association only in the T2D&CAD group. The rs174537 SNP is located in an intron and is in linkage disequilibrium with rs174546 (r2 = 0.99) and rs3834458 (r2 = 0.98), which can influence gene expression directly [17, 42]. Therefore, it is possible that this variant of the rs174537 SNP is a marker of other functional polymorphisms or is in linkage with currently unidentified causal variants that affect fatty acid concentrations. In addition, T2D&CAD patients with the rs174357 GG genotype had decreased D5D and elevated D6D and D9D activity. Our previous study of CAD patients also supports this finding [14].
Indeed, we were unable to take “one-size-fits-all” medication therapy for patients and standard energy intake for all subjects, that resulted in failure to adjust for dietary intake and medicine in the statistical analyses, and our HP-INNOWax column assay did not get the trace amounts of several fatty acids in human plasma, but with a large sample size, these factors did have limited influence on our research. Our data indicate that polyunsaturated fatty acid metabolisms, desaturase activity, and FADS polymorphisms contribute to the simultaneous development of T2D and CAD. Our results suggest that rs174537 may be a key SNP in the FADS gene cluster and is associated with plasma fatty acid levels.
Conclusions
It is the first time to show an association of plasma fatty acids, desaturase activities, and FADS genotypes with patients who had both T2D and CAD. Genetic variation in the FADS gene cluster, particularly rs174537, might cause the alterations in plasma fatty acids and desaturase levels, which could provide new insights for diagnostic and therapeutic strategies for patients with both T2D and CAD, and help prevent T2D patients from CAD.
Authors’ contributions
SWL and SML designed research; SWL, YY, PM, WL and HYL conducted research; LZJ and LC analyzed gas chromatography-mass spectrometry; SWL, JW, and SML prepared the initial manuscript draft; JW and SML edited and revised subsequent drafts; SML had primary responsibility for final content. All authors read and approved the final manuscript.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (81271919, 81472023) and the National Basic Research Program of China (973 Program) (2012CB720605). Special thanks to Professor Chuanhua Yu, School of Public Health, Wuhan University, for guidance and support in statistical analysis.
Competing interests
The authors declare that they have no competing interests.
Additional file
10.1186/s12967-016-0834-8 Online Supplemental Tables.
Contributor Information
Si-Wei Li, Email: 2012103030013@whu.edu.cn.
Jin Wang, Email: wangjin@shaphc.org.
Ying Yang, Email: yangying0109@whu.edu.cn.
Zhi-Jie Liu, Email: zjliu@whu.edu.cn.
Lin Cheng, Email: lincheng@hotmail.com.
Huan-Yu Liu, Email: 115634093@qq.com.
Pei Ma, Email: mapei2010@126.com.
Wan Luo, Email: 2012203030040@whu.edu.cn.
Song-Mei Liu, Phone: +86-27-67812610, Email: smliu@whu.edu.cn.
References
- 1.Zhang P, Zhang X, Brown J, Vistisen D, Sicree R, Shaw J, Nichols G. Global healthcare expenditure on diabetes for 2010 and 2030. Diabetes Res Clin Pract. 2010;87:293–301. doi: 10.1016/j.diabres.2010.01.026. [DOI] [PubMed] [Google Scholar]
- 2.Oikawa S, Yokoyama M, Origasa H, Matsuzaki M, Matsuzawa Y, Saito Y, Ishikawa Y, Sasaki J, Hishida H, Itakura H, et al. Suppressive effect of EPA on the incidence of coronary events in hypercholesterolemia with impaired glucose metabolism: Sub-analysis of the Japan EPA Lipid Intervention Study (JELIS) Atherosclerosis. 2009;206:535–539. doi: 10.1016/j.atherosclerosis.2009.03.029. [DOI] [PubMed] [Google Scholar]
- 3.Betteridge DJ. Lipid control in patients with diabetes mellitus. Nat Rev Cardiol. 2011;8:278–290. doi: 10.1038/nrcardio.2011.23. [DOI] [PubMed] [Google Scholar]
- 4.An Y, Zhang P, Wang J, Gong Q, Gregg EW, Yang W, Li H, Zhang B, Shuai Y, Chen Y, Engelgau MM, Cheng Y, Hu Y, Bennett PH, Li G. Cardiovascular and all-cause mortality over a 23-year period among chinese with newly diagnosed diabetes in the da qing igt and diabetes study. Diabetes Care. 2015;34:1365–1371. doi: 10.2337/dc14-2498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Jia WP, Pang C, Chen L, Bao YQ, Lu JX, Lu HJ, Tang JL, Wu YM, Zuo YH, Jiang SY, Xiang KS. Epidemiological characteristics of diabetes mellitus and impaired glucose regulation in a Chinese adult population: the Shanghai diabetes studies, a cross-sectional 3-year follow-up study in Shanghai urban communities. Diabetologia. 2007;50:286–292. doi: 10.1007/s00125-006-0503-1. [DOI] [PubMed] [Google Scholar]
- 6.Xu Y, Wang L, He J, Bi Y, Li M, Wang T, Wang L, Jiang Y, Dai M, Lu J, et al. Prevalence and control of diabetes in Chinese adults. JAMA. 2013;310:948–959. doi: 10.1001/jama.2013.168118. [DOI] [PubMed] [Google Scholar]
- 7.Xu FX, Zhou X, Shen F, Pang R, Liu SM. Decreased peripheral blood mitochondrial DNA content is related to HbA1c, fasting plasma glucose level and age of onset in Type 2 diabetes mellitus. Diabet Med. 2012;29:E47–E54. doi: 10.1111/j.1464-5491.2011.03565.x. [DOI] [PubMed] [Google Scholar]
- 8.Shen F, Qi JH, Xu FX, Ning LP, Pang R, Zhou X, Liu SM. Age-related distributions of nine fasting plasma free fatty acids in a population of Chinese adults. Clin Chim Acta. 2013;415:81–87. doi: 10.1016/j.cca.2012.10.001. [DOI] [PubMed] [Google Scholar]
- 9.Tosi F, Sartori F, Guarini P, Olivieri O, Martinelli N. Delta-5 and delta-6 desaturases: crucial enzymes in polyunsaturated fatty acid-related pathways with pleiotropic influences in health and disease. Adv Exp Med Biol. 2014;824:61–81. doi: 10.1007/978-3-319-07320-0_7. [DOI] [PubMed] [Google Scholar]
- 10.Park WJ, Kothapalli KS, Reardon HT, Lawrence P, Qian SB, Brenna JT. A novel FADS1 isoform potentiates FADS2-mediated production of eicosanoid precursor fatty acids. J Lipid Res. 2012;53:1502–1512. doi: 10.1194/jlr.M025312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hong SH, Kwak JH, Paik JK, Chae JS, Lee JH. Association of polymorphisms in FADS gene with age-related changes in serum phospholipid polyunsaturated fatty acids and oxidative stress markers in middle-aged nonobese men. Clin Interv Aging. 2013;8:585–596. doi: 10.2147/CIA.S42096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Harslof LB, Larsen LH, Ritz C, Hellgren LI, Michaelsen KF, Vogel U, Lauritzen L. FADS genotype and diet are important determinants of DHA status: a cross-sectional study in Danish infants. Am J Clin Nutr. 2013;97:1403–1410. doi: 10.3945/ajcn.113.058685. [DOI] [PubMed] [Google Scholar]
- 13.Cormier H, Rudkowska I, Paradis AM, Thifault E, Garneau V, Lemieux S, Couture P, Vohl MC. Association between polymorphisms in the fatty acid desaturase gene cluster and the plasma triacylglycerol response to an n-3 PUFA supplementation. Nutrients. 2012;4:1026–1041. doi: 10.3390/nu4081026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Li SW, Lin K, Ma P, Zhang ZL, Zhou YD, Lu SY, Zhou X, Liu SM. fads gene polymorphisms confer the risk of coronary artery disease in a chinese han population through the altered desaturase activities: based on high-resolution melting analysis. Plos ONE. 2013;8:e55869. doi: 10.1371/journal.pone.0055869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Merino DM, Johnston H, Clarke S, Roke K, Nielsen D, Badawi A, El-Sohemy A, Ma DW, Mutch DM. Polymorphisms in FADS1 and FADS2 alter desaturase activity in young Caucasian and Asian adults. Mol Genet Metab. 2011;103:171–178. doi: 10.1016/j.ymgme.2011.02.012. [DOI] [PubMed] [Google Scholar]
- 16.Tanaka T, Shen J, Abecasis GR, Kisialiou A, Ordovas JM, Guralnik JM, Singleton A, Bandinelli S, Cherubini A, Arnett D, et al. Genome-wide association study of plasma polyunsaturated fatty acids in the inchianti study. Plos Genetics. 2009;5:e1000338. doi: 10.1371/journal.pgen.1000338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Dixon AL, Liang L, Moffatt MF, Chen W, Heath S, Wong KCC, Taylor J, Burnett E, Gut I, Farrall M, et al. A genome-wide association study of global gene expression. Nat Genet. 2007;39:1202–1207. doi: 10.1038/ng2109. [DOI] [PubMed] [Google Scholar]
- 18.American Diabetes Association Standards of Medical Care in Diabetes—2012. Diabetes Care. 2012;35:S11–S63. doi: 10.2337/dc12-s011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Rapaport E. Nomenclature and criteria for diagnosis of ischemic heart-disease—report of the Joint-International-Society-and-Federation-of-Cardiology-World-Health-Organization Task-Force on standardization of clinical nomenclature. Circulation. 1979;59:607–609. doi: 10.1161/01.CIR.59.3.607. [DOI] [PubMed] [Google Scholar]
- 20.Liu ZJ, Gao Y, Chen J, Imanaka T, Bao J, Hua Q. Analysis of metabolic fluxes for better understanding of mechanisms related to lipid accumulation in oleaginous yeast Trichosporon cutaneum. Bioresour Technol. 2013;130:144–151. doi: 10.1016/j.biortech.2012.12.072. [DOI] [PubMed] [Google Scholar]
- 21.Sole X, Guino E, Valls J, Iniesta R, Moreno V. SNPStats: a web tool for the analysis of association studies. Bioinformatics. 2006;22:1928–1929. doi: 10.1093/bioinformatics/btl268. [DOI] [PubMed] [Google Scholar]
- 22.Mozaffarian D, Wu JHY. (n-3) Fatty acids and cardiovascular health: are effects of epa and dha shared or complementary? J Nutr. 2012;142:614s–625s. doi: 10.3945/jn.111.149633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Cottin SC, Sanders TA, Hall WL. The differential effects of EPA and DHA on cardiovascular risk factors. Proceedings of the Nutrition Society. 2011;70:215–231. doi: 10.1017/S0029665111000061. [DOI] [PubMed] [Google Scholar]
- 24.Yamagishi K, Hori M, Iso H. Fish and omega-3 polyunsaturated fatty acids in relation to risk of cardiovascular disease. Nihon Rinsho. 2013;71:1552–1557. [PubMed] [Google Scholar]
- 25.Ohnishi H, Saito Y. Eicosapentaenoic acid (EPA) reduces cardiovascular events: relationship with the EPA/arachidonic acid ratio. J Atheroscler Thromb. 2013;20:861–877. doi: 10.5551/jat.18002. [DOI] [PubMed] [Google Scholar]
- 26.Saito Y, Yokoyama M, Origasa H, Matsuzaki M, Matsuzawa Y, Ishikawa Y, Oikawa S, Sasaki J, Hishida H, Itakura H, et al. Effects of EPA on coronary artery disease in hypercholesterolemic patients with multiple risk factors: sub-analysis of primary prevention cases from the Japan EPA lipid intervention study (JELIS) Atherosclerosis. 2008;200:135–140. doi: 10.1016/j.atherosclerosis.2008.06.003. [DOI] [PubMed] [Google Scholar]
- 27.Investigators OT, Bosch J, Gerstein HC, Dagenais GR, Diaz R, Dyal L, Jung H, Maggiono AP, Probstfield J, Ramachandran A, et al. n-3 Fatty acids and cardiovascular outcomes in patients with dysglycemia. N Engl J Med. 2012;367:309–318. doi: 10.1056/NEJMoa1203859. [DOI] [PubMed] [Google Scholar]
- 28.Chi MD, Vansomphone SS, Liu IL, Cheetham C, Green KR, Scott RD, Reynolds K. Adherence to statins and LDL-cholesterol goal attainment. Am J Manag Care. 2014;20:e105–e112. [PubMed] [Google Scholar]
- 29.Rosenson RS, Underberg JA. Systematic review: evaluating the effect of lipid-lowering therapy on lipoprotein and lipid values. Cardiovasc Drugs Ther. 2013;27:465–479. doi: 10.1007/s10557-013-6477-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Bokor S, Dumont J, Spinneker A, Gonzalez-Gross M, Nova E, Widhalm K, Moschonis G, Stehle P, Amouyel P, De Henauw S, et al. Single nucleotide polymorphisms in the FADS gene cluster are associated with delta-5 and delta-6 desaturase activities estimated by serum fatty acid ratios. J Lipid Res. 2010;51:2325–2333. doi: 10.1194/jlr.M006205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kathiresan S, Willer CJ, Peloso GM, Demissie S, Musunuru K, Schadt EE, Kaplan L, Bennett D, Li Y, Tanaka T, et al. Common variants at 30 loci contribute to polygenic dyslipidemia. Nat Genet. 2009;41:56–65. doi: 10.1038/ng.291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Warensjo E, Rosell M, Hellenius ML, Vessby B, De Faire U, Riserus U. Associations between estimated fatty acid desaturase activities in serum lipids and adipose tissue in humans: links to obesity and insulin resistance. Lipids Health Dis. 2009;8:1. doi: 10.1186/1476-511X-8-37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Kroger J, Schulze MB. Recent insights into the relation of Delta 5 desaturase and Delta 6 desaturase activity to the development of type 2 diabetes. Curr Opin Lipidol. 2012;23:4–10. doi: 10.1097/MOL.0b013e32834d2dc5. [DOI] [PubMed] [Google Scholar]
- 34.Hodge AM, English DR, O’Dea K, Sinclair AJ, Makrides M, Gibson RA, Giles GG. Plasma phospholipid and dietary fatty acids as predictors of type 2 diabetes: interpreting the role of linoleic acid. Am J Clin Nutr. 2007;86:189–197. doi: 10.1093/ajcn/86.1.189. [DOI] [PubMed] [Google Scholar]
- 35.Patel PS, Sharp SJ, Jansen E, Luben RN, Khaw KT, Wareham NJ, Forouhi NG. Fatty acids measured in plasma and erythrocyte-membrane phospholipids and derived by food-frequency questionnaire and the risk of new-onset type 2 diabetes a pilot study in the European Prospective Investigation into Cancer and Nutrition (EPIC)-Norfolk cohort. Am J Clin Nutr. 2010;92:1214–1222. doi: 10.3945/ajcn.2010.29182. [DOI] [PubMed] [Google Scholar]
- 36.Kroger J, Zietemann V, Enzenbach C, Weikert C, Jansen EHJM, Doring F, Joost HG, Boeing H, Schulze MB. Erythrocyte membrane phospholipid fatty acids, desaturase activity, and dietary fatty acids in relation to risk of type 2 diabetes in the European Prospective Investigation into Cancer and Nutrition (EPIC)-Potsdam Study. Am J Clin Nutr. 2011;93:127–142. doi: 10.3945/ajcn.110.005447. [DOI] [PubMed] [Google Scholar]
- 37.Martinelli N, Girelli D, Malerba G, Guarini P, Illig T, Trabetti E, Sandri M, Friso S, Pizzolo F, Schaeffer L, et al. FADS genotypes and desaturase activity estimated by the ratio of arachidonic acid to linoleic acid are associated with inflammation and coronary artery disease. Am J Clin Nutr. 2008;88:941–949. doi: 10.1093/ajcn/88.4.941. [DOI] [PubMed] [Google Scholar]
- 38.Cormier H, Rudkowska I, Thifault E, Lemieux S, Couture P, Vohl MC. Polymorphisms in fatty acid desaturase (fads) gene cluster: effects on glycemic controls following an omega-3 polyunsaturated fatty acids (pufa) supplementation. Genes (Basel) 2013;4:485–498. doi: 10.3390/genes4030485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Sergeant S, Hugenschmidt CE, Rudock ME, Ziegler JT, Ivester P, Ainsworth HC, Vaidya D, Case LD, Langefeld CD, Freedman BI, et al. Differences in arachidonic acid levels and fatty acid desaturase (FADS) gene variants in African Americans and European Americans with diabetes or the metabolic syndrome. Br J Nutr. 2012;107:547–555. doi: 10.1017/S0007114511003230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Mathias RA, Vergara C, Gao L, Rafaels N, Hand T, Campbell M, Bickel C, Ivester P, Sergeant S, Barnes KC, Chilton FH. FADS genetic variants and omega-6 polyunsaturated fatty acid metabolism in a homogeneous island population. J Lipid Res. 2010;51:2766–2774. doi: 10.1194/jlr.M008359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Burghardt KJ, Gardner KN, Johnson JW, Ellingrod VL. Fatty acid desaturase gene polymorphisms and metabolic measures in schizophrenia and bipolar patients taking antipsychotics. Cardiovasc Psychiatry Neurol. 2013;2013:596945. doi: 10.1155/2013/596945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Simopoulos AP. Genetic variants in the metabolism of omega-6 and omega-3 fatty acids: their role in the determination of nutritional requirements and chronic disease risk. Exp Biol Med (Maywood) 2010;235:785–795. doi: 10.1258/ebm.2010.009298. [DOI] [PubMed] [Google Scholar]